A Classification Method for Fish Swimming Behaviors under Incremental Water Velocity for Fishway Hydraulic Design


Abstract
1. Introduction
2. Materials and Methods
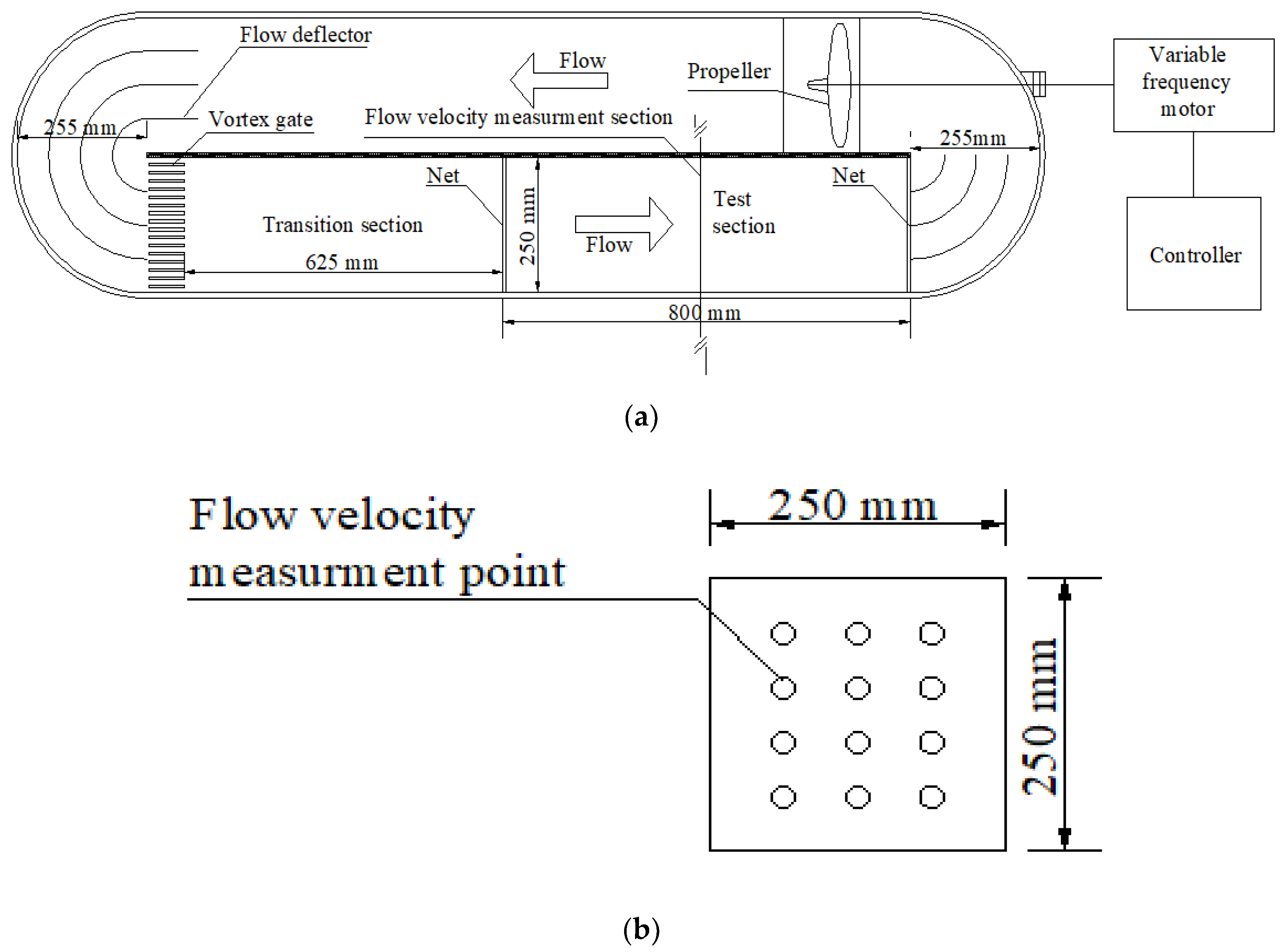
2.1. Test Equipment
2.2. Tested Fish and Environment
2.3. Test Methods
2.3.1. Incremental Water Velocity Method
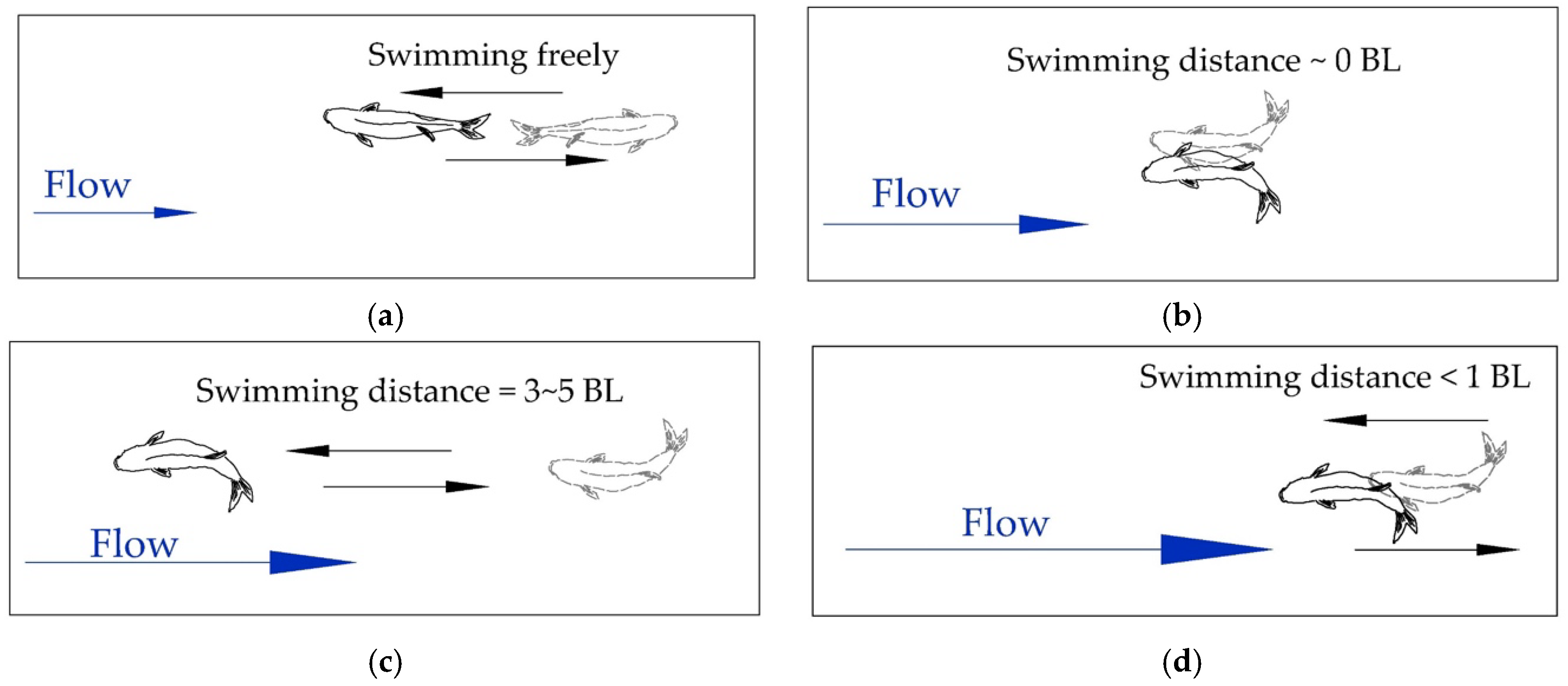
2.3.2. Classification of Fish Swimming Behaviors
3. Results
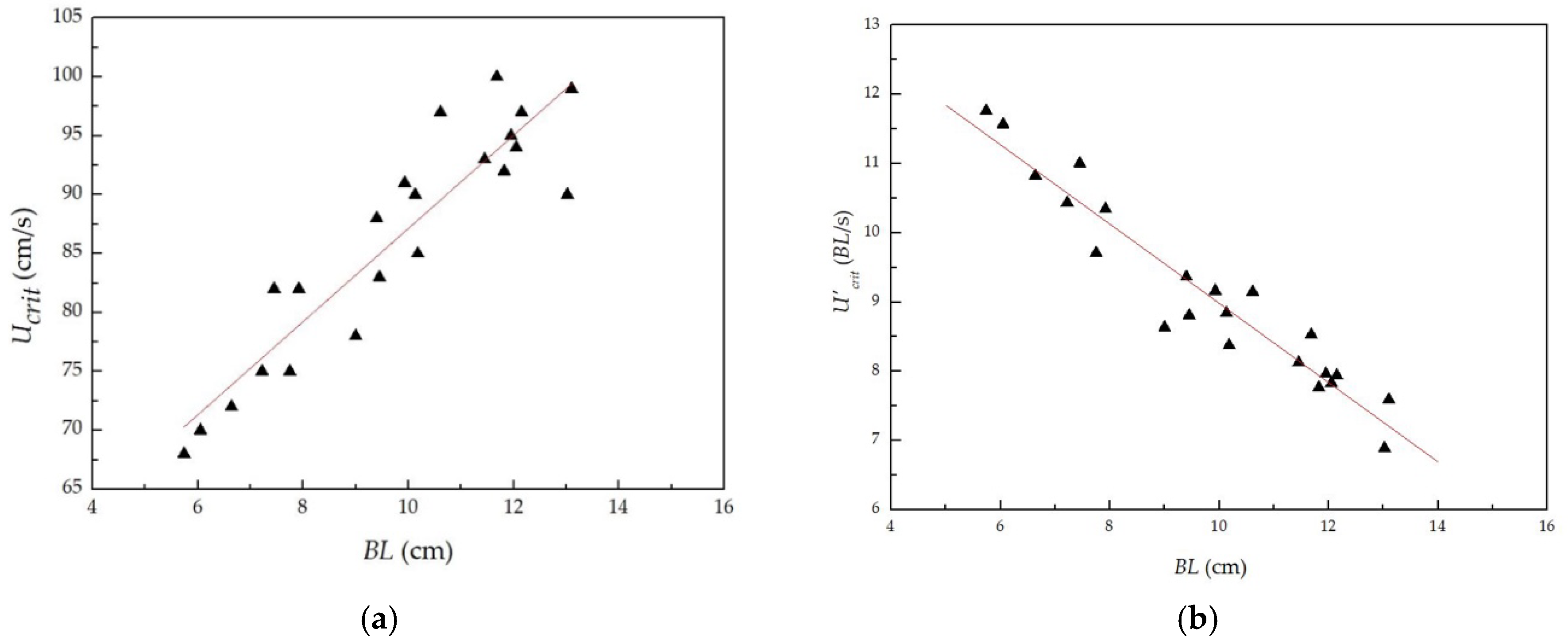
3.1. Relationship between Critical Swimming Speed and Body Length
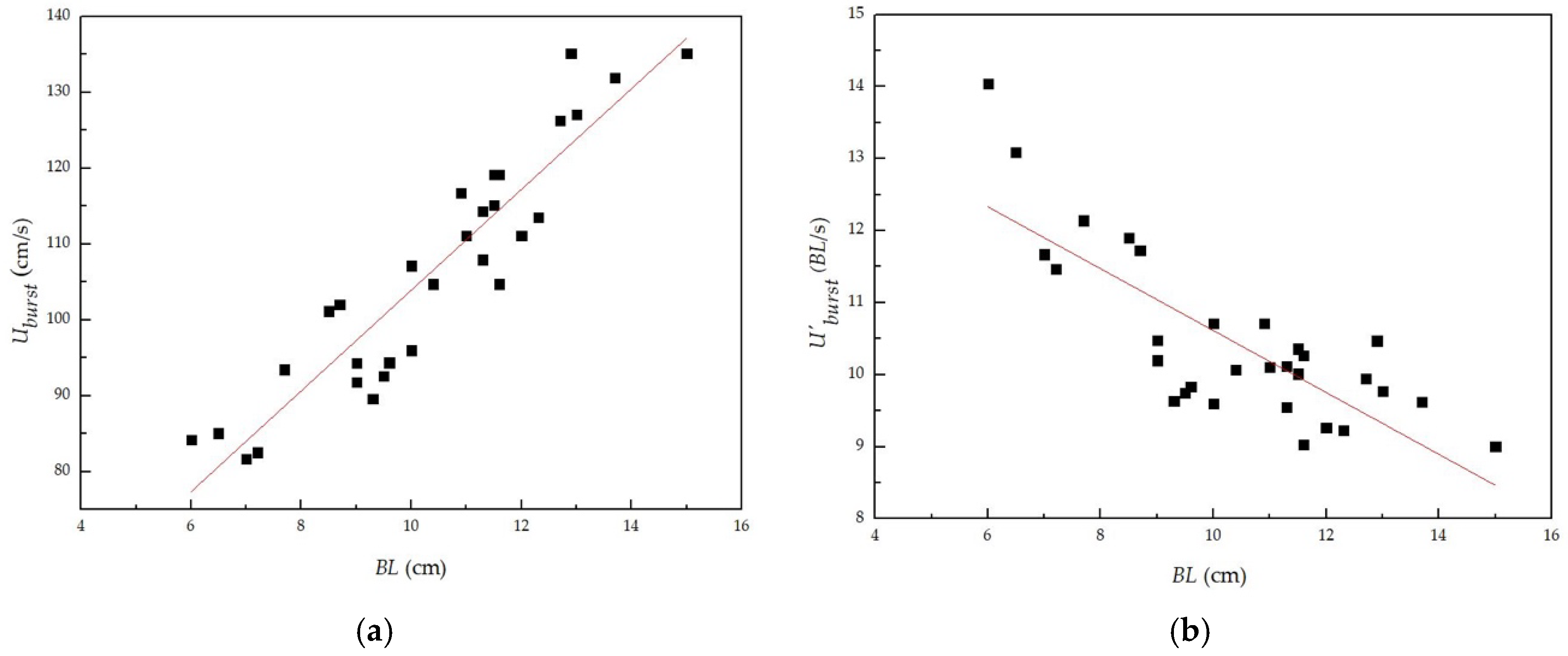
3.2. Relationship between Burst Swimming Speed and Body Length
3.3. Relationship between Absolute Critical and Burst Swimming Speeds
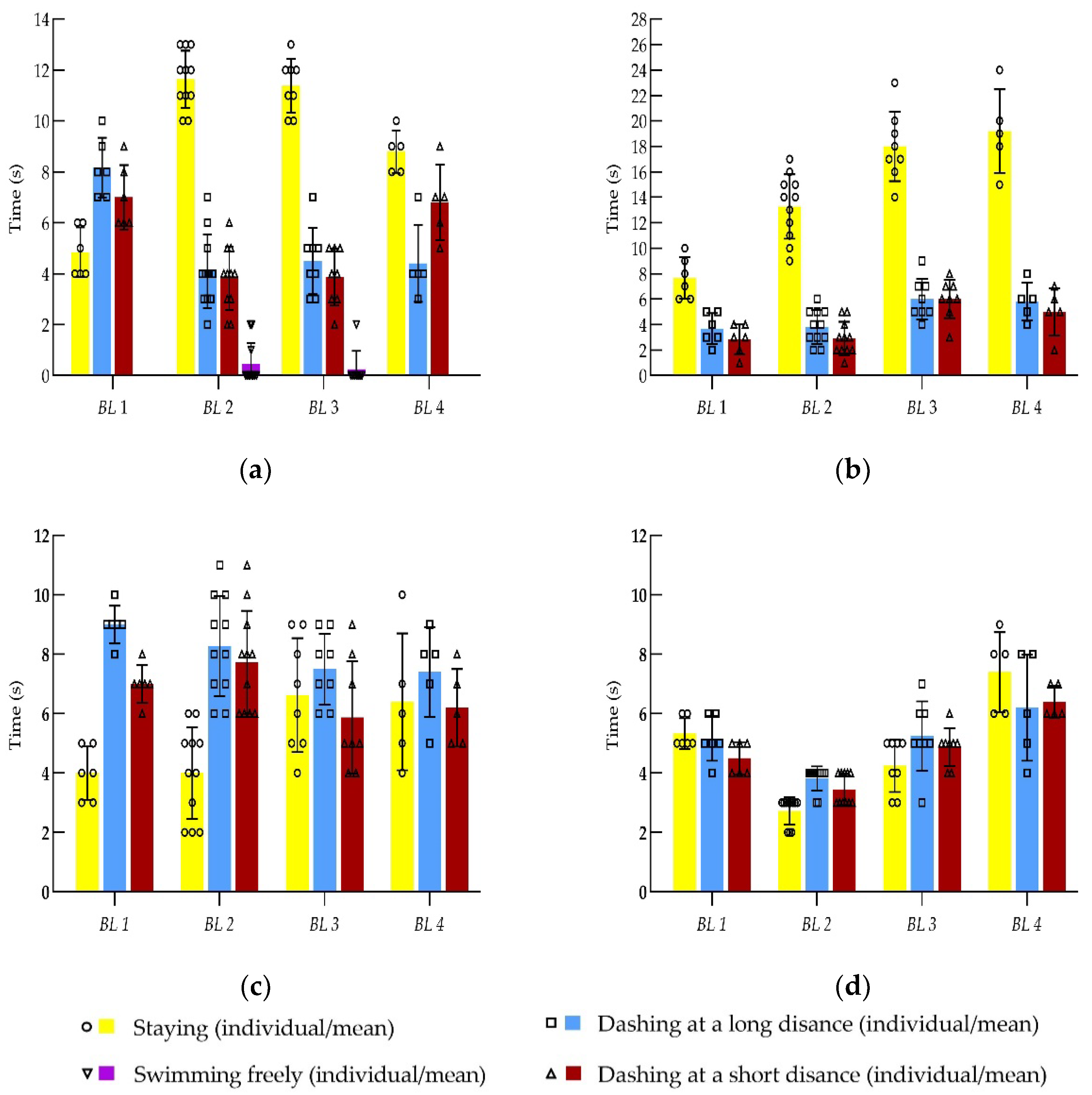
3.4. Fish Swimming Behaviors in Different Stages
4. Discussion
4.1. Comparison with Other Studies
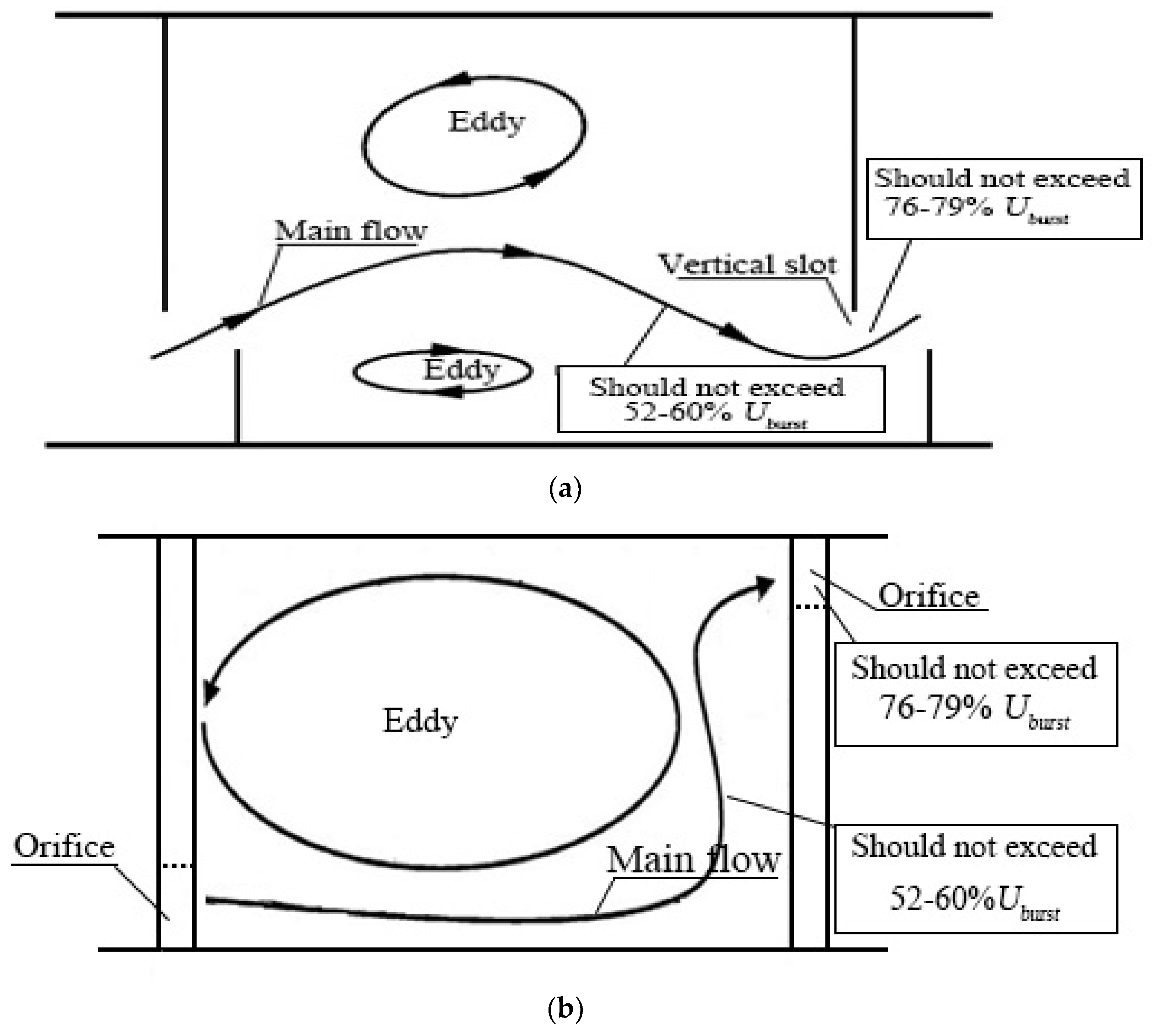
4.2. Implications for Fishway Hydraulic Design
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Unevenness Coefficient
Appendix B. Critical and Burst Swimming Speeds
References
- Wu, X.; Rao, J.; He, B. The history of the Chinese freshwater fisheries. In Cultivation of the Chinese Freshwater Fishes; Science Press: Beijing, China, 1992; pp. 5–29. [Google Scholar]
- Newbold, L.R.; Shi, X.T.; Hou, Y.; Han, D.; Kemp, P.S. Swimming performance and behavior of bighead carp (Hypophthalmichthys nobilis) application to fish passage and exclusion criteria. Ecol. Eng. 2016, 95, 690–698. [Google Scholar] [CrossRef]
- Shi, X.T.; Kynard, B.; Liu, D.F.; Qiao, Y.; Chen, Q.W. Development of fish passage in China. Fisheries 2015, 40, 161–169. [Google Scholar] [CrossRef]
- Mu, X.P.; Zhen, W.Y.; Li, X.; Cao, P.; Gong, L.; Xu, F.R. A study of the impact of different flow velocities and light colors at the entrance of a fish collection system on the upstream swimming behavior of juvenile grass carp. Water 2019, 11, 322. [Google Scholar] [CrossRef]
- Tan, J.; Tao, L.; Gao, Z.; Dai, H.; Shi, X.T. Modeling fish movement trajectories in relation to hydraulic response relationships in an experimental fishway. Water 2018, 10, 1511. [Google Scholar] [CrossRef]
- Laine, A.; Kamula, R.; Hooli, J. Fish and lamprey passage in a combined Denil and vertical slot fishway. Fish. Manag. Ecol. 1998, 5, 31–44. [Google Scholar] [CrossRef]
- Zhang, J.M.; Wu, Z.Q.; Hu, M.L. Resource status of four major Chinese carps in the Xiajiang reach of Ganjiang river. J. Hydroecol. 2010, 3, 34–37. [Google Scholar]
- Ministry of Environmental Protection of the People’s Republic of China. Bulletin on Ecological and Environmental Monitoring of the Three Gorges Project of the Yangtze River (BEEMTGP); Ministry of Environmental Protection of the People’s Republic of China: Beijing, China, 2013. Available online: http://www.cnemc.cn/jcbg/zjsxgcstyhjjcbg/ (accessed on 12 October 2019).
- Castro-Santos, T.; Cotel, A.J.; Webb, P.W. Fishway evaluations for better bioengineering: An integrative approach. In American Fisheries Society Symposium; American Fisheries Society: Bethesda, MD, USA, 2009; Volume 69, pp. 557–575. [Google Scholar]
- Chen, K.Q.; Tao, J.; Chang, Z.N.; Cao, X.H.; Ge, H.F. Difficulties and prospects of fishways in china: An overview of the construction status and operation practice since 2000. Ecol. Eng. 2014, 70, 82–91. [Google Scholar] [CrossRef]
- Castro-Santos, T.; Sanz-Ronda, F.J.; Ruiz-Legazpi, J. Breaking the speed limit—Comparative sprinting performance of brook trout (Salvelinus fontinalis) and brown trout (Salmo trutta). Can. J. Fish. Aquat. Sci. 2012, 70, 280–293. [Google Scholar] [CrossRef]
- Kim, J.H.; Yoon, J.D.; Baek, S.H.; Park, S.H.; Lee, J.W.; Lee, J.A.; Jang, M.H. An efficiency analysis of a nature-like fishway for freshwater fish ascending a large Korean river. Water 2016, 8, 3. [Google Scholar] [CrossRef]
- Bermudez, M.; Rico, A.; Rodriguez, A.; Pena, L.; Rabunal, J.R.; Puertas, J.; Castillo, M. FishPath: Computer application for design of vertical slot fishways. Ing. Del Agua 2015, 19, 179–191. [Google Scholar]
- Sanz-Ronda, F.J.; Ruiz-Legazpi, J.; Bravo-Córdoba, F.J.; Makrakis, S.; Castro-Santos, T. Sprinting performance of two Iberian fish: Luciobarbus bocagei and Pseudochondrostoma duriense in an open channel flume. Ecol. Eng. 2015, 83, 61–70. [Google Scholar] [CrossRef]
- Hammer, C. Fatigue and exercise tests with fish. Comp. Biochem. Physiol. Part A Physiol. 1995, 112, 1–20. [Google Scholar] [CrossRef]
- Brett, J.R. The respiratory metabolism and swimming performance of young sockeye salmon (Oncorhynchus nerka). J. Fish. Board Can. 1964, 21, 1183–1226. [Google Scholar] [CrossRef]
- Beamish, F.W.H. Swimming capacity. Fish Physiol. 1978, 7, 101–187. [Google Scholar]
- Mateus, C.S.; Quintella, B.R.; Almeida, P.R. The critical swimming speed of Iberian barbel Barbus bocagei in relation to size and sex. J. Fish Biol. 2008, 73, 1783–1789. [Google Scholar] [CrossRef]
- Adams, S.R.; Adams, G.L.; Parsons, G.R. Critical swimming speed and behavior of juvenile shovelnose sturgeon and pallid sturgeon (Scaphirhynchus platorynchus). Trans. Am. Fish. Soc. 2003, 132, 392–397. [Google Scholar] [CrossRef]
- Fang, M.; Cai, L.; Gao, Y.; Tu, Z.Y.; Wang, C.F.; Huang, Y.P. Effect of temperature on swimming capability and oxygen consumption of juvenile Hypophthalmichthys molitrix. J. Hydroecol. 2013, 34, 49–53. [Google Scholar]
- Xian, X.M.; Cao, Z.D.; Fu, S.J. The comparison of critical swimming speed and endurance at high speed of four species of juvenile fish. J. Chongqing Norm. Univ. 2010, 27, 16–20. [Google Scholar]
- Cai, L.; Tu, Z.Y.; Yuan, X.; Liu, G.Y.; Liu, D.F.; Shi, X.T.; Huang, Y.P. Swimming capability and swimming behaviour of juvenile Aristichthys nobilis. Resour. Environ. Yangtze Basin 2013, 21, 89–95. [Google Scholar]
- Xiong, F.; Wang, C.J.; Liu, D.F.; Kou, F.L.; Mo, W.J.; Liu, W.; Shi, X.T. Comparative study of burst swimming speed of black carp, grass carp, silver carp and bighead carp from Songhua river. Ecol. Sci. 2014, 33, 339–343. [Google Scholar]
- Li, C.W.; Bo, L.U.; Chen, T.; Liu, L.; Zhu, H.F.; Liu, D.F.; Wang, X.; Shi, X.T. The burst-coast behavior in escape response of black carp. Chin. J. Zool. 2014, 49, 860–866. [Google Scholar]
- Niu, S.; Lu, B.; Luo, J.; He, L.; Niu, J.T.; Chen, T.; Liu, D.F.; Shi, X.T. Swimming performance of fast escapement of silver carp. Acta Hydrobiol. Sin. 2015, 2, 394–398. [Google Scholar]
- Lu, B.; Liu, W.; Liang, Y.; Chen, Q.; Huang, Y.; Pan, L.; Liu, D.F.; Shi, X.T. The burst-coast swimming behavior of grass carp (Ctenopharyngodon idellus) during fast-start. J. Fish. China 2014, 38, 829–834. [Google Scholar]
- Sanz-Ronda, F.J.; Bravo-Córdoba, F.J.; Fuentes-Pérez, J.F.; Castro-Santos, T. Ascent ability of brown trout, Salmo trutta, and two Iberian cyprinids− Iberian barbel, Luciobarbus bocagei, and northern straight-mouth nase, Pseudochondrostoma duriense—In a vertical slot fishway. Knowl. Manag. Aquat. Ecosyst. 2016, 10. [Google Scholar] [CrossRef]
- Katopodis, C. Developing a toolkit for fish passage, ecological flow management and fish habitat works. J. Hydraul. Res. 2005, 43, 451–467. [Google Scholar] [CrossRef]
- Hu, M.L. Characteristics of Water Level, Water Environment and Effects on Fish Communication and Migration in the Hukou of Poyang Lake; Nanchang University: Nanchang, China, 2009. [Google Scholar]
- He, G.; Fang, C.L.; Chen, W.J.; Fu, P.F.; Zhou, H.M.; Zhang, Y.P.; Wu, B.; Wang, S. Community structure and variation of migration fishes in Pingfeng area of Poyang lake channel. Hubei Agric. Sci. 2015, 54, 926–930. [Google Scholar]
- Santos, J.M.; Silva, A.T.; Katopodis, C.; Pinheiro, P.; Pinheiro, A.N.; Bochechas, J.; Ferreira, M.T. Ecohydraulics of pool-type fishways: Getting past the barriers. Ecol. Eng. 2012, 48, 38–50. [Google Scholar] [CrossRef]
- Plaut, I. Critical swimming speed: Its ecological relevance. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2001, 131, 41–50. [Google Scholar] [CrossRef]
- Blank, M.D.; Cahoon, J.; McMahon, T. Using three dimensional hydrodynamic modeling and fish swimming energetics to assess culverts as potential physical barriers to upstream fish movement. In Proceedings of the Transportation Research Board 88th Annual Meeting, Washington, DC, USA, 11–15 January 2009. [Google Scholar]
- Blake, R.W. Functional design and burst-and-coast swimming in fishes. Rev. Can. Zool. 1983, 61, 2491–2494. [Google Scholar] [CrossRef]
- Gong, L.; Wu, Y.H.; Baiyin, B.L.G.; Mu, X.P. Experimental study on swimming capability and swimming behavior of juvenile grass carp. J. China Inst. Water Resour. Hydropower Res. 2015, 13, 211–216. [Google Scholar]
- Cao, P.; Mu, X.P.; Baiyin, B.L.G.; Wang, X.Y.; Chen, Y.F. Study on swimming behavior of juvenile grass carp for the fish channel hydraulic design. J. Hydraul. Eng. 2017, 48, 1456–1464. [Google Scholar]
- Bell, W.H.; Terhune, L.D.B. Water Tunnel Design for Fisheries Research; Fisheries Research Board of Canada, Biological Station: Ottawa, ON, Canada, 1970.
- Xu, G.F.; Yin, J.S.; Ha, Y.; Ma, B.; Mou, Z.B. Effects of different acclimation conditions on sustainable swimming performance of juvenile lenok, Brachymystax lenok (Pallas). Acta Ecol. Sin. 2015, 35, 1938–1946. [Google Scholar]
- Farlinger, S.; Beamish, F.W.H. Effects of time and velocity increments on the critical swimming speed of largemouth bass (Micropterus salmoides). Trans. Am. Fish. Soc. 1977, 106, 436–439. [Google Scholar] [CrossRef]
- Kolok, A.S. Interindividual variation in the prolonged locomotor performance of ectothermic vertebrates: A comparison of fish and herpetofaunal methodologies and a brief review of the recent fish literature. Can. J. Fish. Aquat. Sci. 1999, 56, 700–710. [Google Scholar] [CrossRef]
- Bestgen, K.R.; Mefford, B.; Bundy, J.M.; Walford, C.D.; Compton, R.I. Swimming performance and fishway model passage success of Rio Grande silvery minnow. Trans. Am. Fish. Soc. 2010, 139, 433–448. [Google Scholar] [CrossRef]
- Kern, P.; Cramp, R.L.; Gordos, M.A.; Watson, J.R.; Franklin, C.E. Measuring Ucrit and endurance: Equipment choice influences estimates of fish swimming performance. J. Fish Biol. 2018, 92, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Farrell, A.P. Comparisons of swimming performance in rainbow trout using constant acceleration and critical swimming speed tests. J. Fish Biol. 2008, 72, 693–710. [Google Scholar] [CrossRef]
- Lyon, J.P.; Ryan, T.J.; Scroggie, M.P. Effects of temperature on the fast-start swimming performance of an Australian freshwater fish. Ecol. Freshw. Fish 2008, 17, 184–188. [Google Scholar] [CrossRef]
- Kieffer, J.D. Limits to exhaustive exercise in fish. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2000, 126, 161–179. [Google Scholar] [CrossRef]
- Randall, D.; Brauner, C. Effects of environmental factors on exercise in fish. J. Exp. Biol. 1991, 160, 113–126. [Google Scholar]
- Rome, L.C.; Alexander, R.M. The influence of temperature on muscle velocity and sustained performance in swimming carp. J. Exp. Biol. 1990, 154, 163–178. [Google Scholar] [PubMed]
- Gong, L.; Wu, Y.H.; Baiyin, B.L.G.; Mu, X.P.; Jin, Z.J. Study on impact of water flow at fishway entrance on migratory behavior of juvenile grass carp. Water Resour. Hydropower Eng. 2016, 106, 89–93. [Google Scholar]
- Fang, M.; Cai, L.; Gao, Y.; Tu, Z.Y.; Wang, C.F.; Huang, Y.P. Effect of activity cost on swimming capability of grass carp (Ctenopharyngodon idellus). Resour. Environ. Yangtze Basin 2014, 23, 816–820. [Google Scholar]
- Silva, A.T.; Katopodis, C.; Santos, J.M.; Ferreira, M.T.; Pinheiro, A.N. Cyprinid swimming behaviour in response to turbulent flow. Ecol. Eng. 2012, 44, 314–328. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | BL (cm) | W (g) | Uburst (m/s) | Tested Fish Number |
|---|---|---|---|---|
| BL 1 | 6.00–8.50 | 3.80–12.21 | <0.90 m/s | 6 |
| BL 2 | 8.71–11.01 | 10.40–15.48 | 0.90–1.10 m/s | 11 |
| BL 3 | 11.30–12.30 | 15.00–20.71 | 1.10–1.20 m/s | 8 |
| BL 4 | 12.70–15.00 | 22.17–35.00 | >1.20 m/s | 5 |
| Classification | BL 1 | BL 2 | ||
| Uburst (m/s) | Uburst (%) | Uburst (m/s) | Uburst (%) | |
| Stage 1 | 0.56 ± 0.08 | 58 ± 10.50 | 0.58 ± 0.10 | 55.70 ± 8.50 |
| Stage 2 | 0.76 ± 0.08 | 78 ± 11 | 0.79 ± 0.11 | 75.80 ± 9 |
| Stage 3 | 0.84 ± 0.12 | 90.50 ± 1 | 0.97 ± 0.13 | 95.8 ± 2.6 |
| Stage 4 | 0.91 ± 0.10 | 100 | 1.03 ± 0.13 | 100 |
| Classification | BL 3 | BL 4 | ||
| Uburst (m/s) | Uburst (%) | Uburst (m/s) | Uburst (%) | |
| Stage 1 | 0.64 ± 0.07 | 60 ± 8 | 0.69 ± 0.01 | 52 ± 8 |
| Stage 2 | 0.84 ± 0.07 | 78.40 ± 9 | 0.92 ± 0.07 | 79 ± 6 |
| Stage 3 | 1.09 ± 0.06 | 95.70 ± 2.80 | 1.18 ± 0.13 | 96 ± 3 |
| Stage 4 | 1.11 ± 0.07 | 100 | 1.31 ± 0.04 | 100 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mu, X.; Cao, P.; Gong, L.; Baiyin, B.; Li, X. A Classification Method for Fish Swimming Behaviors under Incremental Water Velocity for Fishway Hydraulic Design. Water 2019, 11, 2131. https://doi.org/10.3390/w11102131
Mu X, Cao P, Gong L, Baiyin B, Li X. A Classification Method for Fish Swimming Behaviors under Incremental Water Velocity for Fishway Hydraulic Design. Water. 2019; 11(10):2131. https://doi.org/10.3390/w11102131
Chicago/Turabian StyleMu, Xiangpeng, Ping Cao, Li Gong, Baoligao Baiyin, and Xiang Li. 2019. "A Classification Method for Fish Swimming Behaviors under Incremental Water Velocity for Fishway Hydraulic Design" Water 11, no. 10: 2131. https://doi.org/10.3390/w11102131
APA StyleMu, X., Cao, P., Gong, L., Baiyin, B., & Li, X. (2019). A Classification Method for Fish Swimming Behaviors under Incremental Water Velocity for Fishway Hydraulic Design. Water, 11(10), 2131. https://doi.org/10.3390/w11102131