
Relationship between Greenhouse Gas Budget and Soil Carbon Storage Measured on Site in Zhalainuoer Grassland Mining Area
Abstract
:1. Introduction
2. Materials and Methods
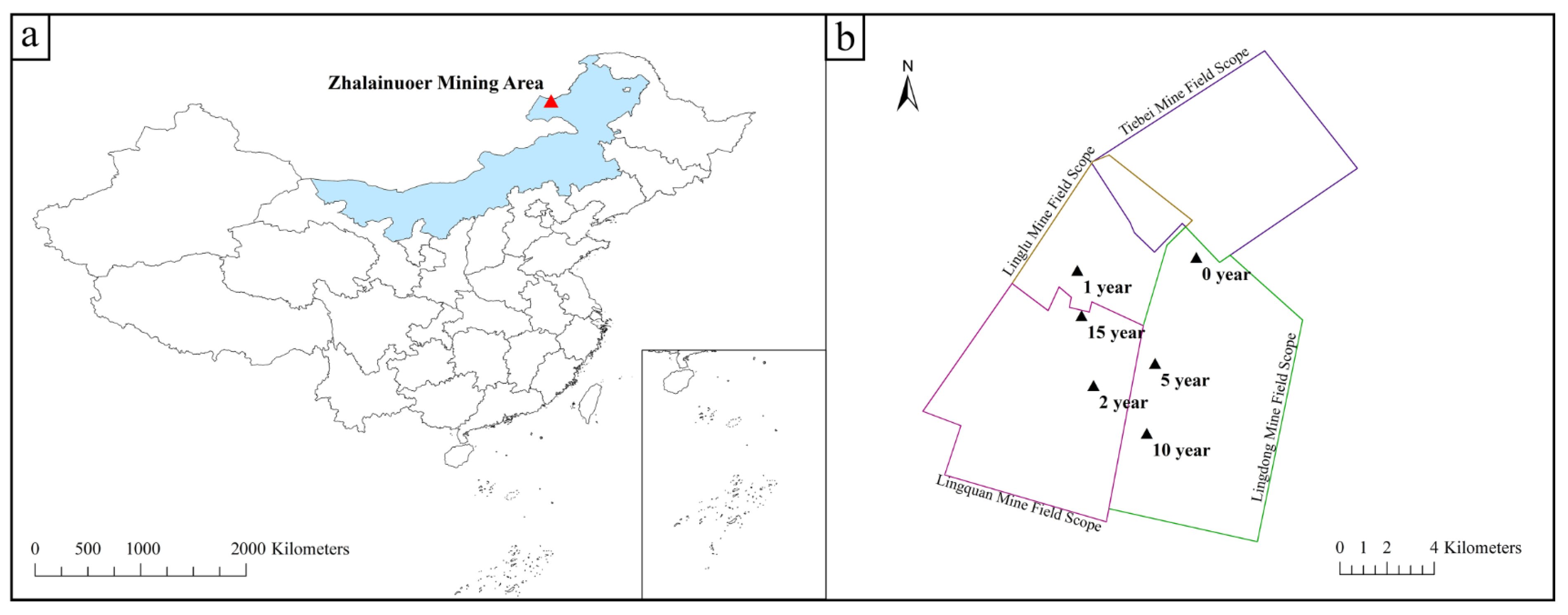
2.1. Overview of the Study Area
2.2. Research Methods
2.2.1. Sample Collection
2.2.2. Determination of the Soil’s Physical and Chemical Properties
2.2.3. Determination of Soil Microorganisms
- 1.
- DNA extraction and PCR amplification
- 2.
- Illumina PE300/PE250 sequencing
- 3.
- Amplicon sequence processing
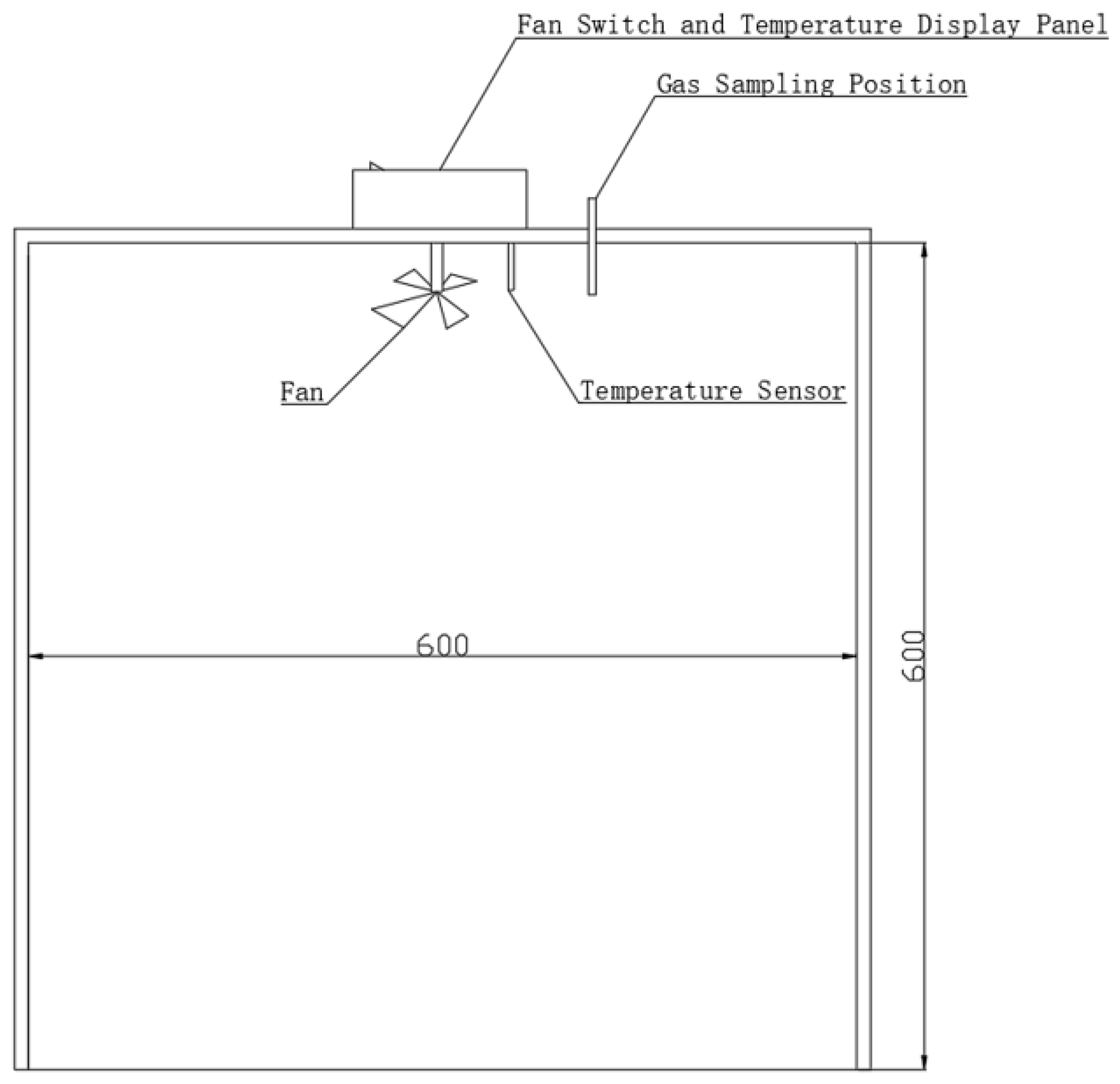
2.2.4. Determination of Gas
2.2.5. Calculation of CS and the GHG Budget
- CS
- (1)
- Selecting a carbon pool
- (2)
- Calculation of CS
- Caboveground: The aboveground CS of the herb layer (tC);
- Baboveground: The mean biomass per unit area of the aboveground part of the herb layer (t.d.m/hm2);
- CFherb: The average carbon content of herbs (tC/t.d.m). The reference value of herbaceous vegetation is 0.47.
- Csoil: Soil organic carbon storage per unit area (tC/hm2);
- SOCC: Soil organic carbon density (gC/(100 g of soil));
- BD: Volume weight of soil (g/cm3);
- F: Volume percentage of gravel, roots, and other dead residues with a soil layer diameter greater than 2 mm in the sample plot (%);
- Depth: Soil thickness (cm).
- 2.
- GHG Budget
- (1)
- Calculation of CO2 and CH4 emission
- A: Average change rate of target gas concentration in gas chamber during sealing of sampling box.
- k1 − k2 × C0: Instantaneous change rate of target gas concentration in the gas chamber at the beginning of sealing the sampling box.
- C0: Gas concentration in the air chamber of the sampling box at the beginning of sealing.
- V: Air chamber volume of sampling box.
- A: Sampling box bottom area.
- M: Molar mass of target gas.
- V0: Molar volume of target gas under standard conditions.
- T and P: They are the air temperature and air pressure in the air chamber when collecting gas samples, respectively. Considering that the sampling box has an insulation design and an air pressure balance tube, the measured values of air temperature and air pressure outside the box can be used instead.
- T0 and P0: Air temperature (273 K) and air pressure (1013 hPa) under standard conditions.
- C: Dimension conversion coefficient.
- (2)
- The calculation of the GHG budget
- GHG = CO2 + (CH4 × 25);
- GHG: The GHG flux in and out of the atmosphere, taking the GWP value of CH4 as 25;
- CO2: The carbon dioxide mission flux;
- CH4: CH4 emission.
2.3. Data Processing and Analysis
3. Results
3.1. Physical and Chemical Properties of Vegetation and Soil and Distribution Characteristics of Microbial Communities
3.1.1. Distribution Characteristics of Physical and Chemical Properties of Vegetation and Soil
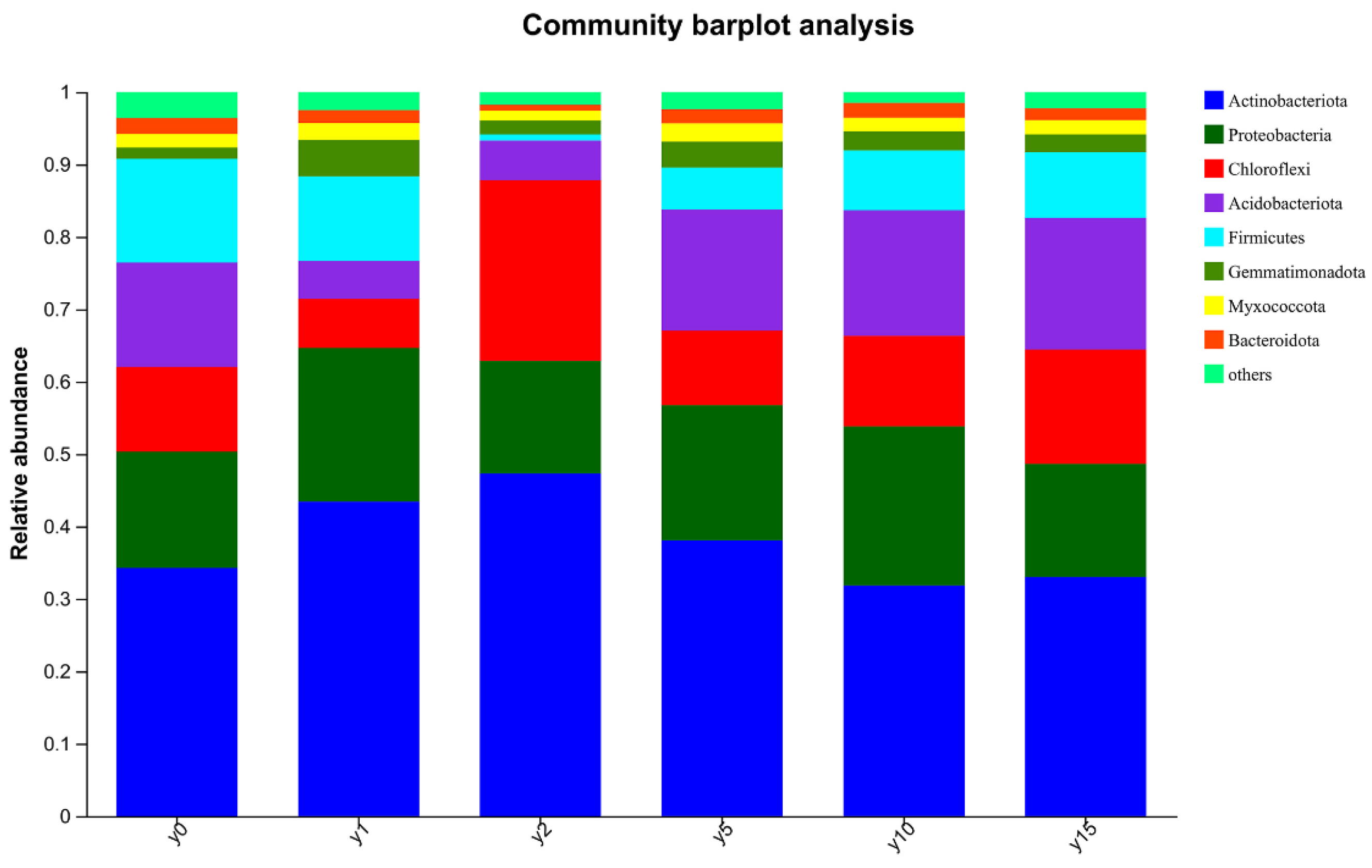
3.1.2. Distribution Characteristics of Soil Microbial Community
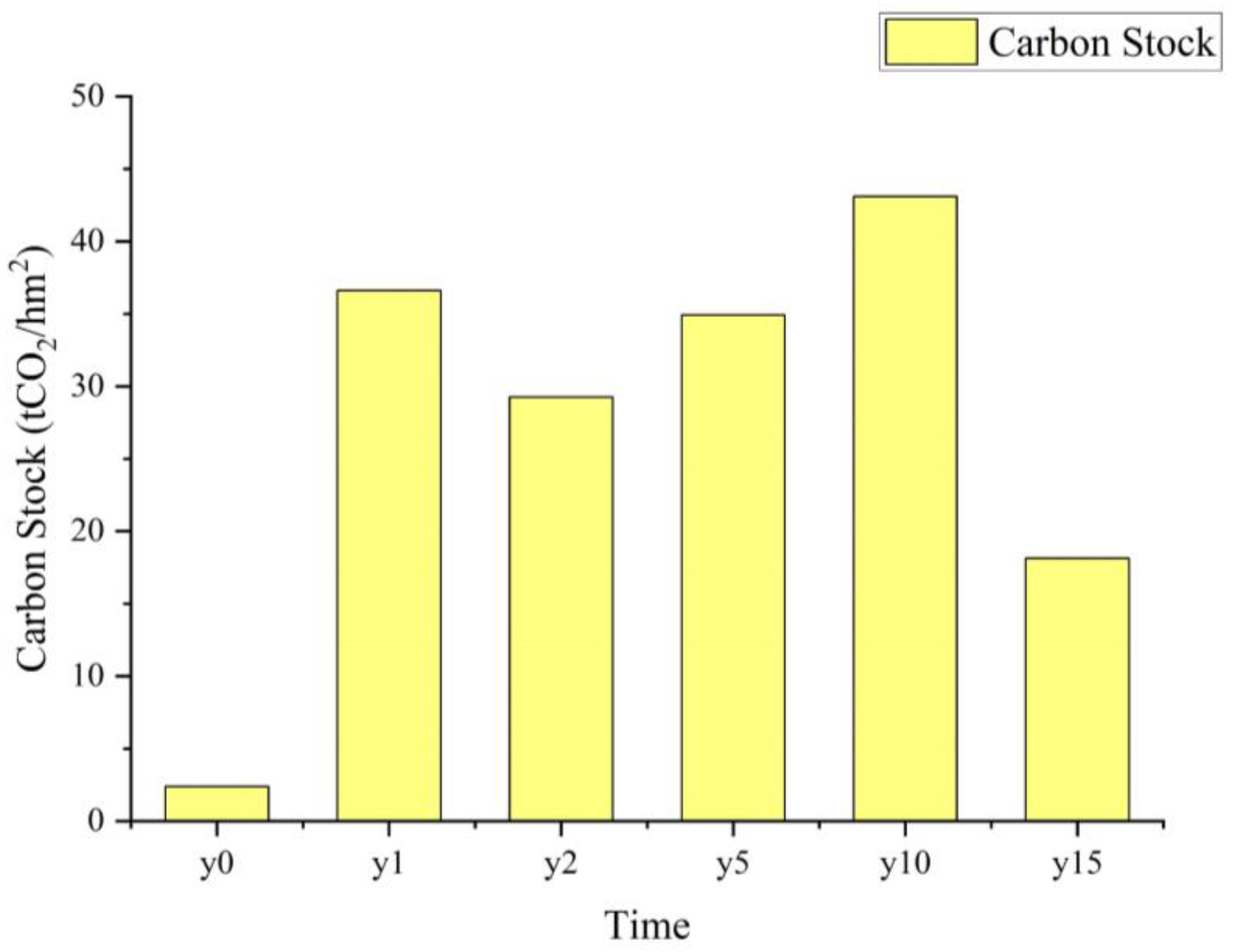
3.2. Distribution Characteristics of CS
3.3. Distribution Characteristics of the GHG Budget
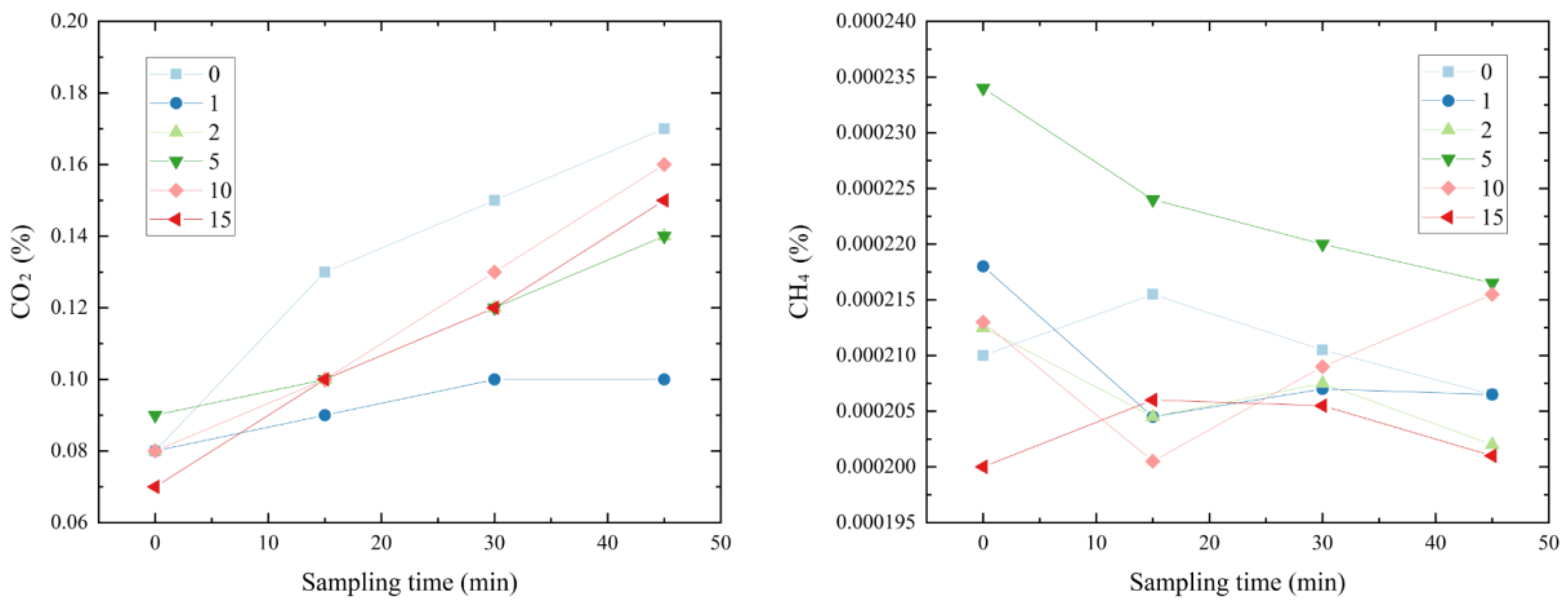
3.3.1. Distribution Characteristics of CO2 and CH4 Concentrations
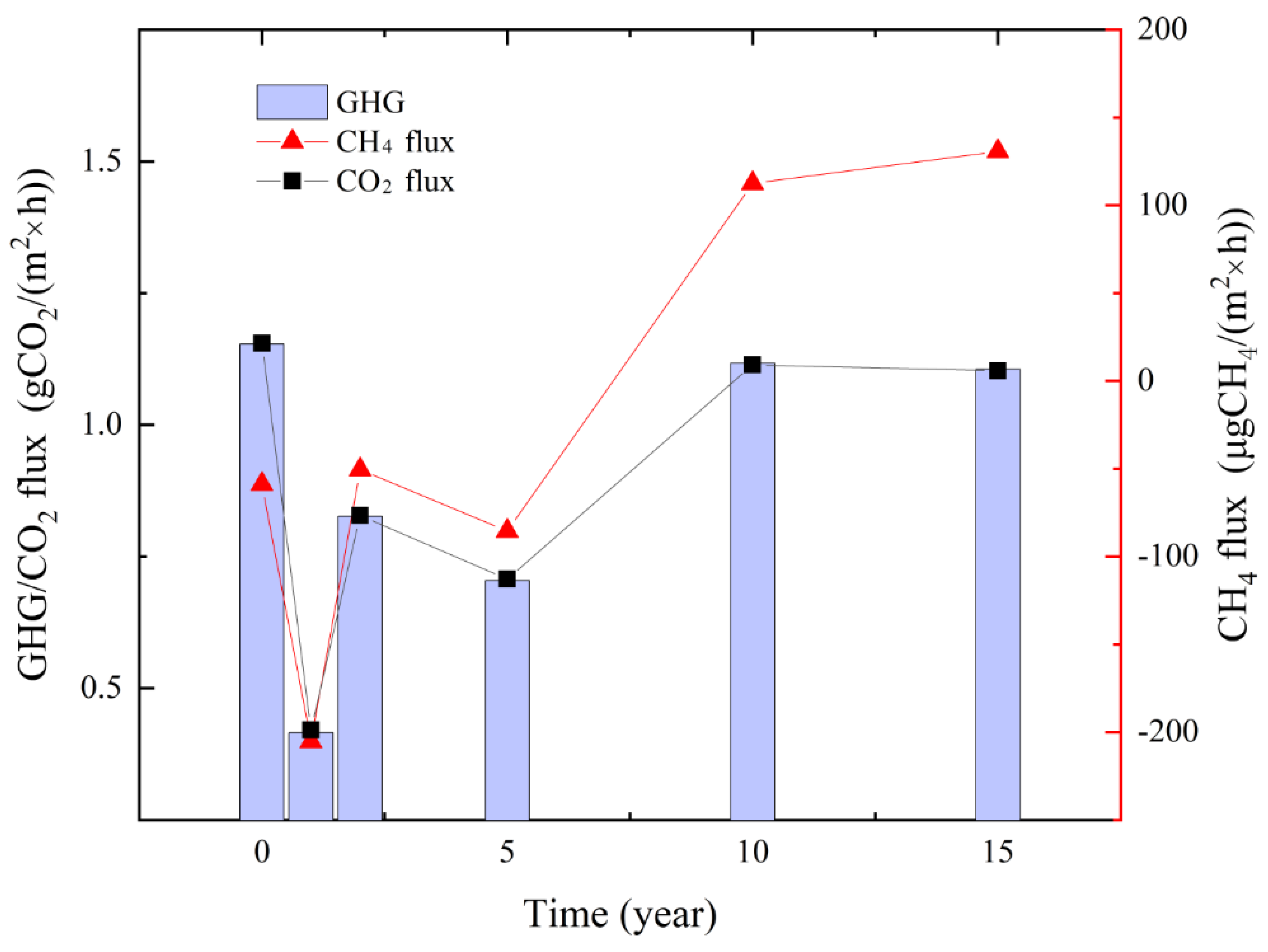
3.3.2. Characteristics of the GHG Budget
3.4. Correlation Analysis Results
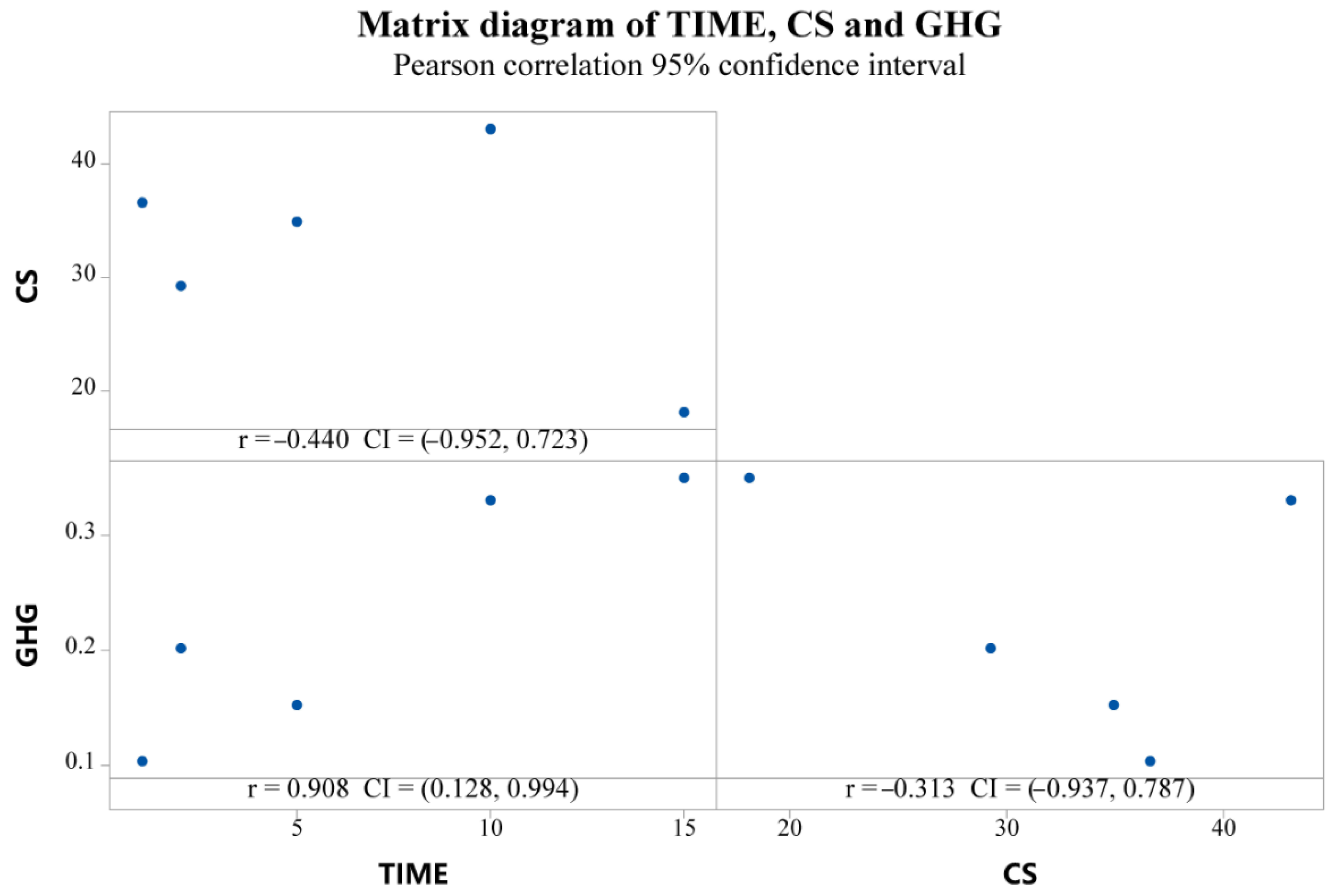
3.4.1. Correlation between CS and the GHG Budget
3.4.2. Correlation of CS and the GHG Budget with Physical and Chemical Properties of Vegetation and Soil
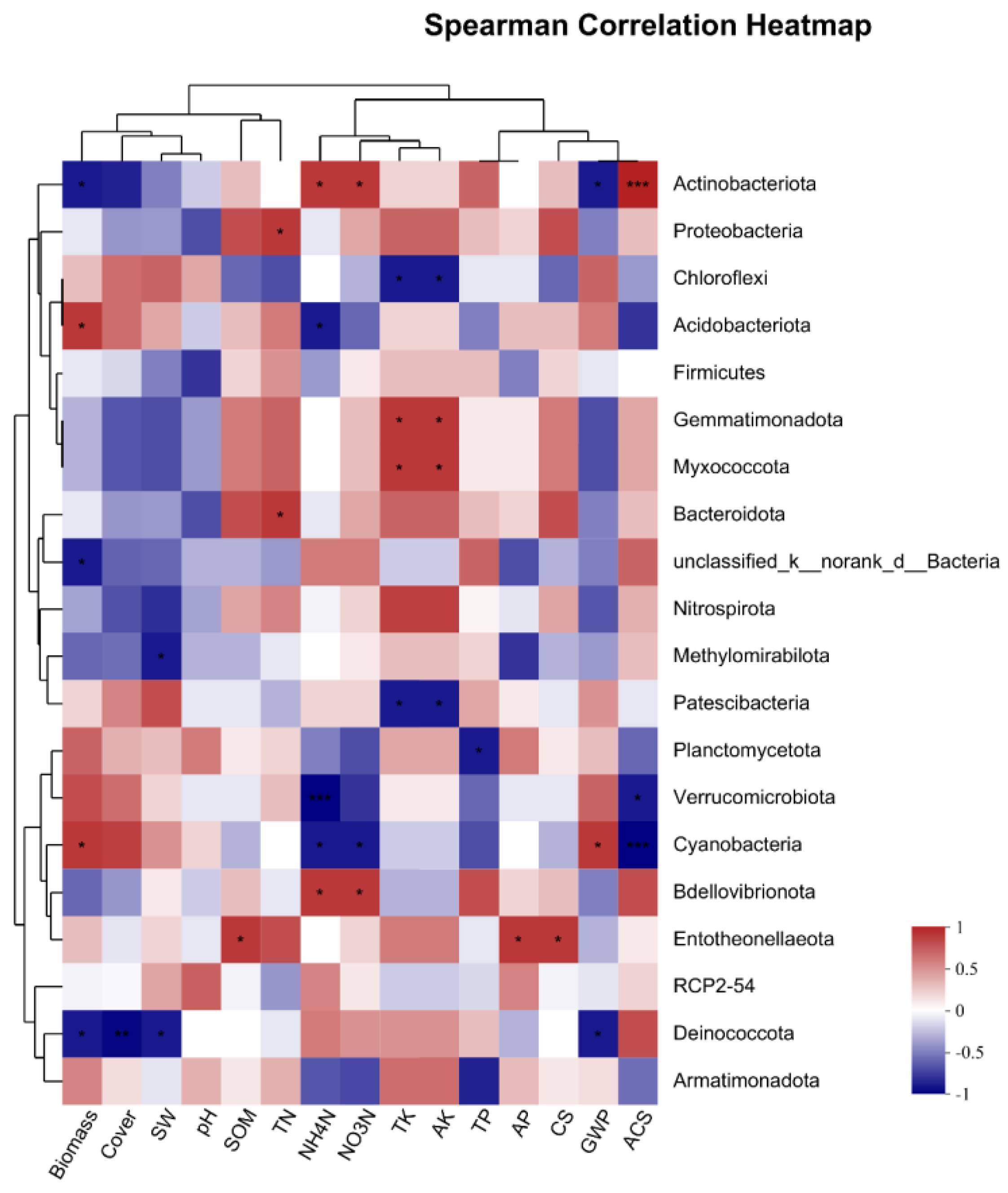
3.4.3. Correlation of CS and the GHG Budget with Soil Microorganisms
4. Discussion
4.1. Relationship between CS and the GHG Budget
4.2. Influencing Factors of CS
4.3. Influencing Factors of the GHG Budget
4.4. Inspiration for Carbon Sequestration in Grassland Mining Area
5. Conclusions
- There is a significant negative correlation between grassland CS and the GHG budget in different subsidence periods. The accurate and universal parameters to describe their relationship need more measured data to compare and verify.
- The CS in grasslands during various subsidence periods exhibits a significant positive correlation with both soil organic matter and available phosphorus, as well as with Entotheonellaeota bacteria. The underlying mechanism may involve organic matter and available phosphorus acting as substrates to facilitate the biological activity of Entotheonellaeota, thereby influencing the CS dynamics. Therefore, soil has the main role of storing carbon.
- The GHG budget is mainly affected by CO2 emission, which is positively correlated with plant biomass and coverage, negatively correlated with the bacteria Actinobacteriota and Deinococcota, and positively correlated with Cyanobacteria and with the fungus Ascomycota. Acidobacteria has carbon fixation genes. The fungus Ascomycota and Cyanobacteria coexist, and they jointly affect the GHG budget by affecting the CO2 emission. Therefore, sequestration is a physiological process conducted by plants (i.e., grassland) and organisms (including some soil organisms but not all of them).
- In line with the prevailing notion of the ecosystem CS, understanding the relationship between CS and GHG flux can aid in estimating the extent of grassland restoration required. Enhancing soil texture or applying phosphate fertilizer can boost grassland CS. The bacterium Entotheonellaeota, along with soil physicochemical properties, can serve as an indicator of microorganism activity and soil CS. In addition to increasing plant biomass, the activities of carbon-fixing microorganisms like cyanobacteria and Ascomycota can directly reduce GHG emissions.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luo, Y.X.; Li, Y.X.; Liu, S.W.; Yu, P.J. Effects of vegetation succession on soil organic carbon fractions and stability in a karst valley area, Southwest China. Environ. Monit. Assess. 2022, 194, 562. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Cotrufo, M.F. Grassland soil carbon sequestration: Current understanding, challenges, and solutions. Science 2022, 377, 603–608. [Google Scholar] [CrossRef]
- Khan, M.Z.; Chiti, T. Soil carbon stocks and dynamics of different land uses in Italy using the LUCAS soil database. J. Environ. Manag. 2022, 306, 114452. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, M.W.I.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kogel-Knabner, I.; Lehmann, J.; Manning, D.A.C.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Dang, Z.; Guo, N.; Li, S.; Degen, A.A.; Cao, J.; Deng, B.; Wang, A.; Peng, Z.; Ding, L.; Long, R.; et al. Effect of grazing exclusion on emission of greenhouse gases and soil organic carbon turnover in alpine shrub meadow. Sci. Total Environ. 2023, 858, 159758. [Google Scholar] [CrossRef]
- Li, S.; Chen, P.; Mei, B.; Yue, H.; Zheng, X.; Ren, G.; Aruhan, S. Annual methane uptake of an artificial grassland under different grazing strategies. Nutr. Cycl. Agroecosyst. 2023, 125, 29–42. [Google Scholar] [CrossRef]
- Perez-Quezada, J.F.; Cano, S.; Ibaceta, P.; Aguilera-Riquelme, D.; Salazar, O.; Fuentes, J.P.; Osborne, B. How do land cover changes affect carbon-nitrogen-phosphorus stocks and the greenhouse gas budget of ecosystems in southern Chile? Agric. Ecosyst. Environ. 2022, 340, 108153. [Google Scholar] [CrossRef]
- Jin, Y.; Tian, D.; Li, J.; Wu, Q.; Pan, Z.; Han, M.; Wang, Y.; Zhang, J.; Han, G. Water causes divergent responses of specific carbon sink to long-term grazing in a desert grassland. Sci. Total Environ. 2023, 873, 162166. [Google Scholar] [CrossRef]
- Chang, J.; Ciais, P.; Gasser, T.; Smith, P.; Herrero, M.; Havlík, P.; Obersteiner, M.; Guenet, B.; Goll, D.S.; Li, W. Climate warming from managed grasslands cancels the cooling effect of carbon sinks in sparsely grazed and natural grasslands. Nat. Commun. 2021, 12, 118. [Google Scholar] [CrossRef]
- Rafalska, A.; Walkiewicz, A.; Osborne, B.; Klumpp, K.; Bieganowski, A. Variation in methane uptake by grassland soils in the context of climate change—A review of effects and mechanisms. Sci. Total Environ. 2023, 871, 162127. [Google Scholar] [CrossRef]
- Jin, X.; Wu, F.; Wu, Q.; Hedenec, P.; Peng, Y.; Wang, Z.; Yue, K. Effects of drying-rewetting cycles on the fluxes of soil greenhouse gases. Heliyon 2023, 9, e12984. [Google Scholar] [CrossRef] [PubMed]
- Zuo, X.; Zhang, J.; Zhou, X.; Zhao, X.; Wang, S.; Lian, J.; Lv, P.; Knops, J. Changes in carbon and nitrogen storage along a restoration gradient in a semiarid sandy grassland. Acta Oecol. Int. J. Ecol. 2015, 69, 1–8. [Google Scholar] [CrossRef]
- He, S.; Liang, Z.; Han, R.; Wang, Y.; Liu, G. Soil carbon dynamics during grass restoration on abandoned sloping cropland in the hilly area of the Loess Plateau, China. Catena 2016, 137, 679–685. [Google Scholar] [CrossRef]
- Mu, Y.; Liu, Y.; Tian, F.-P.; Chang, X.-F.; Wu, G.-L. Influence of artificial grassland restoration on soil carbon pool in an arid mining land. J. Soil Sci. Plant Nutr. 2016, 16, 890–900. [Google Scholar] [CrossRef]
- Shrestha, R.K.; Lal, R.; Penrose, C. Greenhouse Gas Emissions and Global Warming Potential of Reclaimed Forest and Grassland Soils. J. Environ. Qual. 2009, 38, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.E.; Campbell, J.E.; Acton, P.M. Carbon Sequestration by Reforesting Legacy Grasslands on Coal Mining Sites. Energies 2020, 13, 6340. [Google Scholar] [CrossRef]
- Qu, R.; He, L.; He, Z.; Wang, B.; Lyu, P.; Wang, J.; Kang, G.; Bai, W. A Study of Carbon Stock Changes in the Alpine Grassland Ecosystem of Zoige, China, 2000–2020. Land 2022, 11, 1232. [Google Scholar] [CrossRef]
- Li, Z.; Tang, Q.; Wang, X.; Chen, B.; Sun, C.; Xin, X. Grassland Carbon Change in Northern China under Historical and Future Land Use and Land Cover Change. Agron. Basel 2023, 13, 2180. [Google Scholar] [CrossRef]
- Xu, Z.; Zhao, Z.; Lu, C. The Contribution of Land Use and Land Cover on Carbon Storage in the North Tibet Plateau, China. J. Anim. Plant Sci. 2021, 31, 1598–1609. [Google Scholar] [CrossRef]
- Conrad, R. Microbial Ecology of Methanogens and Methanotrophs. Adv. Agron. 2007, 96, 1–63. [Google Scholar]
- Li, T.; Zhou, Q.X. The key role of Geobacter in regulating emissions and biogeochemical cycling of soil-derived greenhouse gases. Environ. Pollut. 2020, 266, 115135. [Google Scholar] [CrossRef] [PubMed]
- Davidson, E.A.; Savage, K.; Verchot, L.V.; Navarro, R. Minimizing artifacts and biases in chamber-based measurements of soil respiration. Agric. For. Meteorol. 2002, 113, 21–37. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis; Wiley: Hoboken, NJ, USA, 1996; pp. 961–1010. [Google Scholar]
- Liu, C.S.; Zhao, D.F.; Ma, W.J.; Guo, Y.D.; Wang, A.J.; Wang, Q.L.; Lee, D.J. Denitrifying sulfide removal process on high-salinity wastewaters in the presence of Halomonas sp. Appl. Microbiol. Biotechnol. 2016, 100, 1421–1426. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.F.; Zhou, Y.Q.; Chen, Y.R.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Norderhaug, A.; Clemmensen, K.E.; Kardol, P.; Thorhallsdottir, A.G.; Aslaksen, I. Carbon sequestration potential and the multiple functions of Nordic grasslands. Clim. Chang. 2023, 176, 55. [Google Scholar] [CrossRef]
- Uttaruk, Y.; Laosuwan, T. Development of Prototype Project for Carbon Storage and Greenhouse Gas Emission Reduction from Thailand’s Agricultural Sector. Sains Malays. 2019, 48, 2083–2092. [Google Scholar] [CrossRef]
- Shi, H.; Shen, H.; Dong, S.; Xiao, J.; Mu, Z.; Zhang, R.; Hao, X.; Wang, Z.; Zuo, H. Six Years of Grassland Cultivation Promotes CO2, N2O Emissions and CH4 Uptake with Increasing N Deposition on Qinghai-Tibetan Plateau. Sustainability 2022, 14, 11434. [Google Scholar] [CrossRef]
- Liao, J.; Yang, X.; Dou, Y.; Wang, B.; Xue, Z.; Sun, H.; Yang, Y.; An, S. Divergent contribution of particulate and mineral-associated organic matter to soil carbon in grassland. J. Environ. Manag. 2023, 344, s118536. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.S.; Mir, S.A.; Wani, O.A.; Babu, S.; Yeasin, M.; Bhat, M.A.; Hussain, N.; Wani, A.I.A.; Kumar, R.; Yadav, D.; et al. Land-use systems regulate carbon geochemistry in the temperate Himalayas, India. J. Environ. Manag. 2022, 320, 115811. [Google Scholar] [CrossRef] [PubMed]
- Wani, O.A.; Kumar, S.S.; Hussain, N.; Wani, A.I.A.; Babu, S.; Alam, P.; Rashid, M.; Popescu, S.M.; Mansoor, S. Multi-scale processes influencing global carbon storage and land-carbon-climate nexus: A critical review. Pedosphere 2023, 33, 250–267. [Google Scholar] [CrossRef]
- Hu, L.; Li, Q.; Yan, J.H.; Liu, C.; Zhong, J.X. Vegetation restoration facilitates belowground microbial network complexity and recalcitrant soil organic carbon storage in southwest China karst region. Sci. Total Environ. 2022, 820, 153137. [Google Scholar] [CrossRef]
- Xue, P.P.; Minasny, B.; McBratney, A.; Pino, V.; Fajardo, M.; Luo, Y. Distribution of soil bacteria involved in C cycling across extensive environmental and pedogenic gradients. Eur. J. Soil Sci. 2023, 74, e13337. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Ranalli, M.G.; Haddix, M.L.; Six, J.; Lugato, E. Soil carbon storage informed by particulate and mineral-associated organic matter. Nat. Geosci. 2019, 12, 989–994. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, Q.; Fan, S.; Zhang, Y.; Zhang, M.; Zhang, J. Distinction between Cr and other heavy-metal-resistant bacteria involved in C/N cycling in contaminated soils of copper producing sites. J. Hazard. Mater. 2021, 402, 123454. [Google Scholar] [CrossRef]
- Lu, X.Y.; Wen, L.; Sun, H.Y.; Fei, T.; Liu, H.; Ha, S.N.; Tang, S.M.; Wang, L.X. Responses of soil respiration to phosphorus addition in global grasslands: A meta-analysis. J. Clean. Prod. 2022, 349, 131413. [Google Scholar] [CrossRef]
- Chenyue, Z.; Xia, Z.; Yuchun, X.; Wenjia, T.; Lei, W. Carbon storage and distribution of grassland ecosystems in Qinghai Province. J. Beijing Norm. Univ. Nat. Sci. 2022, 58, 286–292. [Google Scholar]
- Valjavec, M.B.; Carni, A.; Zlindra, D.; Zorn, M.; Marinsek, A. Soil organic carbon stock capacity in karst dolines under different land uses. Catena 2022, 218, 106548. [Google Scholar] [CrossRef]
- Di, S.; Zong, M.M.; Li, S.Y.; Li, H.X.; Duan, C.Q.; Peng, C.H.; Zhao, Y.G.; Bai, J.Y.; Lin, C.; Feng, Y.; et al. The effects of the soil environment on soil organic carbon in tea plantations in Xishuangbanna, southwestern China. Agric. Ecosyst. Environ. 2020, 297, 106951. [Google Scholar] [CrossRef]
- Li, J.H.; Han, Y.W.; Ye, L.F.; Deng, H.D.; Gao, X.T.; Soromotin, A.; Kuzyakov, Y.; Knops, J.M.H.; Abbott, L.K. Effects of nitrogen and phosphorus fertilization on soil organic matter priming and net carbon balance in alpine meadows. Land Degrad. Dev. 2023, 34, 2681–2692. [Google Scholar] [CrossRef]
- Bailey, V.L.; Smith, J.L.; Bolton, H. Fungal-to-bacterial ratios in soils investigated for enhanced C sequestra-tion. Soil Biol. Biochem. 2002, 34, 997–1007. [Google Scholar] [CrossRef]
- Hoertnagl, L.; Barthel, M.; Buchmann, N.; Eugster, W.; Butterbach-Bahl, K.; Diaz-Pines, E.; Zeeman, M.; Klumpp, K.; Kiese, R.; Bahn, M.; et al. Greenhouse gas fluxes over managed grasslands in Central Europe. Glob. Chang. Biol. 2018, 24, 1843–1872. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Cao, S.; Cao, G.; Chen, K.; Peng, C. The Characteristics and Seasonal Variation of Methane Fluxes from an Alpine Wetland in the Qinghai Lake watershed, China. Wetlands 2021, 41, 53. [Google Scholar] [CrossRef]
- Martins, C.S.C.; Nazaries, L.; Macdonald, C.A.; Anderson, I.C.; Singh, B.K. Water availability and abundance of microbial groups are key determinants of greenhouse gas fluxes in a dryland forest ecosystem. Soil Biol. Biochem. 2015, 86, 5–16. [Google Scholar] [CrossRef]
- Karbin, S.; Guillet, C.; Kammann, C.I.; Niklaus, P.A. Effects of Long-Term CO2 Enrichment on Soil-Atmosphere CH4 Fluxes and the Spatial Micro-Distribution of Methanotrophic Bacteria. PLoS ONE 2015, 10, e0131665. [Google Scholar] [CrossRef]
- Liu, Z. Microbial Pathways of Atmospheric Carbon Dioxide Fixation in Soils in the Mu Us Desert. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2019. [Google Scholar]
- Zhang, Q.; Wang, Q.; Ouyang, H.; Lan, S.; Hu, C. Pyrosequencing Reveals Significant Changes in Microbial Communities Along the Ecological Succession of Biological Soil Crusts in the Tengger Desert of China. Pedosphere 2018, 28, 350–362. [Google Scholar] [CrossRef]
- Bellassen, V.; Angers, D.; Kowalczewski, T.; Olesen, A. Soil carbon is the blind spot of European national GHG inventories. Nat. Clim. Chang. 2022, 12, 324–331. [Google Scholar] [CrossRef]
- Ghani, M.U.; Kamran, M.; Ahmad, I.; Arshad, A.; Zhang, C.; Zhu, W.; Lou, S.; Hou, F. Alfalfa-grass mixtures reduce greenhouse gas emissions and net global warming potential while maintaining yield advantages over monocultures. Sci. Total Environ. 2022, 849, 157765. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Plant | Soil | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Biomass | Cover | Water | pH | SOM | TN | NH4N | NO3N | TK | AK | TP | AP | |
| g | % | % | g/kg | g/kg | mg/kg | mg/kg | g/kg | mg/kg | g/kg | mg/kg | ||
| 0 | 21.29 | 40 | 12.5 | 7.06 | 7.52 | 0.60 | 59.42 | 83.20 | 0.91 | 176.50 | 0.13 | 7.25 |
| 1 | 8.20 | 10 | 15.2 | 5.92 | 36.44 | 1.13 | 48.94 | 95.08 | 5.65 | 337.40 | 1.44 | 46.59 |
| 2 | 16.39 | 44 | 17.9 | 7.47 | 31.77 | 0.48 | 53.22 | 59.46 | 2.61 | 180.08 | 0.14 | 51.19 |
| 5 | 21.28 | 40 | 15.9 | 7.51 | 34.53 | 0.66 | 39.44 | 16.24 | 6.84 | 408.79 | 0.10 | 54.12 |
| 10 | 34.85 | 90 | 35.1 | 7.14 | 42.11 | 1.14 | 32.22 | 17.55 | 4.43 | 284.76 | 0.13 | 86.40 |
| 15 | 31.39 | 90 | 16.0 | 7.15 | 20.46 | 0.51 | 31.56 | 8.09 | 3.41 | 200.00 | 0.12 | 44.34 |
| Sample | Ace | Chao | Shannon | Simpson | Sobs | |
|---|---|---|---|---|---|---|
| Bacteria | y0 | 1460 | 1460 | 6.741673 | 0.002301 | 1460 |
| y1 | 933 | 933 | 6.228809 | 0.00399 | 933 | |
| y2 | 654 | 654 | 5.955778 | 0.004601 | 654 | |
| y5 | 922 | 922 | 6.300103 | 0.003976 | 922 | |
| y10 | 932 | 932 | 6.356244 | 0.003038 | 932 | |
| y15 | 1201 | 1201 | 6.575124 | 0.002555 | 1201 |
| Biomass | Cover | Water | pH | SOM | TN | NH4N | NO3N | TK | AK | TP | AP | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Time | 0.889 * | 0.909 * | 0.424 | 0.274 | 0.135 | −0.047 | −0.910 ** | −0.852 * | 0.147 | −0.051 | −0.379 | 0.476 |
| CS | −0.095 | −0.244 | 0.629 | −0.138 | 0.998 ** | 0.640 | −0.500 | −0.276 | 0.779 | 0.665 | 0.301 | 0.903 * |
| GHG | 0.925 * | 0.986 ** | 0.415 | 0.405 | −0.354 | −0.203 | −0.425 | −0.523 | −0.539 | −0.593 | −0.646 | 0.076 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, Y.; Mao, Z.; Lu, D.; Feng, B.; Xu, L.; Zhong, L.; Yu, J. Relationship between Greenhouse Gas Budget and Soil Carbon Storage Measured on Site in Zhalainuoer Grassland Mining Area. Atmosphere 2024, 15, 25. https://doi.org/10.3390/atmos15010025
Tang Y, Mao Z, Lu D, Feng B, Xu L, Zhong L, Yu J. Relationship between Greenhouse Gas Budget and Soil Carbon Storage Measured on Site in Zhalainuoer Grassland Mining Area. Atmosphere. 2024; 15(1):25. https://doi.org/10.3390/atmos15010025
Chicago/Turabian StyleTang, Yan, Zhen Mao, Dongqiang Lu, Bo Feng, Liang Xu, Licun Zhong, and Jinbiao Yu. 2024. "Relationship between Greenhouse Gas Budget and Soil Carbon Storage Measured on Site in Zhalainuoer Grassland Mining Area" Atmosphere 15, no. 1: 25. https://doi.org/10.3390/atmos15010025