Efficient Ammonium Removal by Bacteria Rhodopseudomonas Isolated from Natural Landscape Water: China Case Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization and Identification of the Bacterial Strain
2.2. Effect of Culture Conditions on Bacterial Growth Characteristics
2.3. Application of Smobiisys501 to Ammonium Removal
2.4. Statistical Analysis and Graphical Work
3. Results and Discussion
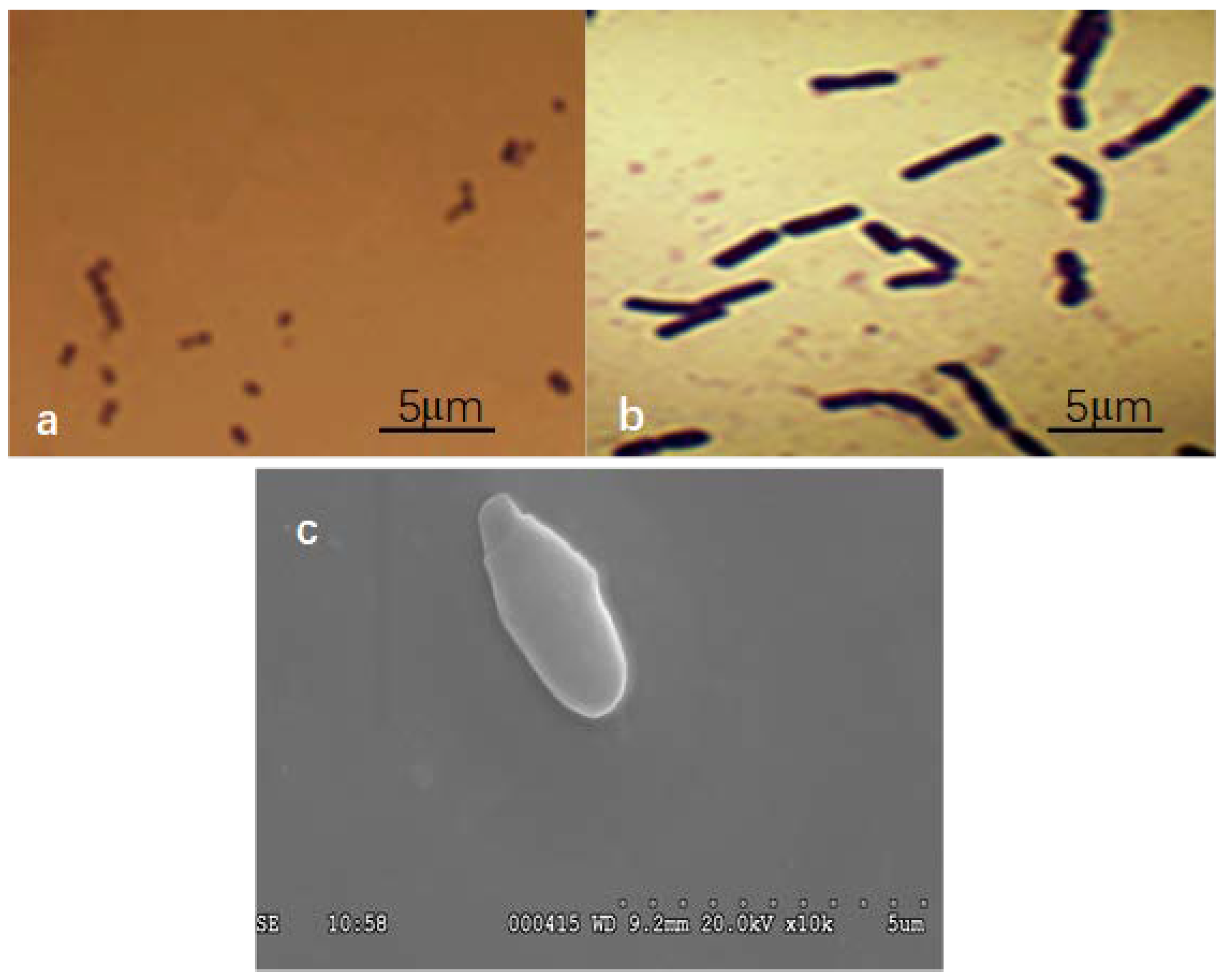
3.1. Characterization of Smobiisys501
3.1.1. Morphological and Absorption Spectra Characteristics
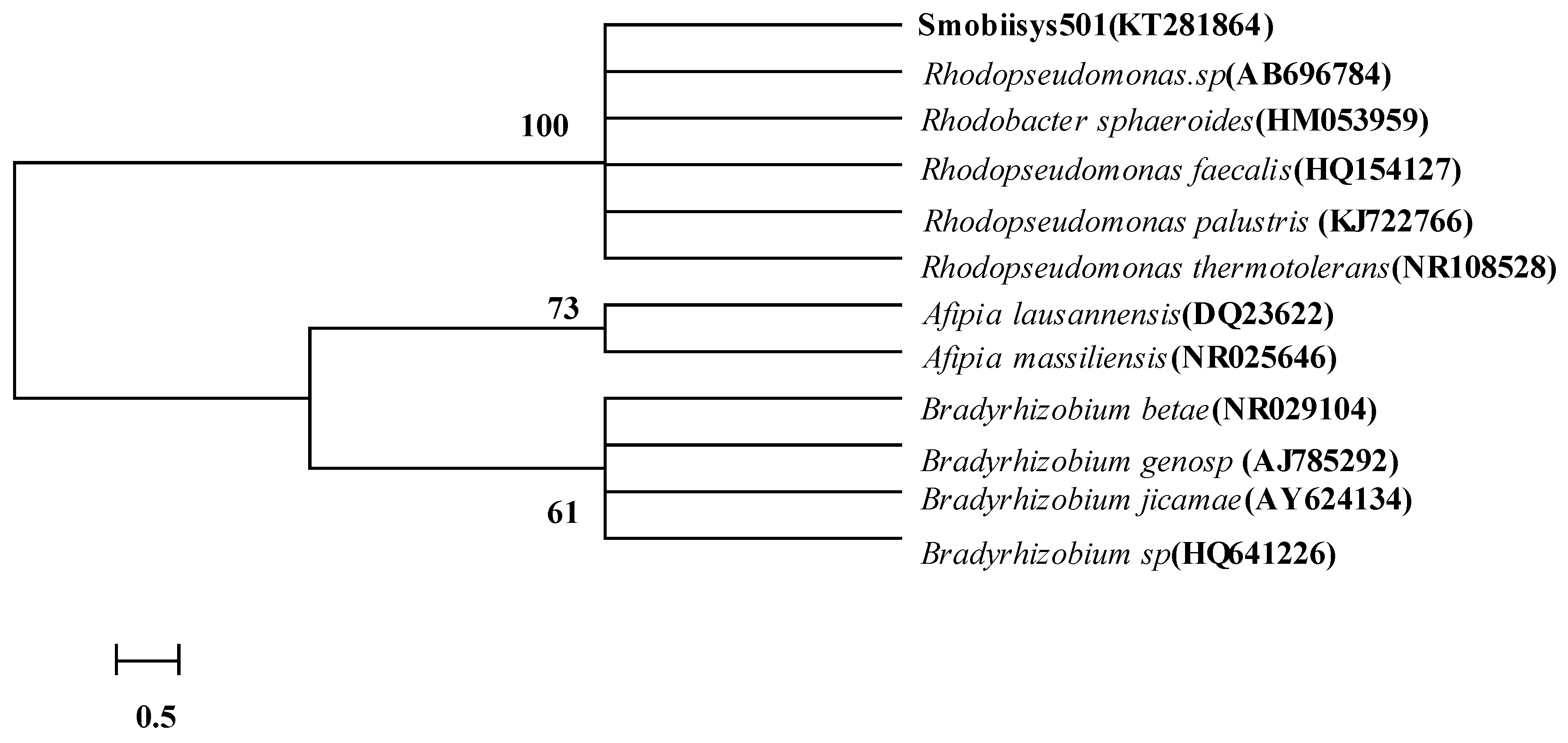
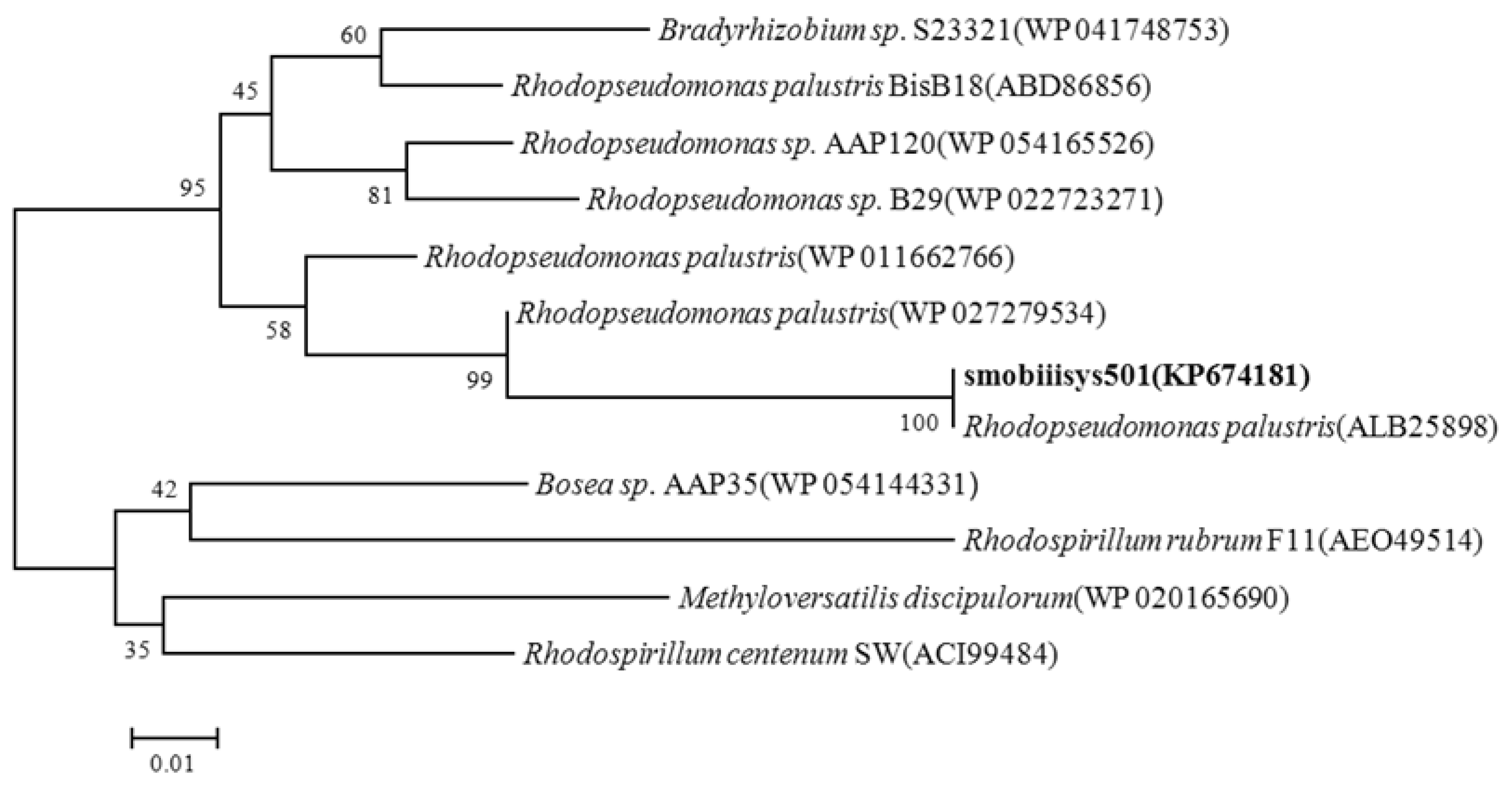
3.1.2. Phylogenetic Analysis
3.1.3. Physiological-Biochemical Characteristics
3.1.4. DNA–DNA Hybridization
3.1.5. Cellular Fatty Acid Analysis
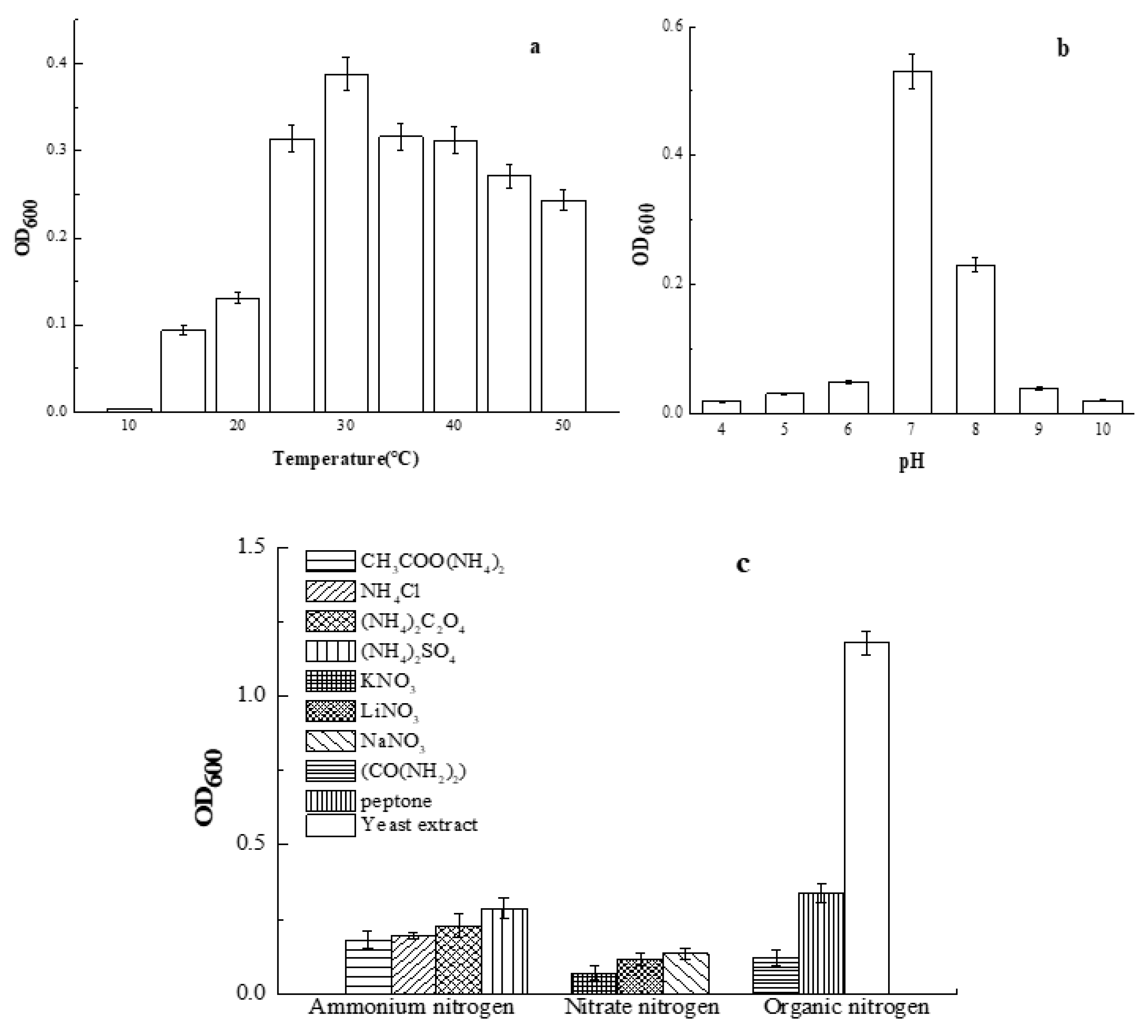
3.2. Effects of Culture Conditions on the Growth of Smobiisys501
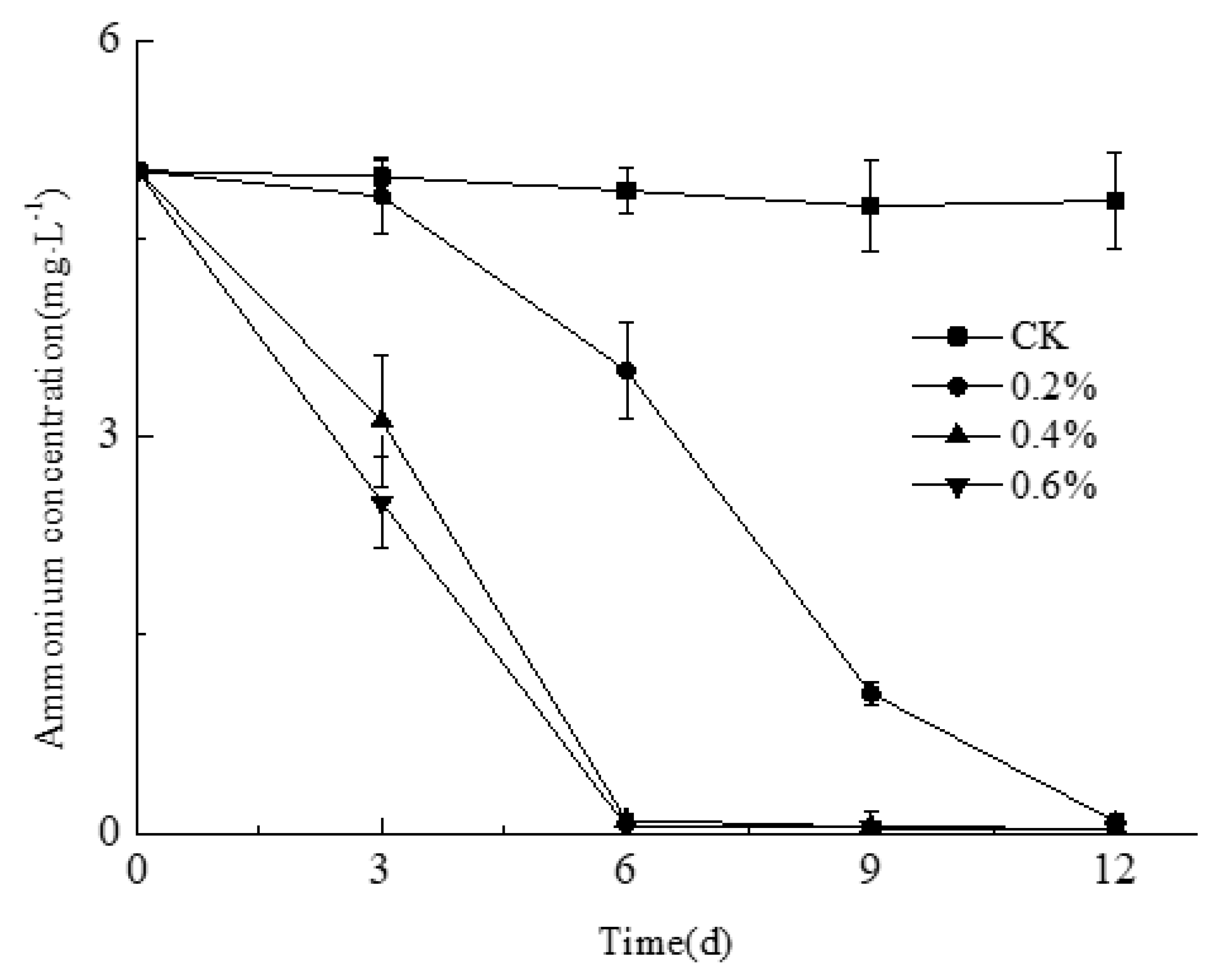
3.3. Assessment of Ammonium Removal by Smobiisys501
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Czurda, V.; Maresch, E. Beitrag zur Kenntnis der Athiorhodobakterien-Gesellschaften. Arch. Mikrobiol. 1937, 8, 99–124. [Google Scholar] [CrossRef]
- Wei, H.; Okunishi, S.; Yoshikawa, T.; Kamei, Y.; Maeda, H. Isolation and characterization of a purple non-sulfur photosynthetic bacterium Rhodopseudomonas faecalis strain a from swine sewage wastewater. Biocontrol Sci. 2016, 21, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Hiraishi, A.; Okamura, K. Rhodopseudomonas telluris sp. nov., a phototrophic alphaproteobacterium isolated from paddy soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 3369–3374. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.F.; Jin, Y.R.; Cui, Q.F.; Xie, G.J.; Wu, Y.N.; Ren, N.Q. Photo-fermentation hydrogen production by Rhodopseudomonas sp. nov. strain A7 isolated from the sludge in a bioreactor. Int. J. Hydrog. Energy 2015, 40, 8661–8668. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Trüper, H.G.; Pfennig, N. Rearrangement of the species and genera of the phototrophic “purple nonsulfur bacteria”. Int. J. Syst. Evol. Microbiol. 1984, 34, 340–343. [Google Scholar] [CrossRef]
- Okamura, K.; Takata, K.; Hiraishi, A. Intrageneric relationships of members of the genus Rhodopseudomonas. J. Gen. Appl. Microbiol. 2009, 55, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.V.; Ramprasad, E.V.V.; Sasikala, C.; Ramana, C.V. Rhodopseudomonas pentothenatexigens sp. nov. and Rhodopseudomonas thermotolerans sp. nov., isolated from paddy soils. Int. J. Syst. Evol. Microbiol. 2013, 63, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Phankhamla, P.; Sawaengkaew, J.; Buasri, P.; Mahakhan, P. Biohydrogen production by a novel thermotolerant photosynthetic bacterium Rhodopseudomonas pentothenatexigens strain KKU-SN1/1. Int. J. Hydrog. Energy 2014, 39, 15424–15432. [Google Scholar] [CrossRef]
- Guan, D.W.; Li, J.; Shen, D.L.; Cao, F.M.; Li, L. Construction and application of photosynthetic bacteria PCR detection technology. Chin. J. Appl. Environ. Biol. 2008, 14, 699–704. [Google Scholar]
- Yutin, N.; Suzuki, M.T.; Rosenberg, M.; Rotem, D.; Madigan, M.T.; Süling, J.; Imhoff, J.F.; Béjà, O. BchY-based degenerate primers target all types of anoxygenic photosynthetic bacteria in a single PCR. Int. J. Syst. Evol. Microbiol. 2009, 75, 7556–7559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.P.; Shu, M.A.; Wang, Y.B.; Fu, L.Q.; Li, W.F.; Deng, B.; Liang, Q.; Shen, W.Y. Effect of photosynthetic bacteria on water quality and microbiota in grass carp culture. World J. Microbiol. Biotechnol. 2014, 30, 2523–2531. [Google Scholar] [CrossRef] [PubMed]
- Hülsen, T.; Batstone, D.J.; Keller, J. Phototrophic bacteria for nutrient recovery from domestic wastewater. Water Res. 2014, 50, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Yang, A.; Wang, H.; Zhang, G.; Li, X.; Zhang, Y.; Zou, Z. One-step treatment and resource recovery of high-concentration non-toxic organic wastewater by photosynthetic bacteria. Bioresour. Technol. 2018, 251, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Talaiekhozani, A.; Rezania, S. Application of photosynthetic bacteria for removal of heavy metals, macro-pollutants and dye from wastewater: A review. J. Water Process Eng. 2017, 19, 312–321. [Google Scholar] [CrossRef]
- Huang, X.J.; Yang, C.; Ni, J.P.; Li, Z.L. Isolation, identification and characteristics of a Rhodopseudomonas with high ammonia-nitrogen removal efficiency. Environ. Sci. 2016, 37, 2276–2283. [Google Scholar]
- Madukasi, E.I.; Chunhua, H.; Zhang, G. Isolation and application of a wild strain photosynthetic bacterium to environmental waste management. Int. J. Environ. Sci. Technol. 2011, 8, 513–522. [Google Scholar] [CrossRef] [Green Version]
- Lakshmi, K.V.N.S.; Sasikala, C.; Ramana, C.V. Rhodoplanes pokkaliisoli sp. Nov., a phototrophic alphaproteobacterium isolated from a waterlogged brackish paddy soil. Int. J. Syst. Evol. Microbiol. 2009, 59, 2153–2157. [Google Scholar] [CrossRef] [PubMed]
- Hot, G.J.; Krieg, R.N.; Sneath, H.A.P. Bergey’s Manual of Determinative Bacteriology; Baltirnore: Washington, DC, USA, 1994. [Google Scholar]
- Dong, Z.X.; Cai, M.Y. Handbook of Common Bacterial System Identification; Thomson Learning Press: Beijing, China, 2001. [Google Scholar]
- Ley, J.; Cattoir, H.; Reynaerts, A. The quantitative measurement of DNA hybridization from renaturation rates. Eur. J. Biochem. 1970, 12, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Shi, W.; Ni, J.; Li, Z.L. Evaluation of laboratory-scale in situ capping sediments with purple parent rock to control the eutrophication. Environ. Sci. Pollut. Res. 2017, 24, 7114–7123. [Google Scholar] [CrossRef] [PubMed]
- Romanenko, L.A.; Tanaka, N.; Svetashev, V.I. Devosia submarina sp. nov., isolated from deep sea surface sediments. Int. J. Syst. Evol. Microbiol. 2013, 63, 3079–3085. [Google Scholar] [CrossRef] [PubMed]
- State Environmental Protection Administration. Methods for Monitoring and Analysis of Water and Wastewater; Chinese Environmental Science Press: Beijing, China, 2012. [Google Scholar]
- Hiraishi, A.; Muramatsu, K.; Urata, K. Characterization of new denitrifying Rhodobacter strainsisolated from photosynthetic sludge for wastewater treatment. J. Ferment. Bioeng. 1995, 79, 39–44. [Google Scholar] [CrossRef]
- Madigan, M.T.; Jung, D.O.; Woese, C.R.; Achenbach, L.A. Rhodoferax antarcticus sp. nov., a moderately psychrophilic purple nonsulfur bacterium isolated from an antarctic microbial mat. Arch. Microbiol. 2000, 173, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Girija, K.; Sasikala, C.; Ramana, C.V.; Spröer, C.; Takaichi, S.; Thiel, V.; Imhoff, J.F. Rhodobacter johrii sp. nov., an endospore-producing cryptic species isolated from semi-arid tropical soils. Int. J. Syst. Evol. Microbiol. 2010, 60, 2099–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramana, V.V.; Chakravarthy, S.K.; Raj, P.S.; Kumar, B.V.; Shobha, E.; Ramaprasad, E.V.V.; Sasikala, C.; Ramana, C.V. Descriptions of Rhodopseudomonas parapalustris sp. nov., Rhodopseudomonas harwoodiae sp. nov. and Rhodopseudomonas pseudopalustris sp. nov., and emended description of Rhodopseudomonas palustris. Int. J. Syst. Evol. Microbiol. 2012, 62, 1790–1798. [Google Scholar] [CrossRef] [PubMed]
- Tindall, B.J.; Rossello-Mora, R.; Busse, H.J.; Ludwig, W.; Kämpfer, P. Notes on the characterization of prokaryote strains for taxonomic purposes. Int. J. Syst. Evol. Microbiol. 2010, 60, 249–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wayne, L.G.; Brenner, D.J.; Colwell, R.R.; Grimont, P.A.D.; Kandler, O.; Krichevsky, M.I.; Moore, H.; Moore, W.E.C.; Murray, R.G.E.; Stackebrandt, E.; et al. Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Evol. Microbiol. 1987, 37, 463–464. [Google Scholar] [CrossRef]
- Arunasri, K.; Ramana, V.V.; Spröer, C.; Sasikala, C.; Ramana, C.V. Rhodobacter megalophilus sp. nov., a phototroph from the Indian Himalayas possessing a wide temperature range for growth. Int. J. Syst. Evol. Microbiol. 2008, 58, 1792–1796. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.P.; Zhang, Y.Q. Removal of nitrogen (N) from hypereutrophic waters by ecological floating beds (EFBs) with various substrates. Ecol. Eng. 2014, 62, 148–152. [Google Scholar] [CrossRef]
- Liang, Z.; Liu, Y.; Ge, F.; Wong, M. A pH-dependent enhancement effect of co-cultured Bacillus licheniformis on nutrient removal by Chlorella vulgaris. Ecol. Eng. 2015, 75, 258–263. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, D.M.; Wang, L.G.; Pan, Z. Biological characteristics and phylogenetic analysis of a denitrifying photosynthetic bacterium. Acta Microbiol. Sin. 2011, 51, 249–255. [Google Scholar]
- Zhou, Q.; Zhang, G.; Zheng, X.; Liu, G. Biological treatment of high NH4+-N wastewater using an ammonia-tolerant photosynthetic bacteria strain (ISASWR2014). Chin. J. Chem. Eng. 2015, 23, 1712–1715. [Google Scholar] [CrossRef]
- Takabatake, H.; Suzuki, K.; Ko, I.B.; Noike, T. Characteristics of anaerobic ammonia removal by a mixed culture of hydrogen producing photosynthetic bacteria. Bioresour. Technol. 2004, 95, 151–158. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological Characteristics | Biochemical Characteristics | ||
|---|---|---|---|
| Carbon source utilization | + | Oxidase | + |
| Sodium acetate | + | Catalase | + |
| methanol | − | Indole | + |
| glucose | − | Oxidization H2S | − |
| Sodium thiosulfate | − | Starch hydrolysis | − |
| Mannitol | − | Gram stain | − |
| Fatty Acid Composition (%) | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| C12:0 | 3.05 | 1.6 | - | 1.7 |
| C16:0 | 19.13 | 20.8 | 5.1 | 14.9 |
| C16:1ω7c/16:1ω6c | 19.29 | 10.2 | 1.9 | 6.2 |
| C18:1ω7c | 38.43 | 51 | 77.2 | 63 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Ni, J.; Yang, C.; Feng, M.; Li, Z.; Xie, D. Efficient Ammonium Removal by Bacteria Rhodopseudomonas Isolated from Natural Landscape Water: China Case Study. Water 2018, 10, 1107. https://doi.org/10.3390/w10081107
Huang X, Ni J, Yang C, Feng M, Li Z, Xie D. Efficient Ammonium Removal by Bacteria Rhodopseudomonas Isolated from Natural Landscape Water: China Case Study. Water. 2018; 10(8):1107. https://doi.org/10.3390/w10081107
Chicago/Turabian StyleHuang, Xuejiao, Jiupai Ni, Chong Yang, Mi Feng, Zhenlun Li, and Deti Xie. 2018. "Efficient Ammonium Removal by Bacteria Rhodopseudomonas Isolated from Natural Landscape Water: China Case Study" Water 10, no. 8: 1107. https://doi.org/10.3390/w10081107
APA StyleHuang, X., Ni, J., Yang, C., Feng, M., Li, Z., & Xie, D. (2018). Efficient Ammonium Removal by Bacteria Rhodopseudomonas Isolated from Natural Landscape Water: China Case Study. Water, 10(8), 1107. https://doi.org/10.3390/w10081107