Genome-Wide Analysis of the GRF Family Reveals Their Involvement in Abiotic Stress Response in Cassava

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Identification and Evolutionary Analysis
2.3. Transcriptome Analysis
2.4. Sequence Analysis
2.5. qRT-PCR Analysis
3. Results
3.1. Identification and Phylogenetic Analysis of GRF Gene Family in Cassava
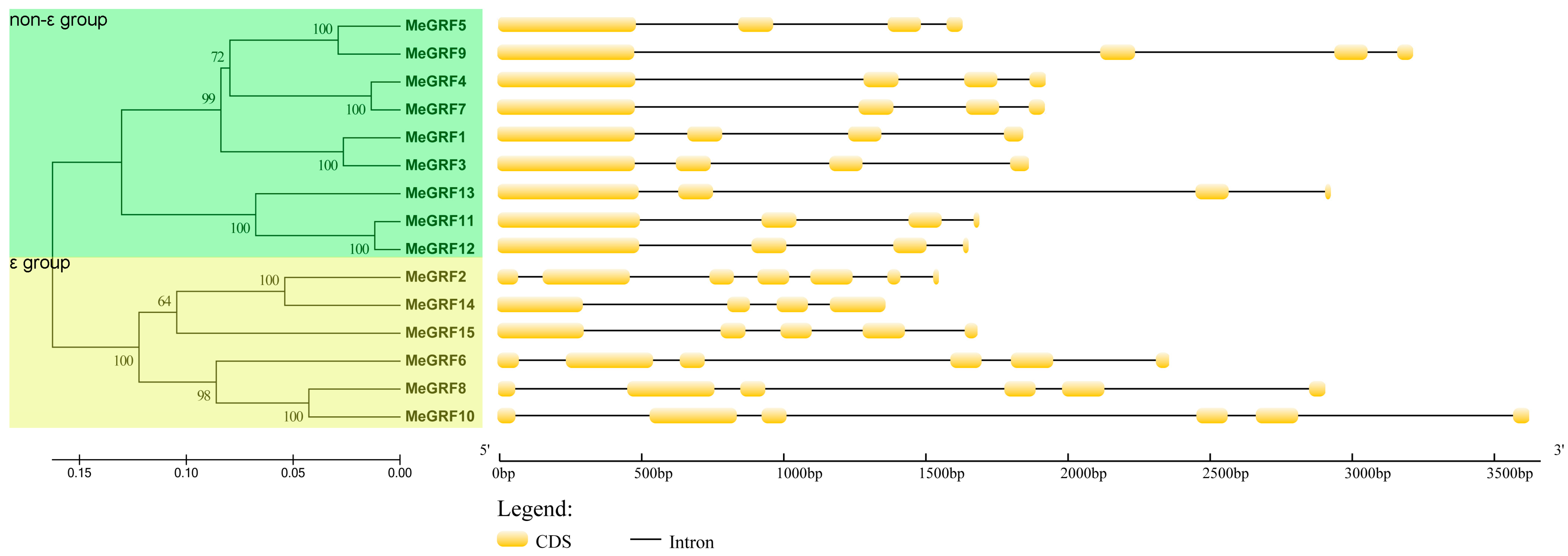
3.2. Conserved Motif and Gene Structure Analyses of GRF Gene Family in Cassava
3.3. Expression Profiles of MeGRF Genes in Different Tissues
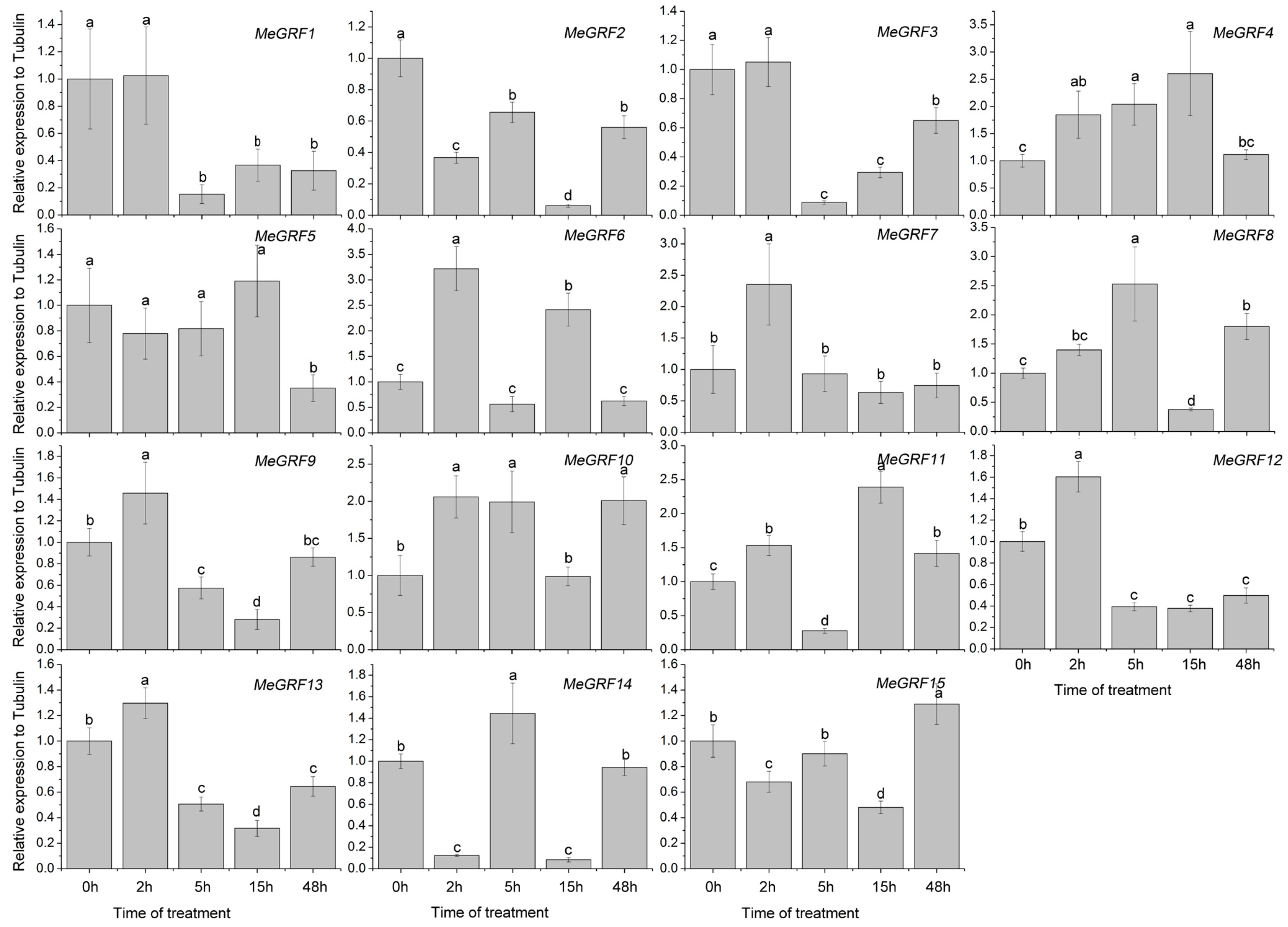
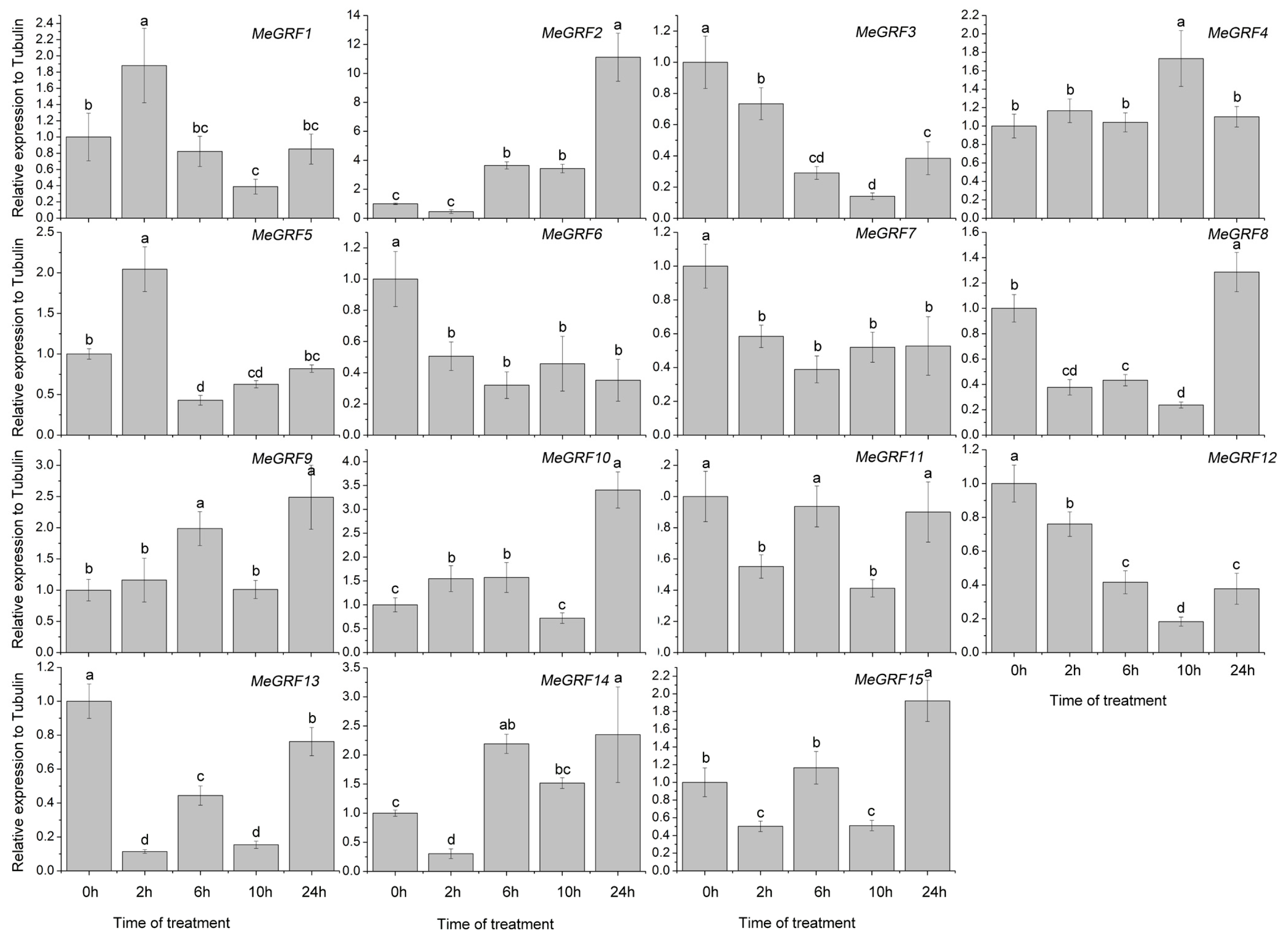
3.4. Differential Expression of MeGRF Genes in Response to Cold, Osmotic, Salt, ABA, and H2O2 Treatments
3.5. Analysis of GRF Family Interaction Network
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sehnke, P.C.; DeLille, J.M.; Ferl, R.J. Consummating signal transduction: the role of 14–3–3 proteins in the completion of signal-induced transitions in protein activity. Plant Cell 2002, 14l, S339–354. [Google Scholar] [CrossRef]
- Chevalier, D.; Morris, E.R.; Walker, J.C. 14–3–3 and FHA domains mediate phosphoprotein interactions. Annu. Rev. Plant Biol. 2009, 60, 67–91. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.L.; Denison, F.C.; Schultz, E.R.; Zupanska, A.K.; Ferl, R.J. 14–3–3 phosphoprotein interaction networks—Does isoform diversity present functional interaction specification? Front. Plant. Sci 2012, 3, 190. [Google Scholar] [CrossRef] [PubMed]
- Cotelle, V.; Leonhardt, N. 14–3–3 Proteins in Guard Cell Signaling. Front. Plant. Sci 2016, 6, 1210. [Google Scholar] [CrossRef] [PubMed]
- De Boer, A.H.; van Kleeff, P.J.; Gao, J. Plant 14–3–3 proteins as spiders in a web of phosphorylation. Protoplasma 2013, 250, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ren, L.; Xu, B.; Yang, X.; Xia, Q.; He, P.; Xiao, S.; Guo, A.; Hu, W.; Jin, Z. Genome-Wide Identification, Phylogeny, and Expression Analyses of the 14–3–3 Family Reveal Their Involvement in the Development, Ripening, and Abiotic Stress Response in Banana. Front. Plant. Sci. 2016, 7, 1442. [Google Scholar] [CrossRef] [PubMed]
- Saponaro, A.; Porro, A.; Chaves-Sanjuan, A.; Nardini, M.; Rauh, O.; Thiel, G.; Moroni, A. Fusicoccin Activates KAT1 Channels by Stabilizing Their Interaction with 14–3–3 Proteins. Plant. Cell. 2017, 29, 2570–2580. [Google Scholar] [CrossRef] [PubMed]
- De Vetten, N.C.; Lu, G.; Feri, R.J. A maize protein associated with the G-box binding complex has homology to brain regulatory proteins. Plant Cell 1992, 4, 1295–1307. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Li, Q.; Sun, L.; He, Z. The rice 14–3–3 gene family and its involvement in responses to biotic and abiotic stress. DNA Res. 2006, 13, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Du, Y.; Jiang, L.; Liu, J.Y. Molecular analysis and expression patterns of the 14–3–3 gene family from Oryza sativa. J. Biochem. Mol. Biol. 2007, 40, 349–357. [Google Scholar] [CrossRef] [PubMed]
- DeLille, J.M.; Sehnke, P.C.; Ferl, R.J. The Arabidopsis 14–3–3 family of signaling regulators. Plant. Physiol. 2001, 126, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Rosenquist, M.; Alsterfjord, M.; Larsson, C.; Sommarin, M. Data mining the Arabidopsis genome reveals fifteen 14–3–3 genes. Expression is demonstrated for two out of five novel genes. Plant. Physiol. 2001, 127, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Dhaubhadel, S. Soybean 14–3–3 gene family: Identification and molecular characterization. Planta 2011, 233, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Xie, F.; Zhang, B. Transcriptome-wide identification and stress properties of the 14–3–3 gene family in cotton (Gossypium hirsutum L.). Funct. Integr. Genom. 2011, 11, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Jiang, X.; Jin, D.; Dhaubhadel, S.; Bian, S.; Li, X. Identification of 14–3–3 Family in Common Bean and Their Response to Abiotic Stress. PLoS ONE 2015, 10, e0143280. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.F.; Shi, W.M. Expression profiling of the 14–3–3 gene family in response to salt stress and potassium and iron deficiencies in young tomato (Solanum lycopersicum) roots: analysis by real-time RT-PCR. Ann. Bot 2006, 98, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Wang, T.; Xie, Y.; Zhang, J.; Hu, J. Genome-wide identification, classification, and expression analysis of 14–3–3 gene family in Populus. PLoS ONE 2015, 10, e0123225. [Google Scholar] [CrossRef] [PubMed]
- Chandna, R.; Augustine, R.; Kanchupati, P.; Kumar, R.; Kumar, P.; Arya, G.C.; Bisht, N.C. Class-Specific Evolution and Transcriptional Differentiation of 14–3–3 Family Members in Mesohexaploid Brassica rapa. Front. Plant. Sci. 2016, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhang, Y.; Chen, L.; Wu, C.; Luo, Q.; Zhang, F.; Wei, Q.; Li, K.; Chang, J.; Yang, G.; et al. Member of the 14-3–3 Gene Family in Brachypodium distachyon, BdGF14d, Confers Salt Tolerance in Transgenic Tobacco Plants. Front. Plant. Sci. 2017, 8, 340. [Google Scholar] [CrossRef] [PubMed]
- Keicher, J.; Jaspert, N.; Weckermann, K.; Moller, C.; Throm, C.; Kintzi, A.; Oecking, C. Arabidopsis 14–3–3 ε members contribute to polarity of PIN auxin carrier and auxin transport-related development. Elife 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Vercruyssen, L.; Tognetti, V.B.; Gonzalez, N.; Van Dingenen, J.; De Milde, L.; Bielach, A.; De Rycke, R.; Van Breusegem, F.; Inze, D. GROWTH REGULATING FACTOR5 stimulates Arabidopsis chloroplast division, photosynthesis, and leaf longevity. Plant. Physiol. 2015, 167, 817–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Kleeff, P.J.; Jaspert, N.; Li, K.W.; Rauch, S.; Oecking, C.; de Boer, A.H. Higher order Arabidopsis 14–3–3 mutants show 14–3–3 involvement in primary root growth both under control and abiotic stress conditions. J. Exp. Bot. 2014, 65, 5877–5888. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wu, J.; Lv, B.; Li, J.; Gao, Z.; Xu, W.; Baluska, F.; Shi, W.; Shaw, P.C.; Zhang, J. Involvement of 14–3–3 protein GRF9 in root growth and response under polyethylene glycol-induced water stress. J. Exp. Bot. 2015, 66, 2271–2281. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Luo, X.; Sun, M.; Chen, C.; Ding, X.; Wang, X.; Yang, S.; Yu, Q.; Jia, B.; Ji, W.; et al. Glycine soja 14–3–3 protein GsGF14o participates in stomatal and root hair development and drought tolerance in Arabidopsis thaliana. Plant. Cell. Physiol. 2014, 55, 99–118. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, Z.T.; Li, M.; Wei, X.Z.; Li, X.J.; Li, B.Y.; Li, X.B. Cotton (Gossypium hirsutum) 14–3–3 proteins participate in regulation of fibre initiation and elongation by modulating brassinosteroid signalling. Plant Biotechnol. J. 2015, 13, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, X.; Chang, S.; Chu, Z.; Wang, H.; Han, S.; Wang, Y. Calcium-dependent protein kinase 21 phosphorylates 14–3–3 proteins in response to ABA signaling and salt stress in rice. Biochem. Biophys. Res. Commun. 2017, 493, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Kaundal, A.; Ramu, V.S.; Oh, S.; Lee, S.; Pant, B.; Lee, H.K.; Rojas, C.M.; Senthil-Kumar, M.; Mysore, K.S. GENERAL CONTROL NONREPRESSIBLE4 Degrades 14–3–3 and the RIN4 Complex to Regulate Stomatal Aperture with Implications on Nonhost Disease Resistance and Drought Tolerance. Plant Cell 2017, 29, 2233–2248. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; You, J.; Wang, Y.; Li, J.; Quan, W.; Yin, M.; Wang, Q.; Chan, Z. Systematic analysis of the G-box Factor 14–3–3 gene family and functional characterization of GF14a in Brachypodium distachyon. Plant Physiol. Biochem. 2017, 117, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Jia, Y.; Ding, Y.; Shi, Y.; Li, Z.; Guo, Y.; Gong, Z.; Yang, S. Plasma Membrane CRPK1-Mediated Phosphorylation of 14–3–3 Proteins Induces Their Nuclear Import to Fine-Tune CBF Signaling during Cold Response. Mol. Cell 2017, 66, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Zidenga, T.; Leyva-Guerrero, E.; Moon, H.; Siritunga, D.; Sayre, R. Extending Cassava root shelf life via reduction of reactive oxygen species production. Plant Physiol. 2012, 159, 1396–1407. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, W.Q.; Zhang, G.L.; Kaminek, M.; Dobrev, P.; Xu, J.; Gruissem, W. Senescence-inducible expression of isopentenyl transferase extends leaf life, increases drought stress resistance and alters cytokinin metabolism in Cassava. J. Integr. Plant Biol. 2010, 52, 653–669. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Duan, X.; Yang, J.; Beeching, J.R.; Zhang, P. Enhanced reactive oxygen species scavenging by overproduction of superoxide dismutase and catalase delays postharvest physiological deterioration of Cassava storage roots. Plant Physiol. 2013, 161, 1517–1528. [Google Scholar] [CrossRef] [PubMed]
- Kawahara, Y.; de la Bastide, M.; Hamilton, J.P.; Kanamori, H.; McCombie, W.R.; Ouyang, S.; Schwartz, D.C.; Tanaka, T.; Wu, J.; Zhou, S.; et al. Improvement of the Oryza sativa Nipponbare reference genome using next generation sequence and optical map data. Rice 2013, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium. UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar] [CrossRef]
- Prochnik, S.; Marri, P.R.; Desany, B.; Rabinowicz, P.D.; Kodira, C.; Mohiuddin, M.; Rodriguez, F.; Fauquet, C.; Tohme, J.; Harkins, T.; et al. The Cassava Genome: Current Progress, Future Directions. Trop. Plant Biol. 2012, 5, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.N.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Hannon Lab. FASTX-Toolkit. Available online: http://hannonlab.cshl.edu/fastx_toolkit (accessed on 25 May 2014).
- Braham Bioinformatics. FastQC. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 17 September 2014).
- John Hopkins University. Center for Computational biology (CCB). TopHat. Available online: http://ccb.jhu.edu/software/tophat/index.shtml/ (accessed on 21 November 2014).
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [PubMed]
- Brown, P.; Baxter, L.; Hickman, R.; Beynon, J.; Moore, J.D.; Ott, S. MEME-LaB: Motif analysis in clusters. Bioinformatics 2013, 29, 1696–1697. [Google Scholar] [CrossRef] [PubMed]
- Mulder, N.; Apweiler, R. InterPro and InterProScan: Tools for protein sequence classification and comparison. Methods Mol. Biol. 2007, 396, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Cheng, L.; Shen, J.; Zhang, Y.; Cao, H.; Lu, D.; Shen, C. Genome-Wide Identification and Expression Analysis of the 14–3–3 Family Genes in Medicago truncatula. Front. Plant Sci. 2016, 7, 320. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, C.; Zhang, C.; Wang, N.; Lu, D.; Wang, J.; Zhang, S.; Wang, Z.X.; Ma, H.; Wang, X. Dual role of BKI1 and 14–3–3 s in brassinosteroid signaling to link receptor with transcription factors. Dev. Cell 2011, 21, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Gampala, S.S.; Kim, T.W.; He, J.X.; Tang, W.; Deng, Z.; Bai, M.Y.; Guan, S.; Lalonde, S.; Sun, Y.; Gendron, J.M.; et al. An essential role for 14–3–3 proteins in brassinosteroid signal transduction in Arabidopsis. Dev. Cell 2007, 13, 177–189. [Google Scholar] [CrossRef] [PubMed]










© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shang, S.; Wu, C.; Huang, C.; Tie, W.; Yan, Y.; Ding, Z.; Xia, Z.; Wang, W.; Peng, M.; Tian, L.; et al. Genome-Wide Analysis of the GRF Family Reveals Their Involvement in Abiotic Stress Response in Cassava. Genes 2018, 9, 110. https://doi.org/10.3390/genes9020110
Shang S, Wu C, Huang C, Tie W, Yan Y, Ding Z, Xia Z, Wang W, Peng M, Tian L, et al. Genome-Wide Analysis of the GRF Family Reveals Their Involvement in Abiotic Stress Response in Cassava. Genes. 2018; 9(2):110. https://doi.org/10.3390/genes9020110
Chicago/Turabian StyleShang, Sang, Chunlai Wu, Chao Huang, Weiwei Tie, Yan Yan, Zehong Ding, Zhiqiang Xia, Wenquan Wang, Ming Peng, Libo Tian, and et al. 2018. "Genome-Wide Analysis of the GRF Family Reveals Their Involvement in Abiotic Stress Response in Cassava" Genes 9, no. 2: 110. https://doi.org/10.3390/genes9020110
APA StyleShang, S., Wu, C., Huang, C., Tie, W., Yan, Y., Ding, Z., Xia, Z., Wang, W., Peng, M., Tian, L., & Hu, W. (2018). Genome-Wide Analysis of the GRF Family Reveals Their Involvement in Abiotic Stress Response in Cassava. Genes, 9(2), 110. https://doi.org/10.3390/genes9020110