Mutation in the pssZ Gene Negatively Impacts Exopolysaccharide Synthesis, Surface Properties, and Symbiosis of Rhizobium leguminosarum bv. trifolii with Clover

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, and Growth Conditions
2.2. DNA Methods and Sequence Analysis
2.3. Isolation of a Rhizobium leguminosarum pssZ Mutant
2.4. Construction of Plasmid pPL1 for Complementation of the pssZ Mutation
2.5. β-glucuronidase Assay
2.6. Isolation and Quantification of Exopolysaccharide
2.7. Determination of Lipopolysaccharide Profiles
2.8. Cell Hydrophobicity Assay
2.9. Aggreagation Assay
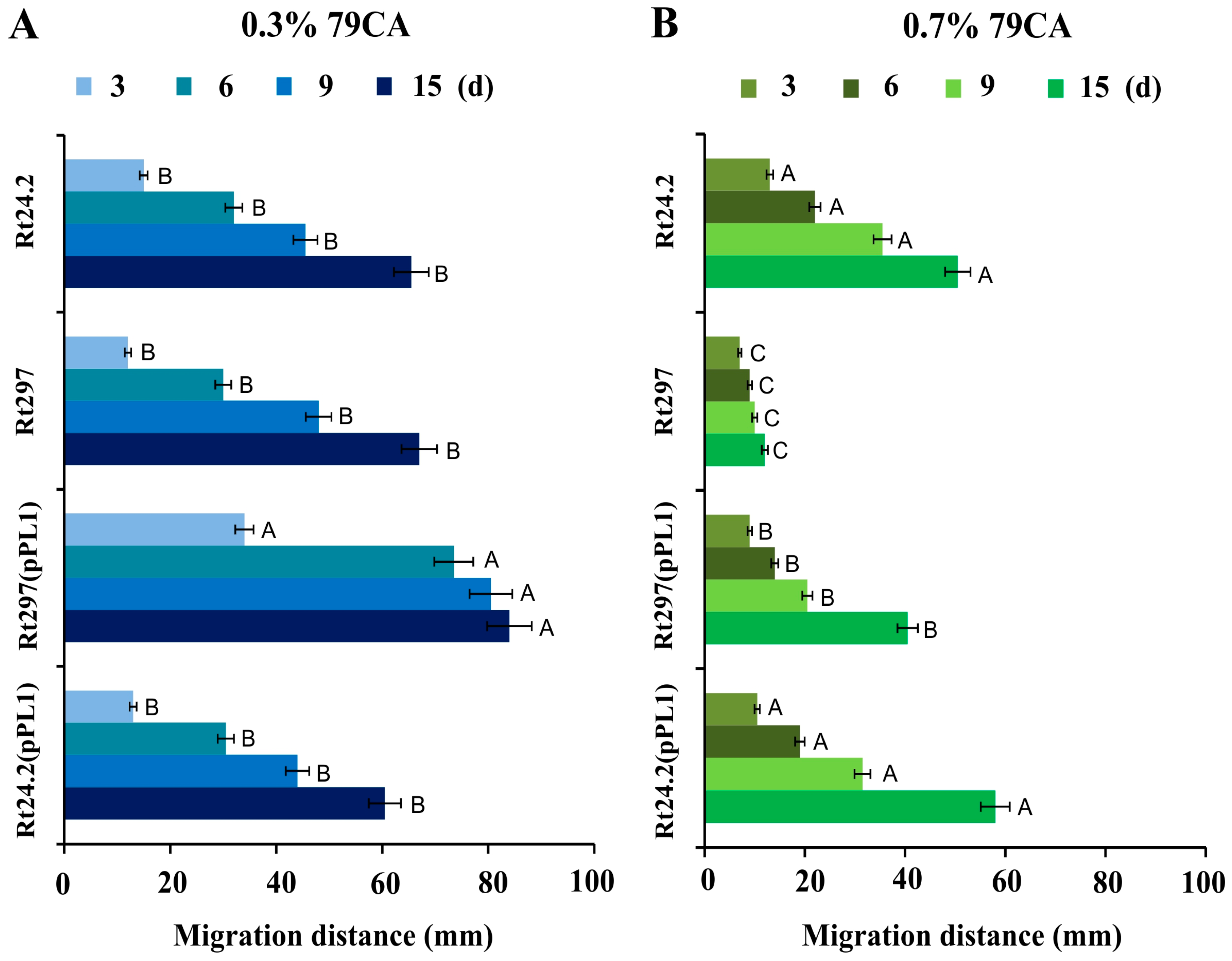
2.10. Determination of Cell Motility
2.11. Determination of Rhizobial Sensitivity to Stress Factors
2.12. Biofilm Production Assay
2.13. Determination of Cell Topology and Properties Using Atomic Force Microscopy
2.14. Plant Experiments
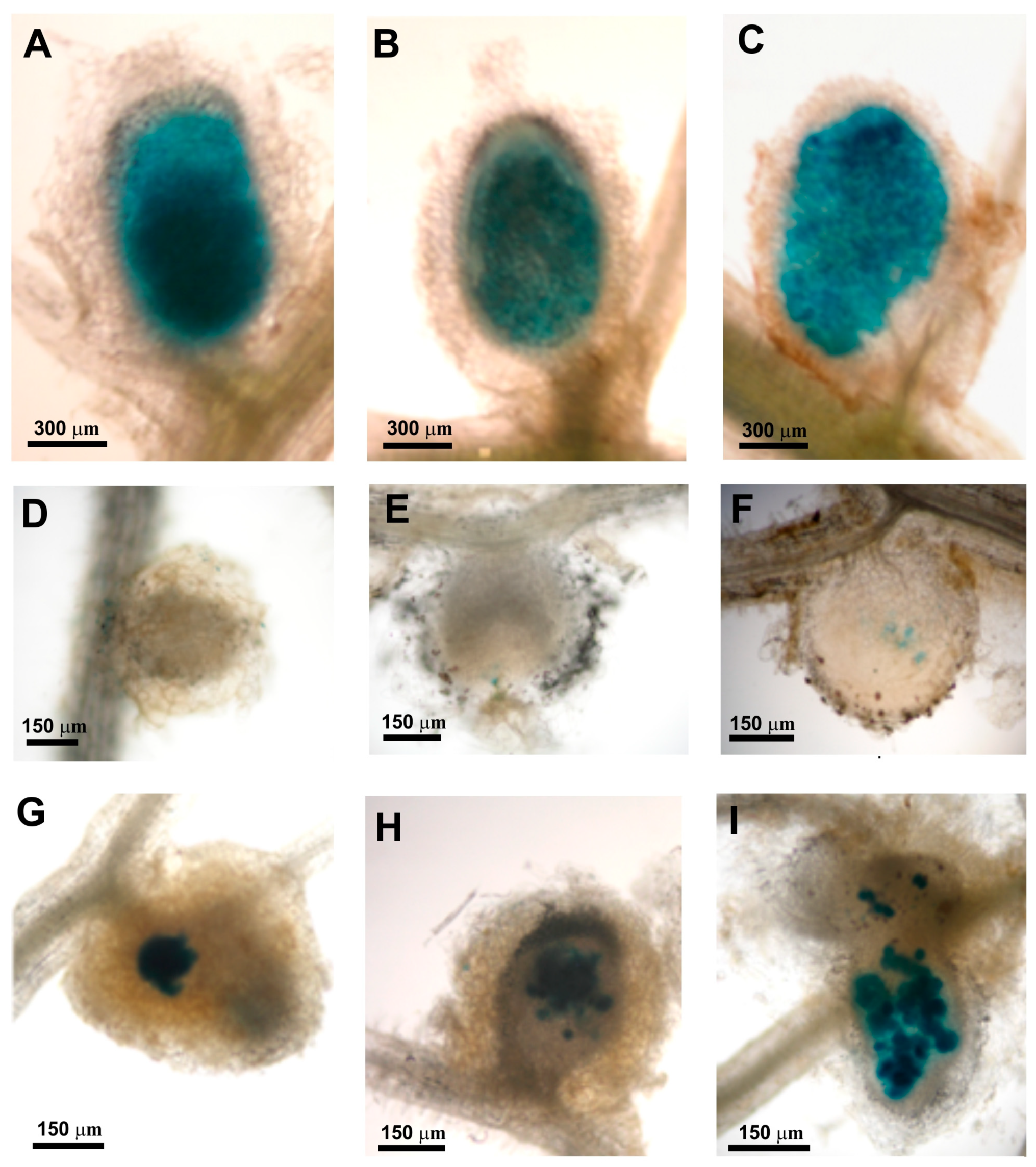
2.15. Nodule Analysis Using Light and Electron Microscopy
2.16. Statistical Analysis
3. Results
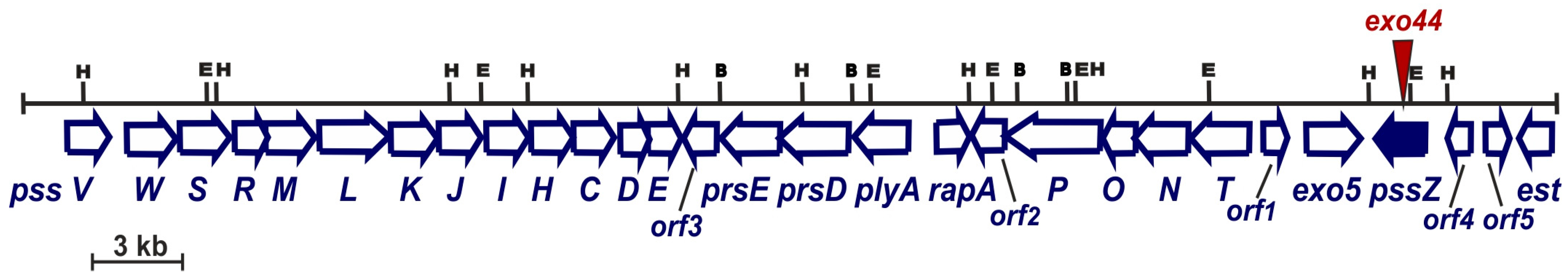
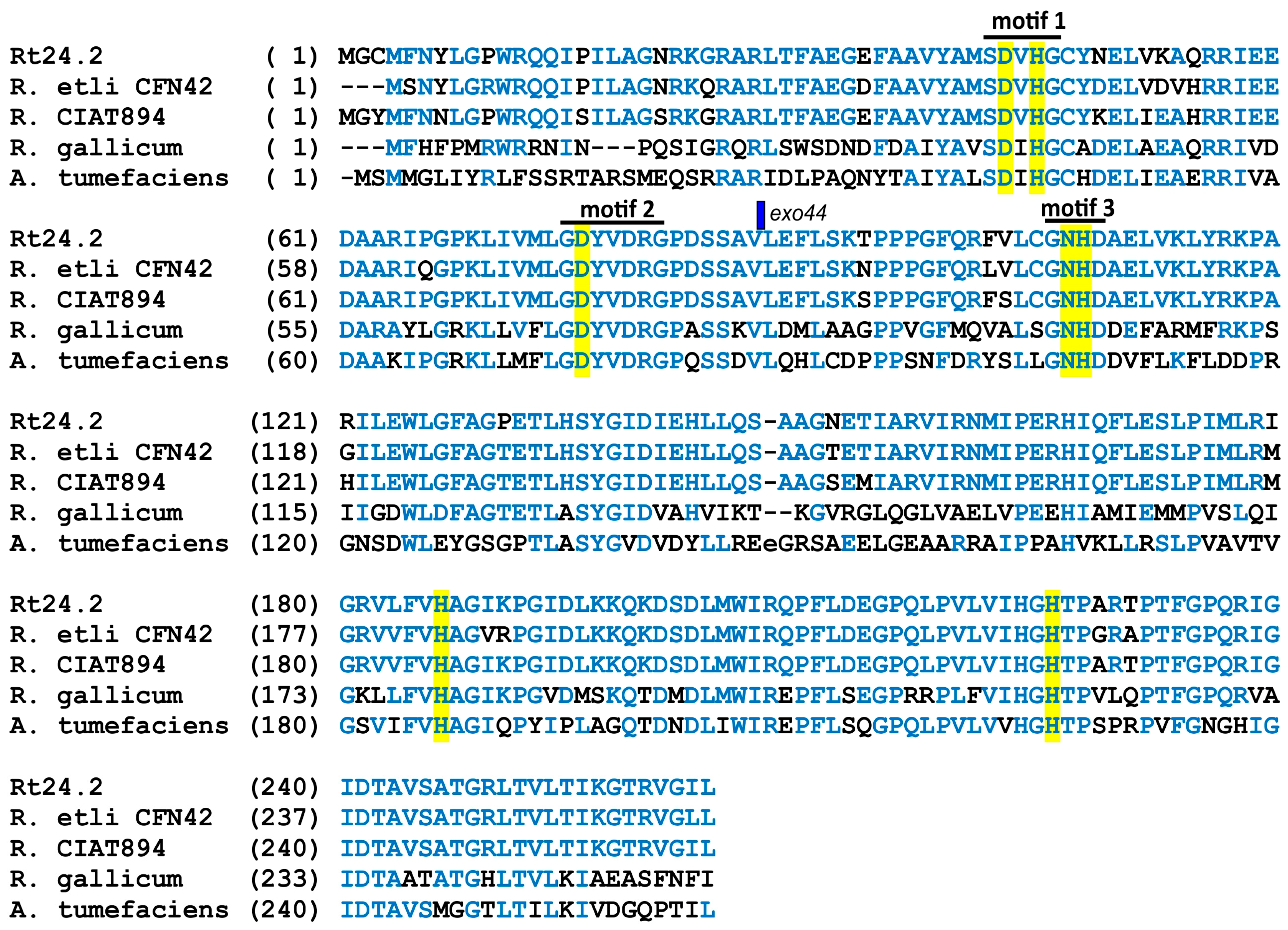
3.1. Genetic Characterization of a Mutant Strain Rt297 and Complementation of a pssZ Mutation
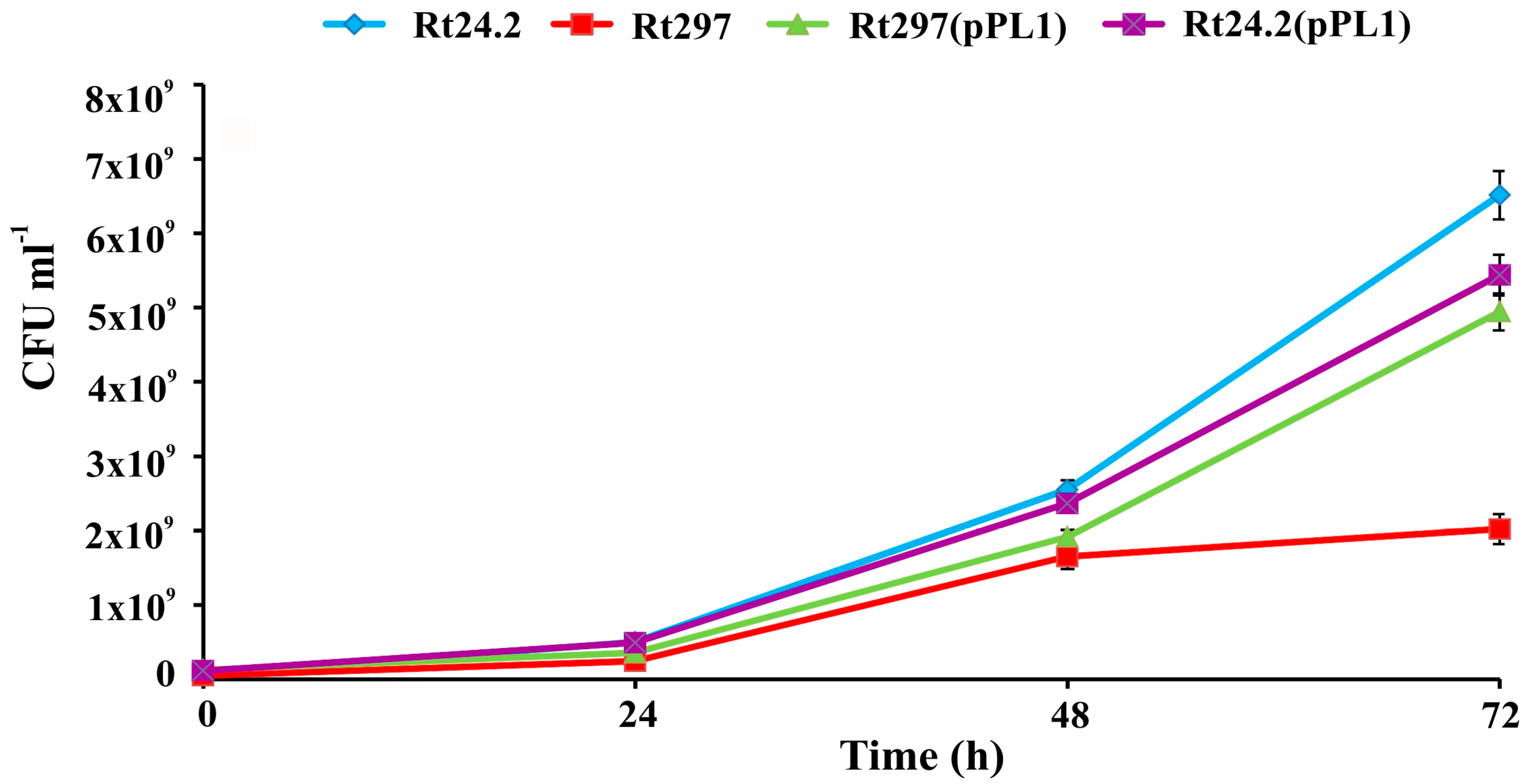
3.2. The exo44 Mutation Negatively Affects Growth and Motility of Rhizobium leguminosarum
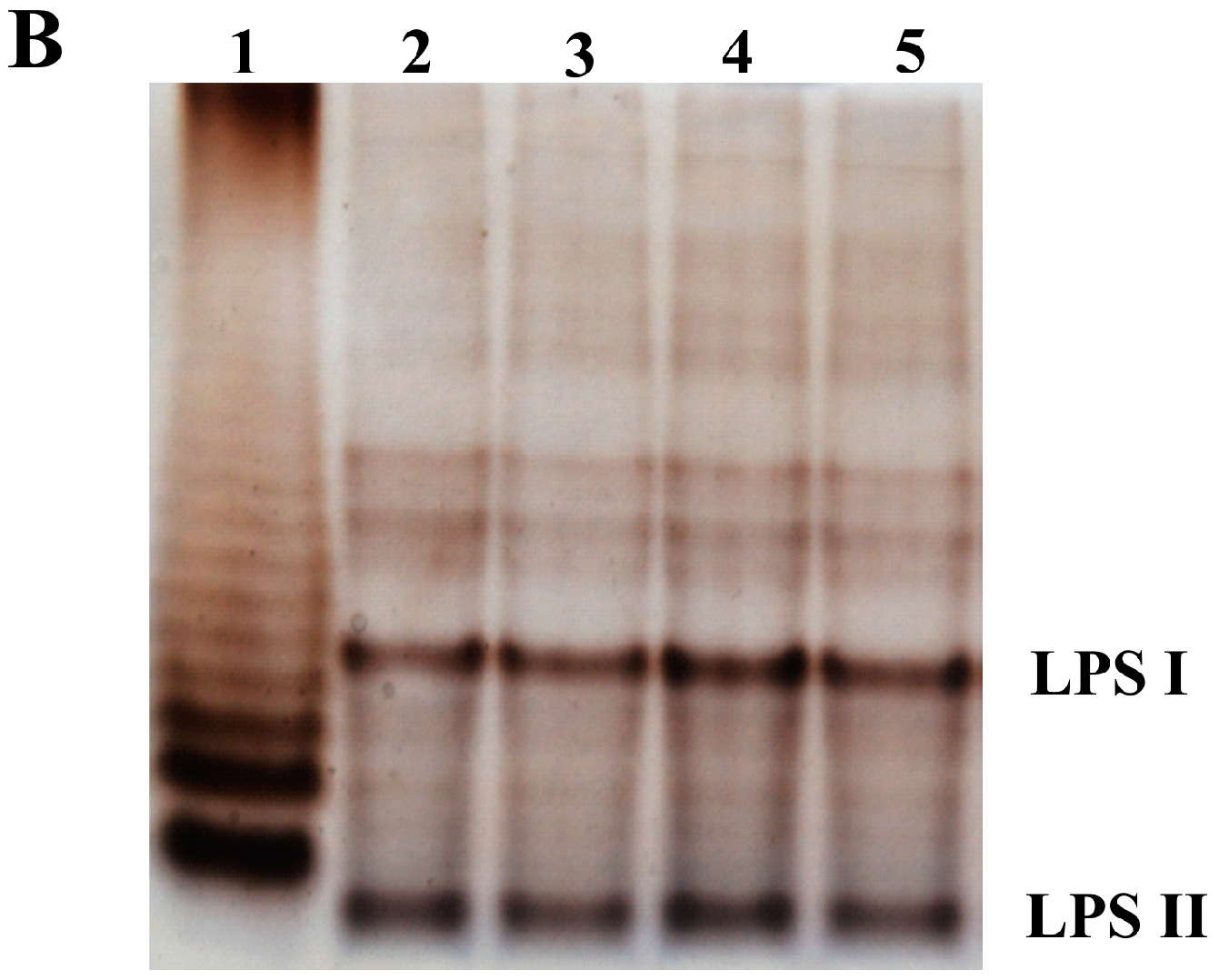
3.3. The exo44 Mutation Affects Surface Properties of Rhizobium leguminosarum Cells
3.4. The exo44 Mutation Leads to Disturbances in Symbiosis of Rhizobium leguminosarum with Clover
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Haag, A.F.; Arnold, M.F.F.; Myka, K.K.; Kerscher, B.; Dall’Angelo, S.; Zanda, M.; Mergaert, P.; Ferguson, G.P. Molecular insights into bacteroid development during Rhizobium-legume symbiosis. FEMS Microbiol. Rev. 2013, 37, 364–383. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Guerrero, I.; Acosta-Jurado, S.; Del Cerro, P.; Navarro-Gómez, P.; López-Baena, F.J.; Ollero, F.J.; Vinardell, J.M.; Pérez-Montaño, F. Transcriptomic studies of the effect of nod gene-inducing molecules in rhizobia: Different weapons, one purpose. Genes 2018, 9, 1. [Google Scholar] [CrossRef] [PubMed]
- Janczarek, M.; Rachwał, K.; Marzec, A.; Grządziel, J.; Palusińska-Szysz, M. Signal molecules and cell-surface components involved in early stages of the legume-rhizobium interactions. Appl. Soil Ecol. 2015, 85, 94–113. [Google Scholar] [CrossRef]
- Marczak, M.; Mazur, A.; Koper, P.; Żebracki, K.; Skorupska, A. Synthesis of rhizobial exopolysaccharides and their importance for symbiosis with legume plants. Genes 2017, 8, 360. [Google Scholar] [CrossRef] [PubMed]
- Janczarek, M.; Skorupska, A. Modulation of rosR expression and exopolysaccharide production in Rhizobium leguminosarum bv. trifolii by phosphate and clover root exudates. Int. J. Mol. Sci. 2011, 12, 4132–4155. [Google Scholar] [CrossRef] [PubMed]
- Ivashina, T.V.; Khmelnitsky, M.I.; Shlyapnikov, M.G.; Kanapin, A.A.; Ksenzenko, V.N. The pss4 gene from Rhizobium leguminosarum biovar viciae VF39: Cloning, sequence and the possible role in polysaccharide production and nodule formation. Gene 1994, 50, 111–116. [Google Scholar] [CrossRef]
- Van Workum, W.A.; Canter Cremers, H.C.; Wijfjes, A.H.; van der Kolk, C.; Wijffelman, C.A.; Kijne, J.W. Cloning and characterization of four genes of Rhizobium leguminosarum bv. trifolii involved in exopolysaccharide production and nodulation. Mol. Plant Microbe Interact. 1997, 10, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.P.; Walker, G.C. Succinoglycan is required for initiation and elongation of the infection threads during nodulation of alfalfa by Rhizobium meliloti. J. Bacteriol. 1998, 180, 5183–5191. [Google Scholar] [PubMed]
- Janczarek, M.; Urbanik-Sypniewska, T. Expression of the Rhizobium leguminosarum bv. trifolii pssA gene involved in exopolysaccharide synthesis is regulated by RosR, phosphate and the carbon source. J. Bacteriol. 2013, 195, 3412–3423. [Google Scholar] [CrossRef] [PubMed]
- Margaret-Oliver, I.; Lei, W.; Parada, M.; Rodríguez-Carvajal, M.A.; Crespo-Rivas, J.C.; Hidalgo, Á.; Gil-Serrano, A.; Moreno, J.; Rodríguez-Navarro, D.N.; Buendía-Clavería, A.; et al. Sinorhizobium fredii HH103 does not strictly require KPS and/or EPS to nodulate Glycyrrhiza uralensis, an indeterminate nodule-forming legume. Arch. Microbiol. 2012, 194, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Rivas, J.C.; Guefrachi, I.; Mok, K.C.; Villaécija-Aguilar, J.A.; Acosta-Jurado, S.; Pierre, O.; Ruiz-Sainz, J.E.; Taga, M.E.; Mergaert, P.; Vinardell, J.M. Sinorhizobium fredii HH103 bacteroids are not terminally differentiated and show altered O-antigen in nodules of the IRLC legume Glycyrrhiza uralensis. Environ. Microbiol. 2016, 8, 2392–2404. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Navarro, D.N.; Rodríguez-Carvajal, M.A.; Acosta-Jurado, S.; Soto, M.J.; Margaret, I.; Crespo-Rivas, J.C.; Sanjuan, J.; Temprano, F.; Gil-Serrano, A.; Ruiz-Sainz, J.E.; et al. Structure and biological roles of Sinorhizobium fredii HH103 exopolysaccharide. PLoS ONE 2014, 18, e115391. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.J.; Muszyński, A.; Kawaharada, Y.; Hubber, A.M.; Sullivan, J.T.; Sandal, N.; Carlson, R.W.; Stougaard, J.; Ronson, C.W. Conditional requirement for exopolysaccharide in the Mesorhizobium-Lotus symbiosis. Mol. Plant Microbe Interact. 2013, 26, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Muszyński, A.; Heiss, C.; Hjuler, C.T.; Sullivan, J.T.; Kelly, S.J.; Thygesen, M.B.; Stougaard, J.; Azadi, P.; Carlson, R.W.; Ronson, C.W. Structures of exopolysaccharides involved in receptor-mediated perception of Mesorhizobium loti by Lotus japonicus. J. Biol. Chem. 2016, 291, 20946–20961. [Google Scholar] [CrossRef] [PubMed]
- Kawaharada, Y.; Kelly, S.; Nielsen, M.W.; Hjuler, C.T.; Gysel, K.; Muszyński, A.; Carlson, R.W.; Thygesen, M.B.; Sandal, N.; Asmussen, M.H.; et al. Receptor-mediated exopolysaccharide perception controls bacterial infection. Nature 2015, 523, 308–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broos, K.; Beyens, H.; Smolders, E. Survival of rhizobia in soil is sensitive to elevated zinc in the absence of the host plant. Soil Biol. Biochem. 2005, 37, 573–579. [Google Scholar] [CrossRef]
- Janczarek, M.; Rachwał, K.; Cieśla, J.; Ginalska, G.; Bieganowski, A. Production of exopolysaccharide by Rhizobium leguminosarum bv. trifolii and its role in bacterial attachment and surface properties. Plant Soil 2015, 388, 211–227. [Google Scholar] [CrossRef]
- Robertsen, B.K.; Aman, P.; Darvill, A.G.; McNeil, M.; Albersheim, P. Host-symbiont interactions: V. The structure of acidic extracellular polysaccharides secreted by Rhizobium leguminosarum and Rhizobium trifolii. Plant Physiol. 1981, 67, 389–400. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, M.A.; Darvill, A.G.; Albersheim, P. The degree of esterification and points of substitution by O-acetyl and O-(3-hydroxybutanoyl) groups in the acidic extracellular polysaccharides secreted by Rhizobium leguminosarum biovars viciae, trifolii, and phaseoli are not related to host range. J. Biol. Chem. 1991, 266, 9549–9555. [Google Scholar] [PubMed]
- Philip-Hollingswort, S.; Hollingsworth, R.I.; Dazzo, F.B. Host-range related structural features of the acidic extracellular polysaccharides of Rhizobium trifolii and Rhizobium leguminosarum. J. Biol. Chem. 1989, 264, 1461–1466. [Google Scholar]
- Breedveld, M.W.; Cremers, H.C.; Batley, M.; Posthumus, M.A.; Zevenhuizen, L.P.; Wijffelman, C.A.; Zehnder, A.J. Polysaccharide synthesis in relation to nodulation behavior of Rhizobium leguminosarum. J. Bacteriol. 1993, 175, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Breedveld, M.W.; Zevenhuizen, L.P.; Canter Cremers, H.C.; Zehnder, A.J. Influence of growth conditions on production of capsular and extracellular polysaccharides by Rhizobium leguminosarum. Antonie Leeuwenhoek 1993, 64, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Pollock, T.J.; van Workum, W.A.; Thorne, L.; Mikolajczak, M.J.; Yamazaki, M.; Kijne, J.W.; Armentrout, R.W. Assignment of biochemical functions to glycosyl transferase genes which are essential for biosynthesis of exopolysaccharides in Sphingomonas strain S88 and Rhizobium leguminosarum. J. Bacteriol. 1998, 180, 586–593. [Google Scholar] [PubMed]
- Ivashina, T.V.; Ksenzenko, V.N. Exopolysaccharide biosynthesis in Rhizobium leguminosarum from genes to functions. In The Complex World of Polysaccharides; Karunaratne, D.N., Ed.; InTech: Rijeka, Croatia, 2012; pp. 99–127. ISBN 978-953-51-0819-1. [Google Scholar]
- Janczarek, M.; Rachwał, K.; Kopcińska, J. Genetic characterization of the Pss region and the role of PssS in exopolysaccharide production and symbiosis of Rhizobium leguminosarum bv. trifolii with clover. Plant Soil 2015, 396, 257–275. [Google Scholar] [CrossRef]
- Janczarek, M.; Rachwał, K.; Turska-Szewczuk, A. A mutation in pssE affects exopolysaccharide synthesis by Rhizobium leguminosarum bv. trifolii, its surface properties, and symbiosis with clover. Plant Soil 2017, 417, 331–347. [Google Scholar] [CrossRef]
- Ivashina, T.V.; Fedorova, E.E.; Ashina, N.P.; Kalinchuk, N.A.; Druzhinina, T.N.; Shashkov, A.S.; Shibaev, V.N.; Ksenzenko, V.N. Mutation in the pssM gene encoding ketal pyruvate transferase leads to disruption of Rhizobium leguminosarum bv. viciae-Pisum sativum symbiosis. J. Appl. Microbiol. 2010, 109, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Mazur, A.; Marczak, M.; Król, J.E.; Skorupska, A. Topological and transcriptional analysis of pssL gene product: A putative Wzx-like exopolysaccharide translocase in Rhizobium leguminosarum bv. trifolii TA1. Arch. Microbiol. 2005, 184, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Marczak, M.; Dźwierzyńska, M.; Skorupska, A. Homo- and heterotypic interactions between Pss proteins involved in the exopolysaccharide transport system in Rhizobium leguminosarum bv. trifolii. Biol. Chem. 2013, 394, 541–559. [Google Scholar] [CrossRef] [PubMed]
- Marczak, M.; Matysiak, P.; Kutkowska, J.; Skorupska, A. PssP2 is a polysaccharide co-polymerase involved in exopolysaccharide chain-length determination in Rhizobium leguminosarum. PLoS ONE 2014, 9, e109106. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.T.; Lam, J.S. Synthesis of bacterial polysaccharides via the Wzx/Wzy-dependent pathway. Can. J. Microbiol. 2014, 60, 697–716. [Google Scholar] [CrossRef] [PubMed]
- Schmid, J.; Sieber, V.; Rehm, B. Bacterial exopolysaccharides: Biosynthesis pathways and engineering strategies. Front. Microbiol. 2015, 6, 496. [Google Scholar] [CrossRef] [PubMed]
- Grangeasse, C.; Terreux, R.; Nessler, S. Bacterial tyrosine-kinases: Structure–function analysis and therapeutic potential. Biochim. Biophys. Acta 2010, 1804, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, D.; Becker, A. The molecular weight distribution of succinoglycan produced by Sinorhizobium meliloti is influenced by specific tyrosine phosphorylation and ATPase activity of the cytoplasmic domain of the ExoP protein. J. Bacteriol. 2001, 183, 5163–5170. [Google Scholar] [CrossRef] [PubMed]
- Tocilj, A.; Munger, C.; Proteau, A.; Morona, R.; Purins, L.; Ajamian, E.; Wagner, J.; Papadopoulos, M.; Van Den Bosch, L.; Rubinstein, J.L.; et al. Bacterial polysaccharide co-polymerases share a common framework for control of polymer length. Nat. Struct. Mol. Biol. 2008, 15, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Marczak, M.; Mazur, A.; Król, J.E.; Gruszecki, W.I.; Skorupska, A. Lipoprotein PssN of Rhizobium leguminosarum bv. trifolii: Subcellular localization and possible involvement in exopolysaccharide export. J. Bacteriol. 2006, 188, 6943–6952. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C. Biosynthesis and assembly of capsular polysaccharides in Escherichia coli. Annu. Rev. Biochem. 2006, 75, 39–68. [Google Scholar] [CrossRef] [PubMed]
- Janczarek, M.; Skorupska, A. The Rhizobium leguminosarum bv. trifolii RosR: Transcriptional regulator involved in exopolysaccharide production. Mol. Plant Microbe Interact. 2007, 20, 867–881. [Google Scholar] [CrossRef] [PubMed]
- Janczarek, M.; Jaroszuk-Ściseł, J.; Skorupska, A. Multiple copies of rosR and pssA genes enhance exopolysaccharide production, symbiotic competitiveness and clover nodulation in Rhizobium leguminosarum bv. trifolii. Antonie Leeuwenhoek 2009, 96, 471–486. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989; pp. 11–26, 94–144, 162–186. ISBN 1936113422. [Google Scholar]
- Simon, R.; Quandt, J.; Klipp, W. New derivatives of transposon Tn5 suitable for mobilization of replicons, generation of operon fusions and induction of genes in Gram-negative bacteria. Gene 1989, 80, 161–169. [Google Scholar] [CrossRef]
- Wilson, K.J.; Sessitsch, A.; Corbo, J.C.; Giller, K.E.; Akkermans, A.D.; Jefferson, R.A. Beta-Glucuronidase (GUS) transposons for ecological and genetic studies of rhizobia and other gram-negative bacteria. Microbiology 1995, 141, 1691–1705. [Google Scholar] [CrossRef] [PubMed]
- Kovach, M.E.; Elzer, P.H.; Hill, D.S.; Robertson, G.T.; Farris, M.A.; Roop, R.M.; Peterson, K.M. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar] [CrossRef]
- Wielbo, J.; Skorupska, A. Construction of improved vectors and cassettes containing gusA and antibiotic resistance genes for studies of transcriptional activity and bacterial localization. J. Microbiol. Methods 2001, 45, 197–205. [Google Scholar] [CrossRef]
- Kowalczuk, E.; Lorkiewicz, Z. Transfer of RP4 and R68.45 factors to Rhizobium. Acta Microbiol. Pol. 1979, 28, 221–229. [Google Scholar] [PubMed]
- FASTA. Available online: https://www.ebi.ac.uk/Tools/sss/fasta/ (accessed on 8 May 2018).
- BLAST. Available online: http://blast.ncbi.nlm.nih.gov/ (accessed on 8 May 2018).
- BDGP Neural Network Promoter Prediction. Available online: http://www.fruitfly.org/ (accessed on 8 May 2018).
- Malign Program. Available online: http://www.genebee.msu.su/services/malign/ (accessed on 8 May 2018).
- Fuzznuc Program. Available online: http://emboss.ch.embnet.org/Pise/ (accessed on 8 May 2018).
- MacLellan, S.R.; MacLean, A.M.; Finan, T.M. Promoter prediction in the rhizobia. Microbiology 2006, 152, 1751–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1972; ISBN 0879691069. [Google Scholar]
- Vanderlinde, E.M.; Yost, C.K. Mutation of the sensor kinase chvG in Rhizobium leguminosarum negatively impacts cellular metabolism, outer membrane stability, and symbiosis. J. Bacteriol. 2012, 194, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Loewus, F.A. Improvement in the anthrone method for determination of carbohydrates. Anal. Chem. 1952, 24, 219–220. [Google Scholar] [CrossRef]
- Lesse, A.J.; Campagnari, A.A.; Bittner, W.E.; Apicella, M.A. Increased resolution of lipopolysaccharides and lipooligosaccharides utilizing tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis. J. Immunol. Methods 1990, 126, 109–117. [Google Scholar] [CrossRef]
- Rachwał, K.; Boguszewska, A.; Kopcińska, J.; Karaś, M.; Tchórzewski, M.; Janczarek, M. The regulatory protein RosR affects Rhizobium leguminosarum bv. trifolii protein profiles, cell surface properties, and symbiosis with clover. Front. Microbiol. 2016, 7, 1302. [Google Scholar] [CrossRef] [PubMed]
- Sorroche, F.G.; Spesia, M.B.; Zorreguieta, A.; Giordano, W. A positive correlation between bacterial autoaggregation and biofilm formation in native Sinorhizobium meliloti isolates from Argentina. Appl. Environ. Microbiol. 2012, 78, 4092–4101. [Google Scholar] [CrossRef] [PubMed]
- Rachwał, K.; Matczyńska, E.; Janczarek, M. Transcriptome profiling of a Rhizobium leguminosarum bv. trifolii rosR mutant reveals the role of the transcriptional regulator RosR in motility, synthesis of cell-surface components, and other cellular processes. BMC Genom. 2015, 16, 1111. [Google Scholar] [CrossRef]
- Rinaudi, L.V.; González, J.E. The low-molecular-weight fraction of exopolysaccharide II from Sinorhizobium meliloti is a crucial determinant of biofilm formation. J. Bacteriol. 2009, 191, 7216–7224. [Google Scholar] [CrossRef] [PubMed]
- Zdybicka-Barabas, A.; Januszanis, B.; Mak, P.; Cytryńska, M. An atomic force microscopy study of Galleria mellonella apolipophorin III effect on bacteria. Biochim. Biophys. Acta 2011, 1808, 1896–1906. [Google Scholar] [CrossRef] [PubMed]
- Horcas, I.; Fernández, R.; Gómez-Rodríguez, J.M.; Colchero, J.; Gómez-Herrero, J.; Baro, A.M. WSXM: A software for scanning probe microscopy and a tool for nanotechnology. Rev. Sci. Instrum. 2007, 78, 013705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vincent, J.M. A Manual for the Practical Study of Root Nodule Bacteria; International Biological Program Handbook No. 15; Blackwell Scientific Publications: Oxford, UK, 1970; pp. 153–159. ISBN 0632064102. [Google Scholar]
- Janczarek, M.; Rachwał, K. Mutation in the pssA gene involved in exopolysaccharide synthesis leads to several physiological and symbiotic defects in Rhizobium leguminosarum bv. trifolii. Int. J. Mol. Sci. 2013, 14, 23711–23735. [Google Scholar] [CrossRef] [PubMed]
- Fujishige, NA.; Kapadia, N.N.; De Hoff, P.L.; Hirsch, A.M. Investigations of Rhizobium biofilm formation. FEMS Microbiol. Ecol. 2006, 56, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Jaszek, M.; Janczarek, M.; Kuczyński, K.; Piersiak, T.; Grzywnowicz, K. The response of the Rhizobium leguminosarum bv. trifolii wild-type and exopolysaccharide-deficient mutants to oxidative stress. Plant Soil 2014, 376, 75–94. [Google Scholar] [CrossRef]
- Sadykov, M.R.; Ivashina, T.V.; Kanapin, A.A.; Shliapnikov, M.G.; Ksenzenko, V.N. Structure-functional organization of exopolysaccharide biosynthetic genes in Rhizobium leguminosarum bv. viciae VF39. Mol. Biol. (Mosk.) 1998, 32, 797–804. [Google Scholar] [PubMed]
- Król, J.E.; Mazur, A.; Marczak, M.; Skorupska, A. Syntenic arrangements of the surface polysaccharide biosynthesis genes in Rhizobium leguminosarum. Genomics 2007, 89, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Laus, M.C.; Logman, T.J.; Van Brussel, A.A.; Carlson, R.W.; Azadi, P.; Gao, M.Y.; Kijne, J.W. Involvement of exo5 in production of surface polysaccharides in Rhizobium leguminosarum and its role in nodulation of Vicia sativa subsp. nigra. J. Bacteriol. 2004, 186, 6617–6625. [Google Scholar] [CrossRef] [PubMed]
- Muszyński, A.; Laus, M.; Kijne, J.W.; Carlson, R.W. Structures of the lipopolysaccharides from Rhizobium leguminosarum RBL5523 and its UDP-glucose dehydrogenase mutant (exo5). Glycobiology 2011, 21, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Kereszt, A.; Kiss, E.; Reuhs, B.L.; Carlson, R.W.; Kondorosi, A.; Putnoky, P. Novel rkp gene clusters of Sinorhizobium meliloti involved in capsular polysaccharide production and invasion of the symbiotic nodule: The rkpK gene codes a UDP-glucose dehydrogenase. J. Bacteriol. 1998, 180, 5426–5431. [Google Scholar] [PubMed]
- Acosta-Jurado, S.; Navarro-Gómez, P.; Crespo-Rivas, J.C.; Medina, C.; Murdoch, P.S.; Cuesta-Berrio, L.; Rodríguez-Carvajal, M.A.; Ruiz-Sainz, J.E.; Vinardell, J.M. The Sinorhizobium (Ensifer) fredii HH103 rkp-2 region is involved in the biosynthesis of lipopolysaccharide and exopolysaccharide but not in K-antigen polysaccharide production. Plant Soil 2017, 417, 415–431. [Google Scholar] [CrossRef]
- Soto, M.J.; Sanjuán, J.; Olivares, J. Rhizobia and plant-pathogenic bacteria: Common infection weapons. Microbiology 2006, 152, 3167–3174. [Google Scholar] [CrossRef] [PubMed]
- Libby, E.A.; Goss, L.A.; Dworkin, J. The eukaryotic-like Ser/Thr kinase PrkC regulates the essential WalRK Two-Component System in Bacillus subtilis. PLoS Genet. 2015, 11, e1005275. [Google Scholar] [CrossRef] [PubMed]
- Brautigan, D.L. Protein Ser/Thr phosphatases—The ugly ducklings of cell signaling. FEBS J. 2013, 280, 324–345. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y. Serine/threonine phosphatases: Mechanism through structure. Cell 2009, 139, 468–484. [Google Scholar] [CrossRef] [PubMed]
- Mijaković, I.; Macek, B. Impact of phosphoproteomics on studies of bacterial physiology. FEMS Microbiol. Rev. 2012, 36, 877–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dworkin, J. Ser/Thr phosphorylation as a regulatory mechanism in bacteria. Curr. Opin. Microbiol. 2015, 24, 47–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macek, B.; Mijakovic, I.; Olsen, J.V.; Gnad, F.; Kumar, C.; Jensen, P.R.; Mann, M. The serine/threonine/tyrosine phosphoproteome of the model bacterium Bacillus subtilis. Mol. Cell. Proteom. 2007, 6, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Macek, B.; Gnad, F.; Soufi, B.; Kumar, C.; Olsen, J.V.; Mijakovic, I.; Mann, M. Phosphoproteome analysis of E. coli reveals evolutionary conservation of bacterial Ser/Thr/Tyr phosphorylation. Mol. Cell. Proteom. 2008, 7, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Tian, C.F.; Chen, W.X. Site-specific Ser/Thr/Tyr phosphoproteome of Sinorhizobium meliloti at stationary phase. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Madec, E.; Laszkiewicz, A.; Iwanicki, A.; Obuchowski, M.; Séror, S. Characterization of a membrane-linked Ser/Thr protein kinase in Bacillus subtilis, implicated in developmental processes. Mol. Microbiol. 2002, 46, 571–586. [Google Scholar] [CrossRef] [PubMed]
- Ravikumar, V.; Shi, L.; Krug, K.; Derouiche, A.; Jers, C.; Cousin, C.; Kobir, A.; Mijakovic, I.; Macek, B. Quantitative phosphoproteome analysis of Bacillus subtilis reveals novel substrates of the kinase PrkC and phosphatase PrpC. Mol. Cell. Proteom. 2014, 13, 1965–1978. [Google Scholar] [CrossRef] [PubMed]
- Beltramini, A.M.; Mukhopadhyay, C.D.; Pancholi, V. Modulation of cell wall structure and antimicrobial susceptibility by a Staphylococcus aureus eukaryote-like serine/threonine kinase and phosphatase. Infect. Immun. 2009, 77, 1406–1416. [Google Scholar] [CrossRef] [PubMed]
- Rajagopal, L.; Clancy, A.; Rubens, C.E. A eukaryotic type serine/threonine kinase and phosphatase in Streptococcus agalactiae reversibly phosphorylate an inorganic pyrophosphatase and affect growth, cell segregation, and virulence. J. Biol. Chem. 2003, 278, 14429–14441. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Agarwal, S.; Pancholi, P.; Pancholi, V. Role of serine/threonine phosphatase (SP-STP) in Streptococcus pyogenes physiology and virulence. J. Biol. Chem. 2011, 286, 41368–41380. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Kapatral, V.; Xu, W.; Chakrabarty, A.M. Characterization of a Hank’s type serine/threonine kinase and serine/threonine phosphoprotein phosphatase in Pseudomonas aeruginosa. J. Bacteriol. 1999, 181, 6615–6622. [Google Scholar] [PubMed]
- Barton, G.J.; Cohen, P.T.; Barford, D. Conservation analysis and structure prediction of the protein serine/threonine phosphatases. Sequence similarity with diadenosine tetraphosphatase from Escherichia coli suggests homology to the protein phosphatases. Eur. J. Biochem. 1994, 220, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Villafranca, J.E.; Kissinger, C.R.; Parge, H.E. Protein serine/threonine phosphatases. Curr. Opin. Biotechnol. 1996, 7, 397–402. [Google Scholar] [CrossRef]
- Bollen, M.; Stalmans, W. The structure, role, and regulation of type 1 protein phosphatases. Crit. Rev. Biochem. Mol. Biol. 1992, 27, 227–281. [Google Scholar] [CrossRef] [PubMed]
- Burnside, K.; Rajagopal, L. Regulation of prokaryotic gene expression by eukaryotic-like enzymes. Curr. Opin. Microbiol. 2012, 15, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umeyama, T.; Naruoka, A.; Horinouchi, S. Genetic and biochemical characterization of a protein phosphatase with dual substrate specificity in Streptomyces coelicolor A3(2). Gene 2000, 258, 55–62. [Google Scholar] [CrossRef]
- Treuner-Lange, A.; Ward, M.J.; Zusman, D.R. Pph1 from Myxococcus xanthus is a protein phosphatase involved in vegetative growth and development. Mol. Microbiol. 2001, 40, 26–40. [Google Scholar] [CrossRef]
- Burnside, K.; Rajagopal, L. Aspects of eukaryotic-like signaling in Gram-positive cocci: A focus on virulence. Future Microbiol. 2011, 6, 747–761. [Google Scholar] [CrossRef] [PubMed]
- Pereira, S.F.; Goss, L.; Dworkin, J. Eukaryote-like serine/threonine kinases and phosphatases in bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 192–212. [Google Scholar] [CrossRef] [PubMed]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains, Plasmids, and Primers | Characteristics | Source or Reference |
|---|---|---|
| R. leguminosarum bv. trifolii | ||
| Rt24.2 | Wild type, Rifr, Nxr | [38] |
| Rt297 | Rt24.2 pssZ::mTn5SSgusA40, Spr | This work |
| Rt297(pPL1) | Rt297 carrying pssZ on pBBR1MCS-2 vector, Kmr | This work |
| Rt24.2(pPL1) | Rt24.2 carrying pssZ on pBBR1MCS-2 vector, Kmr | This work |
| Rt24.2 (pBBR1MCS-2) | Rt24.2 carrying pBBR1MCS-2 vector, Kmr | [39] |
| E. coli | ||
| DH5α | supE44 ΔlacU169 (φ80 lacZΔ M15) hsdR17 recA1endA1gyrA96 thi-1 relA1 | [40] |
| S17-1 | thi pro hsdR− hsdM+ recA RP4-2-Tc::Mu-Km::Tn7 | [41] |
| mTn5SSgusA40 | miniTn5 interposon containing a promoterless gusA gene, Spr | [42] |
| Plasmids | ||
| pBBR1MCS-2 | mob, lacZα, cloning vector, Kmr | [43] |
| pJBA21Tc | pMP220 containing gusA, Tcr | [44] |
| pPL1 | pBBR1MCS-2 containing a 1.8-kb SalI-XbaI fragment with the Rt24.2 pssZ gene, Kmr | This work |
| Primers | Sequence (5′–3′) 1 | |
| gusF1 | GCGTTACAAGAAAGCCGGGCAATT | This work |
| gusR1 | GATCCAGACTGAATGCCCACAGGC | This work |
| gusR2 | CAGCAATTGCCCGGCTTTCTTGTAA | This work |
| gusR3 | GTCTGCCAGTTCAGTTCGTTGTTC | This work |
| Xba-Fw1 | GGGTTTATCTAGACTGGCATCGGCAC | This work |
| Xba-Fw3 | CAATCTCTATCTAGATGTGACCAACACC | This work |
| Xba-Fw4 | GGACGCTCTAGATCTTTCAATCCTC | This work |
| Eco-Rw1 | CCCGGTGAATTCGCCATCGTCAAC | This work |
| J44-Rw4 | CAACCGCAGTTTCCACTTTGCACC | This work |
| J44-Rw5 | GGATCTGAGATTCCTGATCAAGAAATG | This work |
| Sal-Rw2 | CCTTCATATTGTCGACTCTGACCGTT | This work |
| Strain | Minimal Inhibitory Concentration a,b | ||
|---|---|---|---|
| SDS (% w/v) | DOC (% w/v) | Ethanol (% v/v) | |
| Rt24.2 (wt) | 0.035 ± 0.005 B | 0.12 ± 0.05 B | 5.0 ± 0.25 A,B |
| Rt297(pssZ) | 0.025 ± 0.005 B,C | 0.11 ± 0.05 B | 4.25 ± 0.25 C |
| Rt297(pPL1) | 0.045 ± 0.005 A,B | 0.12 ± 0.05 B | 5.0 ± 0.25 A,B |
| Rt24.2(pPL1) | 0.050 ± 0.005 A | 0.14 ± 0.05 A | 5.5 ± 0.25 A |
| Property | Strain | |||
|---|---|---|---|---|
| Rt24.2 | Rt297 | Rt297(pPL1) | Rt24.2(pPL1) | |
| Length (μm) | 2.50 ± 0.21 | 2.83 ± 0.13 | 2.64 ± 0.22 | 2.58 ± 0.24 |
| Width (μm) | 0.78 ± 0.06 | 0.86 ± 0.05 | 0.80 ± 0.07 | 0.82 ± 0.05 |
| Height (μm) | 0.215 ± 0.027 | 0.184 ± 0.022 | 0.178 ± 0.015 | 0.204 ± 0.021 |
| Roughness (nm) | 2.786 ± 0.360 | 0.918 ± 0.240 * | 2.903 ± 0.38 | 2.353 ± 0.26 |
| DMT modulus (elasticity; GPa) | 1.326 ± 0.187 | 2.501 ± 0.561 * | 1.484 ± 0.206 | 1.524 ± 0.195 |
| Adhesion (nN) | 501.93 ± 49.74 | 257.00 ± 33.11 * | 541.71 ± 36.00 | 609.43 ± 43.61 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lipa, P.; Vinardell, J.-M.; Kopcińska, J.; Zdybicka-Barabas, A.; Janczarek, M. Mutation in the pssZ Gene Negatively Impacts Exopolysaccharide Synthesis, Surface Properties, and Symbiosis of Rhizobium leguminosarum bv. trifolii with Clover. Genes 2018, 9, 369. https://doi.org/10.3390/genes9070369
Lipa P, Vinardell J-M, Kopcińska J, Zdybicka-Barabas A, Janczarek M. Mutation in the pssZ Gene Negatively Impacts Exopolysaccharide Synthesis, Surface Properties, and Symbiosis of Rhizobium leguminosarum bv. trifolii with Clover. Genes. 2018; 9(7):369. https://doi.org/10.3390/genes9070369
Chicago/Turabian StyleLipa, Paulina, José-María Vinardell, Joanna Kopcińska, Agnieszka Zdybicka-Barabas, and Monika Janczarek. 2018. "Mutation in the pssZ Gene Negatively Impacts Exopolysaccharide Synthesis, Surface Properties, and Symbiosis of Rhizobium leguminosarum bv. trifolii with Clover" Genes 9, no. 7: 369. https://doi.org/10.3390/genes9070369
APA StyleLipa, P., Vinardell, J.-M., Kopcińska, J., Zdybicka-Barabas, A., & Janczarek, M. (2018). Mutation in the pssZ Gene Negatively Impacts Exopolysaccharide Synthesis, Surface Properties, and Symbiosis of Rhizobium leguminosarum bv. trifolii with Clover. Genes, 9(7), 369. https://doi.org/10.3390/genes9070369