
Genome-Wide Identification and Characterization of Ammonium Transporter (AMT) Genes in Chlamydomonas reinhardtii
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Bioinformatics Analyses of CrAMTs
2.2. Phylogenetic Tree Analysis of CrAMTs
2.3. Gene Structures, Conserved Motifs, and Conserved Protein Domain Analyses of CrAMTs
2.4. Chromosome Localization Analysis and Collinearity Analysis of CrAMTs
2.5. Cis-Element Analysis of CrAMT Promoter Regions
2.6. Expression Patterns of CrAMT Genes under High-Ammonium Conditions
2.7. Construction and Phenotypic Characterization of Transgenic Algas
2.8. Growth Trend of Transgenic algas under High-Ammonium Treatment
2.9. Statistical Analysis
3. Results
3.1. Identification of CrAMTs and the Basic Information of CrAMTs
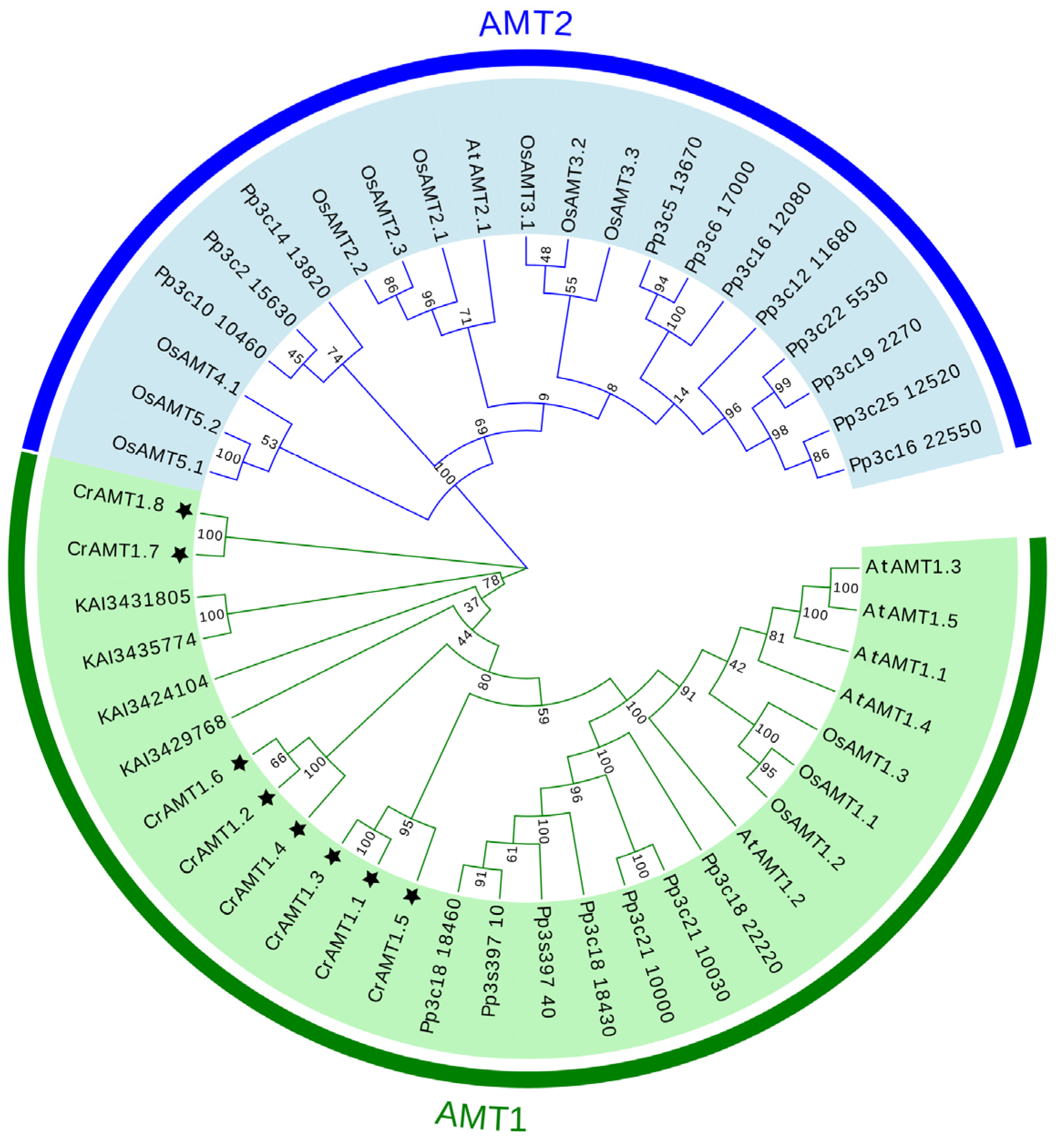
3.2. Phylogenetic Analyses of CrAMT Genes
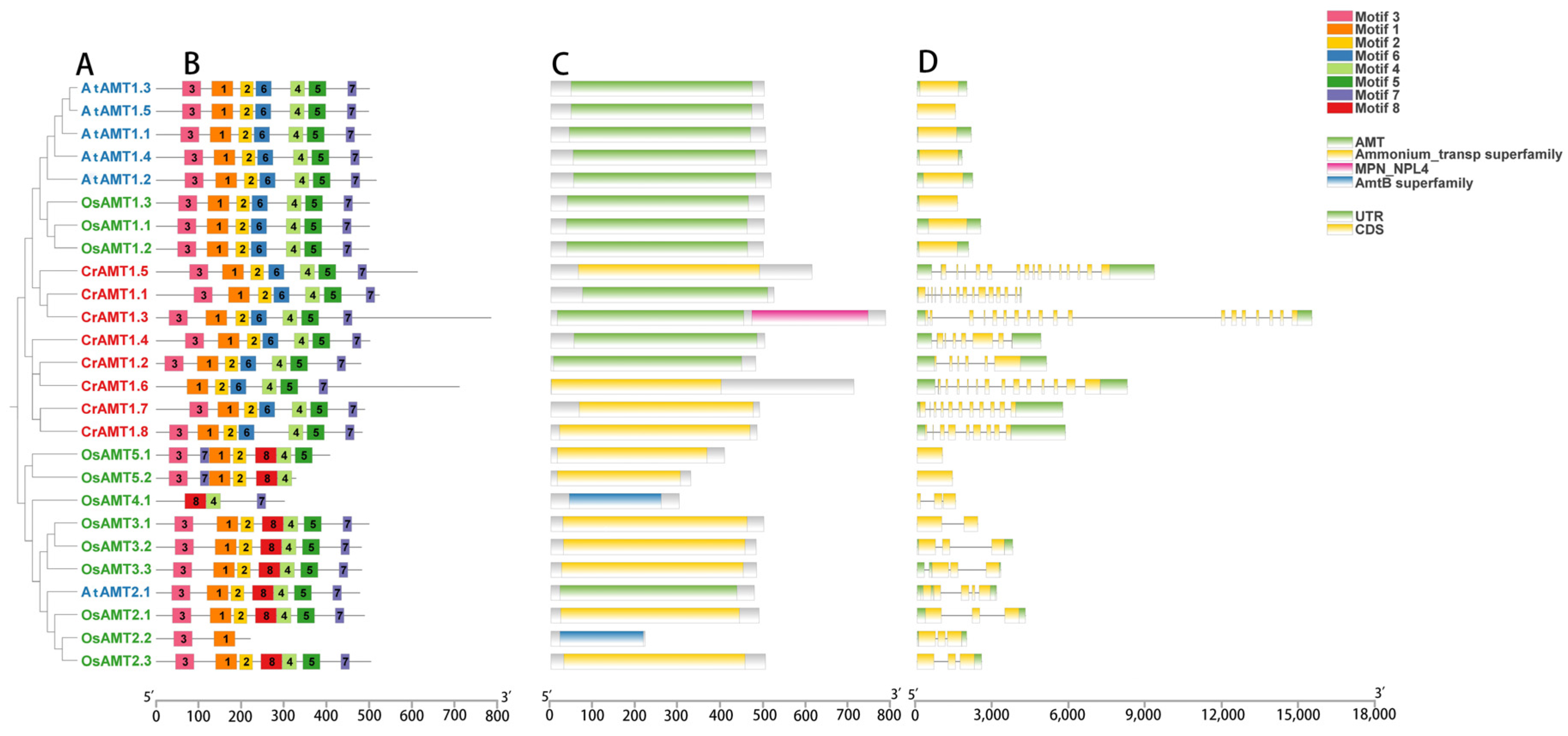
3.3. Characteristics of CrAMT Proteins
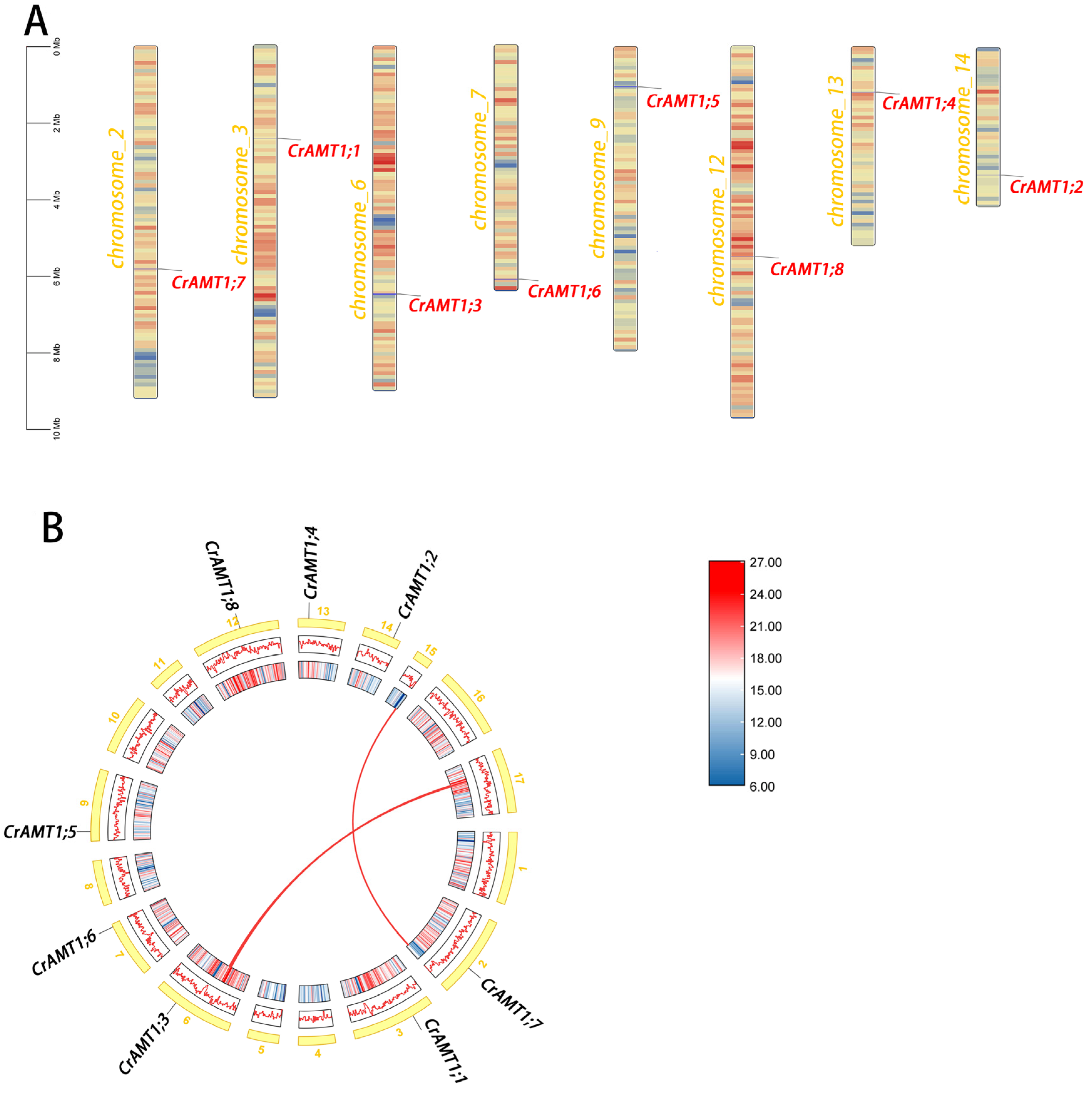
3.4. Chromosome Localization Analysis and Collinearity Analysis of CrAMTs
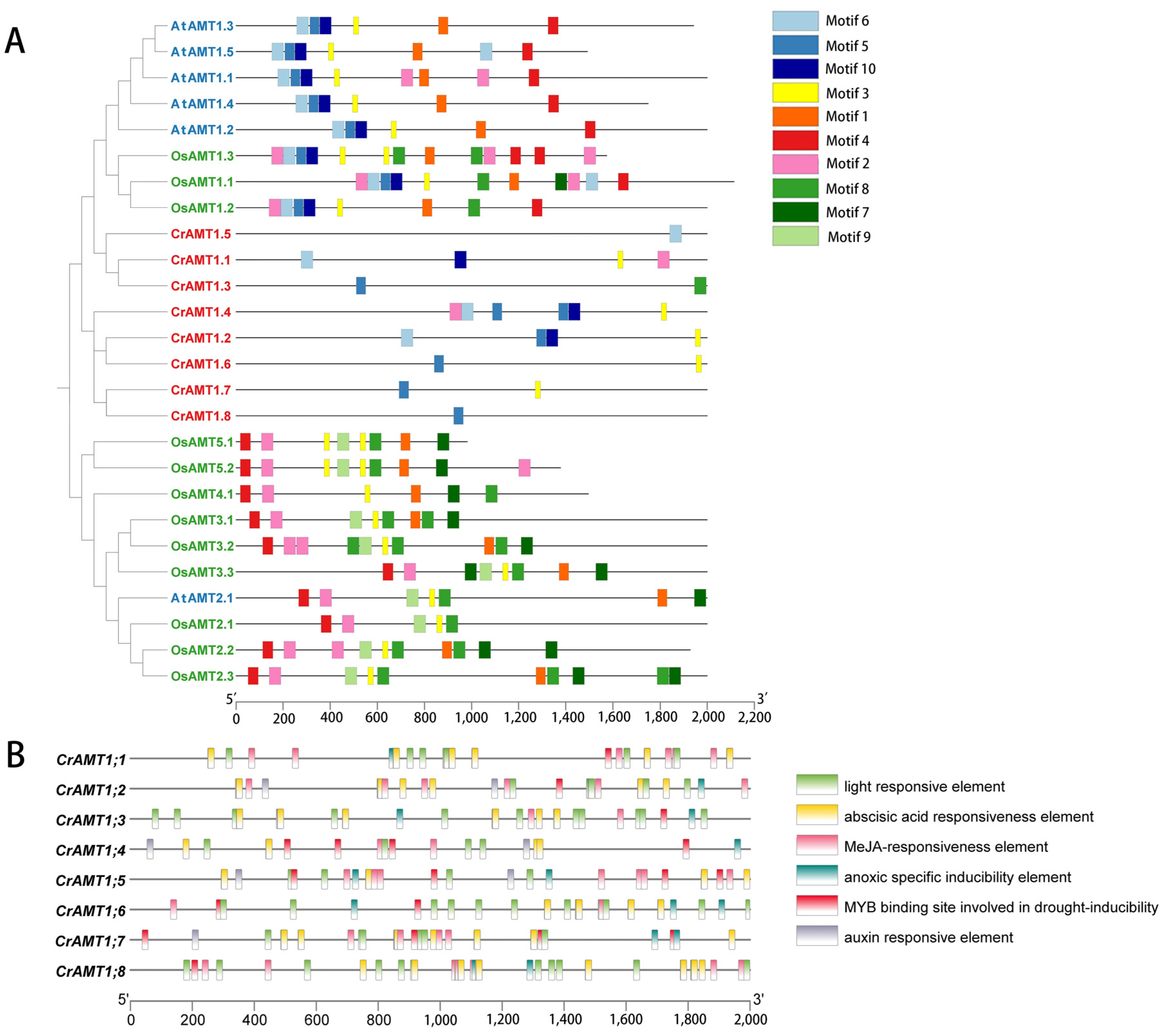
3.5. Promoter Cis-Element Analysis of CrAMTs
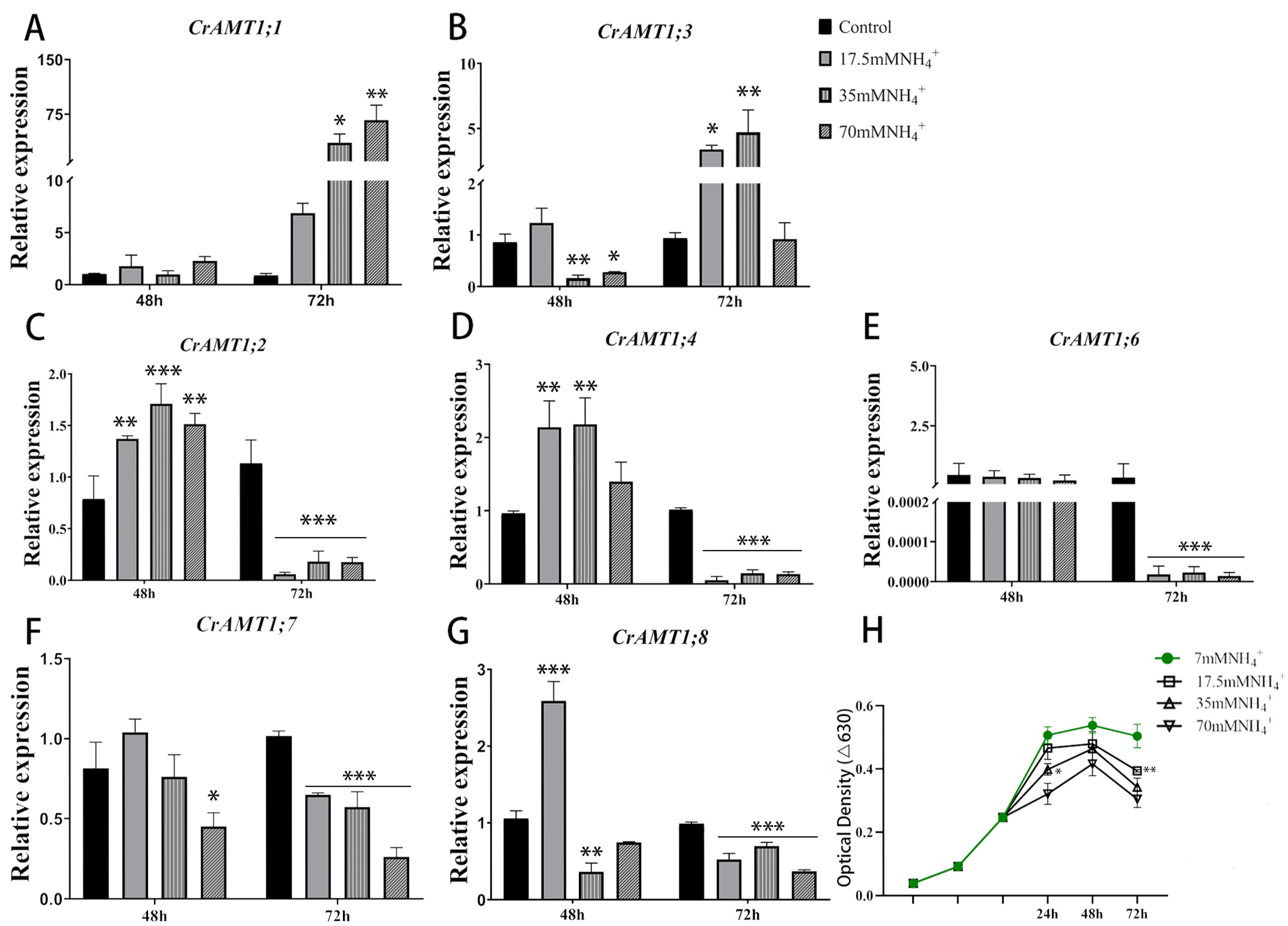
3.6. Expression Patterns of CrAMT Genes under High-Ammonium Conditions
3.7. Identification and Analysis of CrAMT1;7-OE
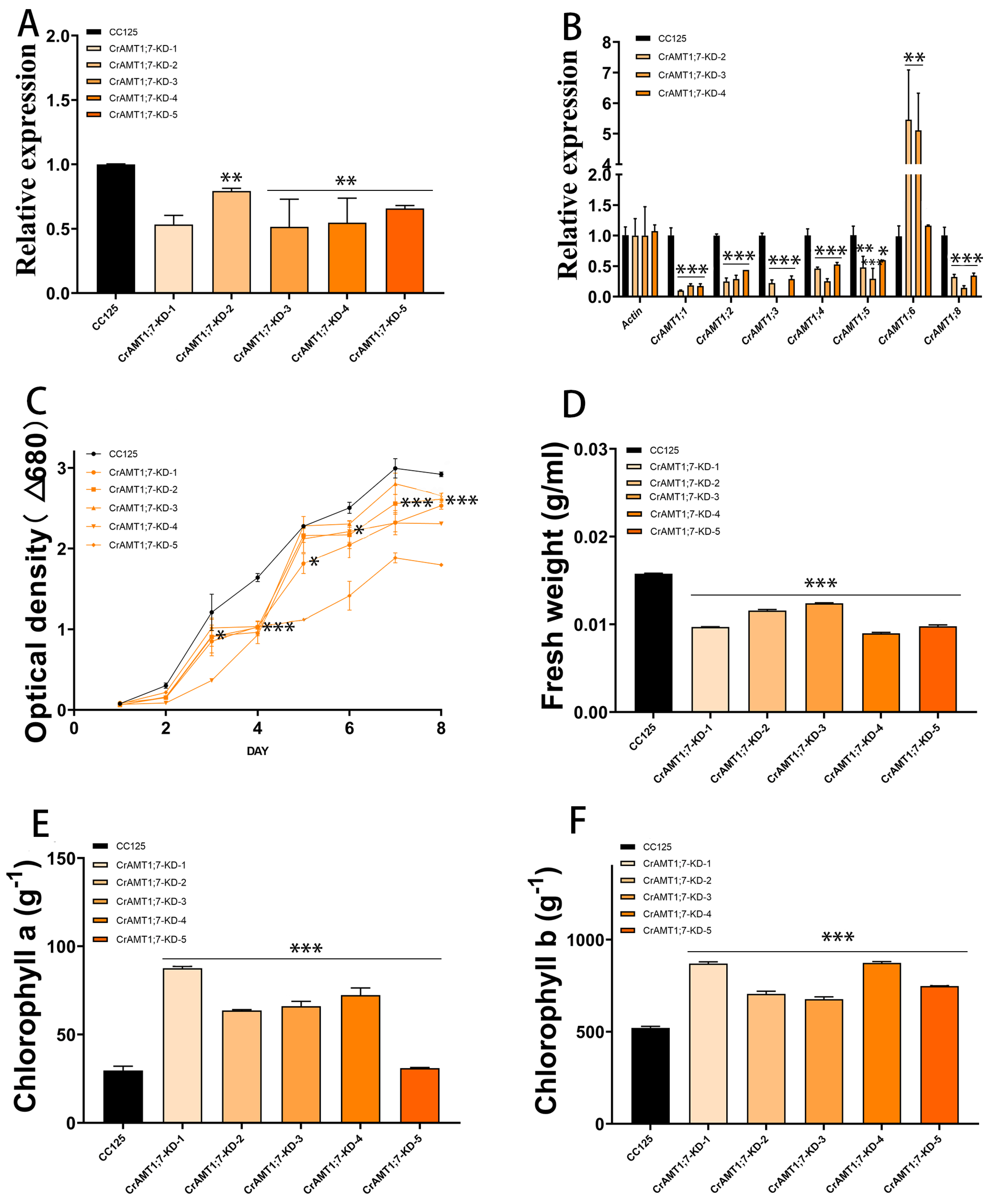
3.8. Identification and Analysis of CrAMT1;7-KD
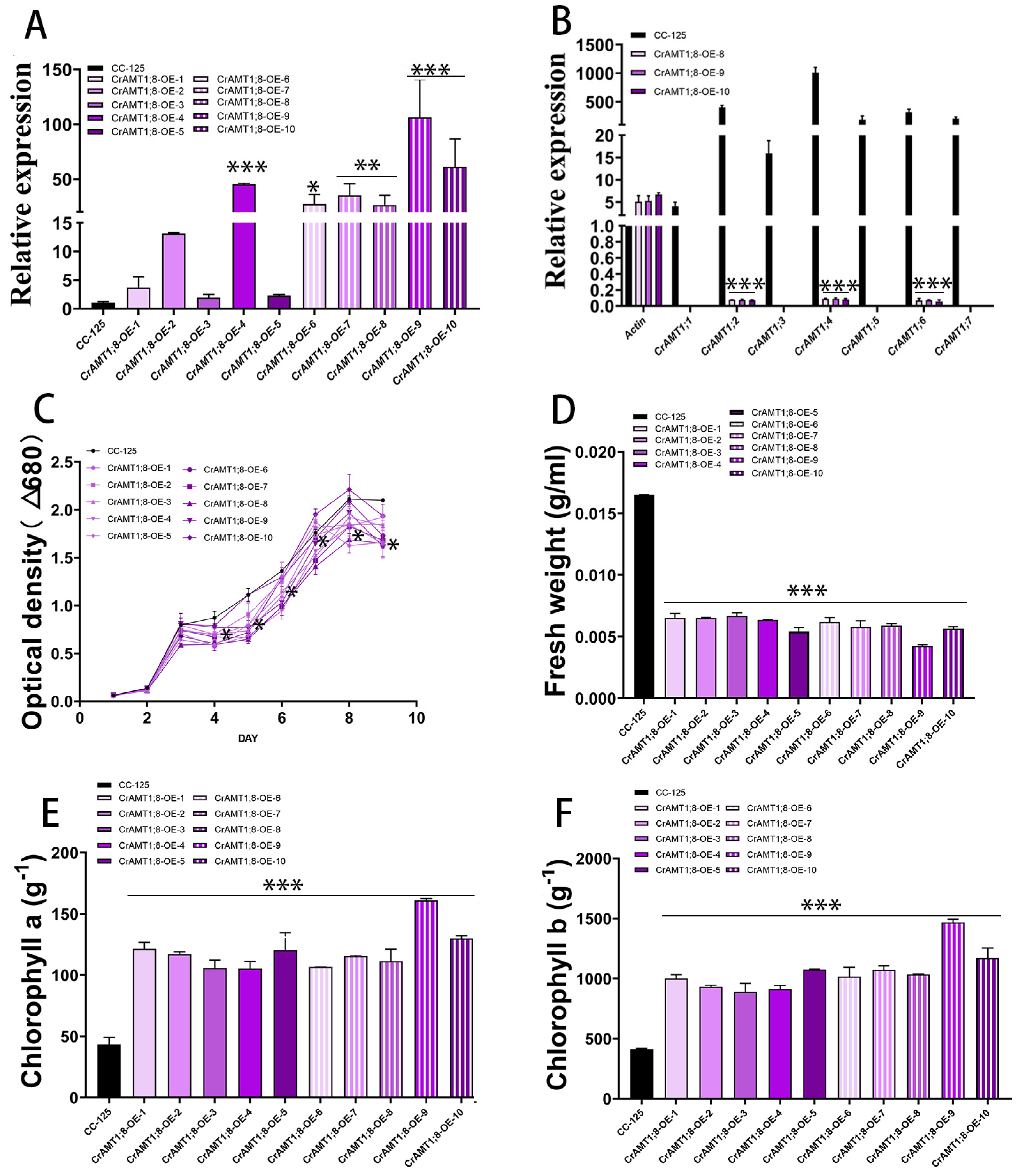
3.9. Identification and Analysis of CrAMT1;8-OE
3.10. Identification and Analysis of CrAMT1;8-KD
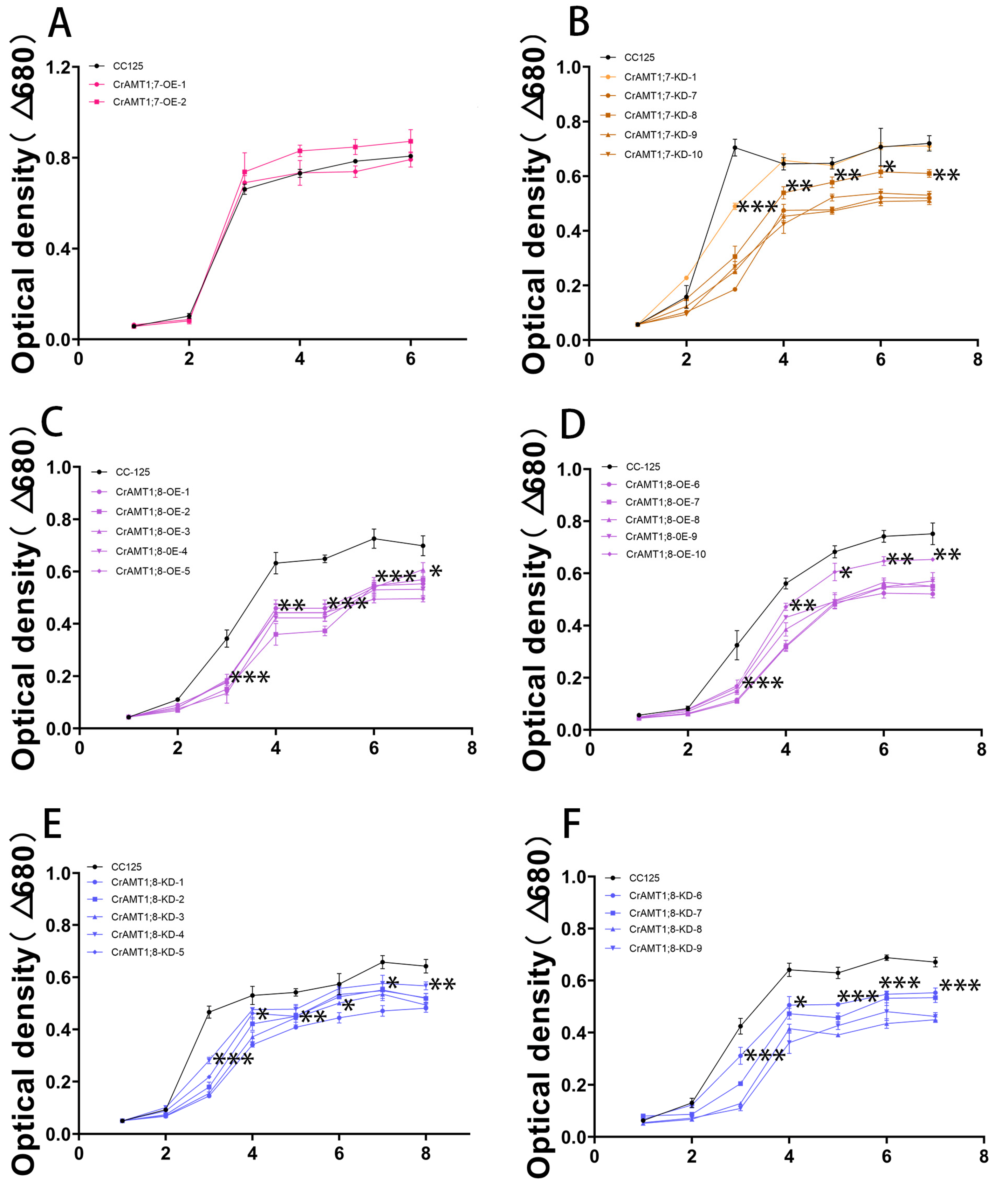
3.11. Growth Trend of Transgenic algas under High-Ammonium Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gazzarrini, S.; Lejay, L.; Gojon, A.; Ninnemann, O.; Frommer, W.B.; von Wirén, N. Three functional transporters for constitutive, diurnally regulated, and starvation-induced uptake of ammonium into Arabidopsis roots. Plant Cell 1999, 11, 937–947. [Google Scholar] [CrossRef]
- Zhang, J.; Cai, Z.; Müller, C. Terrestrial N cycling associated with climate and plant-specific N preferences: A review. Eur. J. Soil Sci. 2018, 69, 488–501. [Google Scholar] [CrossRef]
- Hao, D.L.; Zhou, J.Y.; Yang, S.Y.; Qi, W.; Yang, K.J.; Su, Y.H. Function and regulation of ammonium transporters in plants. Int. J. Mol. Sci. 2020, 21, 3557. [Google Scholar] [CrossRef]
- Yildiztugay, E.; Ozfidan-Konakci, C.; Cavusoglu, H.; Arikan, B.; Alp, F.N.; Elbasan, F.; Kucukoduk, M.; Turkan, I. Nanomaterial sulfonated graphene oxide advances the tolerance against nitrate and ammonium toxicity by regulating chloroplastic redox balance, photochemistry of photosystems and antioxidant capacity in Triticum aestivum. J. Hazard. Mater. 2022, 424, 127310. [Google Scholar] [CrossRef]
- Xiao, Y.; Luan, H.; Lu, S.; Xing, M.; Guo, C.; Qian, R.; Xiao, X. Toxic effects of atmospheric deposition in mining areas on wheat seedlings. Environ. Geochem. Health 2024, 46, 69. [Google Scholar] [CrossRef]
- McDonald, T.R.; Dietrich, F.S.; Lutzoni, F. Multiple horizontal gene transfers of ammonium transporters/ammonia permeases from prokaryotes to eukaryotes: Toward a new functional and evolutionary classification. Mol. Biol. Evol. 2011, 29, 51–60. [Google Scholar] [CrossRef]
- Li, B.; Zhao, Y.; Wang, S.; Zhang, X.; Wang, Y.; Shen, Y.; Yuan, Z. Genome-wide identification, gene cloning, subcellular location and expression analysis of SPL gene family in P. Granatum L. BMC Plant Biol. 2021, 21, 400. [Google Scholar] [CrossRef]
- Ludewig, U.; Neuhäuser, B.; Dynowski, M. Molecular mechanisms of ammonium transport and accumulation in plants. FEBS Lett. 2007, 581, 2301–2308. [Google Scholar] [CrossRef]
- Yuan, L.; Graff, L.; Loqué, D.; Kojima, S.; Tsuchiya, Y.N.; Takahashi, H.; von Wirén, N. AtAMT1;4, a pollen-specific high-affinity ammonium transporter of the plasma membrane in Arabidopsis. Plant Cell Physiol. 2009, 50, 13–25. [Google Scholar] [CrossRef]
- Giehl, R.F.H.; Laginha, A.M.; Duan, F.; Rentsch, D.; Yuan, L.; von Wirén, N. A critical role of AMT2;1 in root-to-shoot translocation of ammonium in Arabidopsis. Mol. Plant 2017, 10, 1449–1460. [Google Scholar] [CrossRef]
- Kronzucker, H.J.; Siddiqi, M.Y.; Glass, A.D.M. Kinetics of NH4+ influx in spruce. Plant Physiol. 1996, 110, 773–779. [Google Scholar] [CrossRef]
- Salvagiotti, F.; Cassman, K.G.; Specht, J.E.; Walters, D.T.; Weiss, A.; Dobermann, A. Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crops Res. 2008, 108, 1–13. [Google Scholar] [CrossRef]
- Chiasson, D.M.; Loughlin, P.C.; Mazurkiewicz, D.; Mohammadidehcheshmeh, M.; Fedorova, E.E.; Okamoto, M.; McLean, E.; Glass, A.D.M.; Smith, S.E.; Bisseling, T.; et al. Soybean SAT1 (Symbiotic Ammonium Transporter 1) encodes a bHLH transcription factor involved in nodule growth and NH4+ transport. Proc. Natl. Acad. Sci. USA 2014, 111, 4814–4819. [Google Scholar] [CrossRef]
- Javelle, A.; Morel, M.; Rodríguez-Pastrana, B.R.; Botton, B.; André, B.; Marini, A.M.; Brun, A.; Chalot, M. Molecular characterization, function and regulation of ammonium transporters (AMT) and ammonium-metabolizing enzymes (GS, NADP-GDH) in the ectomycorrhizal fungus Hebeloma cylindrosporum. Mol. Microbiol. 2003, 47, 411–430. [Google Scholar] [CrossRef]
- Couturier, J.; Montanini, B.; Martin, F.; Brun, A.; Blaudez, D.; Chalot, M. The expanded family of ammonium transporters in the perennial poplar plant. New Phytol. 2007, 174, 137–150. [Google Scholar] [CrossRef]
- Meier-Wagner, J.; Nolden, L.; Jakoby, M.; Siewe, R.; Krämer, R.; Burkovski, A. Multiplicity of ammonium uptake systems in Corynebacterium glutamicum role of AMT and AMTB. Microbiology 2001, 147, 135–143. [Google Scholar] [CrossRef]
- Javelle, A.; Severi, E.; Thornton, J.; Merrick, M. Ammonium sensing in Escherichia coli. J. Biol. Chem. 2004, 279, 8530–8538. [Google Scholar] [CrossRef]
- Loqué, D.; Yuan, L.; Kojima, S.; Gojon, A.; Wirth, J.; Gazzarrini, S.; Fitzpatrick, T.B.; von Wirén, N. Additive contribution of AMT1;1 and AMT1;3 to high-affinity ammonium uptake across the plasma membrane of nitrogen-deficient Arabidopsis roots. Plant J. 2006, 48, 522–534. [Google Scholar] [CrossRef]
- Ninnemann, O.; Jauniaux, J.C.; Frommer, W.B. Identification of a high-affinity NH4+ transportre from plants. Embo J. 1994, 13, 3464–3471. [Google Scholar] [CrossRef]
- Loqué, D.; Lalonde, S.; Looger, L.L.; von Wiren, N.; Frommer, W.B. A cytosolic trans-activation domain essential for ammonium uptake. Nature 2007, 446, 195–198. [Google Scholar] [CrossRef]
- Kaiser, B.N.; Rawat, S.R.; Siddiqi, M.Y.; Masle, J.; Glass, A.D.M. Functional analysis of an Arabidopsis T-DNA "knockout" of the high-affinity NH4+ transporter AtAMT1;1. Plant Physiol. 2002, 130, 1263–1275. [Google Scholar] [CrossRef] [PubMed]
- Konishi, N.; Ma, J.F. Three polarly localized ammonium transporter 1 members are cooperatively responsible for ammonium uptake in rice under low ammonium condition. New Phytol. 2021, 232, 1778–1792. [Google Scholar] [CrossRef]
- Liu, Y.; von Wirén, N. Ammonium as a signal for physiological and morphological responses in plants. J. Exp. Bot. 2017, 68, 2581–2592. [Google Scholar] [CrossRef] [PubMed]
- Westengen, O.T.; Ring, K.H.; Berg, P.R.; Brysting, A.K. Modern maize varieties going local in the semi-arid zone in Tanzania. BMC Evol. Biol. 2014, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Loqué, D.; Kojima, S.; Rauch, S.; Ishiyama, K.; Inoue, E.; Takahashi, H.; von Wirén, N. The organization of high-affinity ammonium uptake in Arabidopsis roots depends on the spatial arrangement and biochemical properties of AMT1-type transporters. Plant Cell 2007, 19, 2636–2652. [Google Scholar] [CrossRef] [PubMed]
- Koltun, A.; Maniero, R.A.; Vitti, M.; de Setta, N.; Giehl, R.F.H.; Lima, J.E.; Figueira, A. Functional characterization of the sugarcane (Saccharum spp.) ammonium transporter AMT2;1 suggests a role in ammonium root-to-shoot translocation. Front. Plant Sci. 2022, 13, 1039041. [Google Scholar] [CrossRef] [PubMed]
- Maniero, R.A.; Koltun, A.; Vitti, M.; Factor, B.G.; de Setta, N.; Camara, A.S.; Lima, J.E.; Figueira, A. Identification and functional characterization of the sugarcane (Saccharum spp.) AMT2-type ammonium transporter ScAMT3;3 revealed a presumed role in shoot ammonium remobilization. Front. Plant Sci. 2023, 14, 1299025. [Google Scholar] [CrossRef]
- Xia, J.; Wang, Y.; Zhang, T.; Pan, C.; Ji, Y.; Zhou, Y.; Jiang, X. Genome-wide identification, expression profiling, and functional analysis of ammonium transporter 2 (AMT2) gene family in cassava (Manihot esculenta crantz). Front. Genet. 2023, 14, 1145735. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lin, L.; Wang, T.; Chen, M.; Song, B.; Sun, W. Genome-wide identification of AMT2-type ammonium transporters reveal that CsAMT2.2 and CsAMT2.3 potentially regulate NH4+ absorption among three different cultivars of Camellia sinensis. Int. J. Mol. Sci. 2022, 23, 15661. [Google Scholar] [CrossRef]
- Li, B.Z.; Mike, M.; Li, S.M.; Li, H.Y.; Zhu, S.W.; Shi, W.M.; Su, Y.-H. Molecular basis and regulation of ammonium transporter in rice. Rice Sci. 2009, 16, 314–322. [Google Scholar] [CrossRef]
- Suenaga, A.; Moriya, K.; Sonoda, Y.; Ikeda, A.; von Wirén, N.; Hayakawa, T.; Yamaguchi, J.; Yamaya, T. Constitutive expression of a novel-type ammonium transporter OsAMT2 in rice plants. Plant Cell Physiol. 2003, 44, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Gaur, V.S.; Singh, U.S.; Gupta, A.K.; Kumar, A. Influence of different nitrogen inputs on the members of ammonium transporter and glutamine synthetase genes in two rice genotypes having differential responsiveness to nitrogen. Mol. Biol. Rep. 2012, 39, 8035–8044. [Google Scholar] [CrossRef] [PubMed]
- Li, S.M.; Li, B.Z.; Shi, W.M. Expression patterns of nine ammonium transporters in Rice in response to N status. Pedosphere 2012, 22, 860–869. [Google Scholar] [CrossRef]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; Maréchal-Drouard, L.; et al. The Chlamydomonas genome reveals the evolution of key animal and plant functions. Science 2007, 318, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Gaur, V.S.; Singh, U.S.; Gupta, A.K.; Kumar, A. Understanding the differential nitrogen sensing mechanism in Rice genotypes through expression analysis of high and low affinity ammonium transporter genes. Mol. Biol. Rep. 2011, 39, 2233–2241. [Google Scholar] [CrossRef] [PubMed]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef]
- Chen, J.; Lin, A.H.; Chen, Z.P.; Wang, W.; Zhang, T.; Cai, H.; Cai, B. Ammonium sulfate gradient loading of brucine into liposomes: Effect of phospholipid composition on entrapment efficiency and physicochemical properties in vitro. Drug Dev. Ind. Pharm. 2010, 36, 245–253. [Google Scholar] [CrossRef]
- Weng, L.; Zhang, M.; Wang, K.; Chen, G.; Ding, M.; Yuan, W.; Zhu, Y.; Xu, W.; Xu, F. Potassium alleviates ammonium toxicity in Rice by reducing its uptake through activation of plasma membrane H+-ATPase to enhance proton extrusion. Plant Physiol. Biochem. 2020, 155, 429–437. [Google Scholar] [CrossRef]
- Xun, Z.; Guo, X.; Li, Y.; Wen, X.; Wang, C.; Wang, Y. Quantitative proteomics analysis of tomato growth inhibition by ammonium nitrogen. Plant Physiol. Biochem. 2020, 154, 129–141. [Google Scholar] [CrossRef]
- Hachiya, T.; Inaba, J.; Wakazaki, M.; Sato, M.; Toyooka, K.; Miyagi, A.; Kawai-Yamada, M.; Sugiura, D.; Nakagawa, T.; Kiba, T.; et al. Excessive ammonium assimilation by plastidic glutamine synthetase causes ammonium toxicity in arabidopsis thaliana. Nat. Commun. 2021, 12, 4944–4953. [Google Scholar] [CrossRef]
- Andrews, M.; Raven, J.A.; Lea, P.J. Do plants need nitrate? The mechanisms by which nitrogen form affects plants. Ann. Appl. Biol. 2013, 163, 174–199. [Google Scholar] [CrossRef]
- Li, T.; Liao, K.; Xu, X.; Gao, Y.; Wang, Z.; Zhu, X.; Jia, B.; Xuan, Y. Wheat ammonium transporter (AMT) gene family: Diversity and possible role in host–pathogen interaction with stem rust. Front. Plant Sci. 2017, 8, 1637. [Google Scholar] [CrossRef] [PubMed]
- Baskin, T.I.; Fang, G.; Yang, J.; Sun, T.; Wang, X.; Li, Y. Evidence that synergism between potassium and nitrate enhances the alleviation of ammonium toxicity in rice seedling roots. PLoS ONE 2021, 16, e0248796. [Google Scholar] [CrossRef]
- Du, W.; Zhang, Y.; Si, J.; Zhang, Y.; Fan, S.; Xia, H.; Kong, L. Nitrate alleviates ammonium toxicity in wheat (triticum aestivuml.) by regulating tricarboxylic acid cycle and reducing rhizospheric acidification and oxidative damage. Plant Signal. Behav. 2021, 16, e1991687. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Lali, M.N.; Xiong, H.; Luo, Y.; Wang, Y.; Wang, Y.; He, X.; Shi, X.; Zhang, Y. Seedlings of poncirus trifoliata exhibit tissue-specific detoxification in response to NH4+ toxicity. Plant Biol. 2024, 26, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Szczerba, M.W.; Britto, D.T.; Kronzucker, H.J. Rapid, futile k+ cycling and pool-size dynamics define low-affinity potassium transport in barley. Plant Physiol. 2006, 141, 1494–1507. [Google Scholar] [CrossRef]
- Jia, Z.; Zhang, J.; Jiang, W.; Wei, M.; Zhao, L.; Li, G. Different nitrogen concentrations affect strawberry seedlings nitrogen form preferences through nitrogen assimilation and metabolic pathways. Sci. Hortic. 2024, 332, 113236–113243. [Google Scholar] [CrossRef]
- Liu, Y.; Maniero, R.A.; Giehl, R.F.H.; Melzer, M.; Steensma, P.; Krouk, G.; Fitzpatrick, T.B.; von Wirén, N. Pdx1.1-dependent biosynthesis of vitamin b6 protects roots from ammonium-induced oxidative stress. Mol. Plant 2022, 15, 820–839. [Google Scholar] [CrossRef]
- Dai, G.Z. Ecological Effect and Molecular Mechanism of Ammonium Toxicity on Cyanobacteria. Ph.D. Thesis, Central China Normal University, Wuhan, China, 2012. [Google Scholar]
- Li, Y.; Zhou, J.; Hao, D.; Yang, S.; Su, Y. Arabidopsis under ammonium over-supply: Characteristics of ammonium toxicity in relation to the activity of ammonium transporters. Pedosphere 2020, 30, 314–325. [Google Scholar] [CrossRef]
- Xiao, C.; Fang, Y.; Wang, S.; He, K. The alleviation of ammonium toxicity in plants. J. Integr. Plant Biol. 2023, 65, 1362–1368. [Google Scholar] [CrossRef]
- Zhang, J.X.; Shi, J.P.; Yuan, C.Y.; Liu, X.C.; Du, G.L.; Fan, R.M.; Zhang, B. MicroRNA expression profile analysis of Chlamydomonas reinhardtii during lipid accumulation process under nitrogen deprivation stresses. Bioengineering 2021, 9, 6. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, W.; Wu, J.; Xie, K.; Li, X. Ljamt2; 2 promotes ammonium nitrogen transport during arbuscular mycorrhizal fungi symbiosis in lotus japonicus. Int. J. Mol. Sci. 2022, 23, 9522–9535. [Google Scholar] [CrossRef]
- Xu, F.; Pan, J. Potassium channel KCN11 is required for maintaining cellular osmolarity during nitrogen starvation to control proper cell physiology and TAG accumulation in Chlamydomonas reinhardtii. Biotechnol. Biofuels 2020, 13, 1–11. [Google Scholar] [CrossRef]
- Sui, J.K.; Cui, Y.X.; Zhang, J.K.; Li, S.Y.; Zhao, Y.; Bai, M.K.; Feng, G.; Wu, H. Enhanced biomass production and harvesting efficiency of Chlamydomonas reinhardtii under high-ammonium conditions by powdered oyster shell. Bioresour. Technol. 2024, 403, 130904. [Google Scholar] [CrossRef]
- Pagnuco, I.A.; Revuelta, M.; Victoria Bondino, H.G.; Brun, M.; Have, A.T. HMMER cut-off threshold tool (HMMERCTTER): Supervised classification of superfamily protein sequences with a reliable cut-off threshold. PLoS ONE 2018, 13, e0193757. [Google Scholar] [CrossRef]
- González-Ballester, D.; Camargo, A.; Fernández, E. Ammonium transporter genes in Chlamydomonas: The nitrate-specific regulatory gene NIT2 is involved in AMT1;1 expression. Plant Mol. Biol. 2004, 56, 863–878. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; I Hurwitz, D.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Xu, G.X.; Guo, C.C.; Shan, H.Y.; Kong, H.Z. Divergence of duplicate genes in exon–intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative pcr and the 2−δδct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef]
- Yang, W.; Dong, X.; Yuan, Z.; Zhang, Y.; Li, X.; Wang, Y. Genome-wide identification and expression analysis of the ammonium transporter family genes in soybean. Int. J. Mol. Sci. 2023, 24, 3991. [Google Scholar] [CrossRef]
- Gu, R.; Duan, F.; An, X.; Zhang, F.; von Wirén, N.; Yuan, L. Characterization of AMT-mediated high-affinity ammonium uptake in roots of maize (Zea mays L.). Plant Cell Physiol. 2013, 54, 1515–1524. [Google Scholar] [CrossRef]
- Xiaoxue, Y.; Yajun, S.; Panpan, L.; ImShik, L. Evolutionary analysis of AMT (ammonium transporters) family in Arabidopsis thaliana and Oryza sativa. Mol. Soil Biol. 2016, 7, 1–7. [Google Scholar] [CrossRef]
- Shaul, O. How introns enhance gene expression. Int. J. Biochem. Cell Biol. 2017, 91, 145–155. [Google Scholar] [CrossRef]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217, 109–119. [Google Scholar] [CrossRef]
- Jo, B.-S.; Choi, S.S. Introns: The functional benefits of introns in genomes. Genom. Inform. 2015, 13, 112–118. [Google Scholar] [CrossRef]
- Dai, G.Z.; Qiu, B.S.; Forchhammer, K. Ammonium tolerance in the cyanobacterium synechocystis sp. Strain pcc 6803 and the role of the psba multigene family. Plant Cell Environ. 2013, 37, 840–851. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5 ‘→3’) |
|---|---|
| Actin | F:CAGTAGGAGGCATAGGGTTTGG |
| R:TCAACGAATTGGGGTGTGTG | |
| a-tubulin | F:TGCTGTGGGACCTGGCTGA |
| R:GCCTTCTTGCTGGTGATGTTG | |
| CrAMT1;1 | F:ACGGTGCCTACGGATTCGGAAT |
| R:AGCCCAGCCTTGTTGAGGATGA | |
| CrAMT1;2 | F:GCCACACATTCACGCATCCCA |
| R:CGTCCAGCCTTCTCGCAATCAG | |
| CrAMT1;3 | F:AGCTGAGCCTCTGTCACATGGT |
| R:GGACGCAGCAAGTCAACCGATT | |
| CrAMT1;4 | F:GCCGACATGACAACGCACATTG |
| R:GCCACGCCCTTTCTCCAAGATG | |
| CrAMT1;5 | F:CATCCAGTACGGCGATCCTCCA |
| R:TGACACCCACCCTAAGCAGCAA | |
| CrAMT1;6 | F:CGGTGCTGCCTCGACATAAGTG |
| R:GCTCCGCTTGCTGTCATCTCTG | |
| CrAMT1;7 | F:CGCTGCTGGTTGATGCCTGAA |
| R:GCACGCACTGCTTCCACTCTAT | |
| CrAMT1;8 | F:GTGTGAGGTGGGCGTGTTCAAA |
| R:TGCCAGCGAAGACCGATAACCA |
| Gene Name | Protein Name | Gene locus | No. of Amino Acids (a.a.) | Length of CDS (bp) | Transmembrane Zone | Subcellular Localization | |
|---|---|---|---|---|---|---|---|
| Plasma Membrane | Vacuolar Membrane | ||||||
| CrAMT1;1 | CrAMT1.1 | Cre03.g159254 | 521 | 1566 | 11 | 12 | 1 |
| CrAMT1;2 | CrAMT1.2 | Cre14.g629920 | 478 | 1437 | 10 | 9 | 3 |
| CrAMT1;3 | CrAMT1.3 | Cre06.g293051 | 782 | 2349 | 11 | 11 | 2 |
| CrAMT1;4 | CrAMT1.4 | Cre13.g569850 | 499 | 1500 | 11 | 10 | 2 |
| CrAMT1;5 | CrAMT1.5 | Cre09.g400750 | 610 | 1833 | 10 | 10 | 1 |
| CrAMT1;6 | CrAMT1.6 | Cre07.g355650 | 708 | 2127 | 11 | 7 | 6 |
| CrAMT1;7 | CrAMT1.7 | Cre02.g111050 | 487 | 1464 | 11 | 9 | 1 |
| CrAMT1;8 | CrAMT1.8 | Cre12.g531000 | 481 | 1446 | 10 | 4 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, W.; Wang, D.; Zhao, S.; Ji, J.; Yang, J.; Wan, Y.; Yu, C. Genome-Wide Identification and Characterization of Ammonium Transporter (AMT) Genes in Chlamydomonas reinhardtii. Genes 2024, 15, 1002. https://doi.org/10.3390/genes15081002
Hu W, Wang D, Zhao S, Ji J, Yang J, Wan Y, Yu C. Genome-Wide Identification and Characterization of Ammonium Transporter (AMT) Genes in Chlamydomonas reinhardtii. Genes. 2024; 15(8):1002. https://doi.org/10.3390/genes15081002
Chicago/Turabian StyleHu, Wenhui, Dan Wang, Shuangshuang Zhao, Jiaqi Ji, Jing Yang, Yiqin Wan, and Chao Yu. 2024. "Genome-Wide Identification and Characterization of Ammonium Transporter (AMT) Genes in Chlamydomonas reinhardtii" Genes 15, no. 8: 1002. https://doi.org/10.3390/genes15081002