Effects of Reclaimed Water Irrigation on Microbial Diversity and Composition of Soil with Reducing Nitrogen Fertilization

 ,
, 

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Water and Soil Analyses
2.3. DNA Extraction and PCR Amplification
2.4. The Plant Biomass and Yield of Tomato
2.5. Statistical Analysis
3. Results
3.1. Changes in Soil Chemical Properties under RW Irrigation
3.2. Soil Bacterial Community Richness and Diversity under RW Irrigation
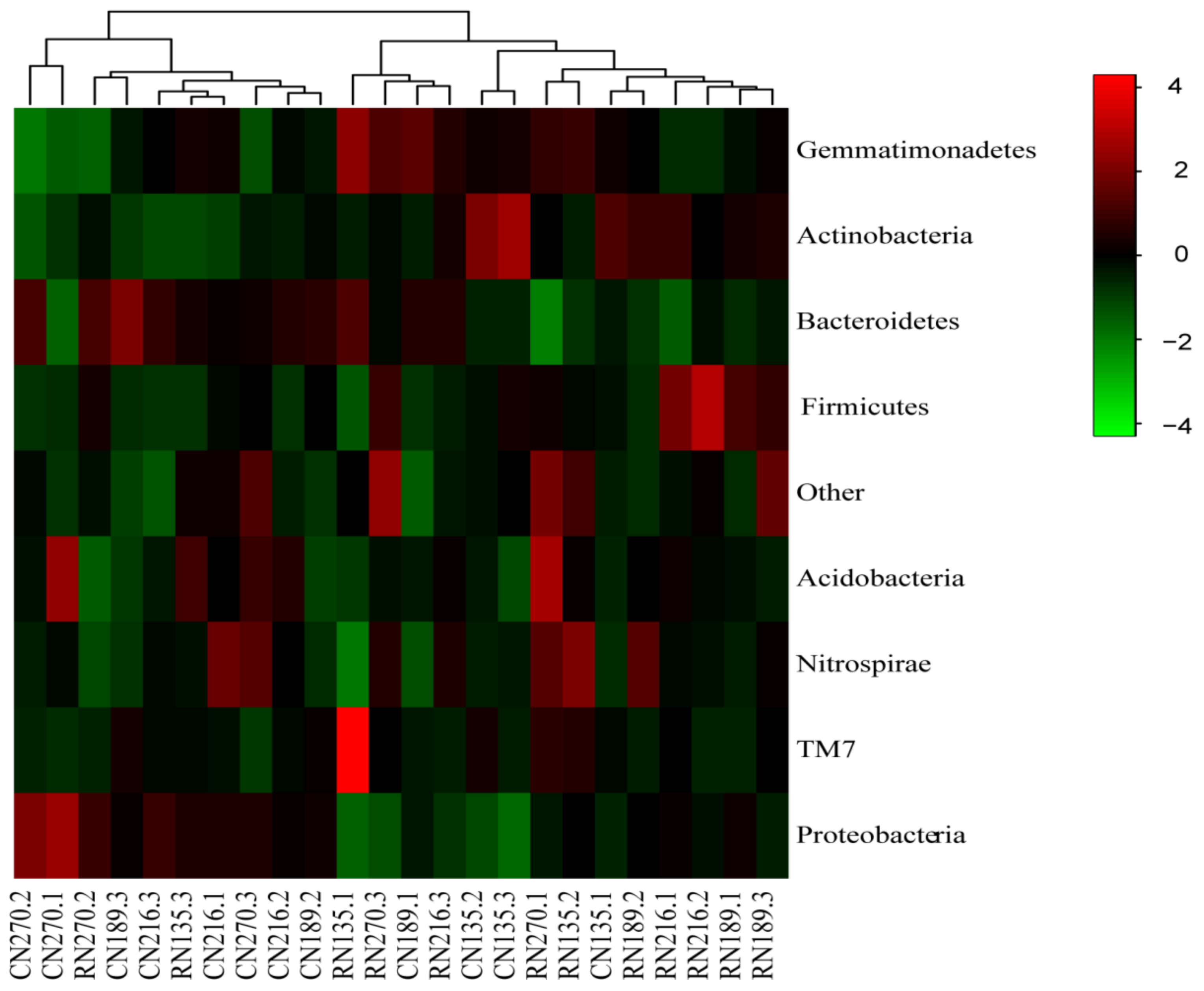
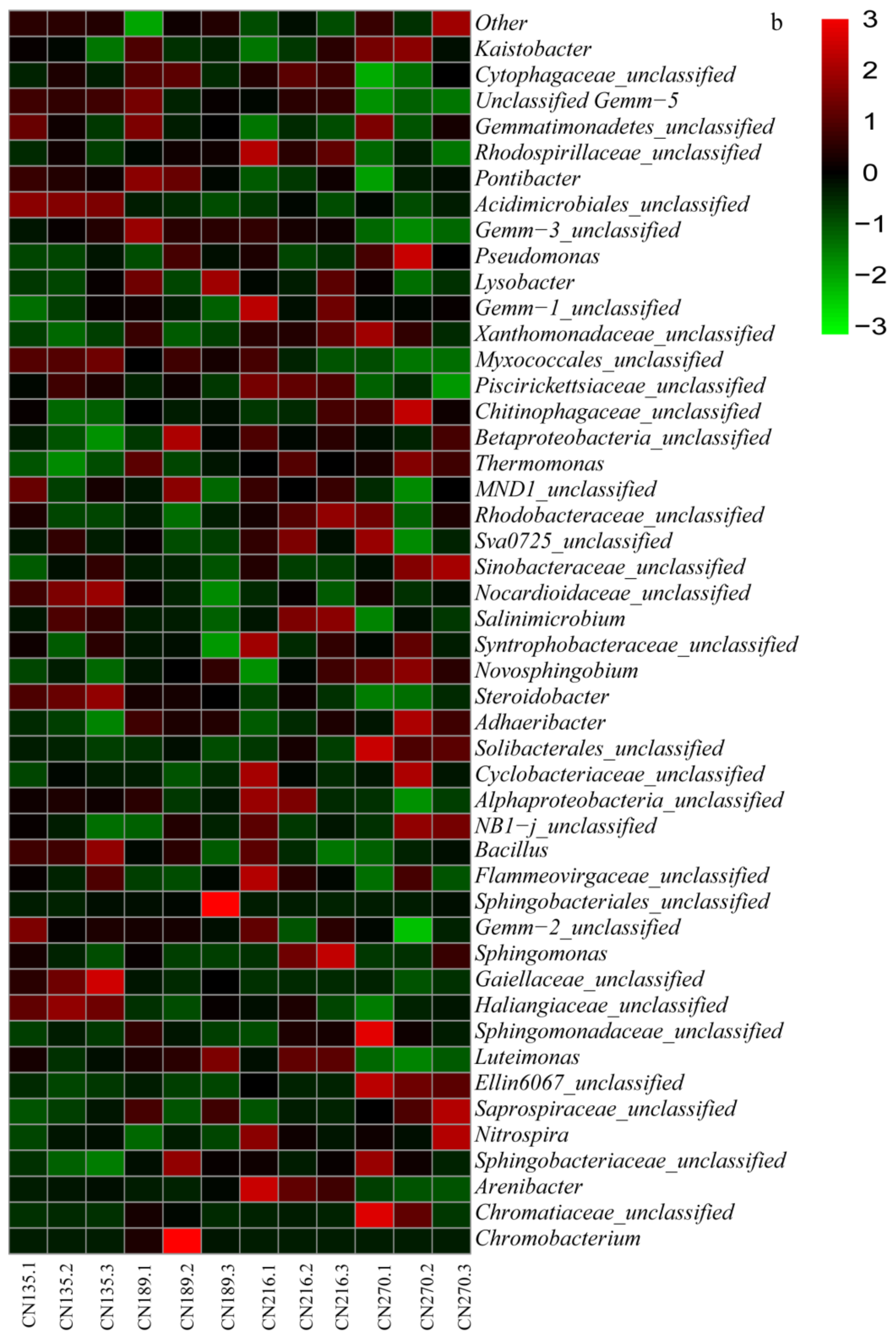
3.3. Taxonomic Evaluation of Bacteria under RW Irrigation by HTS
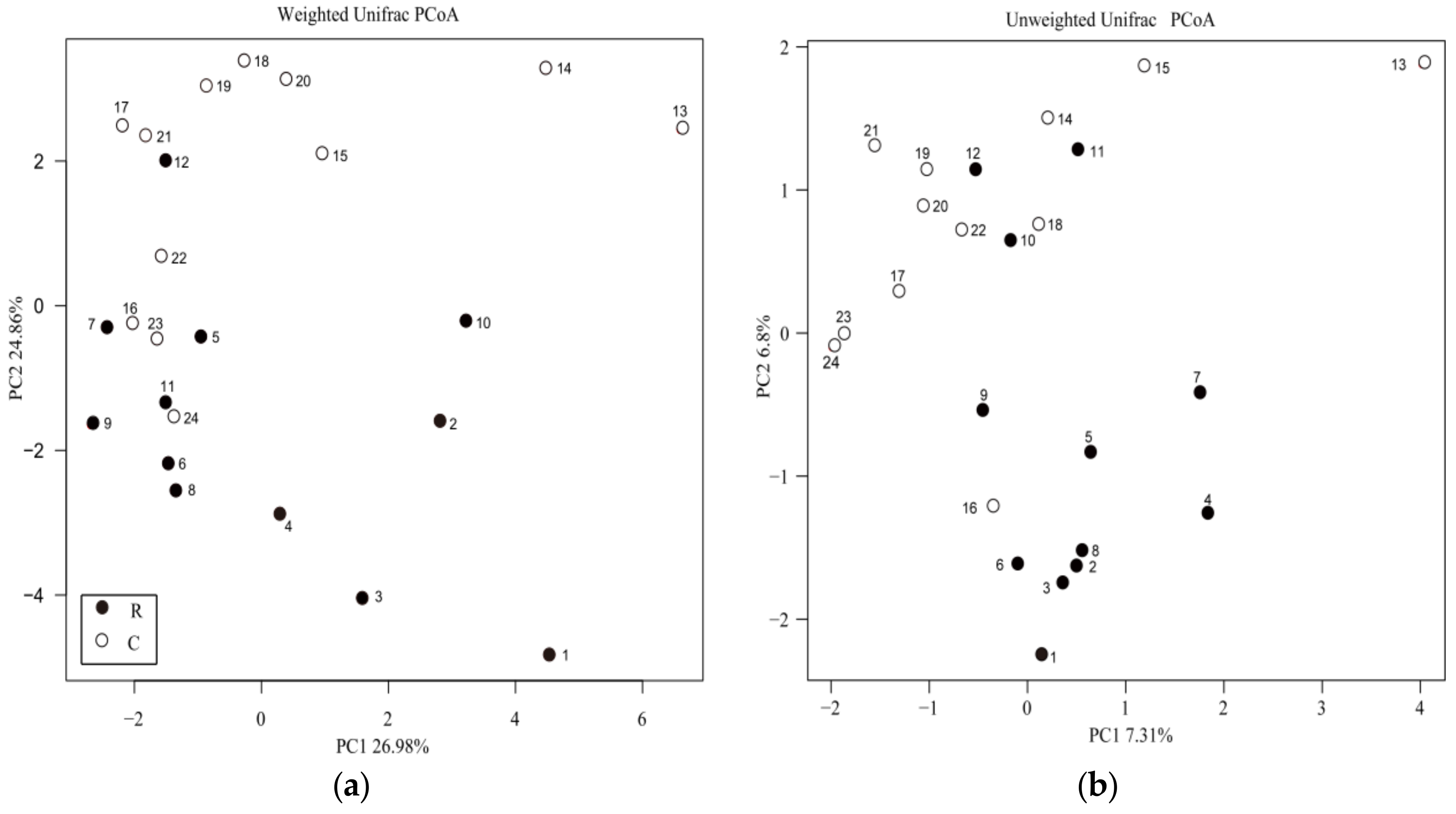
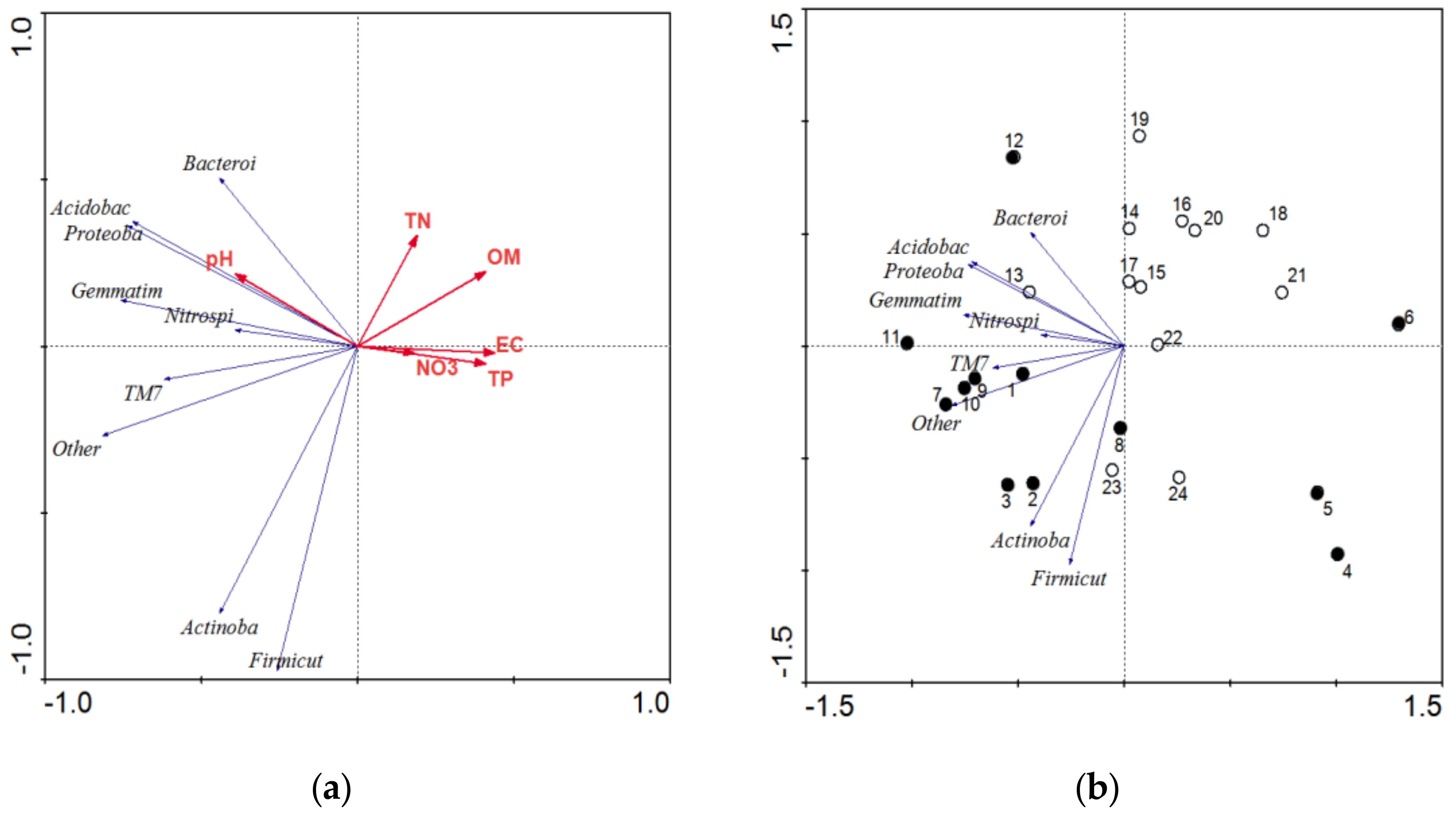
3.4. Phylogenetic Analysis of Soil Bacterial Community Composition and Structure under RW Irrigation
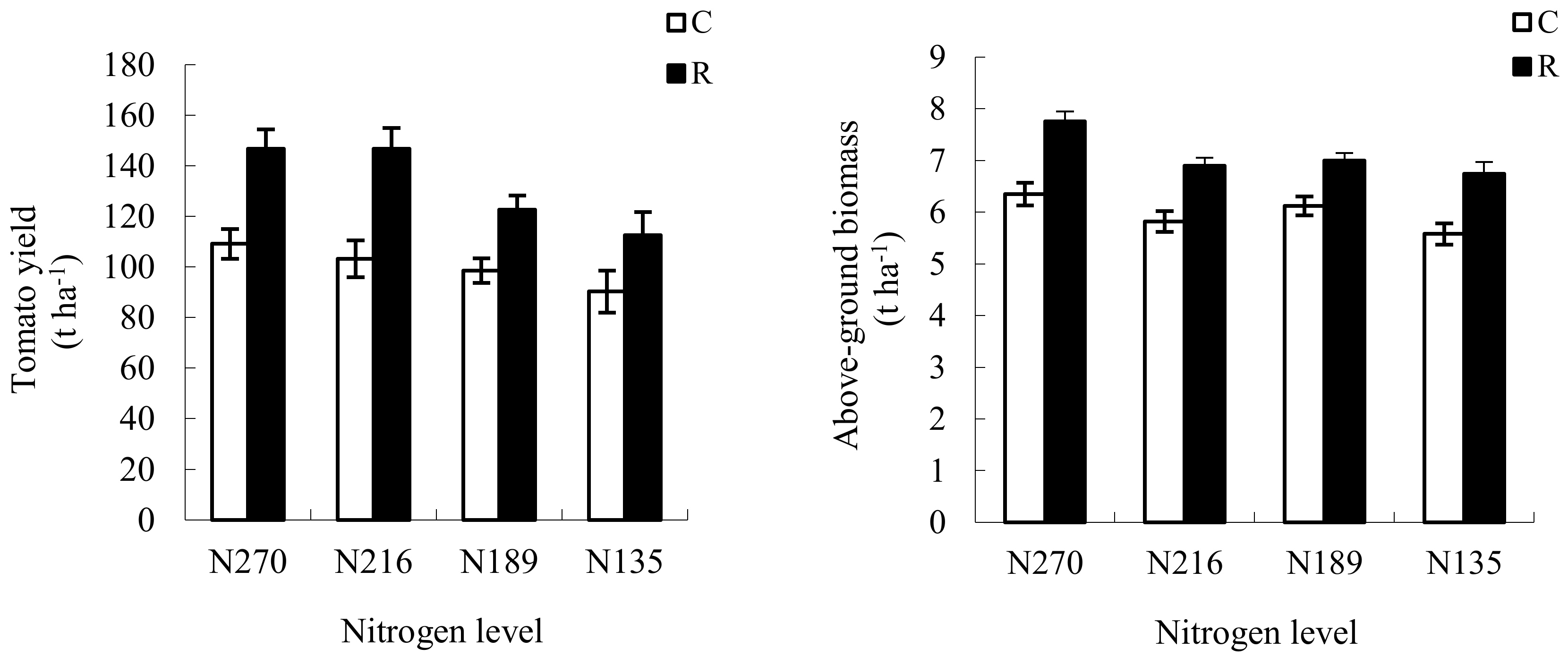
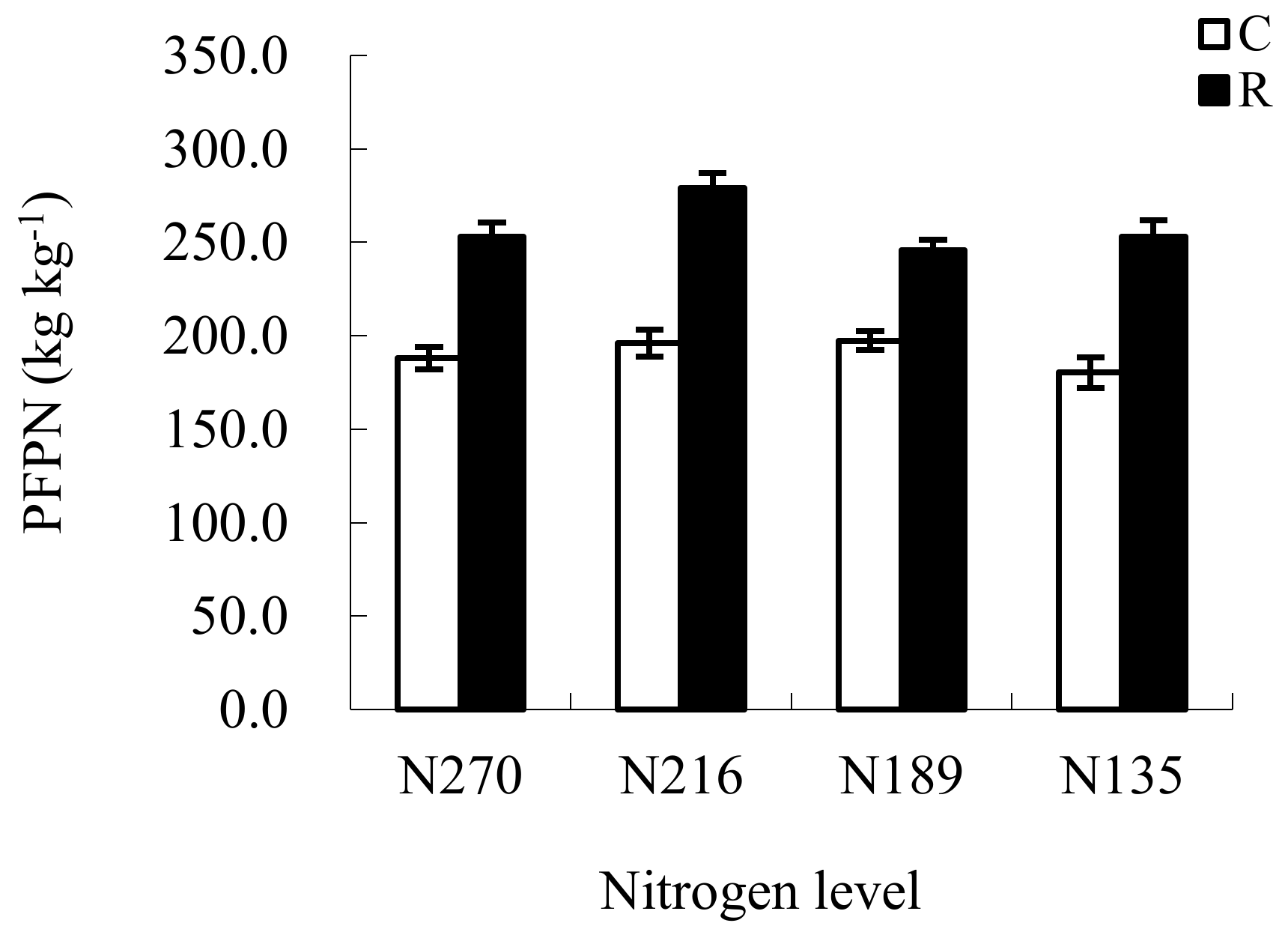
3.5. Changes in Tomato Yield and Partial Factor Productivities from the Applied N (PFPN) under RW Irrigation
4. Discussion
4.1. Changes in Soil Chemical Properties under RW Irrigation
4.2. Effects of RW Irrigation on Soil Bacterial Diversity
4.3. Effects of RW Irrigation on Soil Bacterial Community Strcture and Composition
4.4. Effects of RW Irrigation on Tomato Yield and PFPN
5. Conclusions
Supplementary Materials
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Daquiado, A.R.; Kuppusamy, S.; Kim, S.Y.; Kim, J.H.; Yoon, Y.E.; Kim, P.J.; Oh, S.H.; Kwak, Y.S.; Lee, Y.B. Pyrosequencing analysis of bacterial community diversity in long-term fertilized paddy field soil. Appl. Soil Ecol. 2016, 108, 84–91. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Srivastava, D.S.; Duffy, J.E.; Wright, J.P.; Downing, A.L.; Sankaran, M.; Jouseau, C. Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 2006, 443, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.L.; Paixão, S.M.; Caçador, I.; Carolino, M. CLPP and EEA profiles of microbial communities in salt marsh sediments. J. Soil Sediments 2007, 7, 418–425. [Google Scholar] [CrossRef]
- Frenk, S.; Hadar, Y.; Minz, D. Resilience of soil bacterial community to irrigation with water of different qualities under Mediterranean climate. Environ. Microbiol. 2014, 16, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Angin, I.; Yaganoglu, A.V.; Turan, M. Effects of long-term wastewater irrigation on soil properties. J. Sustain. Agric. 2005, 26, 31–42. [Google Scholar] [CrossRef]
- Lubello, C.; Gori, R.; Nicese, F.P.; Ferrini, F. Municipal-treated wastewater reuse for plant nurseries irrigation. Water Res. 2004, 38, 2939–2947. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wu, L.S.; Chang, A.C.; Zhang, Y. Impact of long-term reclaimed wastewater irrigation on agricultural soils, A preliminary assessment. J. Hazard. Mater. 2010, 183, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Adrover, M.; Farrus, E.; Moya, G.; Vadell, J. Chemical properties and biological activity in soils of Mallorca following twenty years of treated wastewater irrigation. J. Environ. Manag. 2012, 95, S188–S192. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.L.; Mecham, B. Long-term effects of recycled wastewater irrigation on soil chemical properties on golf course fairways. Agron. J. 2005, 97, 717–721. [Google Scholar] [CrossRef]
- Bastida, F.; Torres, I.F.; Romero-Trigueros, C.; Baldrian, P.; Vetrovský, T.; Bayona, J.M.; Alarcón, J.J.; Hernández, T.; García, C.; Nicolás, E. Combined effects of reduced irrigation and water quality on the soil microbial community of a citrus orchard under semi-arid conditions. Soil Biol. Biochem. 2017, 104, 226–237. [Google Scholar] [CrossRef]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes. Glob. Chang. Biol. 2012, 18, 1918–1927. [Google Scholar] [CrossRef]
- Lassaletta, L.; Billen, G.; Grizzetti, B.; Anglade, J.; Garnier, J. 50 year trends in nitrogen use efficiency of world cropping systems, the relationship between yield and nitrogen input to cropland. Environ. Res. Lett. 2014, 9, 105011. [Google Scholar] [CrossRef]
- Shen, W.S.; Lin, X.G.; Gao, N.; Zhang, H.Y.; Yin, R.; Shi, W.M.; Duan, Z.Q. Land use intensification affects soil microbial populations, functional diversity and related suppressiveness of cucumber Fusarium wilt in China’s Yangtze River Delta. Plant Soil 2008, 306, 117–127. [Google Scholar] [CrossRef]
- Geisseler, D.; Scow, K.M. Long-term effects of mineral fertilizers on soil microorganisms—A review. Soil Biol. Biochem. 2014, 75, 54–63. [Google Scholar] [CrossRef]
- Ge, Y.; Zhang, J.B.; Zhang, L.M.; Yang, M.; He, J.Z. Long-term fertilization regimes affect bacterial community structure and diversity of an agricultural soil in Northern China. J. Soil Sediments 2008, 8, 43–50. [Google Scholar] [CrossRef]
- Shen, W.S.; Ni, Y.Y.; Gao, N.; Bian, B.Y.; Zheng, S.N.; Lin, X.G.; Chu, H.Y. Bacterial community composition is shaped by soil secondary salinization and acidification brought on by high nitrogen fertilization rates. Appl. Soil Ecol. 2016, 108, 76–83. [Google Scholar] [CrossRef]
- Huang, X.R.; Xiong, W.; Liu, W.; Guo, X.Y. Effect of reclaimed water effluent on bacterial community structure in the Typha angustifolia L. rhizosphere soil of urbanized riverside wetland, China. J. Environ. Sci. 2017, 55, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Greaver, T.L. A global perspective on belowground carbon dynamics under nitrogen enrichment. Ecol. Lett. 2010, 13, 819–828. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.D. Soil and Agro-Chemical Analysis Methods; China Agriculture Press: Beijing, China, 2000; pp. 25–38. (In Chinese) [Google Scholar]
- Liu, Z.; Lozupone, C.; Hamady, M.; Bushman, F.; Knight, R. Short pyrosequencing reads suffice for accurate microbial community analysis. Nucleic Acids Res. 2007, 35, e120. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, G.; Bates, S.; Eilers, K.; Lauber, C.; Caporaso, J.; Walters, W. The under-recognized dominance of Verrucomicrobia in soil bacterial communities. Soil Biol. Biochem. 2011, 43, 1450–1455. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur, open source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; Huttley, G.A.; Kelley, S.T.; Knights, D.; Koenig, J.E.; Ley, R.E.; Lozupone, C.A.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Hamady, M.; Lozupone, C.; Knight, R. Fast UniFrac, Facilitating high-throughput phylogenetic analyses of microbial communities including analysis of pyrosequencing and PhyloChip data. ISME J. 2010, 4, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Lauber, C.L.; Ramirez, K.S.; Zaneveld, J.; Bradford, M.A.; Knight, R. Comparative metagenomic, phylogenetic and physiological analyses of soil microbial communities across nitrogen gradients. ISME J. 2012, 6, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Hadidi, M.E.; Ruscheweyh, H.J.; Huson, D.H. Improved metagenome analysis using MEGAN5. In Joint, 21st Annual International Conference on ISMB and 12th ECCB; ISCB: Bethesda, MD, USA, 2013; Volume 4. [Google Scholar]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [PubMed]
- Aldrete-Tapia, A.; Escobar-Ramírez, M.C.; Tamplin, M.L.; Hernández-Iturriaga, M. High-throughput sequencing of microbial communities in Poro cheese, an artisanal Mexican cheese. Food Microbiol. 2014, 44, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, M.; Fegatella, F.; Wasinger, V.; Guihaus, M.; Corthals, L.G.; Cavicchioli, R. Cross-species identification of proteins from proteome profiles of the marine oligotrophic ultramicrobacterium, Sphingopyxis alaskensis. Proteomics 2004, 4, 1779–1788. [Google Scholar] [CrossRef] [PubMed]
- García-Orenes, F.; Caravaca, F.; Morugán-Coronado, A.; Roldán, A. Prolonged irrigation with municipal wastewater promotes a persistent and active soil microbial community in a semiarid agroecosystem. Agric. Water Manag. 2015, 149, 115–122. [Google Scholar] [CrossRef]
- Gelsomino, A.; Badalucco, L.; Ambrosoli, R.; Crecchio, C.; Puglisi, E.; Meli, S.M. Changes in chemical and biological soil properties as induced by anthropogenic disturbance, a case study of an agricultural soil under recurrent flooding by wastewaters. Soil Biol. Biochem. 2006, 38, 2069–2080. [Google Scholar] [CrossRef]
- Bach, E.M.; Baer, S.G.; Meyer, C.K.; Six, J. Soil texture affects soil microbial and structural recovery during grassland restoration. Soil Biol. Biochem. 2010, 42, 2182–2191. [Google Scholar] [CrossRef]
- Xun, W.B.; Xu, Z.H.; Wei, L.; Ren, Y.; Huang, T.; Ran, W.; Wang, B.R.; Shen, Q.R.; Zhang, R.F. Long-term organic-inorganic fertilization ensures great soil productivity and bacterial diversity after natural-to-agricultural ecosystem conversion. J. Microbiol. 2016, 54, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Sylvia, D.M.; Fuhrmann, J.J.; Hartel, P.; Zuberer, D.A. Principles and applications of soil microbiology. New Age Int. 1998, 34, 11–68. [Google Scholar]
- Waldrop, M.P.; Balser, T.C.; Firestone, M.K. Linking microbial composition to function in a tropical soil. Soil Biol. Biochem. 2000, 32, 1837–1846. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity, a synthesis. Ecol. Appl. 2010, 20, 30–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, J.; Liu, X.J.; Song, L.; Lin, X.G.; Zhang, H.Z.; Shen, C.C.; Chu, H.Y. Nitrogen fertilization directly affects soil bacterial diversity and indirectly affects bacterial community composition. Soil Biol. Biochem. 2016, 92, 41–49. [Google Scholar] [CrossRef]
- Becerra-Castro, C.; Lopes, A.R.; Vaz-Moreira, I.; Silva, E.F.; Manaia, C.M.; Nunes, O.C. Wastewater reuse in irrigation, a microbiological perspective on implications in soil fertility and human and environmental health. Environ. Int. 2015, 75, 117–135. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.H.; Gong, H.L.; Guo, X.Y. Rhizosphere bacterial community of Typha angustifolia L. and water quality in a river wetland supplied with reclaimed water. Appl. Microbiol. Biotechnol. 2015, 99, 2883–2893. [Google Scholar] [CrossRef] [PubMed]
- Häni, H.; Siegenthaler, A.; Candinas, T. Soil effects due to sewage sludge application in agriculture. Fertil. Res. 1995, 43, 149–156. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.A.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority, soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Sui, Y.Y.; Yu, Z.H.; Yu, S.; Chu, H.Y.; Jin, J.; Liu, X.B.; Wang, G.H. High throughput sequencing analysis of biogeographical distribution of bacterial communities in the black soils of northeast China. Soil Biol. Biochem. 2014, 70, 113–122. [Google Scholar] [CrossRef]
- Guo, W.; Andersen, M.N.; Qi, X.B.; Li, P.; Li, Z.Y.; Fan, X.Y.; Zhou, Y. Effects of reclaimed water irrigation and nitrogen fertilization on the chemical properties and microbial community of soil. J. Integr. Agric. 2017, 16, 679–690. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.S.; Shen, F.T. Bioprospecting of facultatively oligotrophic bacteria from non-rhizospheric soils. Appl. Soil Ecol. 2016, 108, 315–324. [Google Scholar] [CrossRef]
- Tada, Y.; Kobata, T.; Nakaoka, C. A simple and easy method for the monitoring of environmental pollutants using oligotrophic bacteria. Lett. Appl. Microbiol. 2001, 32, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, K.S.; Lauber, C.L.; Knight, R.; Bradford, M.A.; Fierer, N. Consistent effects of nitrogen fertilization on soil bacterial communities in contrasting systems. Ecology 2010, 91, 3463–3470. [Google Scholar] [CrossRef] [PubMed]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cooper, J.M.; Lin, Z.A.; Li, Y.; Yang, X.; Zhao, B. Soil microbial community structure and function are significantly affected by long-term organic and mineral fertilization regimes in the North China Plain. Appl. Soil Ecol. 2015, 96, 75–87. [Google Scholar] [CrossRef]
- Truu, M.; Truu, J.; Heinsoo, K. Changes in soil microbial community under willow coppice: The effect of irrigation with secondary-treated municipal wastewater. Ecol. Eng. 2009, 35, 1011–2010. [Google Scholar] [CrossRef]
- Zhu, J.H.; Li, X.L.; Zhang, F.S.; Li, J.L.; Christie, P. Response of greenhouse tomato and pepper yields and nitrogen dynamics to applied compound fertilizers. Pedosphere 2004, 14, 213–222. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | NO3−-N | NH4+-N | TN | TP | Cu | Cd | Cr |
| (mg L−1) | (mg L−1) | (mg L−1) | (mg L−1) | (mg L−1) | (µg L−1) | (µg L−1) | |
| CW | 1.7 | 0.86 | 3.9 | 2.88 | 0.005 | 0.68 | 6.38 |
| RW | 20.62 | 11.11 | 45.14 | 2.94 | 0.02 | 3.33 | 20.08 |
| RMI | - | - | - | - | 1.0 | 10.0 | 100 |
| Index | CODMn | pH | EC | K | Na | Ca | Mg |
| (mg L−1) | (ds m−1) | (mg L−1) | (mg L−1) | (mg L−1) | (mg L−1) | ||
| CW | 7.86 | 7.52 | 0.63 | 0.88 | 53.8 | 10.21 | 11.0 |
| RW | 13.37 | 7.4 | 1.7 | 5.81 | 178.6 | 41.73 | 35.7 |
| RMI | 60 | 5.5–8.5 | 1–2 | - | - | - | - |
| Treatments 1) | TNg kg−1 | TPg kg−1 | OM 2) g kg−1 | NO3−-N mg kg−1 | EC μs cm−1 | pH 3) | C/N 4) |
|---|---|---|---|---|---|---|---|
| CN270 | 1.48 ± 0.21a | 1.88 ± 0.08a | 33.32 ± 1.97a | 55.04 ± 3.94c | 812.00 ± 22.91cd | 8.63 ± 0.03b | 13.06 |
| RN270 | 0.97 ± 0.06c | 1.65 ± 0.06b | 24.45 ± 0.83b | 109.77 ± 10.08a | 978.33 ± 67.88b | 8.41 ± 0.02d | 14.62 |
| CN216 | 1.30 ± 0.03ab | 1.80 ± 0.03a | 33.04 ± 2.67a | 73.62 ± 12.28b | 970.00 ± 38.12b | 8.55 ± 0.03c | 14.74 |
| RN216 | 1.15 ± 0.05bc | 1.92 ± 0.13a | 32.52 ± 3.10a | 78.63 ± 1.96b | 1061.00 ± 26.21a | 8.33 ± 0.02e | 16.40 |
| CN189 | 1.45 ± 0.26a | 1.87 ± 0.02a | 33.23 ± 1.35a | 83.46 ± 6.78b | 711.67 ± 57.73de | 8.66 ± 0.01c | 13.29 |
| RN189 | 0.95 ± 0.01c | 1.64 ± 0.12a | 29.94 ± 3.35a | 97.13 ± 0.85b | 805.00 ± 32.19cd | 8.54 ± 0.02b | 18.28 |
| CN135 | 1.21 ± 0.01bc | 1.93 ± 0.10a | 32.65 ± 0.62a | 52.05 ± 0.14c | 706.00 ± 34.39de | 8.74 ± 0.05a | 15.65 |
| RN135 | 1.08 ± 0.12bc | 1.63 ± 0.01b | 28.86 ± 3.00a | 60.73 ± 0.06c | 746.00 ± 34.39de | 8.57 ± 0.01c | 15.50 |
| Significance based on two-way analysis of variance (ANOVA) (F value) | |||||||
| W (water) | 36.44 ** | 22.47 ** | 18.58 ** | 3.76 | 12.23 ** | 4.29 | |
| N (nitrogen) | 0.542 | 1.91 | 2.94 | 43.14 ** | 71.67 ** | 25.98 ** | |
| W × N | 4.02 * | 8.07 ** | 3.33 * | 73.22 ** | 88.99 ** | 34.20 ** | |
| Treatments | OTUs | ACE | Chao | Shannon(H) | Simpson | Coverage |
|---|---|---|---|---|---|---|
| CN270 | 2544b | 3157.07b | 3402.55b | 6.84b | 0.002414ab | 0.9820a |
| RN270 | 2894ab | 3724.00ab | 3995.85ab | 6.99ab | 0.001985ab | 0.9744ab |
| CN216 | 2971ab | 3820.15ab | 4099.65ab | 6.99ab | 0.002006ab | 0.9665b |
| RN216 | 2726ab | 3561.71ab | 3754.39ab | 6.94ab | 0.002036ab | 0.9681b |
| CN189 | 3067ab | 3886.37ab | 4162.58ab | 6.97ab | 0.002641a | 0.9688b |
| RN189 | 2881ab | 3816.72ab | 4085.08ab | 6.98ab | 0.001888ab | 0.9682b |
| CN135 | 3387a | 4294.39a | 4605.83a | 7.10a | 0.001777b | 0.9724ab |
| RN135 | 3057ab | 3995.39ab | 4295.40ab | 6.99ab | 0.002077ab | 0.9748ab |
| Significance based on two-way analysis of variance (ANOVA) (F value) | ||||||
| W (water) | 0.005 | 0.023 | 0.000 | 1.679 | 0.183 | |
| N (nitrogen) | 1.859 | 1.891 | 2.274 | 0.901 | 4.164 * | |
| W × N | 0.856 | 0.884 | 2.574 | 2.038 | 0.851 | |
| OTU | ACE | Chao | Shannon | Simpson | Coverage | EC (ds/m) | pH | NO3—-N (mg/kg) | OM (g/kg) | TP (g/kg) | TN (g/kg) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OTU | 1.00 | |||||||||||
| ACE | 0.985 ** | 1.00 | ||||||||||
| Chao | 0.980 ** | 0.991 ** | 1.00 | |||||||||
| Shannon | 0.803 ** | 0.833 ** | 0.834 ** | 1.00 | ||||||||
| Simpson | −0.144 | −0.225 | −0.214 | −0.585 ** | 1.00 | |||||||
| Coverage | −0.224 | −0.300 | −0.316 | −0.406 * | 0.215 | 1.00 | ||||||
| EC (ds/m) | −0.024 | −0.055 | −0.081 | 0.031 | 0.245 | −0.266 | 1.00 | |||||
| pH | 0.120 | 0.115 | 0.146 | 0.033 | −0.011 | 0.232 | −0.814 ** | 1.00 | ||||
| NO3−-N (mg/kg) | −0.017 | −0.054 | −0.062 | 0.009 | 0.125 | −0.071 | 0.820 * | −0.719 ** | 1.00 | |||
| OM (g/kg) | 0.112 | 0.095 | 0.098 | 0.013 | 0.173 | −0.350 | 0.115 | 0.064 | −0.235 | 1.00 | ||
| TP (g/kg) | −0.005 | −0.089 | −0.08 | −0.076 | 0.207 | −0.098 | 0.443 * | −0.323 | 0.151 | 0.552 ** | 1.00 | |
| TN (g/kg) | −0.097 | −0.204 | −0.171 | −0.275 | 0.323 | 0.177 | 0.193 | 0.087 | 0.111 | 0.554 ** | 0.572 ** | 1.00 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, W.; Qi, X.; Xiao, Y.; Li, P.; Andersen, M.N.; Zhang, Y.; Zhao, Z. Effects of Reclaimed Water Irrigation on Microbial Diversity and Composition of Soil with Reducing Nitrogen Fertilization. Water 2018, 10, 365. https://doi.org/10.3390/w10040365
Guo W, Qi X, Xiao Y, Li P, Andersen MN, Zhang Y, Zhao Z. Effects of Reclaimed Water Irrigation on Microbial Diversity and Composition of Soil with Reducing Nitrogen Fertilization. Water. 2018; 10(4):365. https://doi.org/10.3390/w10040365
Chicago/Turabian StyleGuo, Wei, Xuebin Qi, Yatao Xiao, Ping Li, Mathias Neumann Andersen, Yan Zhang, and Zhijuan Zhao. 2018. "Effects of Reclaimed Water Irrigation on Microbial Diversity and Composition of Soil with Reducing Nitrogen Fertilization" Water 10, no. 4: 365. https://doi.org/10.3390/w10040365
APA StyleGuo, W., Qi, X., Xiao, Y., Li, P., Andersen, M. N., Zhang, Y., & Zhao, Z. (2018). Effects of Reclaimed Water Irrigation on Microbial Diversity and Composition of Soil with Reducing Nitrogen Fertilization. Water, 10(4), 365. https://doi.org/10.3390/w10040365