Stable Isotopes Reveal Climate Signal Hidden in Tree Rings of Endemic Balkan Pines


Abstract
1. Introduction
2. Material and Methods
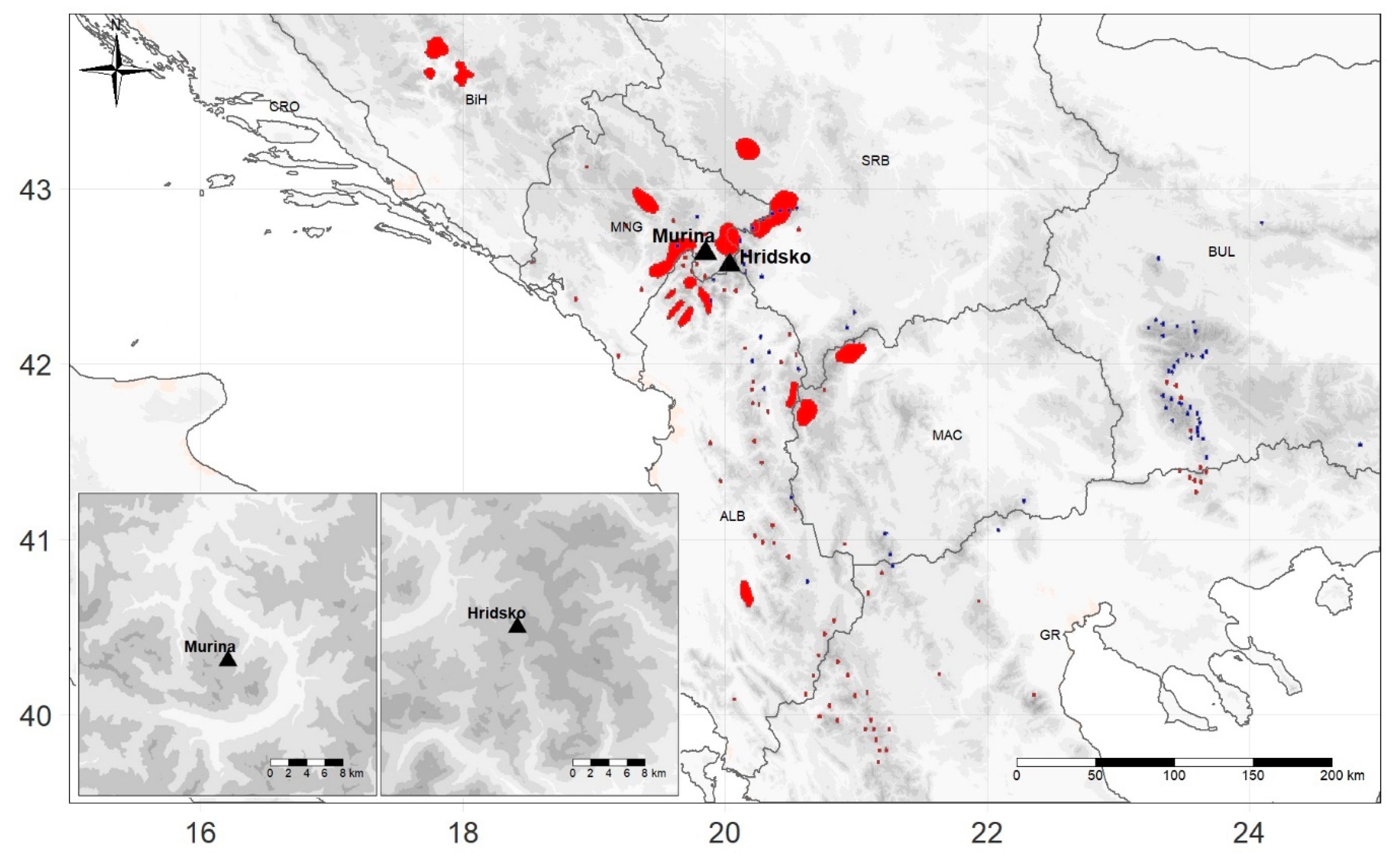
2.1. Sampling Locations
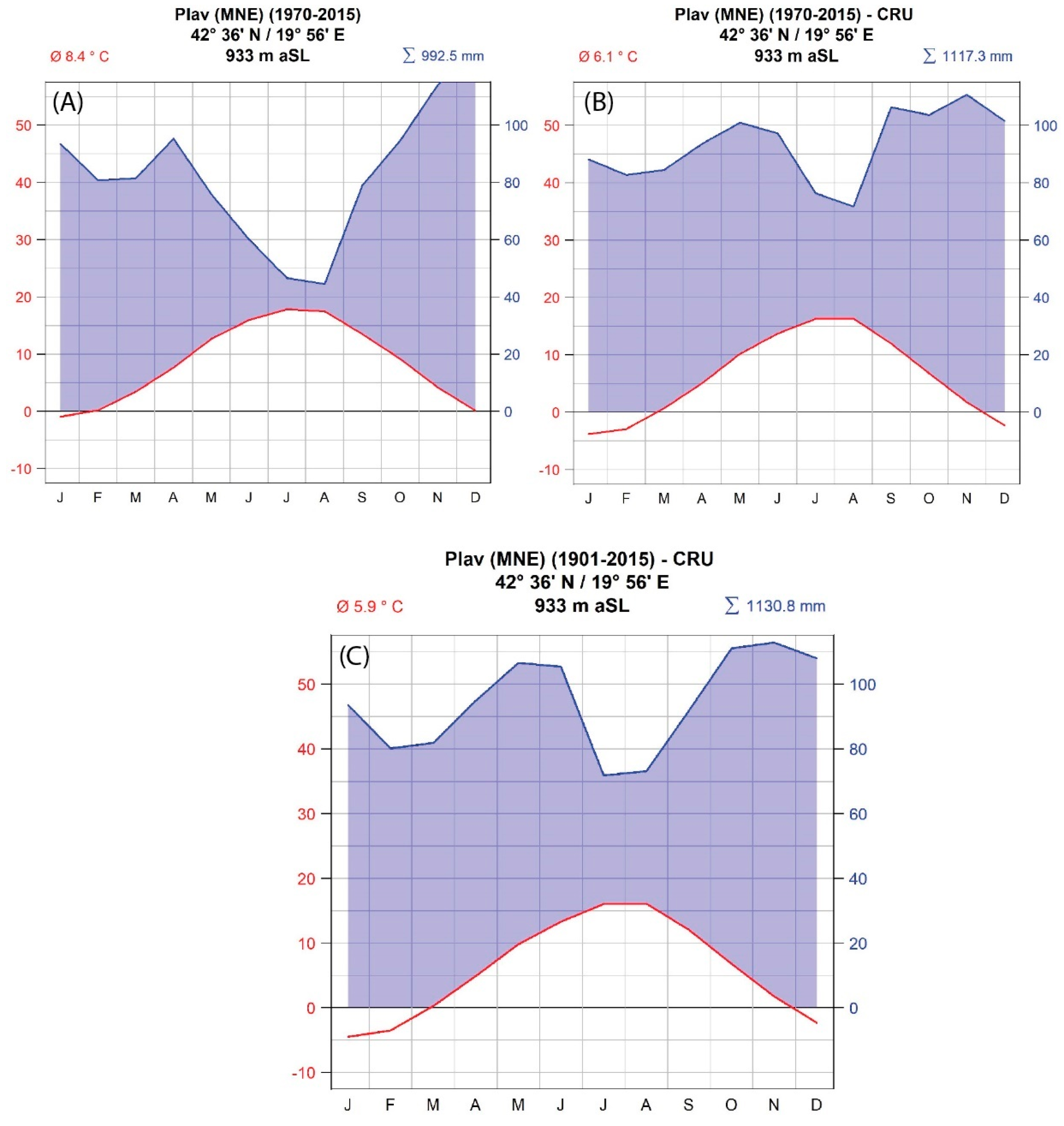
2.2. Meteorological Data
2.3. Sample Collection and Tree-Ring Width Analysis
2.4. Stable Carbon Isotope Analysis
2.5. Analysis of the Climate-Growth Relationship
3. Results
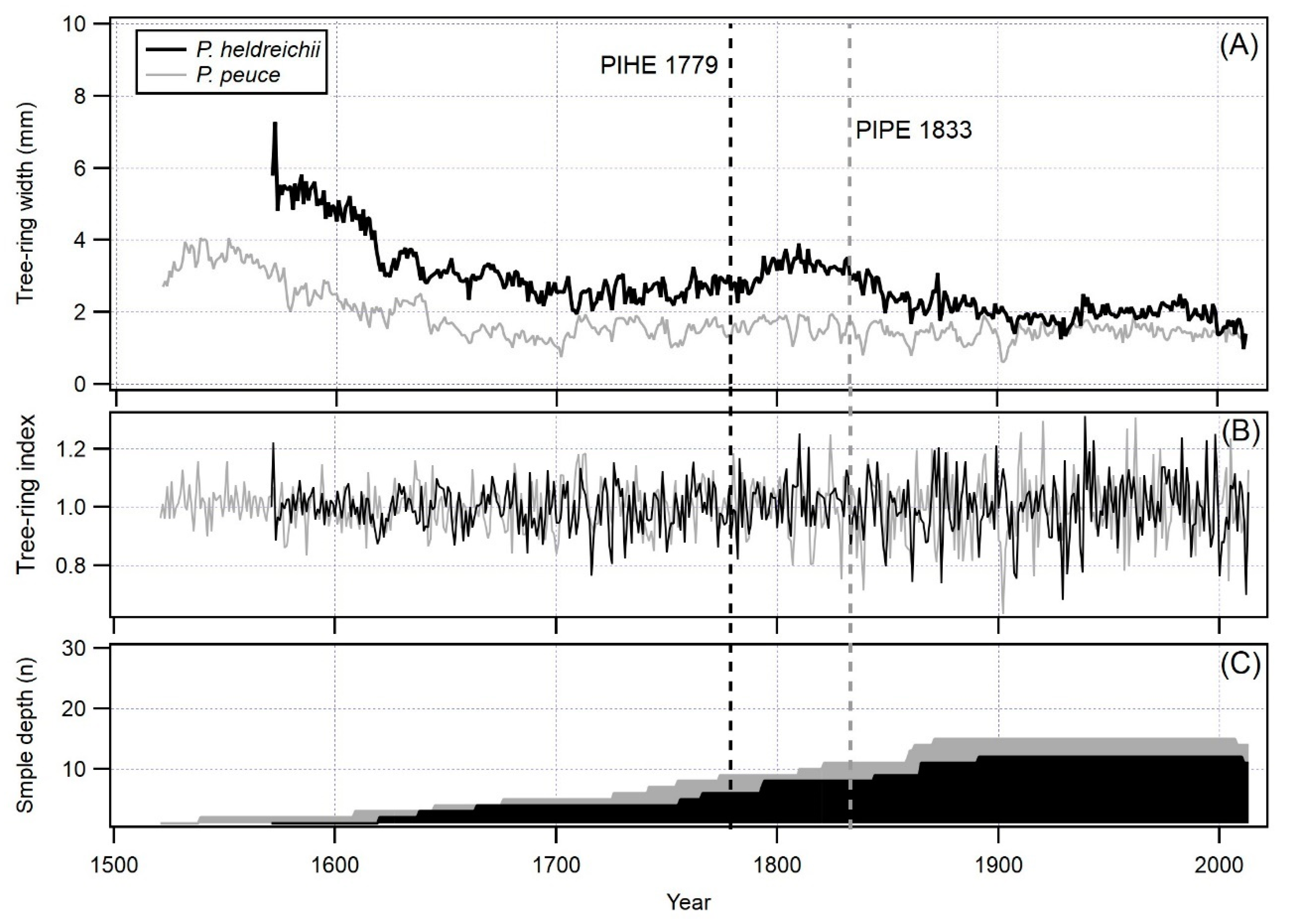
3.1. Long TRW Chronology
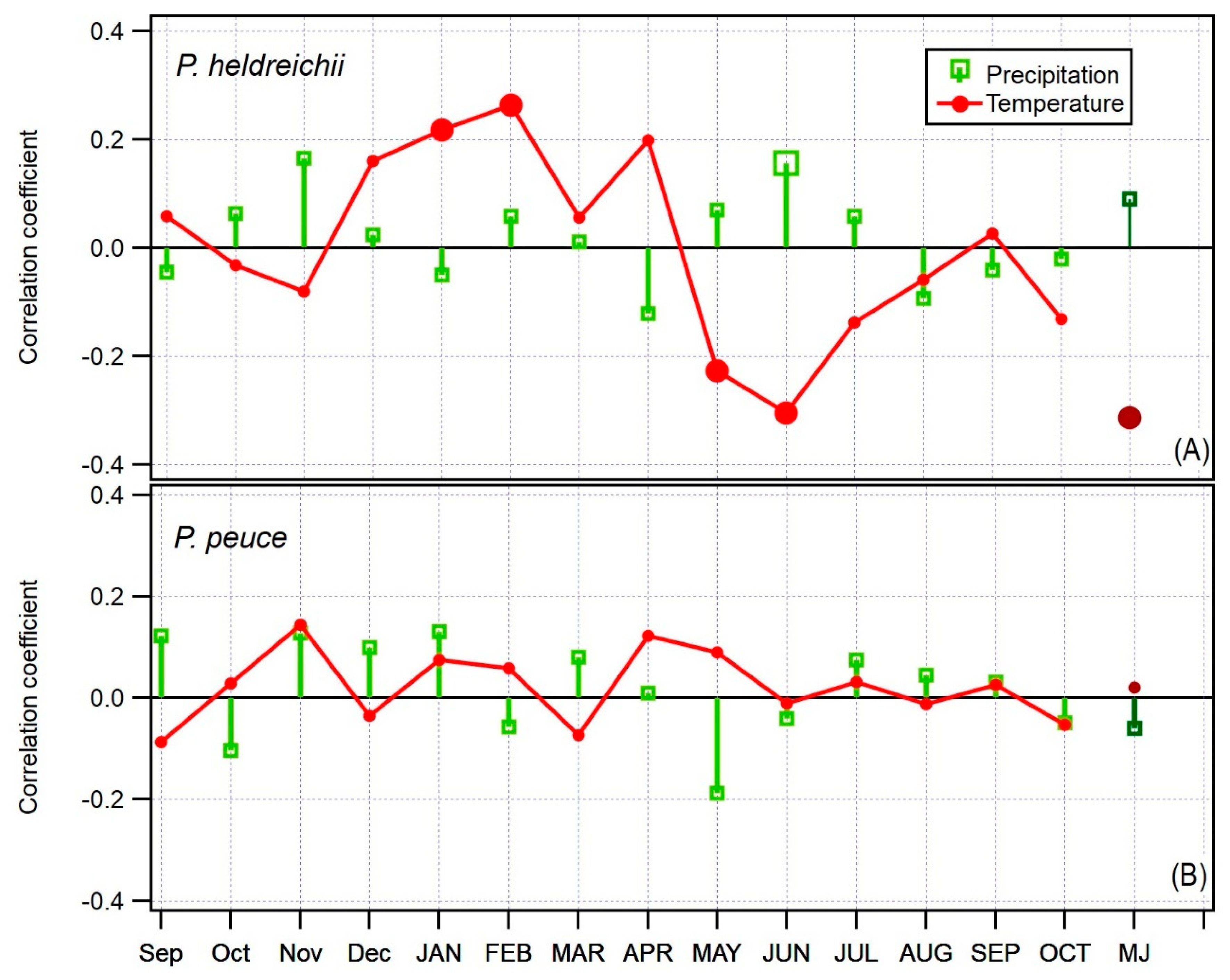
3.2. Climate Signal in P. peuce and P. heldreichii Tree-Ring Widths
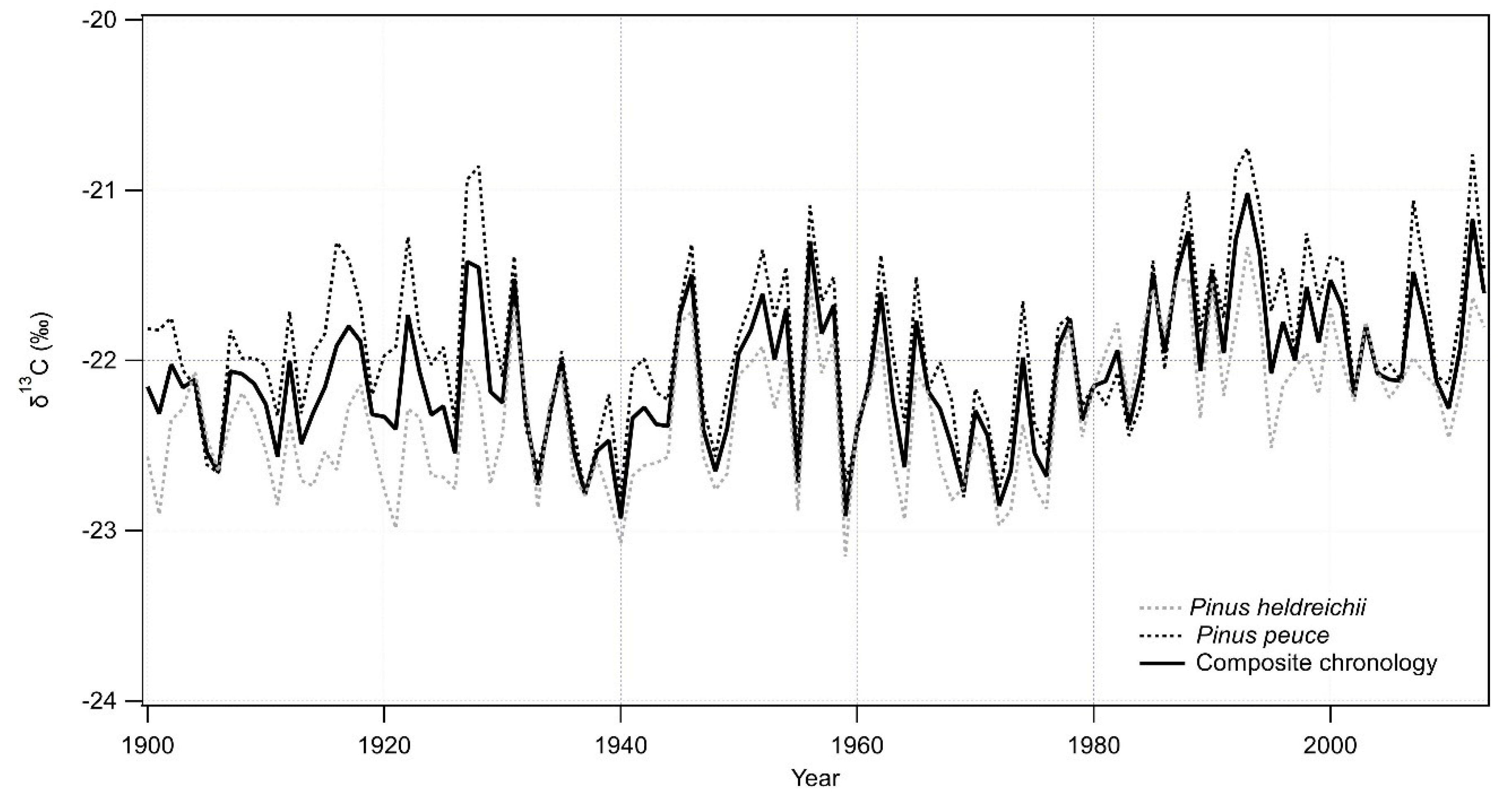
3.3. P. heldreichii and P. peuce Stable Isotope Ratio Chronologies
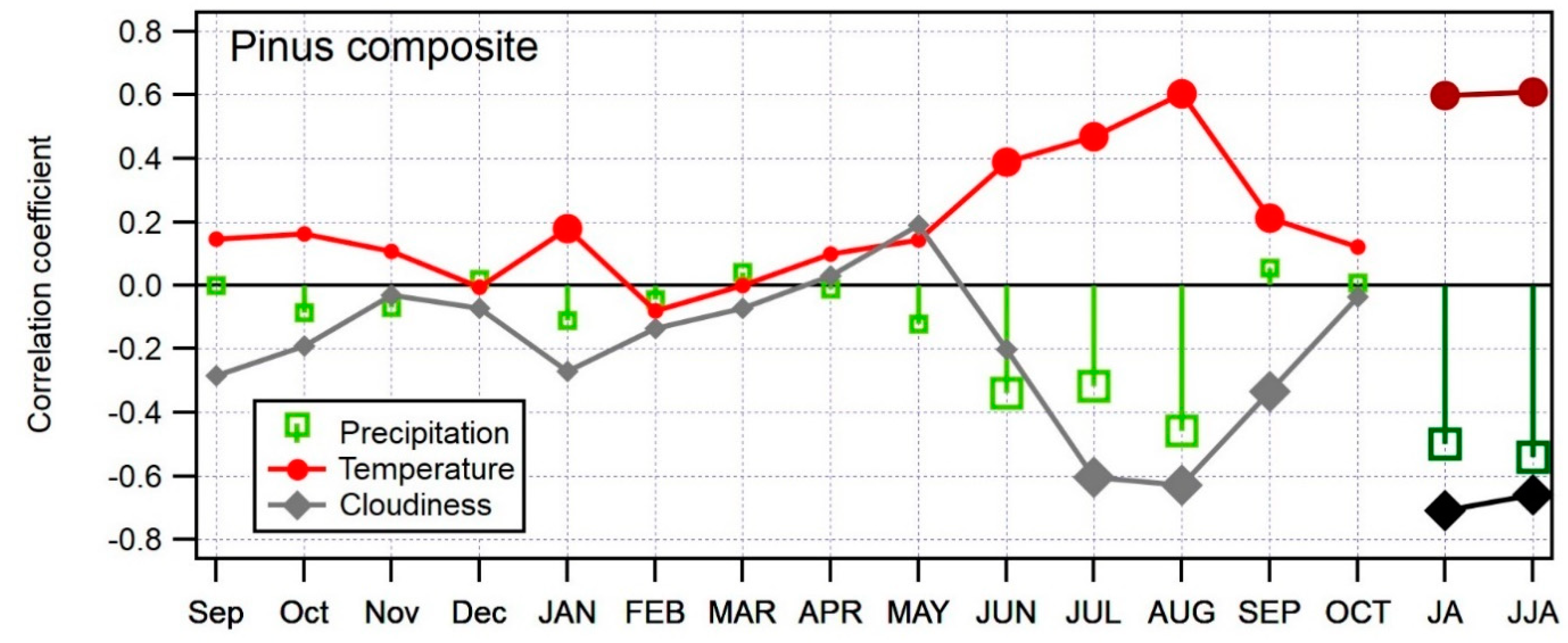
3.4. Relationship between Climate and Composite δ13C Chronology
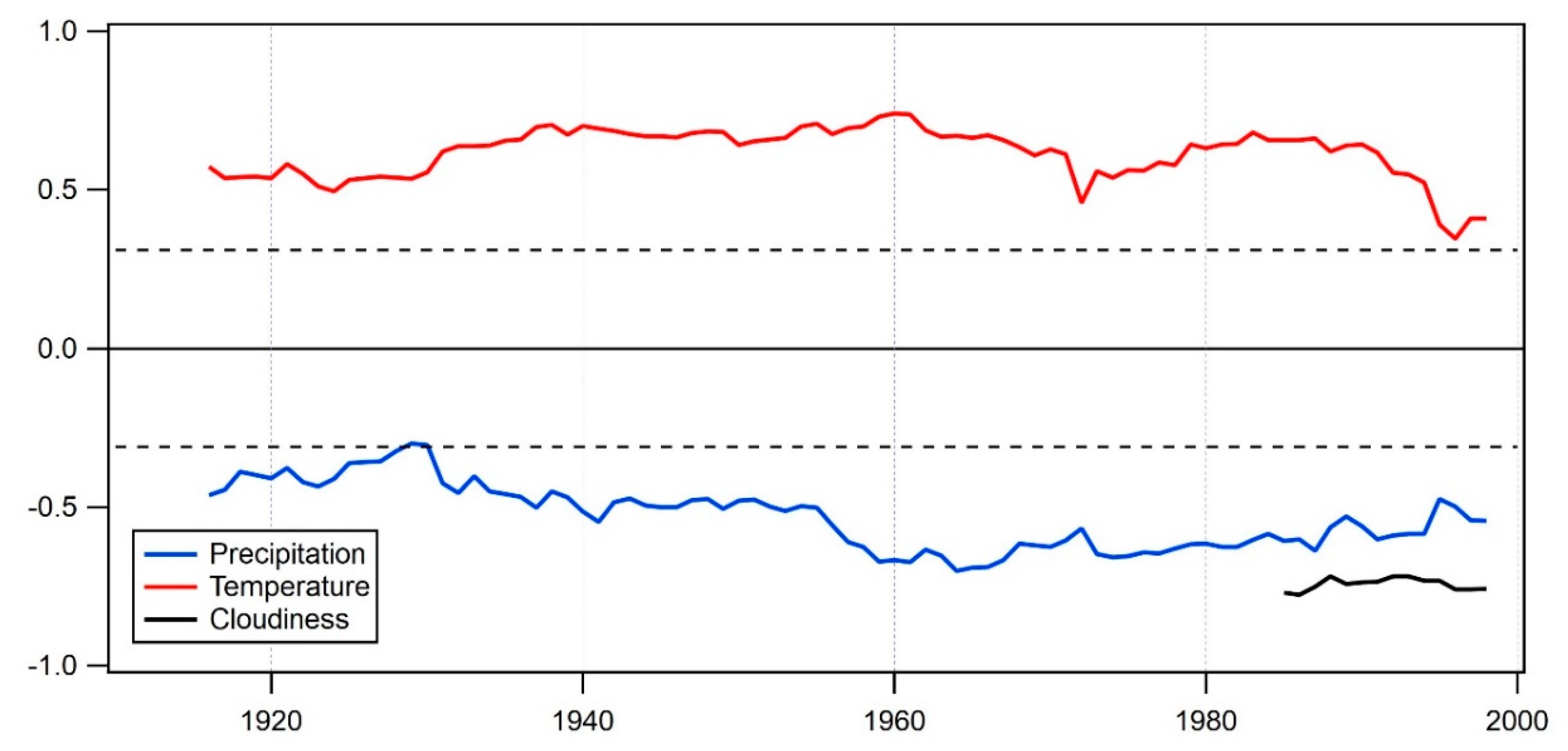
3.5. Temporal Stability of the Climate Signal
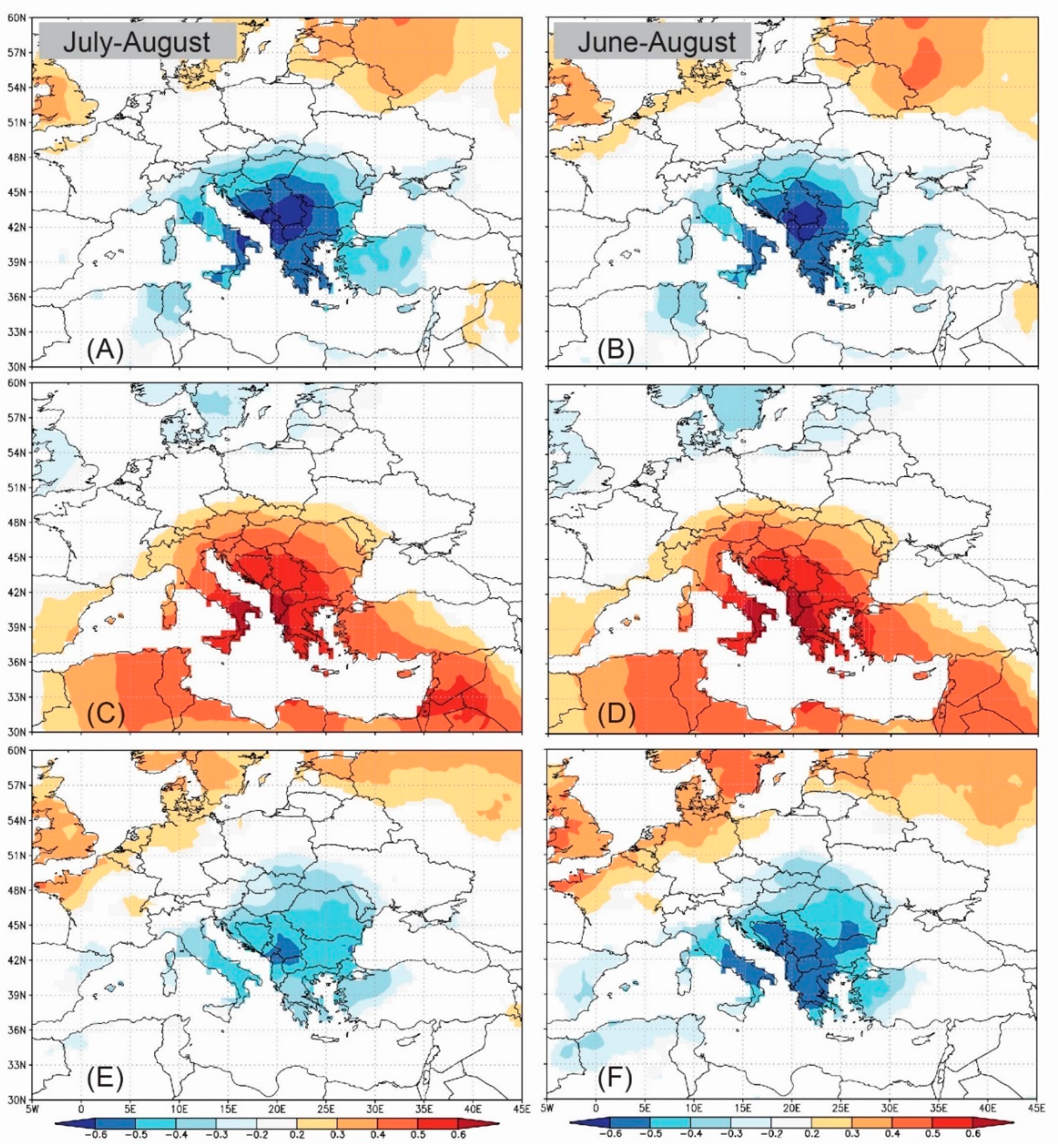
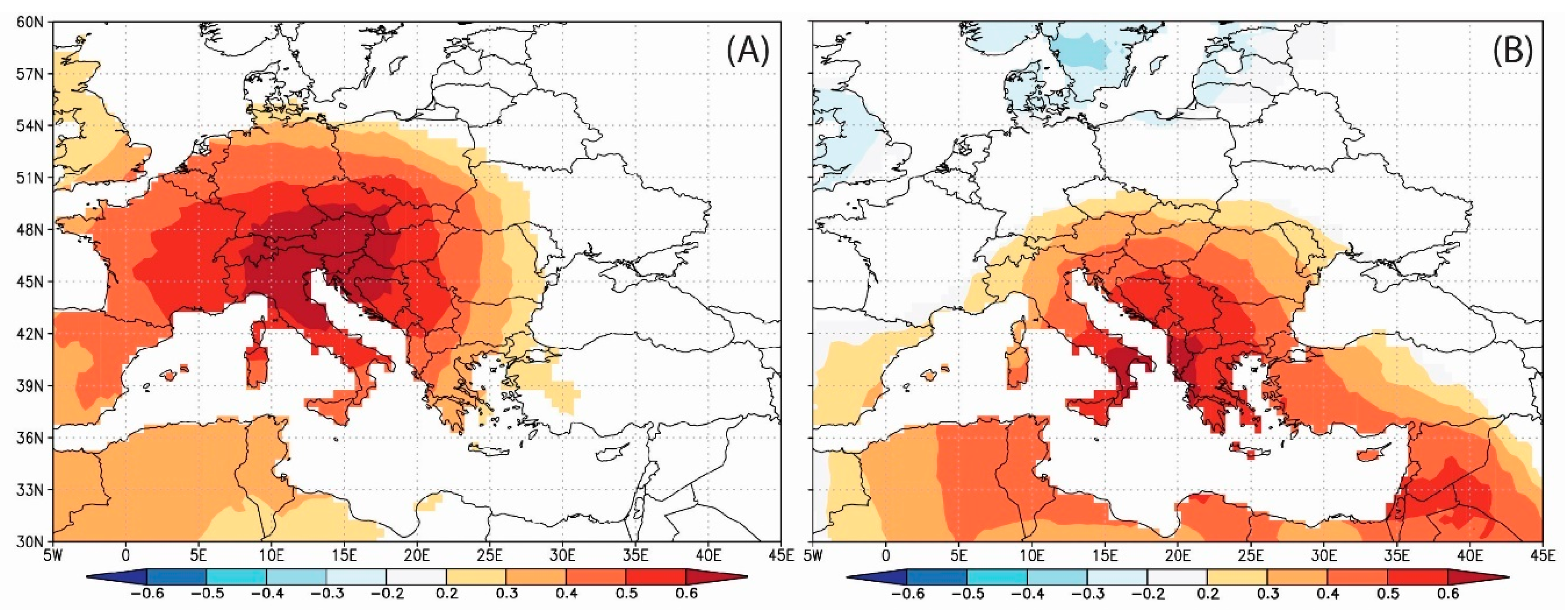
3.6. Spatial Pattern of Correlation between δ13C and Climate and Spatial Outreach of the Climate Signal
3.7. Potential for Climate Reconstruction
4. Discussion
4.1. Long TRW Chronologies and Their Potential for Dating Wooden Objects and Constructions
4.2. Climate Signal in Tree-Ring Widths
4.3. Climate Signal in Stable Carbon Isotopes in Tree Rings
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change. Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Solomon, S., Qin, D., Manning, M., Chen, Z., Marquis, M., Averyt, K.B., Tignor, M., Miller, H.L., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Matyas, C. Climate Change Imapcts on Forest Management in Eastern Europe and Central Asia—Dimensions, Impacts and Adaptation Policies; Food and Agricultural Organization of the United Nations: Rome, Italy, 2010. [Google Scholar]
- Bennett, K.D.; Tzedakis, P.C.; Willis, K.J. Quaternary Refugia of North European Trees. J. Biogeogr. 1991, 18, 103–115. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Willis, K. The vegetational history of the Balkans. Quat. Sci. Rev. 1994, 13, 769–788. [Google Scholar] [CrossRef]
- Opała-Owczarek, M. Warm-season temperature reconstruction from high-elevation juniper tree rings over the past millennium in the Pamir region. Palaeogeogr. Palaeoclim. Palaeoecol. 2019, 532, 109248. [Google Scholar] [CrossRef]
- Corona, C.; Guiot, J.; Edouard, J.L.; Chalié, F.; Büntgen, U.; Nola, P.; Urbinati, C. Millennium-long summer temperature variations in the European Alps as reconstructed from tree rings. Clim. Past 2010, 6, 379–400. [Google Scholar] [CrossRef]
- Cerrato, R.; Salvatore, M.C.; Gunnarson, B.E.; Linderholm, H.W.; Carturan, L.; Brunetti, M.; De Blasi, F.; Baroni, C. A Pinus cembra L. tree-ring record for late spring to late summer temperature in the Rhaetian Alps, Italy. Dendrochronologia 2019, 53, 22–31. [Google Scholar] [CrossRef]
- Saulnier, M.; Corona, C.; Stoffel, M.; Guibal, F.; Edouard, J.-L. Climate-growth relationships in a Larix decidua Mill. network in the French Alps. Sci. Total. Environ. 2019, 664, 554–566. [Google Scholar] [CrossRef]
- Panayotov, M.; Bebi, P.; Trouet, V.; Yurukov, S. Climate signal in tree-ring chronologies of Pinus peuce and Pinus heldreichii from the Pirin Mountains in Bulgaria. Trees 2010, 24, 479–490. [Google Scholar] [CrossRef]
- Trouet, V.; Panayotov, M.; Ivanova, A.; Frank, D. A pan-European summer teleconnection mode recorded by a new temperature reconstruction from the northeastern Mediterranean (ad 1768–2008). Holocene 2012, 22, 887–898. [Google Scholar] [CrossRef]
- Panayotov, M.; Tsavkov, E.; Zhelev, P.; Yurukov, S.; Ivanova, A.; Russeva, M.; Todorova, Y.; Trouet, V. Tree ring and anatomical studies in Pinus heldreichii forests in Pirin mountains, Bulgaria. For. Ideas 2011, 17, 66–71. [Google Scholar]
- Scheithauer, J.; Grunewald, K.; Helle, G.; Guenther, B.; Koenig, J.; Gikov, A. Dednroecological studies on Bosnian Pine (Pirin Mtns., Bulgaria). In Proceedings of the TRACE 2008, Zakopane, Poland, 27–30 April 2008; pp. 142–150. [Google Scholar]
- Scheithauer, J.; Grunewald, K.; Helle, G.; Günther, B.; Gikov, A. Bosnian Pine (Pinus Heldreichii) as Geoarchive at the Timberline in the Pirin Mountains and on the Balkan Peninsula. Biotechnol. Biotechnol. Equip. 2014, 23, 96–99. [Google Scholar] [CrossRef]
- Bojaxhi, F.; Toromani, E. Spatial and temporal variation in Bosnian pine (Pinus heldreichii Christ.) growth due to climate in Kosovo. Int. J. Dev. Sustain. 2017, 6, 1–15. [Google Scholar]
- Bojaxhi, F.; Toromani, E. The Growth of Bosnian Pine (Pinus hedreichii Christ.) at Tree-Line Locations from Kosovo and its Response to Climate. South-East Eur. For. 2016, 7, 109–118. [Google Scholar] [CrossRef][Green Version]
- Seim, A.; Büntgen, U.; Fonti, P.; Haska, H.; Herzig, F.; Tegel, W.; Trouet, V.; Treydte, K. Climate sensitivity of a millennium-long pine chronology from Albania. Clim. Res. 2012, 51, 217–228. [Google Scholar] [CrossRef]
- Seim, A.; Treydte, K.; Büntgen, U.; Esper, J.; Fonti, P.; Haska, H.; Herzig, F.; Tegel, W.; Faust, D. Exploring the potential of Pinus heldreichii Christ. for long-term climate reconstruction in Albania. In Proceedings of TRACE—Tree rings in Archaeology, Climatology and Ecology, Otočec, Slovenija, 16–19 April 2009; pp. 75–82. [Google Scholar]
- Klippel, L.; Krusic, P.J.; Brandes, R.; Hartl-Meier, C.; Trouet, V.; Meko, M.; Esper, J. High-elevation inter-site differences in Mount Smolikas tree-ring width data. Dendrochronologia 2017, 44, 164–173. [Google Scholar] [CrossRef]
- Konter, O.; Krusic, P.; Trouet, V.; Esper, J. Meet Adonis, Europe’s oldest dendrochronologically dated tree. Dendrochronologia 2017, 42. [Google Scholar] [CrossRef]
- Nagavciuc, V.; Kern, Z.; Ionita, M.; Hartl, C.; Konter, O.; Esper, J.; Popa, I. Climate signals in carbon and oxygen isotope ratios of Pinus cembra tree-ring cellulose from the Călimani Mountains, Romania. Int. J. Clim. 2019. [Google Scholar] [CrossRef]
- Szymczak, S.; Joachimski, M.M.; Brauning, A.; Hetzer, T.; Kuhlemann, J. A 560 yr summer temperature reconstruction for the Western Mediterranean basin based on stable carbon isotopes from Pinus nigra ssp. laricio (Corsica/France). Clim. Past Discuss. 2012, 8, 1737–1749. [Google Scholar] [CrossRef]
- Szymczak, S.; Joachimski, M.; Bräuning, A.; Hetzer, T.; Kuhlemann, J. Comparison of whole wood and cellulose carbon and oxygen isotope series from Pinus nigra ssp. laricio (Corsica/France). Dendrochronologia 2011, 29, 219–226. [Google Scholar] [CrossRef]
- Hafner, P.; McCarroll, D.; Robertson, I.; Loader, N.; Gagen, M.; Young, G.; Bale, R.; Sonninen, E.; Levanič, T. A 520 year record of summer sunshine for the eastern European Alps based on stable carbon isotopes in larch tree rings. Clim. Dyn. 2014, 43, 971–980. [Google Scholar] [CrossRef]
- Kern, Z.; Patkó, M.; Kázmér, M.; Fekete, J.; Kele, S.; Pályi, Z. Multiple tree-ring proxies (earlywood width, latewood width and δ13C) from pedunculate oak (Quercus robur L.), Hungary. Quat. Int. 2013, 293, 257–267. [Google Scholar] [CrossRef]
- Vendramin, G.G.; Fineschi, S.; Fady, B. EUFORGEN Technical Guidelines for genetic conservation and use for Bosnian pine (Pinus heldreichii); EUFORGENE c/o Biodiversity International: Rome, Italy, 2008; p. 6. [Google Scholar]
- Aleksandrov, A.H.; Andonovski, V. EUFORGEN Technical Guidelines for genetic conservation and use of Macedonian pine (Pinus peuce); EUFORGEN c/o Biodiversity International: Rome, Italy, 2011; p. 6. [Google Scholar]
- Brus, R. Dendrologija za gozdarje: University book, 2nd ed.; Biotechnical Faculty, Department for Forestry and Renewable Forest Resources: Ljubljana, Slovenia, 2008; p. 408. [Google Scholar]
- Mitchell, T.D.; Jones, P.D. An improved method of constructing a database of monthly climate observations and associated high-resolution grids. Int. J. Clim. 2005, 25, 693–712. [Google Scholar] [CrossRef]
- Levanič, T. Atrics—A New System for Image Acquisition in Dendrochronology. Tree-Ring Res. 2007, 63, 117–122. [Google Scholar] [CrossRef]
- Cook, E.R. A Time Series Analysis Approach to Tree-Ring Standardization; University of Arizona: Tucson, AZ, USA, 1985. [Google Scholar]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Bunn, A.G. Statistical and visual crossdating in R using the dplR library. Dendrochronologia 2010, 28, 251–258. [Google Scholar] [CrossRef]
- Loader, N.J.; Robertson, I.; Barker, A.C.; Switsur, V.R.; Waterhouse, J.S. An improved technique for the batch processing of small wholewood samples to a-cellulose. Chem. Geol. 1997, 136, 313–317. [Google Scholar] [CrossRef]
- McCarroll, D.; Loader, N.J. Stable isotopes in tree rings. Quat. Sci. Rev. 2004, 23, 771–801. [Google Scholar] [CrossRef]
- McCarroll, D.; Gagen, M.H.; Loader, N.J.; Robertson, I.; Anchukaitis, K.J.; Los, S.; Young, G.H.; Jalkanen, R.; Kirchhefer, A.J.; Waterhouse, J.S. Correction of tree ring stable carbon isotope chronologies for changes in the carbon dioxide content of the atmosphere. Geochim. Et Cosmochim. Acta 2009, 73, 1539–1547. [Google Scholar] [CrossRef]
- Gagen, M.; McCarroll, D.; Loader, N.J.; Robertson, I.; Jalkanen, R.; Anchukaitis, K.J. Exorcising the ‘segment length curse’: summer temperature reconstruction since AD 1640 using non-detrended stable carbon isotope ratios from pine trees in northern Finland. Holocene 2007, 17, 435–446. [Google Scholar] [CrossRef]
- Gagen, M.; McCarroll, D.; Robertson, I.; Loader, N.J.; Jalkanen, R. Do tree ring δ13C series from Pinus sylvestris in northern Fennoscandia contain long-term non-climatic trends? Chem. Geol. 2008, 252, 42–51. [Google Scholar] [CrossRef]
- Loader, N.; Santillo, P.; Woodman-Ralph, J.; Rolfe, J.; Hall, M.; Gagen, M.; Robertson, I.; Wilson, R.; Froyd, C.; McCarroll, D. Multiple stable isotopes from oak trees in southwestern Scotland and the potential for stable isotope dendroclimatology in maritime climatic regions. Chem. Geol. 2008, 252, 62–71. [Google Scholar] [CrossRef]
- Young, G.H.; McCarroll, D.; Loader, N.J.; Kirchhefer, A.J. A 500-year record of summer near-ground solar radiation from tree-ring stable carbon isotopes. Holocene 2010, 20, 315–324. [Google Scholar] [CrossRef]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the Average Value of Correlated Time Series, with Applications in Dendroclimatology and Hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Zang, C.; Biondi, F. Treeclim: An R package for the numerical calibration of proxy-climate relationships. Ecography 2015, 38, 431–436. [Google Scholar] [CrossRef]
- Van Oldenborgh, G.J.; Balmaseda, M.A.; Ferranti, L.; Stockdale, T.N.; Anderson, D.L.T. Evaluation of Atmospheric Fields from the ECMWF Seasonal Forecasts over a 15-Year Period. J. Clim. 2005, 18, 3250–3269. [Google Scholar] [CrossRef][Green Version]
- Trouet, V.; Van Oldenborgh, G.J. KNMI Climate Explorer: A Web-Based Research Tool for High-Resolution Paleoclimatology. Tree-Ring Res. 2013, 69, 3–13. [Google Scholar] [CrossRef]
- North, G.B.; Biondi, F.; Bloomfield, P.; Christy, J.R.; Cuffey, K.M.; Dickinson, R.E.; Druffel, E.R.M.; Nychka, D. Surface Temperature Reconstructions for the last 2000 Years; National Academies Press: Washington, DC, USA, 2006; p. 160. [Google Scholar]
- Panayotov, M.; Yurukov, S. Tree ring chronology of Pinus peuce from the Pirin Mts and possibilities to use it for climate analysis. Phytol. Balc. 2007, 13, 313–320. [Google Scholar]
- Büntgen, U.; Frank, D.; Trouet, V.; Esper, J. Diverse climate sensitivity of Mediterranean tree-ring width and density. Trees-Struct. Funct. 2010, 24, 261–273. [Google Scholar] [CrossRef]
- Jevšenak, J.; Levanič, T.; Jernej, J.; Tom, L. dendroTools: R package for studying linear and nonlinear responses between tree-rings and daily environmental data. Dendrochronologia 2018, 48, 32–39. [Google Scholar] [CrossRef]
- Touchan, R.; Shishov, V.V.; Meko, D.M.; Nouiri, I.; Grachev, A. Process based model sheds light on climate sensitivity of Mediterranean tree-ring width. Biogeosciences 2012, 9, 965–972. [Google Scholar] [CrossRef]
- Jevšenak, J. Daily climate data reveal stronger climate-growth relationships for an extended European tree-ring network. Quat. Sci. Rev. 2019, 221, 221. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pinus Heldreichii | Pinus Peuce | |
|---|---|---|
| Local site name | Murina | Hridsko |
| Coordinates | N 42.57°, E 20.03° | N 42.63°, E 19.85° |
| Elevation | 1710 m | 1960 m |
| Slope | 40–50° | 10–40° |
| Exposition | W-NW | W-NW |
| Soil type | Brown soil on limestone | Acidic soil on ultrabasic |
| Number of cores for tree-ring analysis | 21 cores (11 trees) | 27 cores (16 trees) |
| Number of cores for δ13C | 6 cores (6 trees) | 9 cores (9 trees) |
| Parameter | Pinus Heldreichii | Pinus Peuce |
|---|---|---|
| EPS ≥ 0.85 | 1779 | 1833 |
| RBAR | 0.397 | 0.353 |
| SNR | 1.464 | 2.416 |
| Time period | 1571–2013 | 1521–2013 |
| Number of rings | 443 | 493 |
| Average TRW | 1.71 | 1.24 |
| Standard deviation | 0.82 | 0.56 |
| 1st order autocorrelation | 0.84 | 0.86 |
| Mean sensitivity | 0.17 | 0.17 |
| Pinus peuce(Hridsko, MNE) | ||||
| Location | tBP | GLK% | Overlap | Time Period |
| Begova česma (MKD) | 0.60 | 54.20 | 189 | 1825–2014 |
| Lure (ALB) | 3.05 | 54.70 | 451 | 1100–1971 |
| Pinus heldreichii(Murina, MNE) | ||||
| Location | tBP | GLK% | Overlap | Time Period |
| Boričevac (BIH) | 5.89 | 71.20 | 172 | 1839–2010 |
| Prenj (BIH) | 9.99 | 66.60 | 438 | 1573–2010 |
| Šar planina (XKX) | 10.60 | 72.30 | 157 | 1857–2014 |
| Koritnik (XKX) | 6.28 | 70.60 | 182 | 1831–2012 |
| Streoške planine (XKX) | 15.50 | 71.20 | 274 | 1741–2014 |
| Prevalla (XKX) | 8.20 | 70.70 | 135 | 1879–2014 |
| Seasonal variable | Entire Period | 1902–1957 | 1958–2013 | |||||
|---|---|---|---|---|---|---|---|---|
| r2 | RE | CE | MSE | RE | CE | MSE | ||
| TRW P. heldreichii | MJ | 0.097 | −0.065 | −0.315 | 1.6 | 0.009 | −0.18 | 1.767 |
| P. Peuce | P. Heldreichii | Pinus Composite | |
|---|---|---|---|
| n | 5 | 6 | 11 |
| rBT | 0.31 | 0.37 | 0.33 |
| EPS | 0.69 | 0.78 | 0.84 |
| average | −22,3 | −21.9 | −22.1 |
| standard deviation | 0.40 | 0.47 | 0.41 |
| minimum | −23.2 | −22.8 | −22.9 |
| maximum | −21.3 | −20.8 | −21.0 |
| Seasonal | 1902–1957 | 1958–2013 | |||||
|---|---|---|---|---|---|---|---|
| Variable | r2 | RE | CE | MSE | RE | CE | MSE |
| JJA | 0.38 | 0.43 | 0.39 | 0.65 | 0.37 | 0.34 | 0.87 |
| JA | 0.37 | 0.31 | 0.29 | 0.96 | 0.40 | 0.39 | 1.03 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Levanič, T.; Jevšenak, J.; Hafner, P. Stable Isotopes Reveal Climate Signal Hidden in Tree Rings of Endemic Balkan Pines. Atmosphere 2020, 11, 135. https://doi.org/10.3390/atmos11020135
Levanič T, Jevšenak J, Hafner P. Stable Isotopes Reveal Climate Signal Hidden in Tree Rings of Endemic Balkan Pines. Atmosphere. 2020; 11(2):135. https://doi.org/10.3390/atmos11020135
Chicago/Turabian StyleLevanič, Tom, Jernej Jevšenak, and Polona Hafner. 2020. "Stable Isotopes Reveal Climate Signal Hidden in Tree Rings of Endemic Balkan Pines" Atmosphere 11, no. 2: 135. https://doi.org/10.3390/atmos11020135
APA StyleLevanič, T., Jevšenak, J., & Hafner, P. (2020). Stable Isotopes Reveal Climate Signal Hidden in Tree Rings of Endemic Balkan Pines. Atmosphere, 11(2), 135. https://doi.org/10.3390/atmos11020135