Abundance of Biological Ice Nucleating Particles in the Mississippi and Its Major Tributaries


Abstract
:1. Introduction
2. Materials and Methods
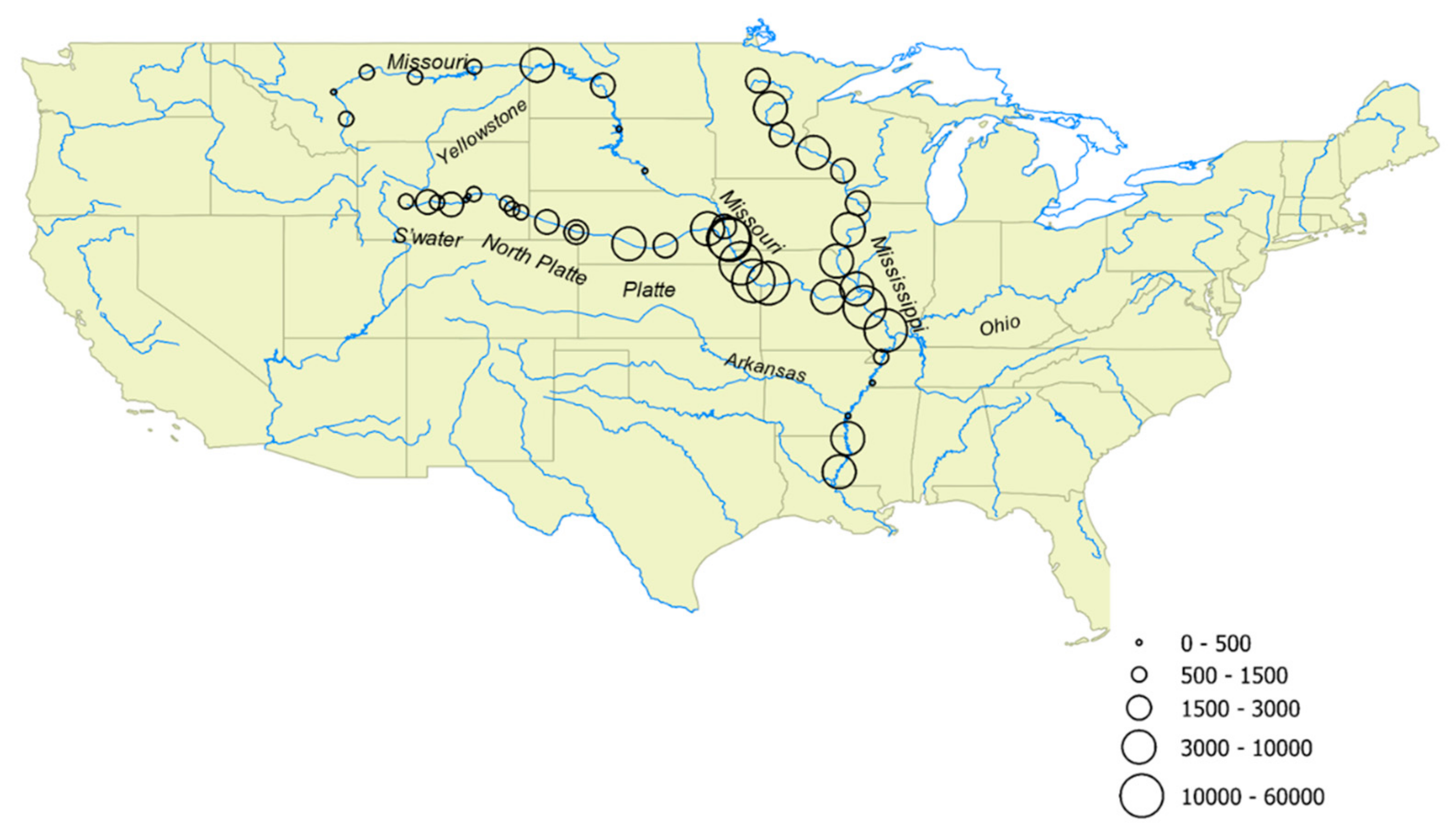
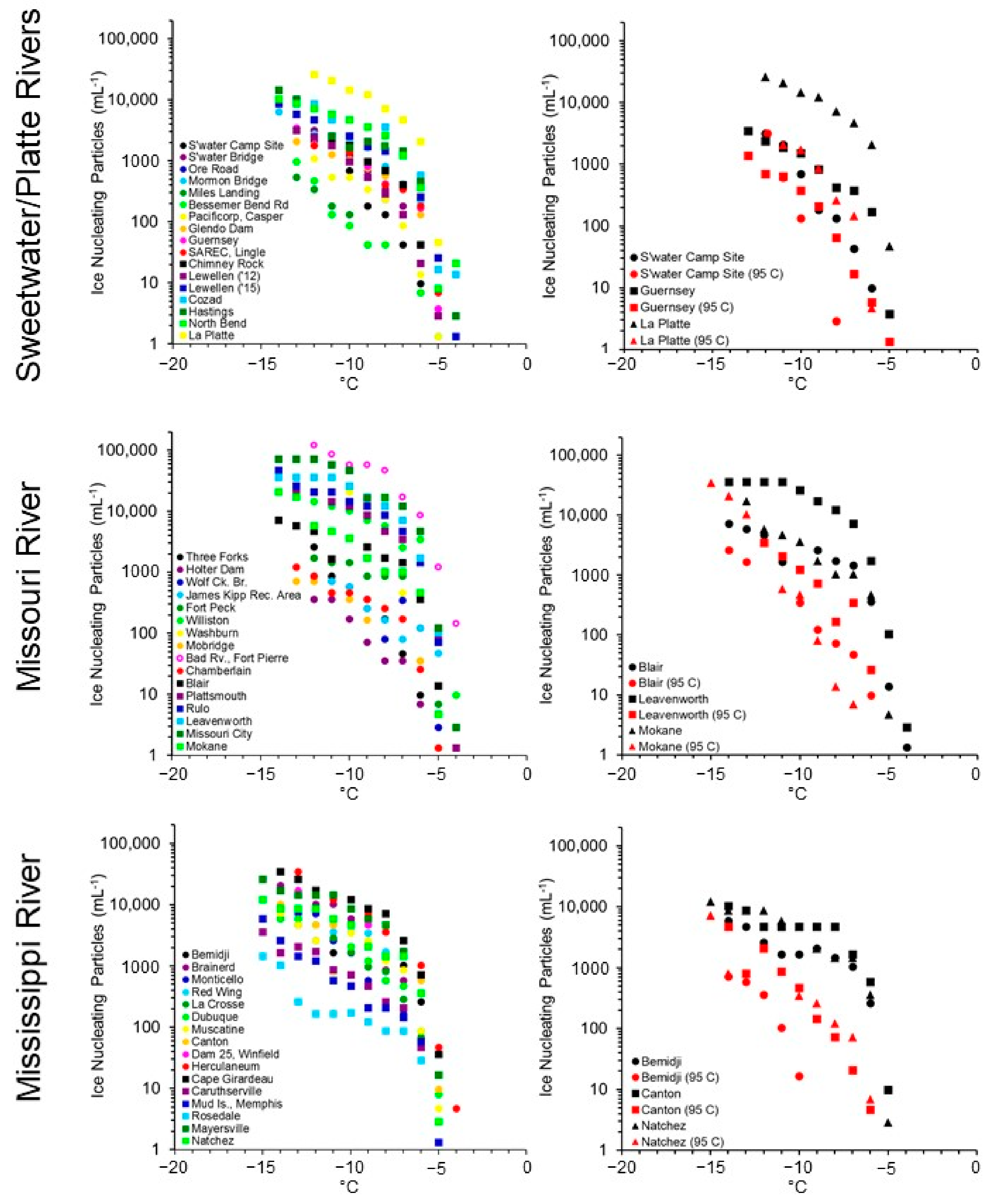
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Petters, M.D.; Wright, T.P. Revisiting ice nucleation from precipitation samples. Geophys. Res. Lett. 2015, 42, 8758–8766. [Google Scholar] [CrossRef]
- Lohmann, U.; Feichter, J. Global indirect aerosol effects: A review. Atmos. Chem. Phys. 2005, 5, 715–737. [Google Scholar] [CrossRef]
- DeMott, P.J.; Prenni, J.A.; Liu, X.; Kreidenweis, S.M.; Petters, M.D.; Twohy, C.H.; Richardson, M.S.; Eidhammer, T.; Rogers, D.C. Predicting global atmospheric ice nuclei distributions and their impacts on climate. Proc. Natl. Acad. Sci. USA 2010, 107, 11217–11222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, B.J.; O’Sullivan, D.; Atkinson, J.D.; Webb, M. Ice nucleation by particles immersed in supercooled cloud droplets. Chem. Soc. Rev. 2012, 41, 6519–6554. [Google Scholar] [CrossRef] [PubMed]
- Boucher, O.; Randall, D.; Artaxo, P.; Bretherton, C.; Feingold, G.; Forster, P.; Kerminen, V.-M.; Kondo, Y.; Liao, H.; Lohmann, U.; et al. Clouds and Aerosols. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- Hoose, C.; Möhler, O. Heterogeneous ice nucleation on atmospheric aerosols: A review of results from laboratory experiments. Atmos. Chem. Phys. 2012, 12, 9817–9854. [Google Scholar] [CrossRef]
- Niemand, M.; Möhler, O.; Vogel, B.; Vogel, H.; Hoose, C.; Connolly, P.; Klein, H.; Bingemer, H.; DeMott, P.; Skrotzki, J.; et al. Parameterization of immersion freezing on mineral dust particles: An application in a regional scale model. J. Atmos. Sci. 2012, 69, 3077–3092. [Google Scholar] [CrossRef]
- Boose, Y.; Welti, A.; Atkinson, J.; Ramelli, F.; Danielczok, A.; Bingemer, H.G.; Plötze, M.; Sierau, B.; Kanji, Z.A.; Lohmann, U. Heterogeneous ice nucleation on dust particles sourced from nine deserts worldwide—Part 1: Immersion freezing. Atmos. Chem. Phys. 2016, 16, 15075–15095. [Google Scholar] [CrossRef]
- Conen, F.; Morris, C.E.; Leifeld, J.; Yakutin, M.V.; Alewell, C. Biological residues define the ice nucleation properties of soil dust. Atmos. Chem. Phys. 2011, 11, 9643–9648. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, D.; Murray, B.J.; Malkin, T.L.; Whale, T.F.; Umo, N.S.; Atkinson, J.D.; Price, H.C.; Baustian, K.J.; Browse, J.; Webb, M.E. Ice nucleation by fertile soil dusts: Relative importance of mineral and biogenic components. Atmos. Chem. Phys. 2014, 14, 1853–1867. [Google Scholar]
- Tobo, Y.; DeMott, P.J.; Hill, T.C.J.; Prenni, A.J.; Swoboda-Colberg, N.G.; Franc, G.D.; Kreidenweis, S.M. Organic matter matters for ice nuclei of agricultural soil origin. Atmos. Chem. Phys. 2014, 14, 8521–8531. [Google Scholar] [CrossRef] [Green Version]
- Hill, T.C.J.; DeMott, P.J.; Tobo, Y.; Fröhlich-Nowoisky, J.; Moffett, B.F.; Franc, G.D.; Kreidenweis, S.M. Sources of organic ice nucleating particles in soils. Atmos. Chem. Phys. 2016, 16, 7195–7211. [Google Scholar] [CrossRef] [Green Version]
- Schnell, R.C.; Vali, G. Biogenic ice nuclei, Part I: Terrestrial and marine sources. J. Atmos. Sci. 1976, 33, 1554–1564. [Google Scholar] [CrossRef]
- Vali, G.; Christensen, M.; Fresh, R.W.; Galyan, E.L.; Maki, L.R.; Schnell, R.C. Biogenic ice nuclei Part II, Bacterial sources. J. Atmos. Sci. 1976, 33, 1565–1570. [Google Scholar]
- Burrows, S.M.; Hoose, C.; Pöschl, U.; Lawrence, M.G. Ice nuclei in marine air: Bioparticles or dust? Atmos. Chem. Phys. 2011, 13, 245–267. [Google Scholar] [CrossRef]
- Wang, X.; Sultana, C.M.; Trueblood, J.; Hill, T.C.J.; Malfatti, F.; Lee, C.; Laskina, O.; Moore, K.A.; Beall, C.M.; McCluskey, C.S.; et al. Microbial control of sea spray aerosol composition: A tale of two blooms. ACS Cent. Sci. 2015, 1, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.W.; Ladino, L.A.; Alpert, P.A.; Breckels, M.N.; Brooks, I.M.; Browse, J.; Burrows, S.M.; Carslaw, K.S.; Huffman, J.A.; Judd, C.; et al. A marine biogenic source of atmospheric ice-nucleating particles. Nature 2015, 525, 234–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeMott, P.J.; Hill, T.C.J.; McCluskey, C.S.; Prather, K.A.; Collins, D.B.; Sullivan, R.C.; Ruppel, M.J.; Mason, R.H.; Irish, V.E.; Lee, T.; et al. Sea spray aerosol as a unique source of ice nucleating particles. Proc. Natl. Acad. Sci. USA 2016, 113, 5797–5803. [Google Scholar] [CrossRef] [PubMed]
- McCluskey, C.S.; Hill, T.C.J.; Malfatti, F.; Sultana, C.M.; Lee, C.; Santander, M.V.; Beall, C.M.; Moore, K.A.; Cornwell, G.C.; Collins, D.B.; et al. A dynamic link between ice nucleating particles released in nascent sea spray aerosol and oceanic biological activity during two mesocosm experiments. J. Atmos. Sci. 2017, 74, 151–166. [Google Scholar] [CrossRef]
- McCluskey, C.S.; Hill, T.C.J.; Sultana, C.M.; Laskina, O.; Trueblood, J.; Santander, M.V.; Beall, C.M.; Michaud, J.M.; Kreidenweis, S.M.; Prather, K.A.; et al. A mesocosm double feature: Insights into the chemical make-up of marine ice nucleating particles. J. Atmos. Sci. 2018, 75, 2405–2423. [Google Scholar] [CrossRef]
- Vergara-Temprado, J.; Miltenberger, A.K.; Furtado, K.; Grosvenor, D.P.; Shipway, B.J.; Hill, A.A.; Wilkinson, J.M.; Field, P.R.; Murray, B.J.; Carslaw, K.S. Strong control of Southern Ocean cloud reflectivity by ice-nucleating particles. P. Natl. Acad. Sci. 2018, 115, 2687–2692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigg, E.K. Ice nucleus concentrations in remote areas. J. Atmos. Sci. 1973, 30, 1153–1157. [Google Scholar] [CrossRef]
- McCluskey, C.S.; Ovadnevaite, J.; Rinaldi, M.; Atkinson, J.; Belosi, F.; Ceburnis, D.; Salvatore, M.; Hill, T.C.J.; Lohmann, U.; Kanji, Z.A.; et al. Marine and terrestrial organic ice-nucleating particles in pristine marine to continentally influenced Northeast Atlantic air masses. J. Geophys. Res. Atmos. 2018, 123. [Google Scholar] [CrossRef]
- Schnell, R.C.; Vali, G. Freezing nuclei in marine waters. Tellus 1975, 27, 321–323. [Google Scholar] [CrossRef]
- Schnell, R. Ice nuclei in seawater, fog water and marine air off the coast of Nova Scotia: Summer 1975. J. Atmos. Sci. 1977, 34, 1299–1305. [Google Scholar] [CrossRef]
- Engel, A.; Bange, H.W.; Cunliffe, M.; Burrows, S.M.; Friedrichs, G.; Galgani, L.; Herrmann, H.; Hertkorn, N.; Johnson, M.; Liss, P.S.; et al. The ocean’s vital skin: Toward an integrated understanding of the sea surface microlayer. Front. Mar. Sci. 2017, 4, 165. [Google Scholar] [CrossRef]
- Morris, C.E.; Sands, D.C.; Vinatzer, B.A.; Glaux, C.; Guilbaud, C.; Buffière, A.; Yan, S.; Dominguez, H.; Thompson, B.M. The life history of the plant pathogen Pseudomonas syringae is linked to the water cycle. Int. Soc. Microb. Ecol. 2008, 2, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.E.; Sands, D.C.; Vanneste, J.L.; Montarry, J.; Oakley, B.; Guilbaud, C.; Glaux, C. Inferring the evolutionary history of the plant pathogen Pseudomonas syringae from its biogeography in headwaters of rivers in North America, Europe, and New Zealand. Am. Soc. Microbol. 2010, 1, 1–11. [Google Scholar] [CrossRef] [PubMed]
- D’souza, N.A.; Kawarasaki, Y.; Gantz, J.D.; Lee, R.D., Jr.; Beall, B.F.N.; Shtarkman, Y.M.; Koçer, Z.A.; Rogers, S.O.; Wildschutte, H.; Bullerjahn, G.S.; et al. Diatom assemblages promote ice formation in large lakes. Int. Soc. Microb. Ecol. 2013, 7, 1632–1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietsch, R.B.; David, R.F.; Marr, L.C.; Vinatzer, B.; Schmale, D.G., III. Aerosolization of two strains (Ice+ and Ice–) of Pseudomonas syringae in a collision nebulizer at different temperatures. Aerosol Sci. Technol. 2015, 49, 159–166. [Google Scholar] [CrossRef]
- Moffett, B.F. Fresh water ice nuclei. Fundam. Appl. Limnol. 2016, 188, 19–23. [Google Scholar] [CrossRef]
- Larsen, J.A.; Conen, F.; Alewell, C. Export of ice nucleating particles from a watershed. R. Soc. Open Sci. 2017, 4, 170213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, T.C.J.; Georgakopoulos, D.G.; DeMott, P.J.; Stump, W.L.; Franc, G.D. Measurement of ice nucleation-active bacteria on plants and in precipitation by quantitative PCR. Appl. Environ. Microbiol. 2014, 80, 256–1267. [Google Scholar] [CrossRef] [PubMed]
- Vali, G. Quantitative evaluation of experimental results on the heterogeneous freezing nucleation of supercooled liquids. J. Atmos. Sci. 1971, 28, 402–409. [Google Scholar] [CrossRef]
- Vali, G. Sizes of atmospheric ice nuclei. Nature 1966, 212, 384–385. [Google Scholar] [CrossRef]
- Rangel-Alvarado, R.B.; Nazarenko, Y.; Ariya, P.A. Snow-borne nanosized particles: Abundance, distribution, composition, and significance in ice nucleation processes. J. Geophys. Res. Atmos. 2015, 120, 11760–11774. [Google Scholar] [CrossRef]
- Šantl-Temkiv, T.; Sahyoun, M.; Finster, K.; Hartmann, S.; Augustin, S.; Stratmann, F.; Wex, H.; Clauss, T.; Nielsen, N.W.; Sørensen, J.H.; et al. Characterization of airborne ice-nucleation-active bacteria and bacterial fragments. Atmos. Environ. 2015, 109, 105–117. [Google Scholar] [CrossRef]
- Fröhlich-Nowoisky, J.; Hill, T.C.J.; Pummer, B.G.; Yordanova, P.; Franc, G.D.; Pöschl, U. Ice nucleation activity in the widespread soil fungus Mortierella alpina. Biogeosciences 2015, 12, 1057–1071. [Google Scholar] [CrossRef]
- Pummer, B.G.; Budke, C.; Augustin-Bauditz, S.; Niedermeier, D.; Felgitsch, L.; Kampf, C.J.; Huber, R.G.; Liedl, K.R.; Loerting, T.; Moschen, T.; et al. Ice nucleation by water-soluble macromolecules. Atmos. Chem. Phys. 2015, 15, 4077–4091. [Google Scholar] [CrossRef]
- O’Sullivan, D.; Murray, B.J.; Ross, J.; Webb, M.E. The adsorption of fungal ice-nucleating proteins on mineral dusts: A terrestrial reservoir of atmospheric ice-nucleating particles. Atmos. Chem. Phys. 2016, 16, 7879–7887. [Google Scholar]
- DeLeon-Rodriguez, N.; Lathem, T.L.; Rodriguez-R, L.M.; Barazesh, J.M.; Anderson, B.E.; Beyersdorf, A.J.; Ziemba, L.D.; Bergin, M.; Nenes, A.; Konstantinidis, K.T. Microbiome of the upper troposphere: Species composition and prevalence, effects of tropical storms, and atmospheric implications. Proc. Natl. Acad. Sci. USA 2013, 110, 2575–2580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maki, T.; Hara, K.; Iwata, A.; Lee, K.C.; Kawai, K.; Kai, K.; Kobayashi, F.; Pointing, S.B.; Archer, S.; Hasegawa, H.; et al. Variations in airborne bacterial communities at high altitudes over the Noto Peninsula (Japan) in response to Asian dust events. Atmos. Chem. Phys. 2017, 17, 11877–11897. [Google Scholar] [CrossRef] [Green Version]
- Pruppacher, H.R.; Klett, J.D. Microphysics of Clouds and Precipitation; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1997. [Google Scholar]
- Leifer, I.; Caulliez, G.; de Leeuw, G. Bubbles generated from wind-steepened breaking waves: 2. Bubble plumes, bubbles, and wave characteristics. J. Geophys. Res. Oceans 2006, 111, C06021. [Google Scholar] [CrossRef]
- Axson, J.L.; May, N.W.; Coloón-Bernal, I.D.; Pratt, K.A.; Ault, A.P. Lake spray aerosol: A chemical signature from individual ambient particles. Environ. Sci. Technol. 2016, 50, 9835–9845. [Google Scholar] [CrossRef] [PubMed]
- Knackstedt, K.A.; Moffett, B.F.; Hartmann, S.; Wex, H.; Hill, T.; Glasgo, E.; Reitz, L.; Augustiin-Bauditz, S.; Deustcher, W.H.; Beall, B.; et al. A terrestrial origin for abundant riverine nanoscale ice-nucleating particles. Environ. Sci. Technol. 2018. submitted. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| River | Location | Date | Lat. | Long. | Water Temp. (°C) | pH | Onset of Freezing (°C) | INPs Active at −7 °C (mL−1) | INPs Active at −10 °C (mL−1) | INPs Heat Labile at −10 °C (%) |
|---|---|---|---|---|---|---|---|---|---|---|
| Sweetwater | Sweetwater Camp Site | 8/15/2012 | 42.564837 | −109.061902 | 11.0 | 6.8 | −5 | 43 | 690 | 81 |
| Sweetwater | Sweetwater Bridge | 8/15/2012 | 42.541721 | −108.18018 | 17.0 | 7.7 | −6 | 180 | 1600 | 99 |
| Sweetwater | Ore Road | 8/15/2012 | 42.524983 | −107.798983 | 21.0 | 8.0 | −6 | 130 | 1200 | 66 |
| Sweetwater | Mormon Bridge | 8/14/2012 | 42.442715 | −107.221725 | - | - | −5 | 420 | 2100 | 78 |
| North Platte | Miles Landing | 8/16/2012 | 42.638502 | −106.619033 | 17.0 | 7.3 | −6 | >69 | 130 | 67 |
| North Platte | Bessemer Bend Rd | 8/16/2012 | 42.771948 | −106.530563 | 17.0 | 7.7 | −5 | >69 | 87 | 89 |
| North Platte | Pacificorp, Casper | 8/16/2012 | 42.865464 | −106.297231 | 19.0 | 8.4 | −4 | 87 | 540 | 76 |
| North Platte | Glendo Dam | 8/17/2012 | 42.478084 | −104.951656 | 17.0 | 7.3 | −4 | 370 | 1100 | 66 |
| North Platte | Guernsey | 8/17/2012 | 42.259881 | −104.74157 | 20.0 | 8.6 | −5 | 370 | 1500 | 75 |
| North Platte | SAREC, Lingle | 8/23/2012 | 42.125186 | −104.378219 | 19.0 | 7.6 | −5 | 340 | 1400 | 87 |
| North Platte | Chimney Rock | 8/24/2012 | 41.729665 | −103.324214 | 18.0 | 7.5 | −5 | 410 | 1800 | 95 |
| North Platte | Lewellen | 8/24/2012 | 41.316284 | −102.126333 | 21.0 | 7.5 | −5 | 130 | 980 | 91 |
| North Platte | Lewellen | 6/14/2015 | 41.316284 | −102.126333 | 21.5 | 8.0 | −4 | 1500 | 2600 | 69 |
| Platte | Cozad | 6/14/2015 | 40.837433 | −99.98696 | 22.5 | 8.0 | −5 | 1500 | 4700 | 99 |
| Platte | Hastings | 6/14/2015 | 40.77126 | −98.492202 | 24.0 | 8.1 | −5 | 1500 | 1700 | 94 |
| Platte | North Bend | 6/14/2015 | 41.452678 | −96.776912 | 24.5 | 8.3 | −5 | 1200 | 4700 | 97 |
| Platte | La Platte | 6/15/2015 | 41.058924 | −95.928276 | - | - | −5 | 4700 | 15,000 | 88 |
| Missouri | Three Forks/Headwaters St. Pk. | 6/28/2015 | 45.926835 | −111.50578 | 22.1 | 8.6 | −6 | 47 | 590 | - |
| Missouri | Wolf Creek Br./Holter Dam | 6/27/2015 | 47.020074 | −112.011922 | 17.3 | 9.0 | −6 | 36 | 170 | - |
| Missouri | Old Fort Benton | 6/27/2015 | 47.820359 | −110.663314 | 24.3 | 8.9 | −4 | 350 | 590 | - |
| Missouri | James Kipp Recreation Area | 6/27/2015 | 47.627895 | −108.697475 | 27.5 | 8.8 | −5 | 81 | 590 | - |
| Missouri | Fort Peck | 6/26/2015 | 48.039842 | −106.290658 | 12.9 | 8.3 | −5 | 870 | 1500 | - |
| Missouri | Williston | 6/26/2015 | 48.110958 | −103.717126 | 21.5 | 8.1 | −4 | 2600 | 10,000 | - |
| Missouri | Washburn | 6/26/2015 | 47.288804 | −101.042415 | 11.0 | 8.4 | −5 | 470 | 2100 | - |
| Missouri | Mobridge | 6/25/2015 | 45.512826 | −100.382602 | 24.2 | 7.3 | −5 | 170 | 360 | - |
| Missouri | Bad River near Fort Pierre | 6/25/2015 | 44.353082 | −100.369376 | 23.2 | 7.8 | −5 | 17,000 | 59,000 | - |
| Missouri | Chamberlain | 6/25/2015 | 43.815016 | −99.326346 | 21.3 | 8.5 | −5 | 170 | 470 | - |
| Missouri | Blair | 6/14/2015 | 41.548381 | −96.097105 | 22.3 | 8.4 | −4 | 1500 | 3000 | 90 |
| Missouri | Plattsmouth | 6/15/2015 | 41.000821 | −95.866735 | 23.2 | 7.9 | −4 | 3500 | 12,000 | 95 |
| Missouri | Rulo | 6/15/2015 | 40.049925 | −95.421274 | 23.0 | 7.9 | −5 | 4700 | 15,000 | 93 |
| Missouri | Leavenworth | 6/15/2015 | 39.330756 | −94.910311 | 23.5 | 7.7 | −4 | 7200 | 26,000 | 95 |
| Missouri | Missouri City | 6/15/2015 | 39.236795 | −94.29664 | 22.7 | 7.7 | −4 | 12,000 | 47,000 | 95 |
| Missouri | Mokane | 6/17/2015 | 38.670188 | −91.874438 | 20.8 | 7.9 | −5 | 1000 | 3600 | 90 |
| Mississippi | Bemidji | 6/23/2015 | 47.483244 | −94.727823 | 20.3 | 8.0 | −6 | 1000 | 1700 | 99 |
| Mississippi | Brainerd | 6/23/2015 | 46.34847 | −94.20721 | 22.7 | 7.9 | −5 | 590 | 5900 | 99 |
| Mississippi | Monticello | 6/22/2015 | 45.294468 | −93.757178 | 25.2 | 8.2 | −5 | 170 | 1700 | 97 |
| Mississippi | Red Wing | 6/22/2015 | 44.562502 | −92.461088 | 23.9 | 8.4 | −5 | 1500 | 3600 | 96 |
| Mississippi | Pettibone Pk, La Crosse | 6/22/2015 | 43.812197 | −91.260732 | 24.7 | 8.4 | −6 | 290 | 1700 | 95 |
| Mississippi | Dubuque | 6/22/2015 | 42.495042 | −90.65702 | 23.2 | 8.1 | −6 | 470 | 2100 | 93 |
| Mississippi | Muscatine | 6/21/2015 | 41.419779 | −91.043757 | 24.4 | 7.9 | −5 | 870 | 3500 | 93 |
| Mississippi | Canton | 6/21/2015 | 40.139674 | −91.515404 | 23.9 | 8.0 | −5 | 1700 | 4700 | 90 |
| Mississippi | Dam 25, Winfield | 6/21/2015 | 39.0054 | −90.691467 | 24.0 | 8.0 | −5 | 2600 | 8700 | 99 |
| Mississippi | Herculaneum | 6/17/2015 | 38.258919 | −90.376007 | 22.8 | 7.7 | −4 | 2600 | 12,000 | 99 |
| Mississippi | Cape Girardeau | 6/17/2015 | 37.31516 | −89.512691 | 25.8 | 7.7 | −5 | 2600 | 12,000 | 97 |
| Mississippi | Caruthersville | 6/17/2015 | 36.232633 | −89.702174 | 28.0 | 8.6 | −5 | 210 | 720 | 76 |
| Mississippi | Mud Island, Memphis | 6/18/2015 | 35.18217 | −90.057939 | 28.6 | 7.4 | −5 | 150 | 470 | 92 |
| Mississippi | Rosedale | 6/18/2015 | 33.827979 | −91.053996 | 27.1 | 7.8 | −6 | 87 | 170 | 90 |
| Mississippi | Mayersville | 6/18/2015 | 32.917046 | −91.061286 | 26.8 | 7.7 | −5 | 1700 | 8700 | 96 |
| Mississippi | Natchez | 6/18/2015 | 31.558579 | −91.413607 | 26.6 | 7.7 | −5 | 1500 | 4700 | 93 |
| Samples | INPs < 0.22 µm Active at −7 °C (%) | INPs < 0.22 µm Active at −10 °C (%) |
|---|---|---|
| Sweetwater (Campsite, Bridge, Ore Road) | 100 | 97 |
| North Platte (Glendo Dam, Guernsey) | 31 | 32 |
| North Platte (SAREC, Lingle) | 44 | 24 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moffett, B.F.; Hill, T.C.J.; DeMott, P.J. Abundance of Biological Ice Nucleating Particles in the Mississippi and Its Major Tributaries. Atmosphere 2018, 9, 307. https://doi.org/10.3390/atmos9080307
Moffett BF, Hill TCJ, DeMott PJ. Abundance of Biological Ice Nucleating Particles in the Mississippi and Its Major Tributaries. Atmosphere. 2018; 9(8):307. https://doi.org/10.3390/atmos9080307
Chicago/Turabian StyleMoffett, Bruce F., Tom C. J. Hill, and Paul J. DeMott. 2018. "Abundance of Biological Ice Nucleating Particles in the Mississippi and Its Major Tributaries" Atmosphere 9, no. 8: 307. https://doi.org/10.3390/atmos9080307