Spatial Variation in Trophic Structure of Dominant Fish Species in Lake Dongting, China during Dry Season

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
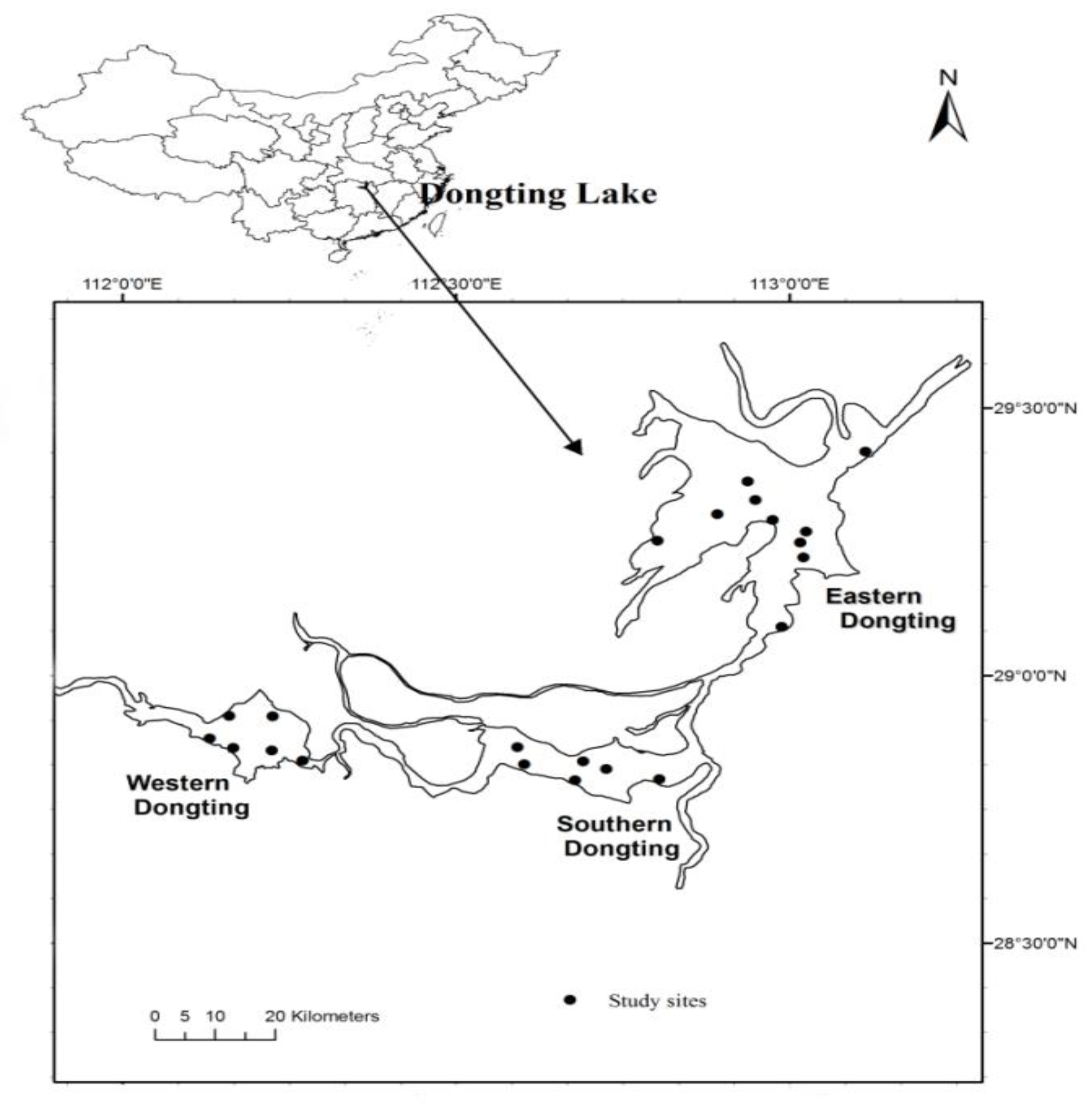
2.1. Study Area
2.2. Sample Processing and Treatment
2.3. Stable Isotope Analyses
2.4. Modeling Trophic Levels
2.5. Bayesian Mixing Model
2.6. Statistical Analyses
3. Results
3.1. Limnological Parameters and Fish Community Structure
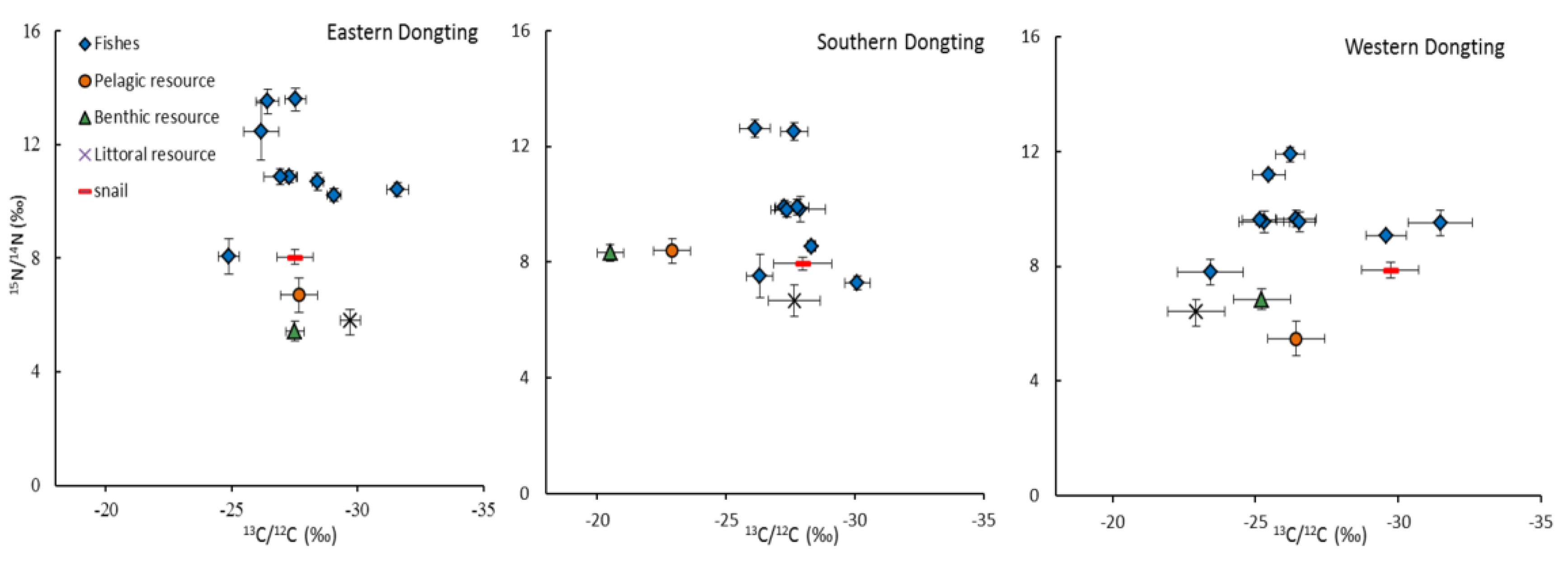
3.2. Isotopic Values
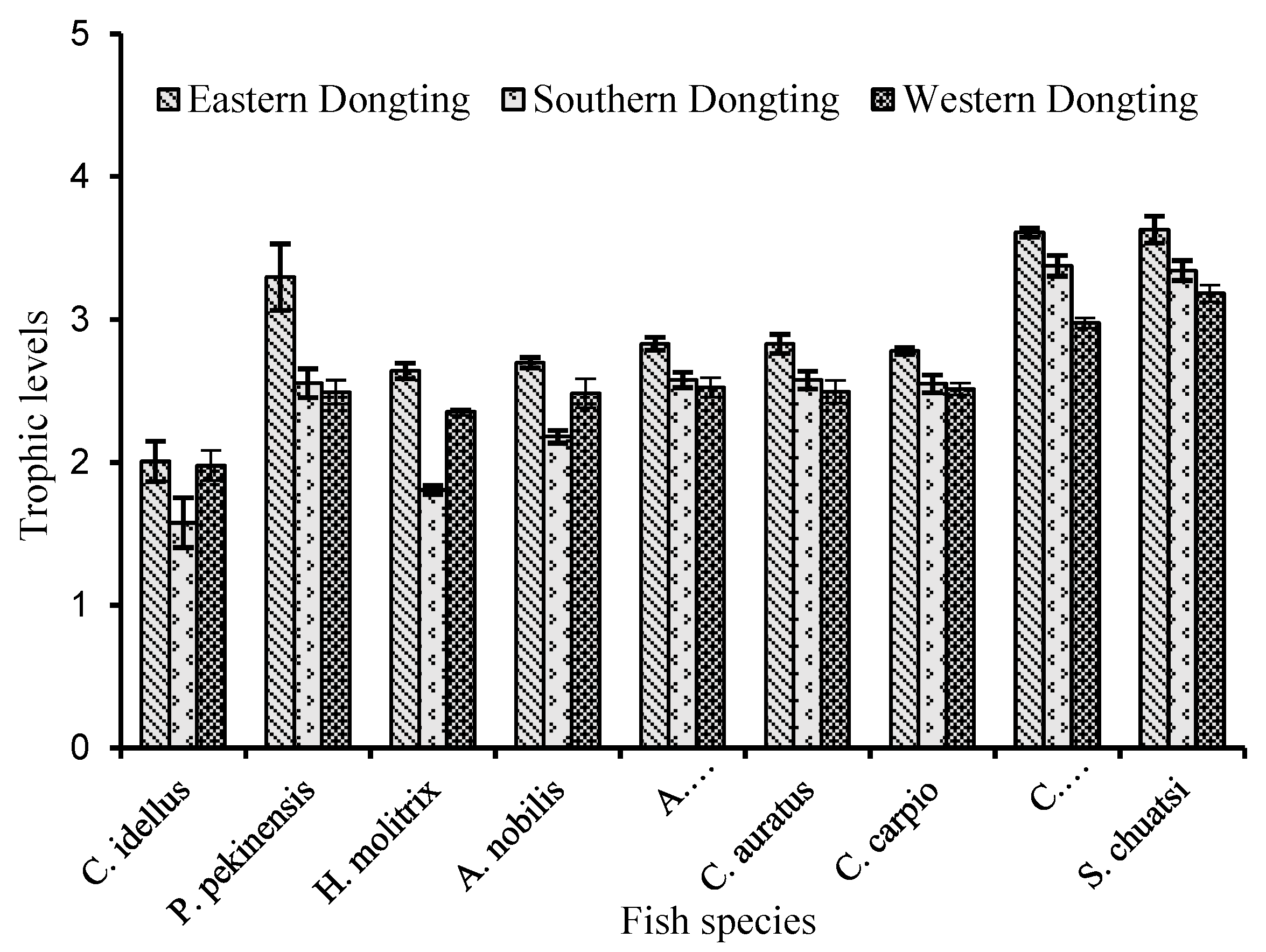
3.3. Trophic Levels of Fishes
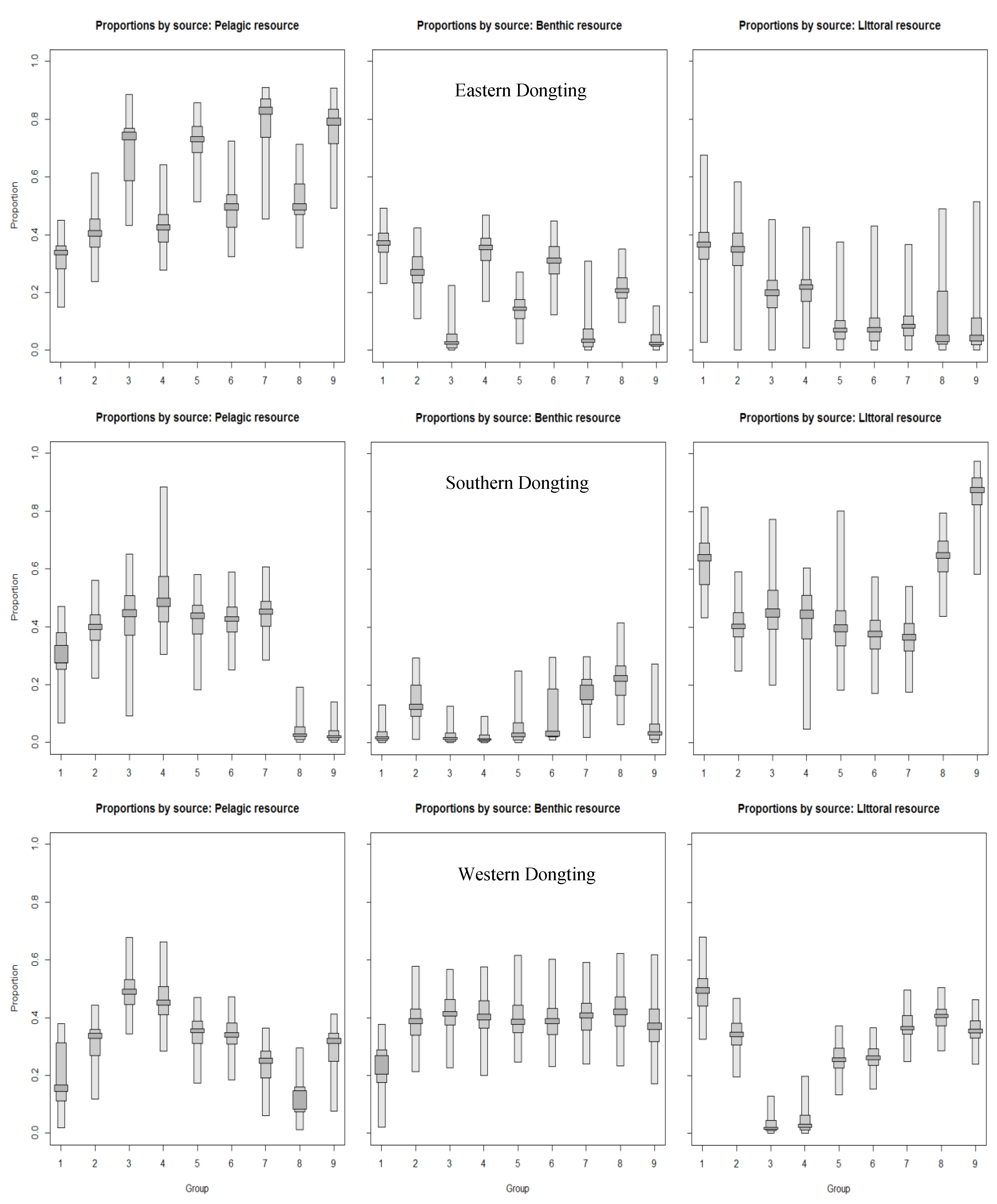
3.4. Contribution Proportions of Primary Food Sources to Fishes
4. Discussion
4.1. Spatial Differences in Fish Community and Top-Down Effects of Omnivorous Fish on Primary Food Sources
4.2. Effect of Primary Food Sources on the Trophic Structure of Fish Consumers
4.3. Different Contribution Proportions of Primary Food Sources
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Post, D.M. Using stable isotopes to estimate trophic position: Models, methods, and assumptions. Ecology 2002, 83, 703–718. [Google Scholar] [CrossRef]
- Karlsson, J.; Jansson, M.; Jonsson, A. Similar relationships between pelagic primary and bacterial production in clear water and humic lakes. Ecology 2002, 83, 2902–2910. [Google Scholar] [CrossRef]
- Vadeboncoeur, Y.; Lodge, D.M. Periphyton production on wood and sediment: Substratum-specific response to laboratory and whole-lake nutrient manipulations. J. N. Am. Benthol. Soc. 2000, 19, 68–81. [Google Scholar] [CrossRef]
- Vadeboncoeur, Y.; Lodge, D.M.; Carpenter, S.R. Whole-lake fertilization effects on distribution of primary production between benthic and pelagic habitats. Ecology 2001, 82, 1065–1077. [Google Scholar] [CrossRef]
- Blumenshine, S.C.; Vadeboncoeur, Y.D.; Lodge, M.; Cottingham, K.L.; Knight, S.E. Benthic-pelagic links: Responses of benthos to water-column nutrient enrichment. J. N. Am. Benthol. Soc. 1997, 16, 466–479. [Google Scholar] [CrossRef]
- Schindler, D.E.; Scheuerell, M.D. Habitat coupling in lake ecosystems. Oikos 2002, 98, 177–189. [Google Scholar] [CrossRef]
- Vadeboncoeur, Y.; Vander Zanden, M.J.; Lodge, D.M. Putting the lake back together: Reintegrating benthic pathways into lake food webs. Bioscience 2002, 52, 44–54. [Google Scholar] [CrossRef]
- Sha, Y.C.; Su, G.H.; Zhang, P.Y.; Zhang, H.; Xu, J. Diverse dietary strategy of lake anchovy Coilia ectenes taiensis in lakes with different trophic status. J. Ichthyol. 2015, 55, 866–873. [Google Scholar] [CrossRef]
- Yao, X.Y.; Huang, G.T.; Xie, P.; Xu, J. Trophic niche differences between coexisting omnivores silver carp and bighead carp in a pelagic food web. Ecol. Res. 2016, 31, 831–839. [Google Scholar] [CrossRef]
- Schindler, D.E.; Hodgson, J.R.; Kitchell, J.F. Density-dependent changes in individual foraging specialization of largemouth bass. Oecologia 1997, 110, 592–600. [Google Scholar] [CrossRef] [PubMed]
- Power, M.E.; Dietrich, W.E. Food webs in river networks. Ecol. Res. 2002, 17, 451–471. [Google Scholar] [CrossRef]
- Lindeman, R.L. The trophic-dynamic aspect of ecology. Ecology 1942, 23, 399–417. [Google Scholar] [CrossRef]
- Dickman, E.M.; Newell, J.M.; Gonzalez, M.J.; Vanni, M.J. Light, nutrients and food-chain length constrain planktonic energy transfer efficiency across multiple trophic levels. Proc. Natl. Acad. Sci. USA 2008, 105, 18408–18412. [Google Scholar] [CrossRef] [PubMed]
- Butchart, S.H.M.; Walpole, M.; Collen, B.; van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global biodiversity: Indicators of recent declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Essington, T.E.; Beaudreau, A.H.; Wiedenmann, J. Fishing through marine food webs. Proc. Natl. Acad. Sci. USA 2006, 103, 3171–3175. [Google Scholar] [CrossRef] [PubMed]
- Branch, T.A.; Watson, R.; Fulton, E.A.; Jennings, S.; McGilliard, C.R.; Pablico, G.T.; Ricard, D.; Tracey, S.R. The trophic fingerprint of marine fisheries. Nature 2010, 468, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, M.; Xie, P. Sympatric variability of isotopic baselines influences modeling of fish trophic patterns. Limnology 2011, 12, 107–115. [Google Scholar] [CrossRef]
- Briones, J.C.A.; Papa, R.D.S.; Cauyan, G.A.; Mendoza, N.; Okuda, N. Fish diversity and trophic interactions in Lake Sampaloc (Luzon Is. Philippines). Trop. Ecol. 2016, 57, 567–581. [Google Scholar]
- De Bernardi, R.; Giussani, G. Are blue-green algae a suitable food for zooplankton? An overview. Hydrobiologia 1990, 61, 29–41. [Google Scholar] [CrossRef]
- Gu, B.; Schelske, C.; Hoyer, M. Stable isotopes of carbon and nitrogen as indicators of diet and trophic structure of the fish community in a shallow hypereutrophic lake. J. Fish. Biol. 1996, 49, 1233–1243. [Google Scholar] [CrossRef]
- Fry, B. Stable isotope diagrams of freshwater food webs. Ecology 1991, 72, 2293–2297. [Google Scholar] [CrossRef]
- Zeug, S.C.; Winemiller, K.O. Evidence supporting the importance of terrestrial carbon in a large-river food web. Ecology 2008, 87, 1733–1743. [Google Scholar] [CrossRef]
- Winemiller, K.O.; Zeug, S.C.; Robertson, C.R.; Winemiller, B.K.; Honeycutt, R.L. Food-web structure of coastal streams in Costa Rica revealed by dietary and stable isotope analyses. J. Trop. Ecol. 2011, 27, 463–476. [Google Scholar] [CrossRef]
- Saito, L.; Rosen, M.R.; Chandra, S.; Fritsen, C.H.; Arufe, J.A.; Redd, C. Using semi-permeable membrane devices and stable nitrogen isotopes to detect anthropogenic influences on the Truckee River, USA. Environ. Eng. Sci. 2008, 25, 585–600. [Google Scholar] [CrossRef]
- Bristow, L.A.; Jickells, T.D.; Weston, K.; Marca-Bell, A.; Parker, R.; Andrews, J.E. Tracing estuarine organic matter sources into the southern North Sea using C and N isotopic signatures. Biogeochemistry 2013, 113, 9–22. [Google Scholar] [CrossRef]
- Li, Y.S.; Li, Y.; Yu, D.B.; Xia, M.; Hu, S.Q.; Xiang, Y.; Zhong, Z.N. A multivariate analysis of the relationship between work ability and S. japonicum infection in Dongting Lake Region, in China. Rev. Inst. Med. Trop. São Paulo 1993, 35, 347–353. [Google Scholar] [PubMed]
- Li, C. Study on the Investigation of Main Economic Fishery Resource and the Law of Its variety in the Dongting Lake. Master’s Thesis, Hunan Agricultural University, Changsha, China, 2006. [Google Scholar]
- Qian, Y.; Zheng, M.; Zhang, B.; Gao, L.; Liu, W. Determination and assessment of HCHs and DDTs residues in sediments from Lake Dongting, China. Environ. Monit. Assess. 2006, 116, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, K.; Yang, Y.; Qi, L.; Hayashi, A.S.; Watanabe, M. Measuring water storage fluctuations in Lake Dongting, China, by Topex/Poseidon satellite altimetry. Environ. Monit. Assess. 2006, 115, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Yu, H.; Jiang, X. Within-lake heterogeneity of environmental factors structuring bacterial community composition in Lake Dongting, China. World J. Microbiol. Biotechnol. 2015, 31, 1683–1689. [Google Scholar] [CrossRef] [PubMed]
- Ru, H.J.; Liu, X.Q.; Huang, X.R.; Ning, Y.Z.; Wang, H.Z. Diversity of fish species and its spatio-temporal variations in Lake Dongting, a large Yangtze connected lake. J. Lake Sci. 2008, 20, 93–99. [Google Scholar]
- Marianna, P.; Mario, A.; Loreen, H.; Nicolas, S.; Loïc, N.M.; Ursula, S.; Krishna, D. Carbon, nitrogen and sulphur isotopic fractionation in captive juvenile hooded seal (Cystophora cristata): Application for diet analysis. Rapid Commun. Mass Spectrom. 2017, 31, 1720–1728. [Google Scholar]
- Li, Y.S.; Sleigh, A.C.; Ross, A.G.P.; Williams, G.M.; Tanner, M.; McManus, D.P. Epidemiology of Schistosoma japonicum in China: Morbidity and strategies for control in the Dongting Lake region. Int. J. Parasitol. 2000, 30, 273–281. [Google Scholar] [CrossRef]
- Li, Y.Z.; Chen, X.S.; Xie, Y.H.; Li, X.; Li, F.; Hou, Z.Y. Effects of young poplar plantations on understory plant diversity in the Dongting Lake wetlands, China. Sci. Rep. UK 2014, 4, 6339. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Chen, W.; Cai, Q. Survey, Observation and Analysis of Lake Ecology; Standard Methods for Observation and Analysis in Chinese Ecosystem Research Network, Series V; Standards Press of China: Beijing, China, 1999. [Google Scholar]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China; Science and Technology Press: Beijing, China, 2006. [Google Scholar]
- Chiang, S.C.; Du, N.S. Fauan Sinica, Crustacea, Freshwater Cladocera; Science Press, Academia Sinica: Beijing, China, 1979. [Google Scholar]
- Shen, J.R. Fauna Sinica, Crustacea, Freshwater Copepoda; Science Press, Academia Sinica: Beijing, China, 1979. [Google Scholar]
- Xu, J.; Zhang, M.; Xie, P. Stable isotope changes in freshwater shrimps (Exopalaemon modestus and Macrobrachium nipponensis): Trophic pattern implications. Hydrobiologia 2008, 605, 45–54. [Google Scholar] [CrossRef]
- Sand-Jensen, K.J.; Borum, P. Interactions among phytoplankton, periphyton, and macrophytes in temperate freshwaters and estuaries. Aquat. Bot. 1991, 41, 137–175. [Google Scholar] [CrossRef]
- Vander Zanden, M.J.; Chandra, S.; Allen, B.C.; Reuter, J.E.; Goldman, C.R. Historical food web structure and restoration of native aquatic communities in the Lake Tahoe (California-Nevada) Basin. Ecosystems 2003, 6, 274–288. [Google Scholar] [CrossRef]
- Parnell, A.C.; Inger, R.; Bearhop, S.; Jackson, A.L. Source Partitioning Using Stable Isotopes: Coping with Too Much Variation. PLoS ONE 2010, 5, 9672. [Google Scholar] [CrossRef] [PubMed]
- Peres-neto, P.R.; Jackson, D.A. How well do multivariate data sets match? The advantages of a Procrustean superimposition approach over the Mantel test. Oecologia 2001, 129, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Vuorio, K.; Meili, M.; Sarvala, J. Taxon-specific variation in the stable isotopic signatures (δ13C and δ15N) of lake phytoplankton. Freshw. Biol. 2006, 51, 807–822. [Google Scholar] [CrossRef]
- Sanchez-Hernandez, J.; Eloranta, A.P.; Finstad, A.G.; Amundsen, P.A. Community structure affects trophic ontogeny in a predatory fish. Ecol. Evol. 2017, 7, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Q. Research Progress of Fish resources in Dongting Lake. J. Anhui Agric. Sci. 2013, 41, 3898–3900. [Google Scholar]
- Iii, J.J.P.; Santucci, V.J.; Wahl, D.H. Effects of adult common carp (Cyprinus carpio) on multiple trophic levels in shallow mesocosms. Can. J. Fish. Aquat. Sci. 2003, 192, 182–192. [Google Scholar]
- Matsuzaki, S.S.; Usio, N.; Takamura, N.; Washitani, I. Effects of common carp on nutrient dynamics and littoral community composition: Roles of excretion and bioturbation. Fundam. Appl. Limnol. 2007, 168, 70176. [Google Scholar] [CrossRef]
- Xu, J.; Wen, Z.; Ke, Z.; Zhang, M.; Zhang, M.; Guo, N.; Hansson, L.-A.; Xie, P. Contrasting energy pathways at the community level as a consequence of regime shifts. Oecologia 2014, 175, 231–241. [Google Scholar] [CrossRef] [PubMed]
- De Bruyn, A.M.H.; Rasmussen, J.B. Quantifying assimilation of sewage derived organic matter by riverine benthos. Ecol. Appl. 2002, 12, 511–520. [Google Scholar] [CrossRef]
- Winemiller, K.O.; Hoeinghaus, D.J.; Pease, A.A.; Esselman, P.C.; Honeycutt, R.L.; Gbanaador, D.G.; Carrera, E.; Payne, J. Stable isotope analysis reveals food web structure and watershed impacts along the fluvial gradient of a Mesoamerican coastal river. River Res. Appl. 2011, 27, 791–803. [Google Scholar] [CrossRef]
- Jeppesen, E.; Meerhoff, M.; Davidson, T.A.; Trolle, D.; Søndergaard, M.; Lauridsen, T.L.; Beklioglu, M.; Brucet, S.; Volta, P.; González-Bergonzoni, I.; et al. Climate change impacts on lakes: An integrated ecological perspective based on a multi-faceted approach, with special focus on shallow lakes. J. Limnol. 2014, 73, 88–111. [Google Scholar] [CrossRef]
- Jeppesen, E.; Søndergaard, M.; Liu, Z. Lake Restoration and Management in a Climate Change Perspective: An Introduction. Water 2017, 9, 122. [Google Scholar] [CrossRef]
- Fox, A.D.; Cao, L.; Barter, M.; Rees, E.C.; Hearn, R.D.; Cong, P.H.; Wang, X.; Zhang, Y.; Dou, S.T.; Shao, X.F. The functional use of East Dongting Lake, China, by wintering geese. Wildfowl 2008, 58, 3–19. [Google Scholar]
- Anderson, C.; Cabana, G. δ15N in riverine food webs: Effects of N inputs from agricultural watersheds. Can. J. Fish. Aquat. Sci. 2005, 62, 333–340. [Google Scholar] [CrossRef]
- Simon, K.S.; Niyogi, D.K.; Frew, R.D.; Townsend, C.R. Nitrogen dynamics in grassland streams along a gradient of agricultural development. Limnol. Oceanoga 2007, 52, 1246–1257. [Google Scholar] [CrossRef]
- Thomas, S.H.; Kiljunen, M.; Malinen, T.; Eloranta, A.P.; Amundsen, P.A.; Lodenius, M.; Kahilnen, K.K. Food-web structure and mercury dynamics in a large subarctic lake following multiple species introductions. Freshw. Biol. 2016, 61, 500–517. [Google Scholar] [CrossRef]
- Yang, G.S.; Zhang, Q.; Wan, R.R.; Lai, X.J.; Jiang, X.; Li, L.; Dai, H.C.; Lei, G.C.; Chen, J.C.; Lu, Y.J. Lake hydrology, water quality and ecology impacts of altered river–lake interactions: Advances in research on the middle Yangtze river. Hydrol. Res. 2016, 47. [Google Scholar] [CrossRef]
- Bricker, S.B.; Rice, K.C.; Bricker, O.P. From Headwaters to Coast: Influence of Human Activities on Water Quality of the Potomac River Estuary. Aquat. Geochem. 2014, 20, 291–323. [Google Scholar] [CrossRef]
- Hu, Y.; Lu, Y.H.; Liu, C.K.; Shang, P.; Liu, J.; Zheng, C.M. Sources and Dynamics of Dissolved Inorganic Carbon, Nitrogen, and Phosphorus in a Large Agricultural River Basin in Arid Northwestern China. Water 2017, 9, 415. [Google Scholar] [CrossRef]
- Xie, Y.H.; Tang, Y.; Chen, X.S.; Li, F.; Deng, Z.M. The impact of Three Gorges Dam on the downstream eco-hydrological environment and vegetation distribution of East Dongting Lake. Ecohydrology 2015, 8, 738–746. [Google Scholar] [CrossRef]
- Barry, J.F.; Murray, E. Periphyton biomass dynamics in gravel bed rivers: The relative effects of flows and nutrients. Freshw. Biol. 1989, 10, 1365–2427. [Google Scholar]
- Jeppesen, E.; Jensen, J.P.; Søndergaard, M.; Lauridsen, T. Trophic structure, species richness and biodiversity in Danish lakes: Changes along a phosphorus gradient. Freshw. Biol. 2000, 201–218. [Google Scholar] [CrossRef]
- Gremberghe, I.V. How Do Diversity and Identity at Different Trophic Levels Influence the Strength of Top-Down Control in a Simple Aquatic Food Web? A Mediation Analysis. Master’s Thesis, Ghent University, Ghent, Belgium, 2016. [Google Scholar]
- Colombo, F.; Costa, V.; Stanislas, F.; Dubois, S.F.; Gianguzza, P.; Mazzola, A.; Vizzini, S. Trophic structure of vermetid reef community: High trophic diversity at small spatial scales. J. Sea Res. 2013, 77, 93–99. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Eastern Dongting | Southern Dongting | Western Dongting | |
|---|---|---|---|
| Sub-lake area (km2) | 1300 | 900 | 340 |
| Water temperature (°C) | 25.28 ± 0.22 a | 24.04 ± 0.35 ab | 23.56 ± 0.35 b |
| ORP (mv) | 79.20 ± 24.22 a | 112.36 ± 22.35 b | 161.17 ± 15.36 c |
| Water depth (m) | 5.47 ± 2.13 a | 2.85 ± 0.74 b | 3.34 ± 0.69 c |
| TN (mg/L) | 1.98 ± 0.14 a | 1.44 ± 0.11 b | 1.78 ± 0.15 ab |
| NO3-N (mg/L) | 1.24 ± 0.15 a | 0.64 ± 0.12 b | 0.74 ± 0.20 ab |
| NH4-N (mg/L) | 0.14 ± 0.02 a | 0.43 ± 0.09 b | 0.36 ± 0.05 c |
| TP (mg/L) | 0.10 ± 0.01 a | 0.10 ± 0.02 a | 0.07 ± 0.01 a |
| Density of phytoplankton (cells/L) | (10.26 ± 4.19) × 106 a | (1.09 ± 0.19) × 106 b | (1.20 ± 0.31) × 106 b |
| Density of zooplankton (inds/L) | 29.09 ± 5.71 a | 3.51 ± 1.40 b | 1.66 ± 0.72 c |
| Eastern and Southern Dongting | Eastern and Western Dongting | Southern and Western Dongting | |||||||
|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | |
| Ctenopharynodon idellus | 17 | 2.914 | 0.107 | 17 | 0.021 | 0.886 | 17 | 3.067 | 0.099 |
| Parabramis pekinensis | 17 | 6.855 | 0.019 | 17 | 8.472 | 0.010 | 17 | 0.181 | 0.676 |
| Hypophthalmichthys molitrix | 17 | 135.099 | <0.001 | 17 | 18.855 | 0.001 | 17 | 150.433 | <0.001 |
| Aristichthys nobilis | 17 | 65.325 | <0.001 | 17 | 3.131 | 0.096 | 17 | 5.983 | 0.026 |
| Acheilognathus macropterus | 17 | 10.299 | 0.005 | 17 | 11.259 | 0.004 | 17 | 0.285 | 0.601 |
| Carassius auratus | 17 | 6.175 | 0.024 | 17 | 8.311 | 0.011 | 17 | 0.501 | 0.489 |
| Cyprinus carpio | 17 | 9.694 | 0.007 | 17 | 25.780 | <0.001 | 17 | 0.190 | 0.669 |
| Cultrichthys erythropterus | 17 | 6.885 | 0.018 | 17 | 136.225 | <0.001 | 17 | 19.543 | <0.001 |
| Siniperca chuatsi | 17 | 4.822 | 0.043 | 17 | 13.181 | 0.002 | 17 | 2.495 | 0.134 |
| Primary Food Sources | Eastern and Southern Dongting | Eastern and Western Dongting | Southern and Western Dongting | One-Way ANOVA |
|---|---|---|---|---|
| Pelagic source | 0.07 | 0.03 | 0.921 | Turkey HSD |
| Benthic source | 0.155 | <0.001 | <0.001 | Dunnett T3 |
| Littoral source | <0.001 | 0.421 | 0.001 | Turkey HSD |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Guo, L.; Zhang, H.; Xu, J.; Hu, H.; Xue, T.; Luo, C.; Yi, C.; Hu, Y.; Silva, K.H.W.L.D.; et al. Spatial Variation in Trophic Structure of Dominant Fish Species in Lake Dongting, China during Dry Season. Water 2018, 10, 602. https://doi.org/10.3390/w10050602
Yu J, Guo L, Zhang H, Xu J, Hu H, Xue T, Luo C, Yi C, Hu Y, Silva KHWLD, et al. Spatial Variation in Trophic Structure of Dominant Fish Species in Lake Dongting, China during Dry Season. Water. 2018; 10(5):602. https://doi.org/10.3390/w10050602
Chicago/Turabian StyleYu, Jia, Longgen Guo, Huan Zhang, Jun Xu, Huaming Hu, Ting Xue, Congqiang Luo, Chunlong Yi, Yufei Hu, Kaluwa Handi Wasana Lalanthi De Silva, and et al. 2018. "Spatial Variation in Trophic Structure of Dominant Fish Species in Lake Dongting, China during Dry Season" Water 10, no. 5: 602. https://doi.org/10.3390/w10050602
APA StyleYu, J., Guo, L., Zhang, H., Xu, J., Hu, H., Xue, T., Luo, C., Yi, C., Hu, Y., Silva, K. H. W. L. D., & Xie, P. (2018). Spatial Variation in Trophic Structure of Dominant Fish Species in Lake Dongting, China during Dry Season. Water, 10(5), 602. https://doi.org/10.3390/w10050602