
Genome-Wide Analysis Reveals the Secondary Metabolome in Streptomyces kanasensis ZX01

Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Plasmids and Growth Conditions
2.2. Genome Sequencing, Annotation, and Analysis
2.3. Quantitative Real-time PCR Analysis
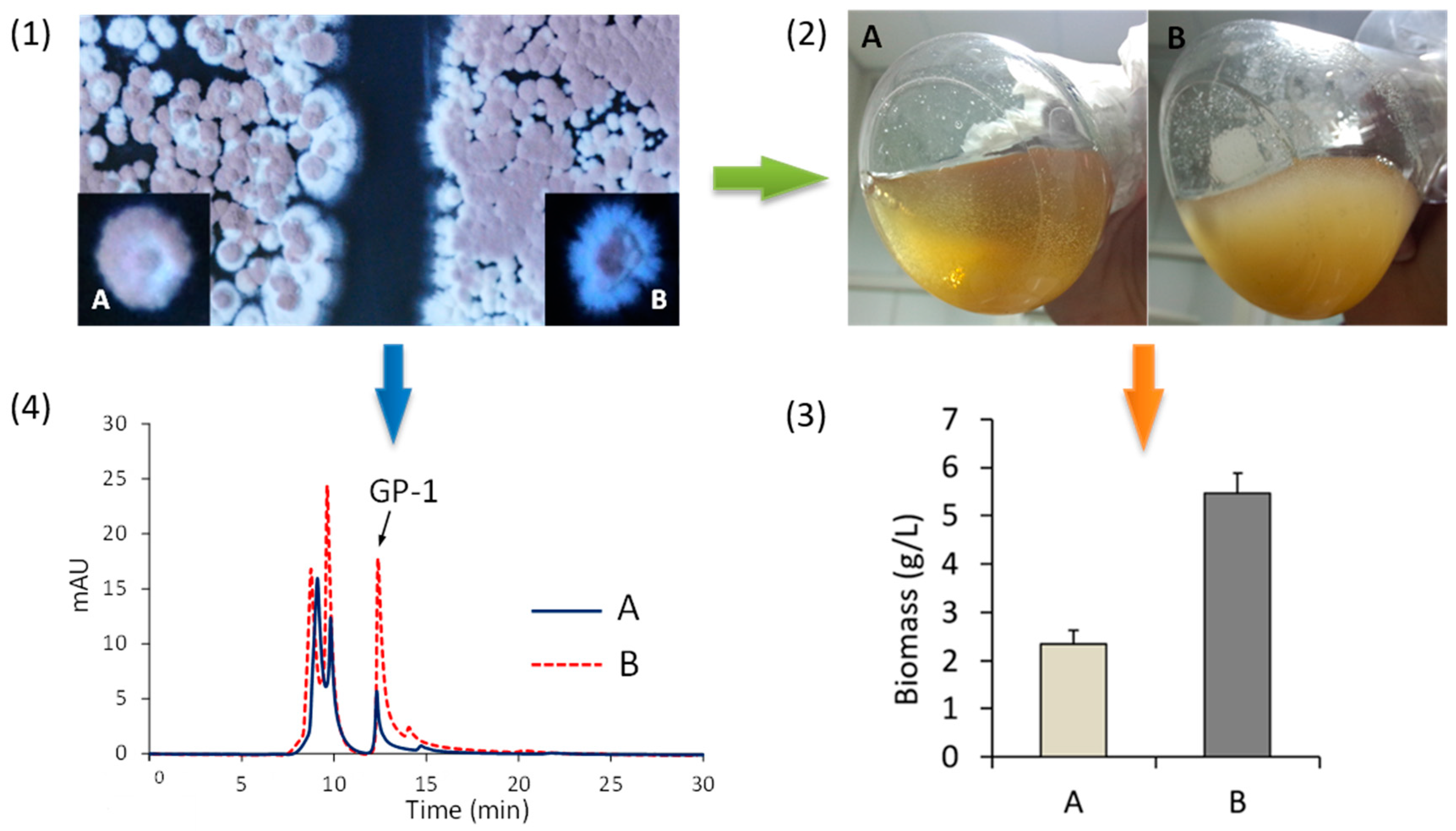
2.4. Construction of nsdA-Null Mutant
2.5. Purification and Determination of Glycoprotein GP-1
2.6. Statistical Analyses
3. Results and Discussion
3.1. Sequencing and Gene Annotation of the S. kanasensis ZX01
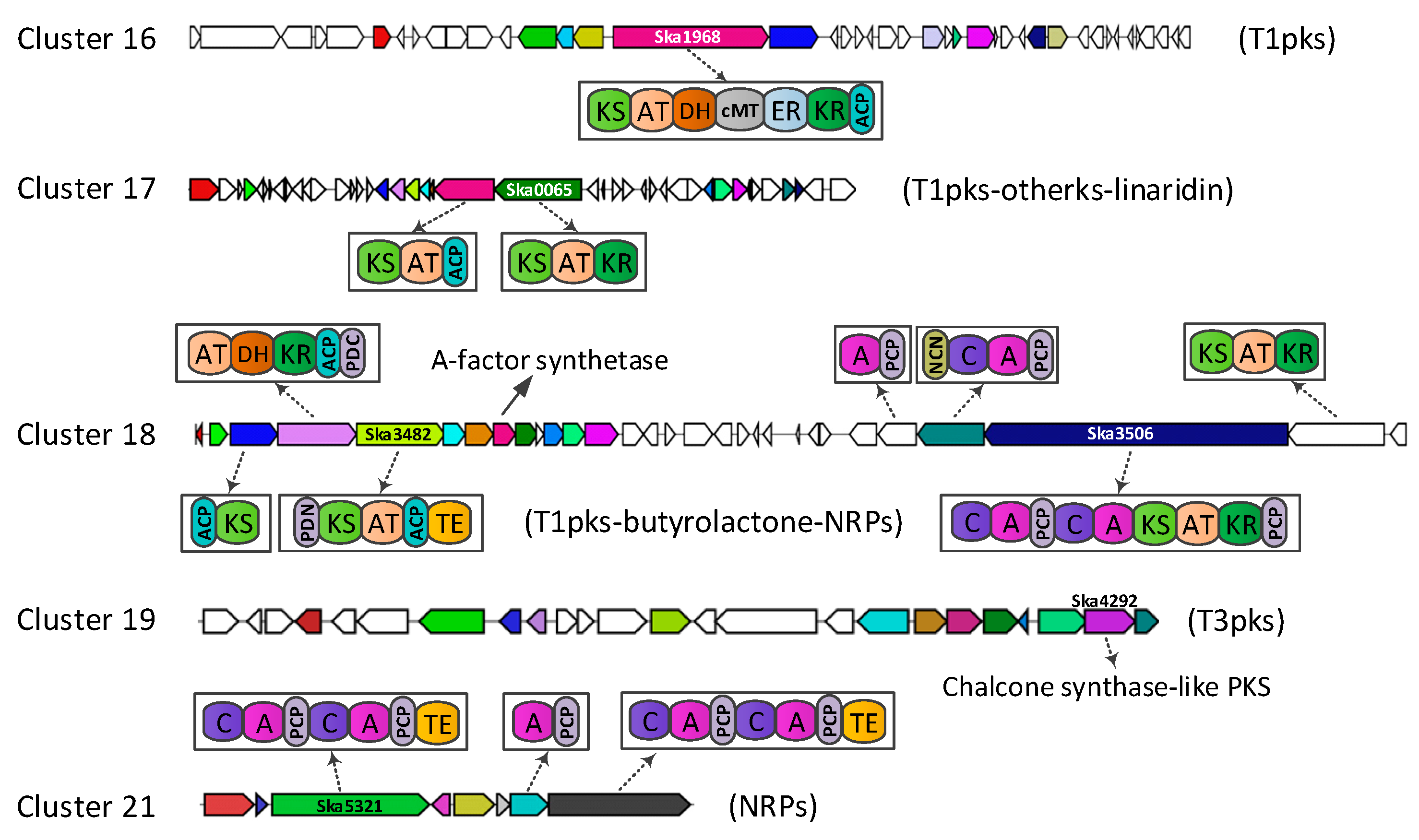
3.2. Gene Clusters for Secondary Metabolites
3.3. Regulators for Secondary Metabolism in S. kanasensis ZX01 Genome
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bentley, S.; Chater, K.; Cerdeno-Tarraga, A.M.; Challis, G.; Thomson, N.; James, K.; Harris, D.; Quail, M.; Kieser, H.; Harper, D. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2). Nature 2002, 417, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Zazopoulos, E.; Huang, K.; Staffa, A.; Liu, W.; Bachmann, B.O.; Nonaka, K.; Ahlert, J.; Thorson, J.S.; Shen, B.; Farnet, C.M. A genomics-guided approach for discovering and expressing cryptic metabolic pathways. Nat. Biotechnol. 2003, 21, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Udwary, D.W.; Zeigler, L.; Asolkar, R.N.; Singan, V.; Lapidus, A.; Fenical, W.; Jensen, P.R.; Moore, B.S. Genome sequencing reveals complex secondary metabolome in the marine actinomycete Salinispora tropica. Proc. Natl. Acad. Sci. USA 2007, 104, 10376–10381. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Chater, K.F.; Chandra, G.; Niu, G.; Tan, H. Molecular regulation of antibiotic biosynthesis in Streptomyces. Microbiol. Mol. Biol. Rev. 2013, 77, 112–143. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zhang, G.; Miao, G.; Zhang, X.; Feng, J. Streptomyces kanasensis sp. nov., an antiviral glycoprotein producing actinomycete isolated from forest soil around Kanas lake of china. Curr. Microbiol. 2015, 71, 627–631. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Han, L.; Zhang, G.; Zhang, X.; Feng, J. Purification and characterization of a novel glycoprotein from Streptomyces sp. ZX01. Int. J. Biol. Macromol. 2015, 78, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Kieser, T. Practical Streptomyces Genetics; John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Wang, L.Y.; Li, S.T.; Li, Y. Identification and characterization of a new exopolysaccharide biosynthesis gene cluster from Streptomyces. FEMS Microbiol. Lett. 2003, 220, 21–27. [Google Scholar] [CrossRef]
- Delcher, A.L.; Harmon, D.; Kasif, S.; White, O.; Salzberg, S.L. Improved microbial gene identification with Glimmer. Nucleic Acids Res. 1999, 27, 4636–4641. [Google Scholar] [CrossRef] [PubMed]
- Lukashin, A.V.; Borodovsky, M. Genemark.HMM: New solutions for gene finding. Nucleic Acids Res. 1998, 26, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Lagesen, K.; Hallin, P.; Rødland, E.A.; Stærfeldt, H.-H.; Rognes, T.; Ussery, D.W. RNAmmer: Consistent and rapid annotation of ribosomal RNA genes. Nucleic Acids Res. 2007, 35, 3100–3108. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Wolf, T.; Chevrette, M.G.; Lu, X.; Schwalen, C.J.; Kautsar, S.A.; Duran, S.; Hernando, G.; de Los Santos, E.L.; Kim, H.U.; et al. antiSMASH 4.0-improvements in chemistry prediction and gene cluster boundary identification. Nucleic Acids Res. 2017, 45, W36–W41. [Google Scholar] [CrossRef] [PubMed]
- Pullan, S.T.; Chandra, G.; Bibb, M.J.; Merrick, M. Genome-wide analysis of the role of GlnR in Streptomyces venezuelae provides new insights into global nitrogen regulation in actinomycetes. BMC Genom. 2011, 12, 175. [Google Scholar] [CrossRef] [PubMed]
- Heckman, K.L.; Pease, L.R. Gene splicing and mutagenesis by PCR-driven overlap extension. Nat. Protoc. 2007, 2, 924–932. [Google Scholar] [CrossRef] [PubMed]
- Skellam, E.J.; Stewart, A.K.; Strangman, W.K.; Wright, J.L. Identification of micromonolactam, a new polyene macrocyclic lactam from two marine Micromonospora strains using chemical and molecular methods: Clarification of the biosynthetic pathway from a glutamate starter unit. J. Antibiot. 2013, 66, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Kato, J.-Y.; Funa, N.; Watanabe, H.; Ohnishi, Y.; Horinouchi, S. Biosynthesis of γ-butyrolactone autoregulators that switch on secondary metabolism and morphological development in Streptomyces. Proc. Natl. Acad. Sci. USA 2007, 104, 2378–2383. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, N.-H.; Söding, J.; Linke, D.; Lange, C.; Hertweck, C.; Wohlleben, W.; Takano, E. ScbA from Streptomyces coelicolor A3(2) has homology to fatty acid synthases and is able to synthesize γ-butyrolactones. Microbiology 2007, 153, 1394–1404. [Google Scholar] [CrossRef] [PubMed]
- Kawachi, R.; Akashi, T.; Kamitani, Y.; Sy, A.; Wangchaisoonthorn, U.; Nihira, T.; Yamada, Y. Identification of an AfsA homologue (BarX) from Streptomyces virginiae as a pleiotropic regulator controlling autoregulator biosynthesis, virginiamycin biosynthesis and virginiamycin M1 resistance. Mol. Microbiol. 2000, 36, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Bibb, M. The regulation of antibiotic production in Streptomyces coelicolor A3(2). Microbiology 1996, 142, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ying, X.; Guo, Y.; Yu, Z.; Zhou, X.; Deng, Z.; Kieser, H.; Chater, K.F.; Tao, M. Identification of a gene negatively affecting antibiotic production and morphological differentiation in Streptomyces coelicolor A3(2). J. Bacteriol. 2006, 188, 8368–8375. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Guo, S.L.; Guo, W.Q.; Xi, D.; Xiang, W.S. Role of nsdA in negative regulation of antibiotic production and morphological differentiation in Streptomyces bingchengensis. J. Antibiot. 2009, 62, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Sola-Landa, A.; Moura, R.; Martin, J. The two-component phoR-phoP system controls both primary metabolism and secondary metabolite biosynthesis in Streptomyces lividans. Proc. Natl. Acad. Sci. USA 2003, 100, 6133–6138. [Google Scholar] [CrossRef] [PubMed]
- Asturias, J.A.; Martin, J.F.; Liras, P. Biosynthesis and phosphate control of candicidin by Streptomyces acrimycini Ji2236: Effect of amplification of the pabAB gene. J. Ind. Microbiol. 1994, 13, 183–189. [Google Scholar] [CrossRef] [PubMed]
- McDowall, K.J.; Thamchaipenet, A.; Hunter, I.S. Phosphate control of oxytetracycline production by Streptomyces rimosus is at the level of transcription from promoters overlapped by tandem repeats similar to those of the DNA-binding sites of the ompr family. J. Bacteriol. 1999, 181, 3025–3032. [Google Scholar] [PubMed]
- Mendes, M.V.; Tunca, S.; Antón, N.; Recio, E.; Sola-Landa, A.; Aparicio, J.F.; Martín, J.F. The two-component phoR-phoP system of Streptomyces natalensis: Inactivation or deletion of phoP reduces the negative phosphate regulation of pimaricin biosynthesis. Metab. Eng. 2007, 9, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Hesketh, A.; Sun, J.; Bibb, M. Induction of ppGpp synthesis in Streptomyces coelicolor A3(2) grown under conditions of nutritional sufficiency elicits actII-ORF4 transcription and actinorhodin biosynthesis. Mol. Microbiol. 2001, 39, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.G.; Jin, W.; Bibb, M.; Lee, K.J. Actinorhodin and undecylprodigiosin production in wild-type and relA mutant strains of Streptomyces coelicolor A3(2) grown in continuous culture. FEMS Microbiol. Lett. 1998, 168, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Takano, E. γ-butyrolactones: Streptomyces signalling molecules regulating antibiotic production and differentiation. Curr. Opin. Microbiol. 2006, 9, 287–294. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cluster | Type | GenBank ID | Location | Most Similar Known Biosynthetic Gene Cluster | Similarity (%) * | |
|---|---|---|---|---|---|---|
| From | To | |||||
| 1 | Siderophore | LNSV01000002 | 180883 | 189602 | Macrotetrolide | 33 |
| 2 | Siderophore | LNSV01000023 | 1 | 11484 | Desferrioxamine B | 100 |
| 3 | Siderophore | LNSV01000114 | 10308 | 18860 | - | - |
| 4 | Terpene | LNSV01000004 | 55721 | 76761 | - | - |
| 5 | Terpene | LNSV01000004 | 119146 | 140210 | - | - |
| 6 | Terpene | LNSV01000006 | 39446 | 61665 | - | - |
| 7 | Terpene | LNSV01000031 | 32044 | 58653 | Hopene | 61 |
| 8 | Terpene | LNSV01000035 | 41851 | 62261 | - | - |
| 9 | Terpene | LNSV01000039 | 39913 | 59882 | Isorenieratene | 85 |
| 10 | Phenazine | LNSV01000013 | 37297 | 57785 | Marinophenazines | 30 |
| 11 | Phosphonate | LNSV01000030 | 1 | 34405 | Azinomycin B | 6 |
| 12 | Bacteriocin | LNSV01000030 | 45699 | 57147 | Enduracidin | 6 |
| 13 | Ectoine | LNSV01000074 | 2911 | 13309 | Ectoine | 100 |
| 14 | Lassopeptide | LNSV01000003 | 18630 | 41325 | Collismycin A | 7 |
| 15 | Thiopeptide | LNSV01000094 | 1 | 22641 | Granaticin | 8 |
| 16 | T1pks | LNSV01000017 | 13669 | 61120 | Monensin | 5 |
| 17 | T1pks-Otherks | LNSV01000001 | 32321 | 84798 | - | - |
| 18 | T1pks-Butyrolactone-NRPs | LNSV01000041 | 1 | 58607 | Micromonolactam | 100 |
| 19 | T3pks | LNSV01000059 | 19531 | 42521 | Merochlorin | 4 |
| 20 | Otherks-Phenazine | LNSV01000010 | 440 | 49063 | Esmeraldin | 64 |
| 21 | NRPs | LNSV01000098 | 1 | 22529 | - | - |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, G.; Yu, D.; Sang, B.; Feng, J.; Han, L.; Zhang, X. Genome-Wide Analysis Reveals the Secondary Metabolome in Streptomyces kanasensis ZX01. Genes 2017, 8, 346. https://doi.org/10.3390/genes8120346
Zhang G, Yu D, Sang B, Feng J, Han L, Zhang X. Genome-Wide Analysis Reveals the Secondary Metabolome in Streptomyces kanasensis ZX01. Genes. 2017; 8(12):346. https://doi.org/10.3390/genes8120346
Chicago/Turabian StyleZhang, Guoqiang, Dailin Yu, Bu Sang, Juntao Feng, Lirong Han, and Xing Zhang. 2017. "Genome-Wide Analysis Reveals the Secondary Metabolome in Streptomyces kanasensis ZX01" Genes 8, no. 12: 346. https://doi.org/10.3390/genes8120346
APA StyleZhang, G., Yu, D., Sang, B., Feng, J., Han, L., & Zhang, X. (2017). Genome-Wide Analysis Reveals the Secondary Metabolome in Streptomyces kanasensis ZX01. Genes, 8(12), 346. https://doi.org/10.3390/genes8120346