Aquatic Angiosperm Transplantation: A Tool for Environmental Management and Restoring in Transitional Water Systems

 ,
, 
 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
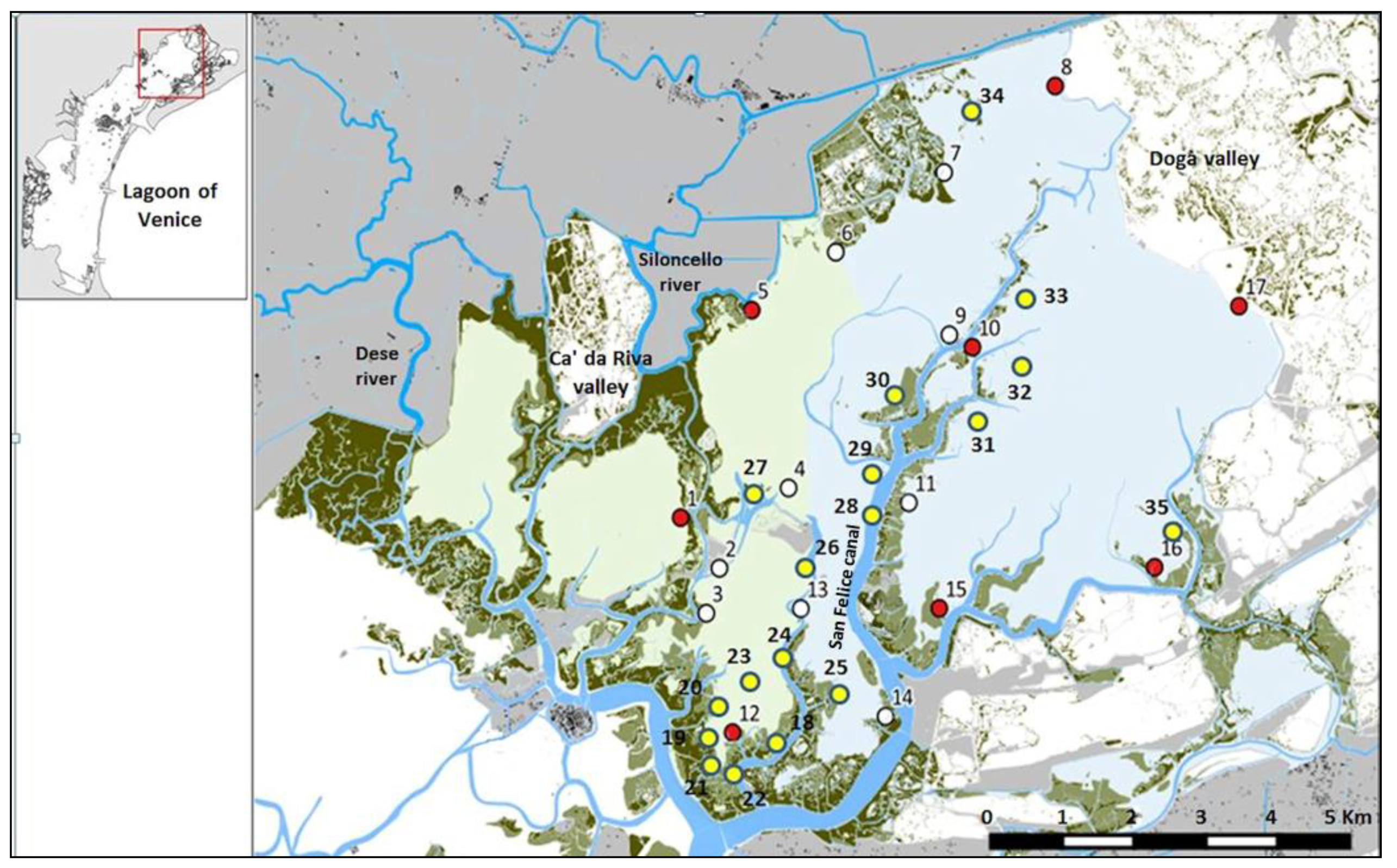
2.1. Study Area
2.2. Angiosperm Transplants
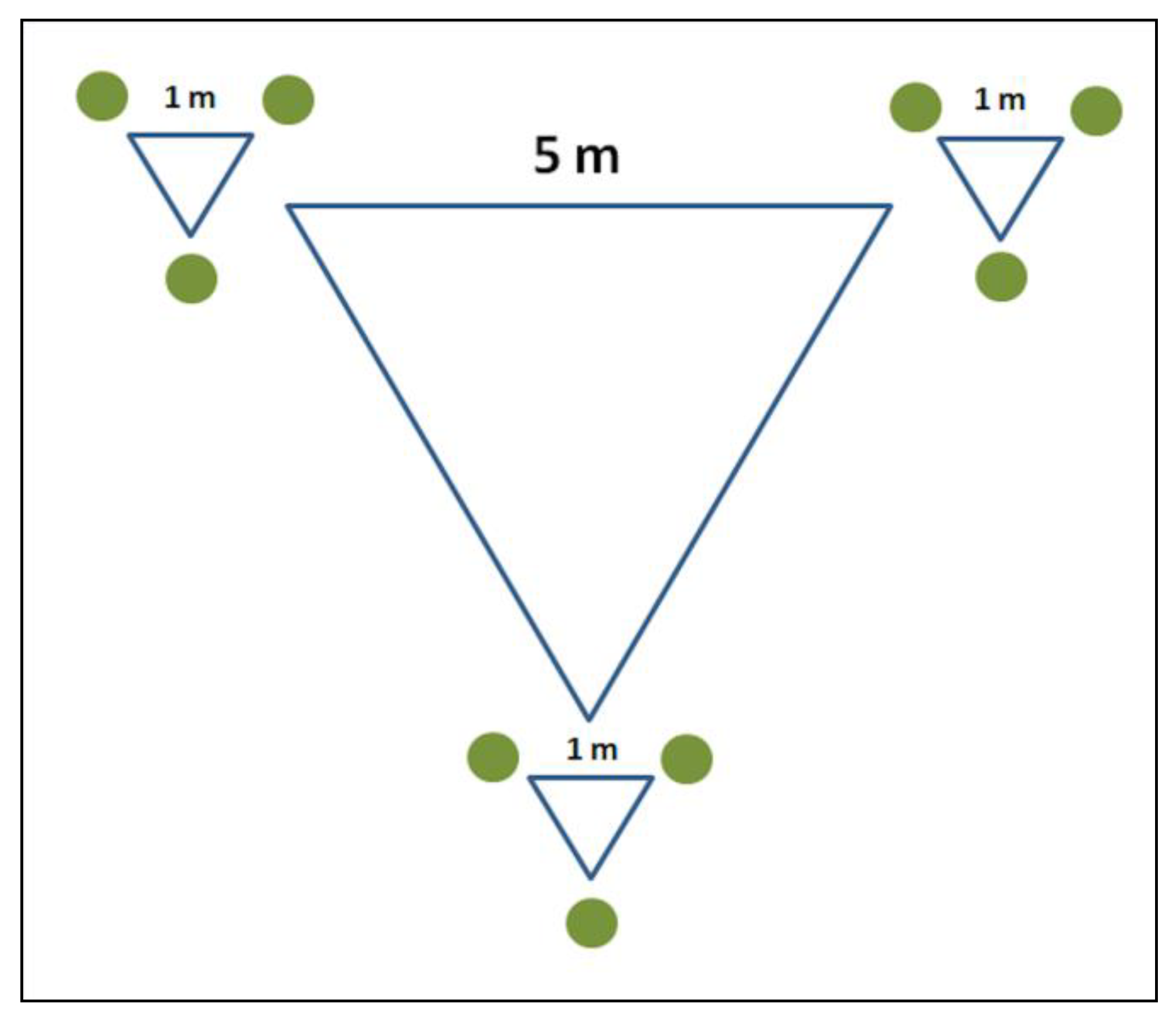
2.3. Angiosperm Monitoring
2.4. Physico-Chemical Parameter Determination
2.5. Statistical Analyses
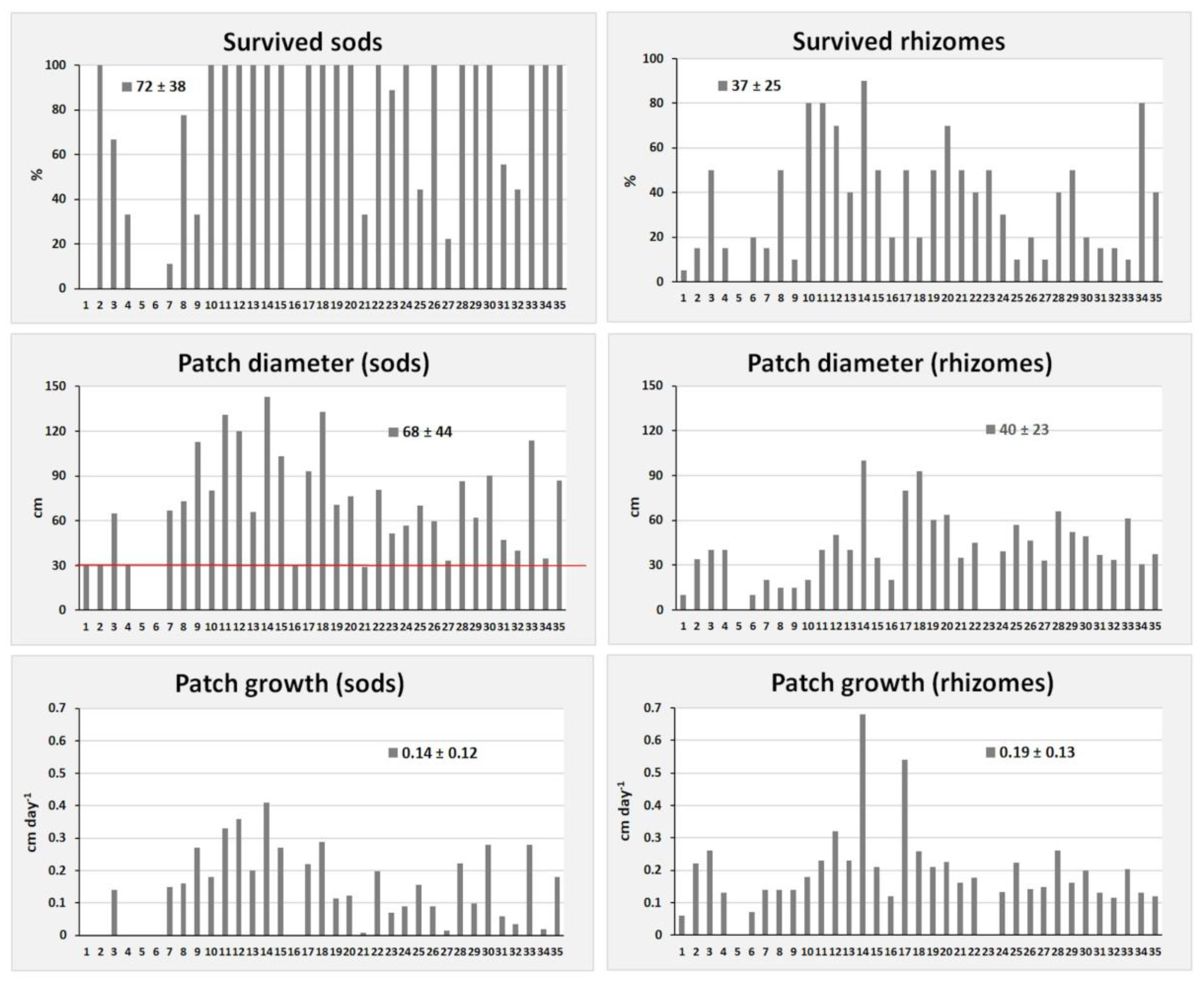
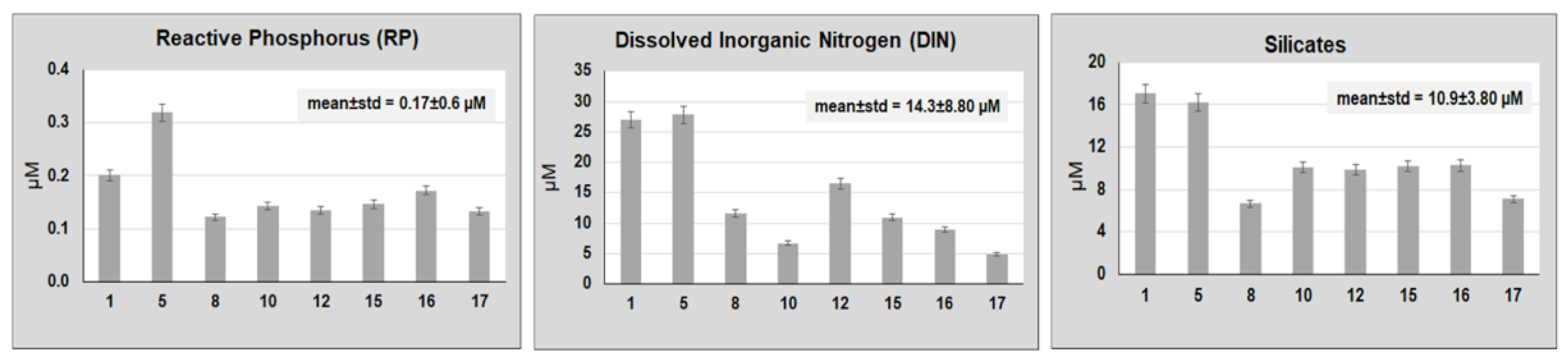
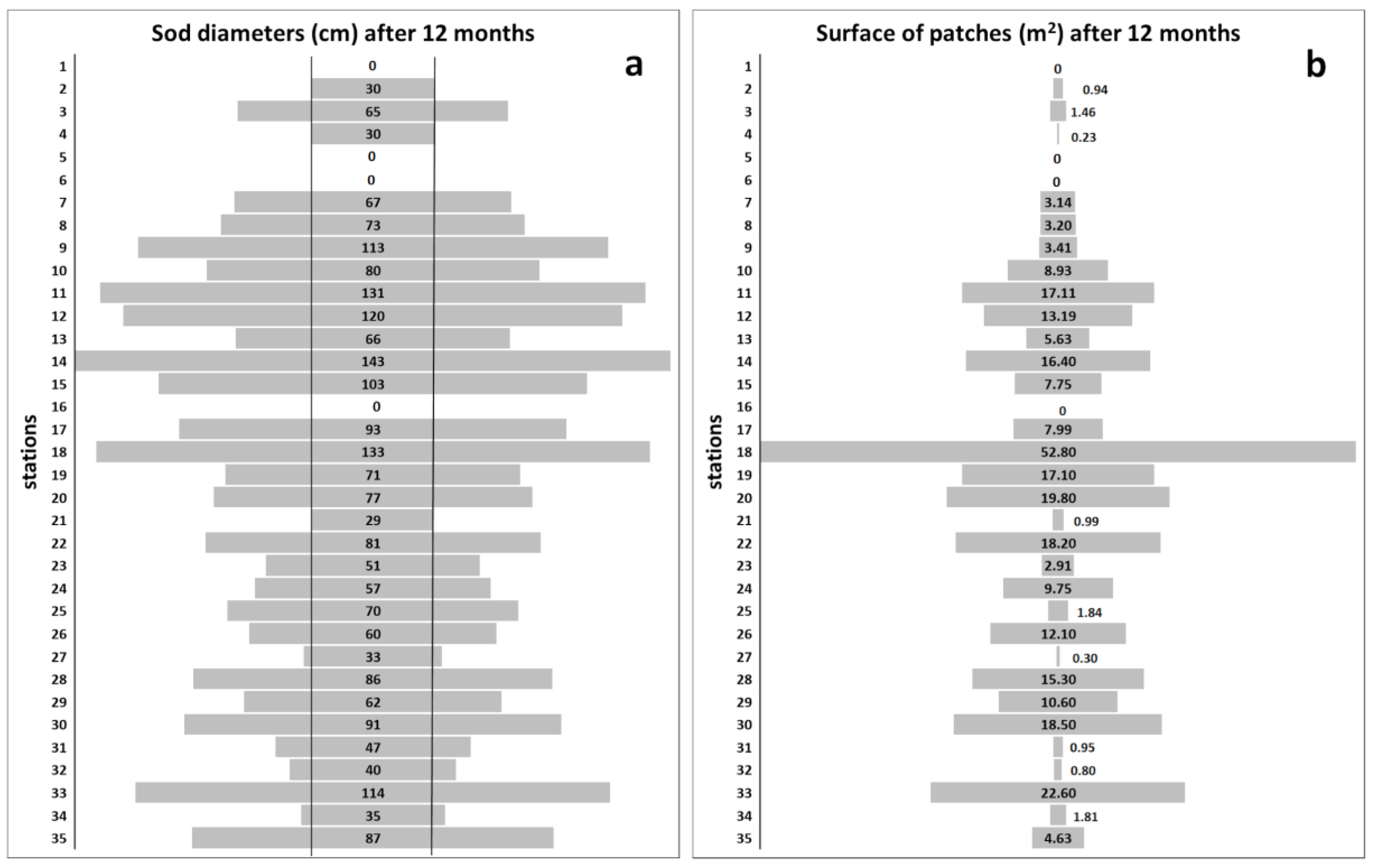
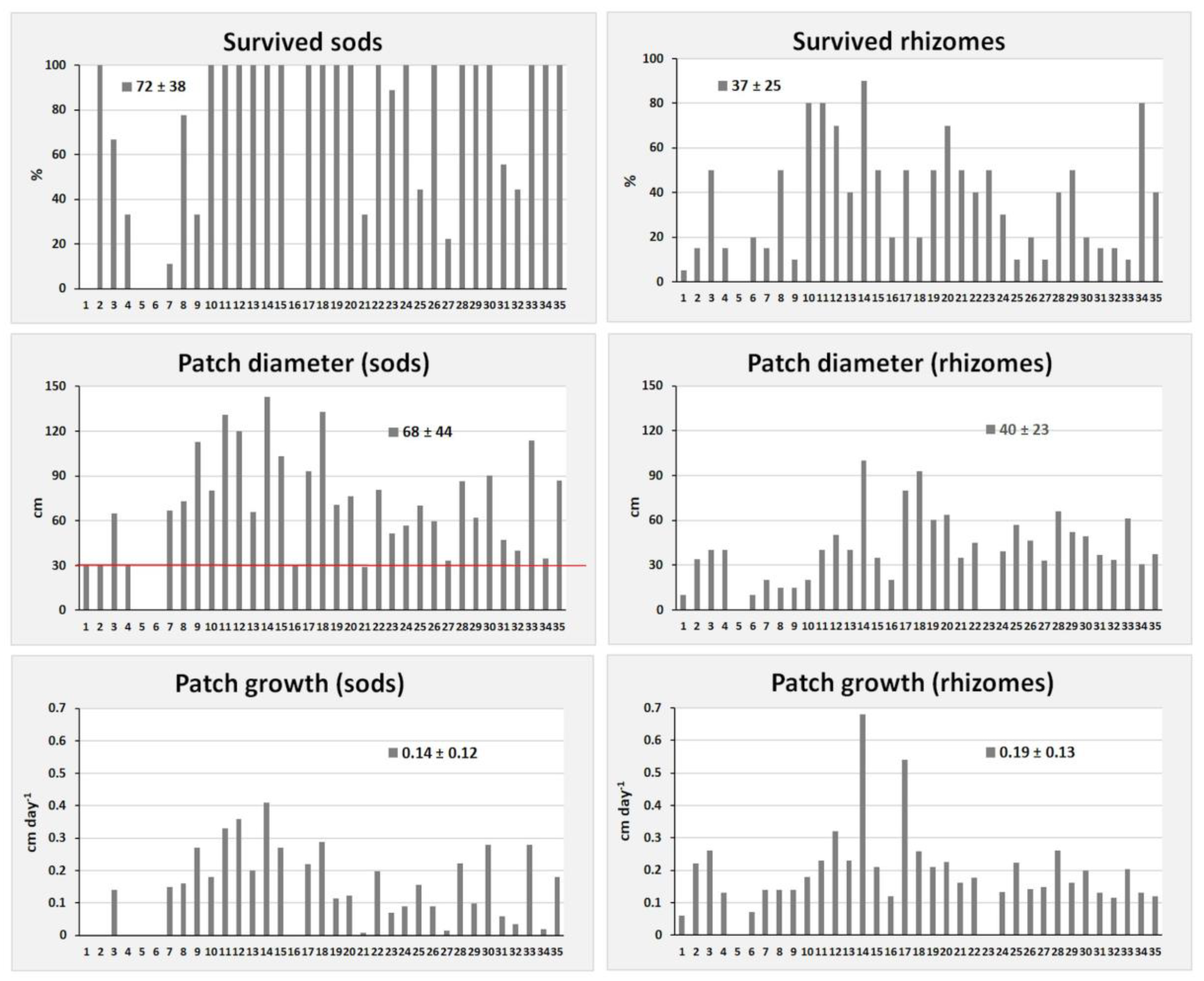
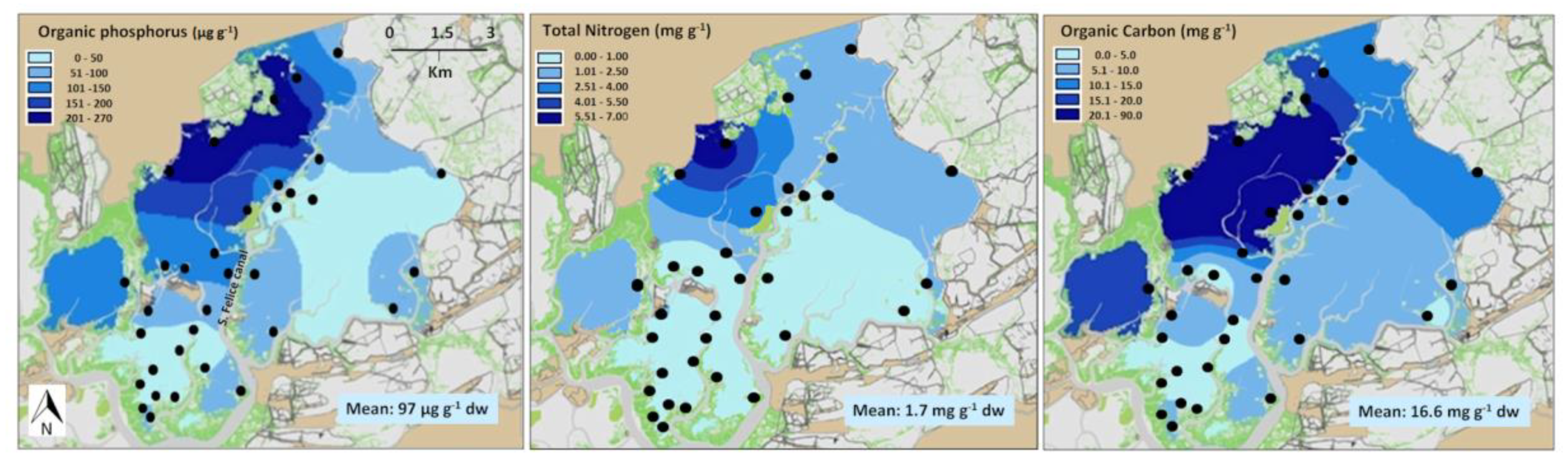
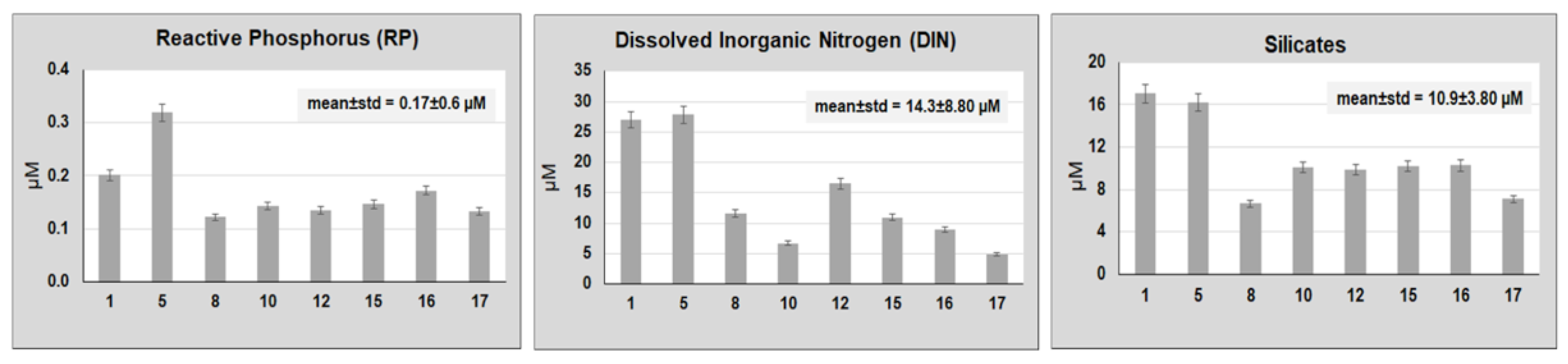
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Larkum, A.W.D.; McComb, A.J.; Shepherd, S.A. Biology of Seagrasses: A Treatise of the Biology of Seagrasses with Special Reference to Australian Region; Elsevier: Amsterdam, The Netherlands, 1989; p. 841. [Google Scholar]
- Ondiviela, B.; Losada, I.J.; Lara, J.L.; Maza, M.; Galván, C.; Bouma, T.J.; van Belzen, J.V. The role of seagrasses in coastal protection in a changing climate. Coast. Eng. 2014, 87, 158–168. [Google Scholar] [CrossRef]
- Duarte, C.M.; Losada, I.J.; Hendriks, I.E.; Mazarrasa, I.; Marba, N. The role of coastal plant communities for climate change mitigation and adaptation. Nat. Clim. Chang. 2013, 3, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.G.; Lawron, J.H.; Shachak, M. Positive and negative effects of organisms as physical ecosystem engineers. Ecology 1997, 78, 1946–1957. [Google Scholar] [CrossRef]
- Horinouchi, M. Horizontal gradient in fish assemblage structures in and around a seagrass habitat: Some implications for seagrass habitat conservation. Ichthyol. Res. 2009, 56, 109–125. [Google Scholar] [CrossRef]
- Jackson, E.L.; Attrill, M.J.; Jones, M.B. Habitat characteristics and spatial arrangement affecting the diversity of fish and decapod assemblages of seagrass (Zostera marina) beds around the coast of Jersey (English Channel). Estuar. Coast. Shelf Sci. 2006, 68, 421–432. [Google Scholar] [CrossRef]
- Sfriso, A.; Buosi, A.; Facca, C.; Sfriso, A.A. Role of environmental factors in affecting macrophyte dominance in transitional environments: The Italian Lagoons as a study case. Mar. Ecol. 2017, 38, e12414. [Google Scholar] [CrossRef]
- Sfriso, A.; Pavoni, B.; Marcomini, A.; Orio, A.A. Macroalgae, nutrient cycles and pollutants in the lagoon of Venice. Estuaries 1992, 15, 517–528. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C. Distribution and production of macrophytes in the lagoon of Venice Comparison of actual and past abundance. Hydrobiologia 2007, 577, 71–85. [Google Scholar]
- Sfriso, A.; Facca, C.; Bon, D.; Giovannone, F.; Buosi, A. Using phytoplankton and macrophytes to assess the trophic and ecological status of some Italian transitional systems. Cont. Shelf Res. 2014, 81, 88–98. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Bon, D.; Buosi, A. Macrophytes and ecological status assessment in the Po delta transitional systems, Adriatic Sea (Italy). Application of Macrophyte Quality Index (MaQI). Acta Adriat. 2016, 57, 209–226. [Google Scholar]
- Sfriso, A.; Facca, C.; Marcomini, A. Sedimentation rates and erosion processes in the lagoon of Venice. Environ. Int. 2005, 31, 983–992. [Google Scholar]
- Sfriso, A.; Buosi, A.; Mistri, M.; Munari, C.; Franzoi, P.; Sfriso, A.A. Long-term changes of the trophic status in transitional ecosystems of the northern Adriatic Sea, key parameters and future expectations: The lagoon of Venice as a study case. In Italian Long-Term Ecological Research for Understanding Ecosystem Diversity and Functioning. Case Studies from Aquatic, Terrestrial and Transitional Domains. Nat. Conserv. 2019, 34, 247–272. [Google Scholar]
- Available online: http://www.arpa.veneto.it/dati-ambientali/open-data/file-e-allegati/2017 (accessed on 10 July 2019).
- Sfriso, A.; Facca, C.; Bonometto, A.; Boscolo Brusà, R. Compliance of the Macrophyte Quality Index (MaQI) with the WFD (2000/60/EC) and ecological status assessment in transitional areas: The Venice lagoon as study case. Ecol. Indic. 2014, 46, 536–547. [Google Scholar] [CrossRef]
- ISPRA. Manuale per la Classificazione Dell’elemento di Qualità Biologica “Fauna Ittica” Nelle Lagune Costiere Italiane Applicazione Dell’indice Nazionale HFBI (Habitat Fish Bio-Indicator) ai Sensi del D.Lgs 152/2006; Manuali e Linee Guida 168/2017; ISPRA: Ispra, Italy, 2017. [Google Scholar]
- Muxika, I.; Borja, A.; Bald, J. Using historical data, expert judgement and multivariate analysis in assessing reference conditions and benthic ecological status, according to the European Water Framework Directive. Mar. Pollut. Bull. 2007, 55, 16–29. [Google Scholar] [CrossRef]
- Sfriso, A.A.; Sfriso, A. In situ biomass production of Gracilariaceae and Ulva rigida: The Venice Lagoon as study case. Bot. Mar. 2017, 60, 271–283. [Google Scholar] [CrossRef]
- Sfriso, A.; Curiel, D.; Rismondo, A. The Venice Lagoon. In Flora and Vegetation of the Italian Transitional Water Systems; Cecere, E., Petrocelli, A., Izzo, G., Sfriso, A., Eds.; CORILA: Venice, Italy, 2009; pp. 17–80. [Google Scholar]
- Consorzio Venezia Nuova, Cantieri Costruzioni Cementi, SELC. Interventions to the Lagoon Inlets for the Regulation of Tidal Flows. Lido Mouth: S. Nicolò-Treporti. Lido-Arsenale Instrumentation and Control Systems. Morphological-Vegetation Restoration of the Area Occupied by the Artificial Island by Transplanting Seagrasses; Final Report; Consorzio Venezia Nuova: Venice, Italy, 2016. [Google Scholar]
- Venice Water Authority, THETIS, SELC. Compensation, Conservation and Environmental Redevelopment Plan for SCI and SPA of the Venice Lagoon. Seagrass Transplant Operations; Concessionaire Consorzio Venezia Nuova: Venice, Italy, 2010. (In Italian) [Google Scholar]
- CORILA—Consortium for the Coordination of Research Related to the Venice Lagoon System. Study B.6.72 B/13. Survey Activities for Monitoring the Effects Produced by the Construction of the Gates on the Lagoon inlets. Monitoring of Marine Phanerogam Transplants; Final Report; CORILA: Venice, Italy, 2019. [Google Scholar]
- Sfriso, A. Chlorophyta Multicellulari e Fanerogame Acquatiche. Ambienti di Transizione Italiani e Litorali Adiacenti. I Quaderni di ARPA; ARPA Emilia-Romagna: Bologna, Italy, 2010; p. 320. [Google Scholar]
- Dooley, F.D.; Wyllie-Echeverria, S.; Rothc, M.B.; Warda, P.D. Tolerance and response of Zostera marina seedlings to hydrogen sulfide. Aquat. Bot. 2013, 105, 7–10. [Google Scholar] [CrossRef]
- Höffle, H.; Thomsen, M.S.; Holmer, M. High mortality of Zostera marina under high temperature regimes but minor effects of the invasive macroalgae Gracilaria vermiculophylla. Estuar. Coast. Shelf Sci. 2011, 92, 35–46. [Google Scholar] [CrossRef]
- Nejrup, L.B.; Pedersen, M.F. Effects of salinity and water temperature on the ecological performance of Zostera marina. Aquat. Bot. 2008, 88, 239–246. [Google Scholar] [CrossRef]
- Mannino, A.M.; Menéndez, M.; Obrador, B.; Sfriso, A.; Triest, L. The genus Ruppia L. (Ruppiaceae) in the Mediterranean region: An overview. Aquat. Bot. 2015, 124, 1–9. [Google Scholar] [CrossRef]
- Available online: https://it.wikipedia.org/wiki/Zostera_marina (accessed on 10 July 2019).
- Sfriso, A. Ruppia maritima L. e Ruppia cirrhosa (Petagna) Grande (Helobiae, Spermatophyta) in laguna di Venezia. Lavori Soc. Veneziana Sci. Nat. 2008, 33, 41–46. [Google Scholar]
- Brodersen, E.K.; Koren, K.; Moßhammer, M.; Ralph, P.J.; Michael Kühl, M.; Santner, J. Seagrass-Mediated Phosphorus and Iron Solubilization in Tropical Sediments. Environ. Sci. Technol. 2017, 51, 14155–14163. [Google Scholar] [CrossRef] [PubMed]
- Sfriso, A.; Marcomini, A. Macrophyte production in a shallow coastal lagoon Part I. Coupling with physico-chemical parameters and nutrient concentrations in waters. Mar. Environ. Res. 1997, 44, 351–375. [Google Scholar] [CrossRef]
- Koch, M.S.; Erskine, J.M. Sulfide as a phytotoxin to the tropical seagrass Thalassia testudinum: Interactions with light, salinity and temperature. J. Exp. Mar. Biol. Ecol. 2001, 266, 81–95. [Google Scholar] [CrossRef]
- Pedersen, M.Ø.; Kristensen, E. Sensitivity of Ruppia maritima and Zostera marina to sulfide exposure around roots. J. Exp. Mar. Biol. Ecol. 2015, 468, 138–145. [Google Scholar] [CrossRef]
- Orfanidis, S.; Panayotidis, P.; Stamatis, N. Ecological evaluation of transitional and coastal waters: A marine benthic macrophytes-base model. Mediterr. Mar. Sci. 2001, 2, 45–65. [Google Scholar] [CrossRef]
- Sfriso, A.; Facca, C.; Ghetti, P.F. Rapid Quality Index (R-MaQI), based mainly on macrophyte associations, to assess the ecological status of Mediterranean transitional environments. Chem. Ecol. 2007, 23, 1–11. [Google Scholar] [CrossRef]
- Viaroli, P.; Bartoli, M.; Giordani, G.; Naldi, M.; Orfanidis, S.; Zaldívar, J.M. Community shifts, alternative stable states, biogeochemical controls and feed-backs in eutrophic coastal lagoons: A brief overview. Aquat. Conserv. 2008, 18, S105–S117. [Google Scholar] [CrossRef]
- Mcleod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Björk, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef]
- Duarte, C.M.; Sintes, T.S.; Marbà, N. Assessing the CO2 capture potential of seagrass restoration projects. J. Appl. Ecol. 2013, 50, 1341–1349. [Google Scholar] [CrossRef]
- Scapin, L.; Zucchetta, M.; Facca, C.; Sfriso, A.; Franzoi, P. Using fish assemblage to identify success criteria for seagrass habitat restoration. Web Ecol. 2016, 16, 33–36. [Google Scholar] [CrossRef] [Green Version]
- Scapin, L.; Zucchetta, M.; Sfriso, A.; Franzoi, P. Local habitat and Seascape Structure Influence Seagrass Fish Assemblages in the Venice Lagoon: The Importance of Conservation at Multiple Spatial Scales. Estuar. Coasts 2018, 41, 2410–2425. [Google Scholar] [CrossRef]
- Scapin, L.; Zucchetta, M.; Sfriso, A.; Franzoi, P. Predicting the response of nekton assemblages to seagrass transplantations in the Venice lagoon: An approach to assess ecological restoration. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 849–864. [Google Scholar] [CrossRef]



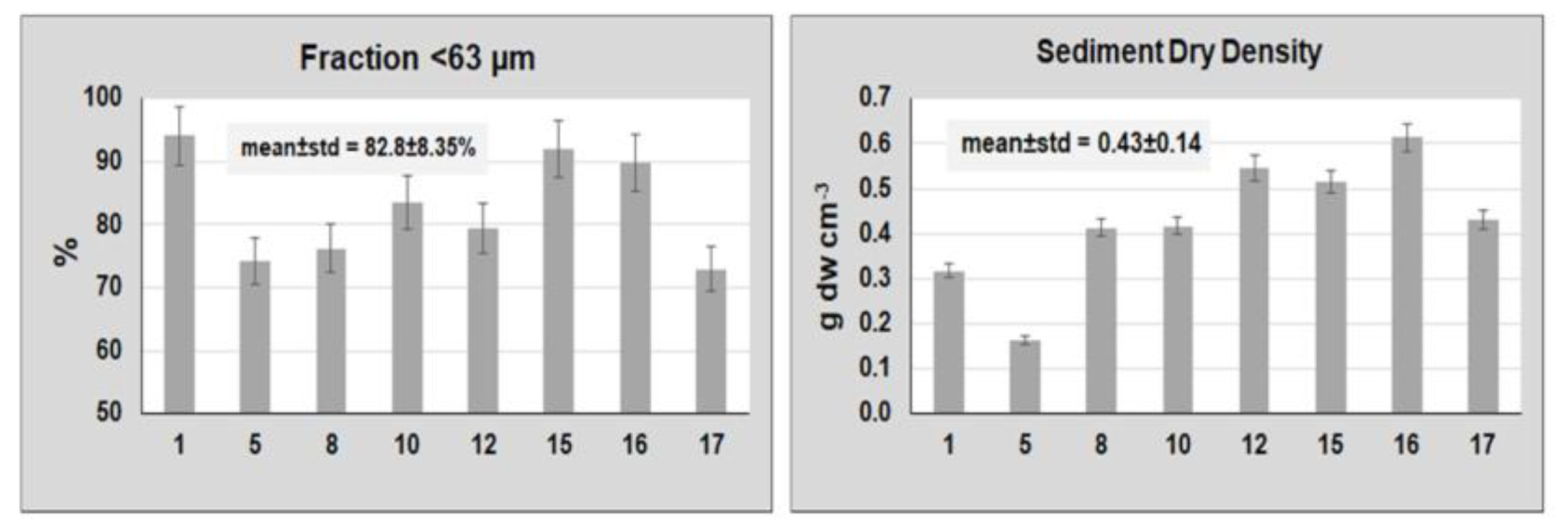
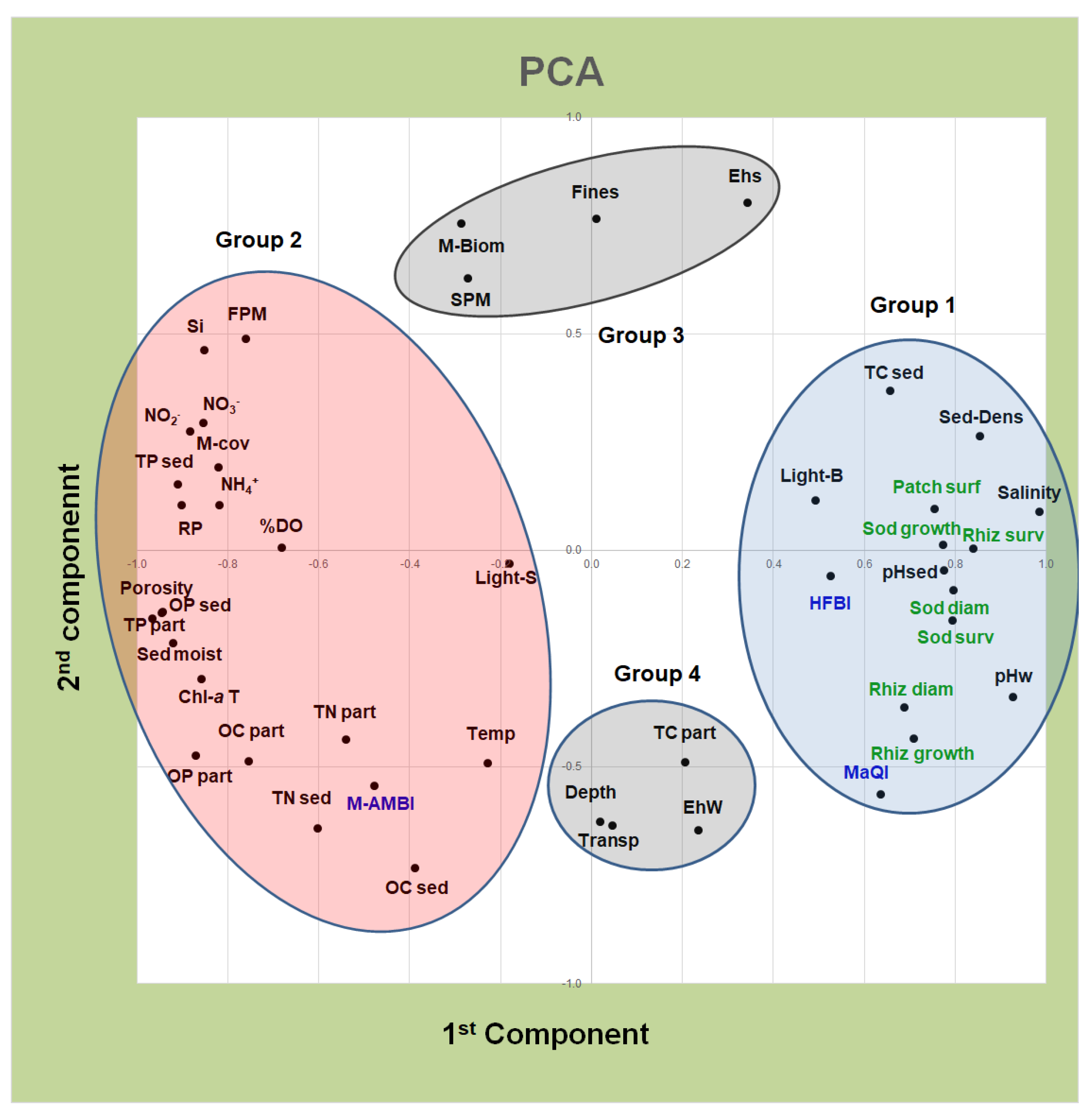

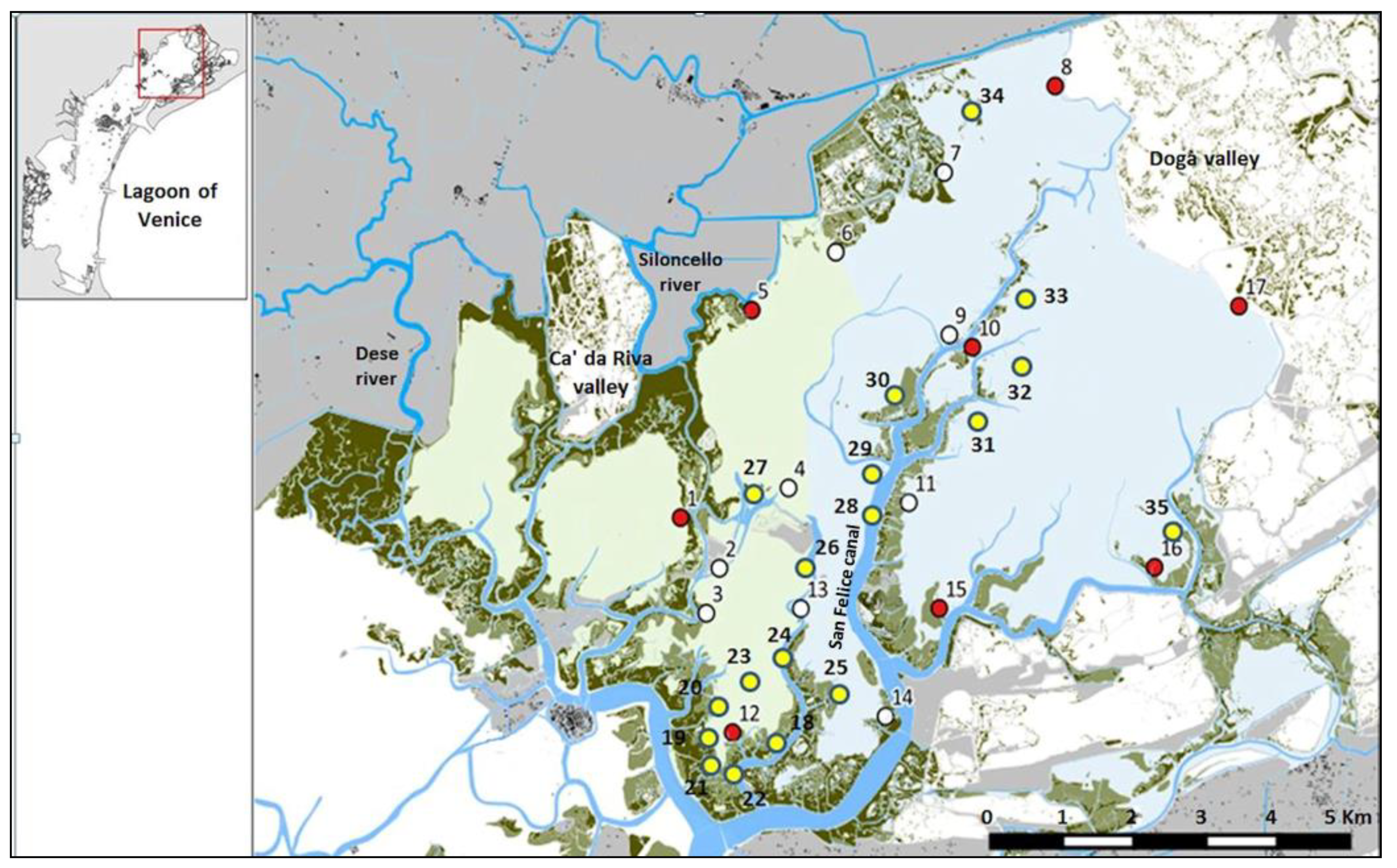{kind=link}
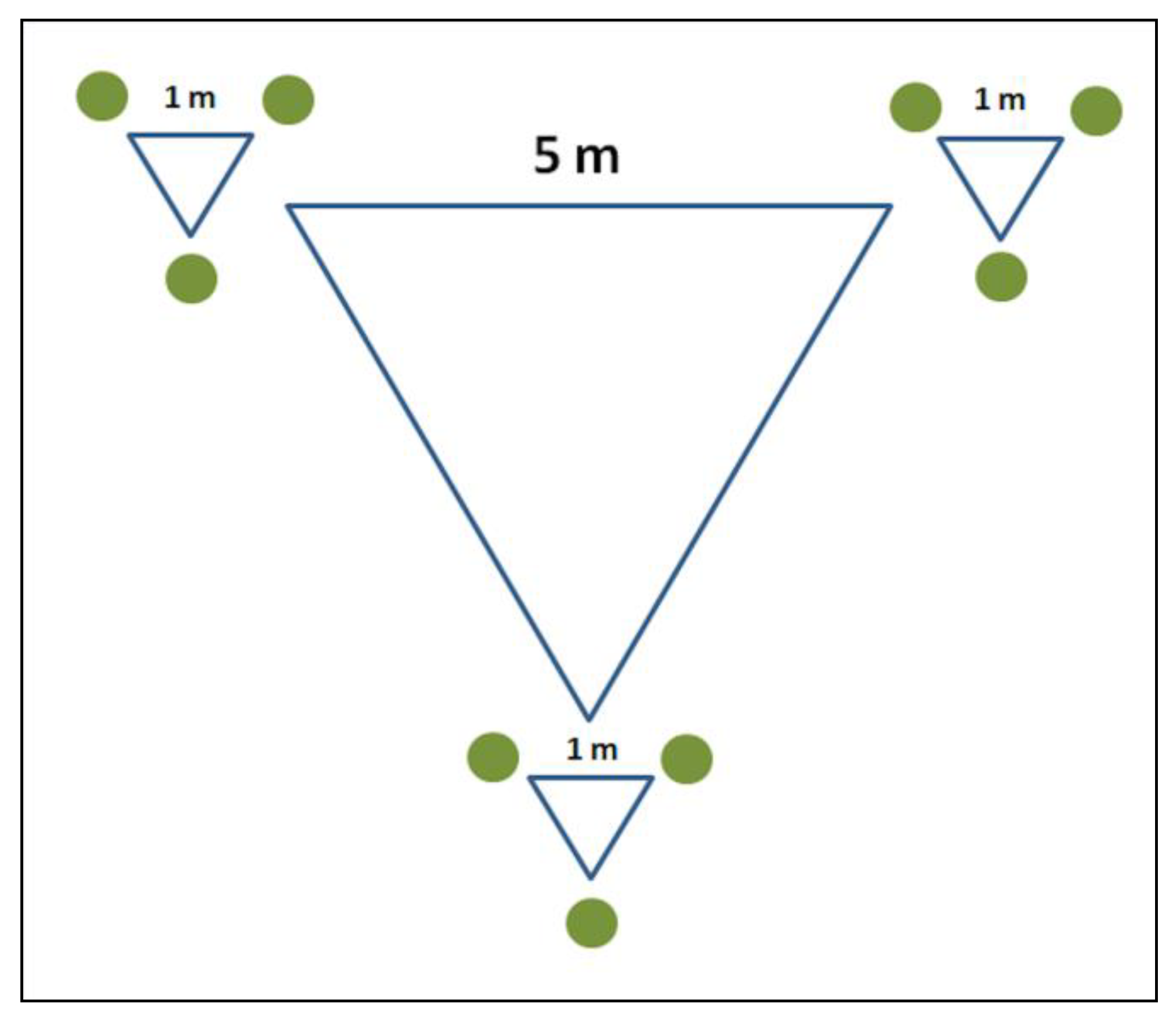{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
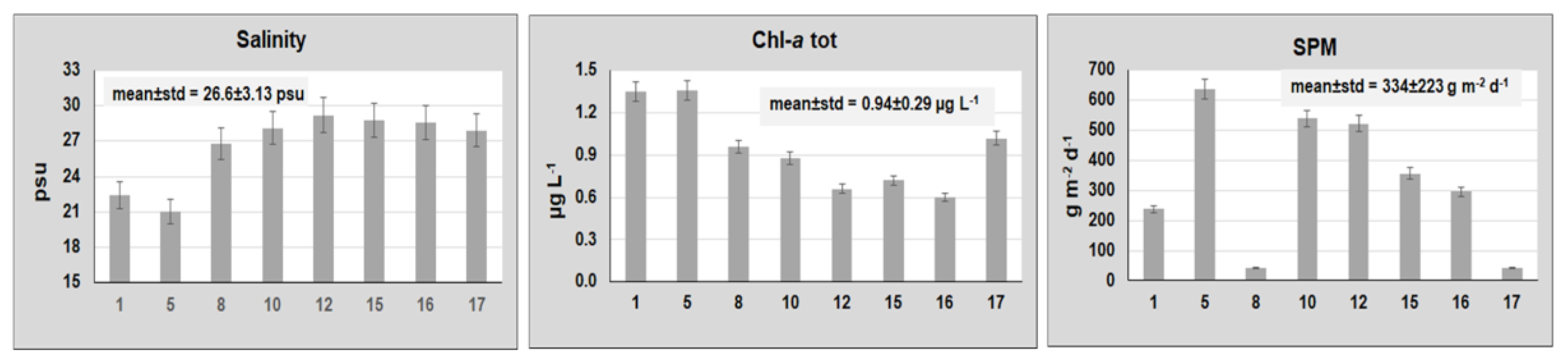{kind=link}
{kind=link}
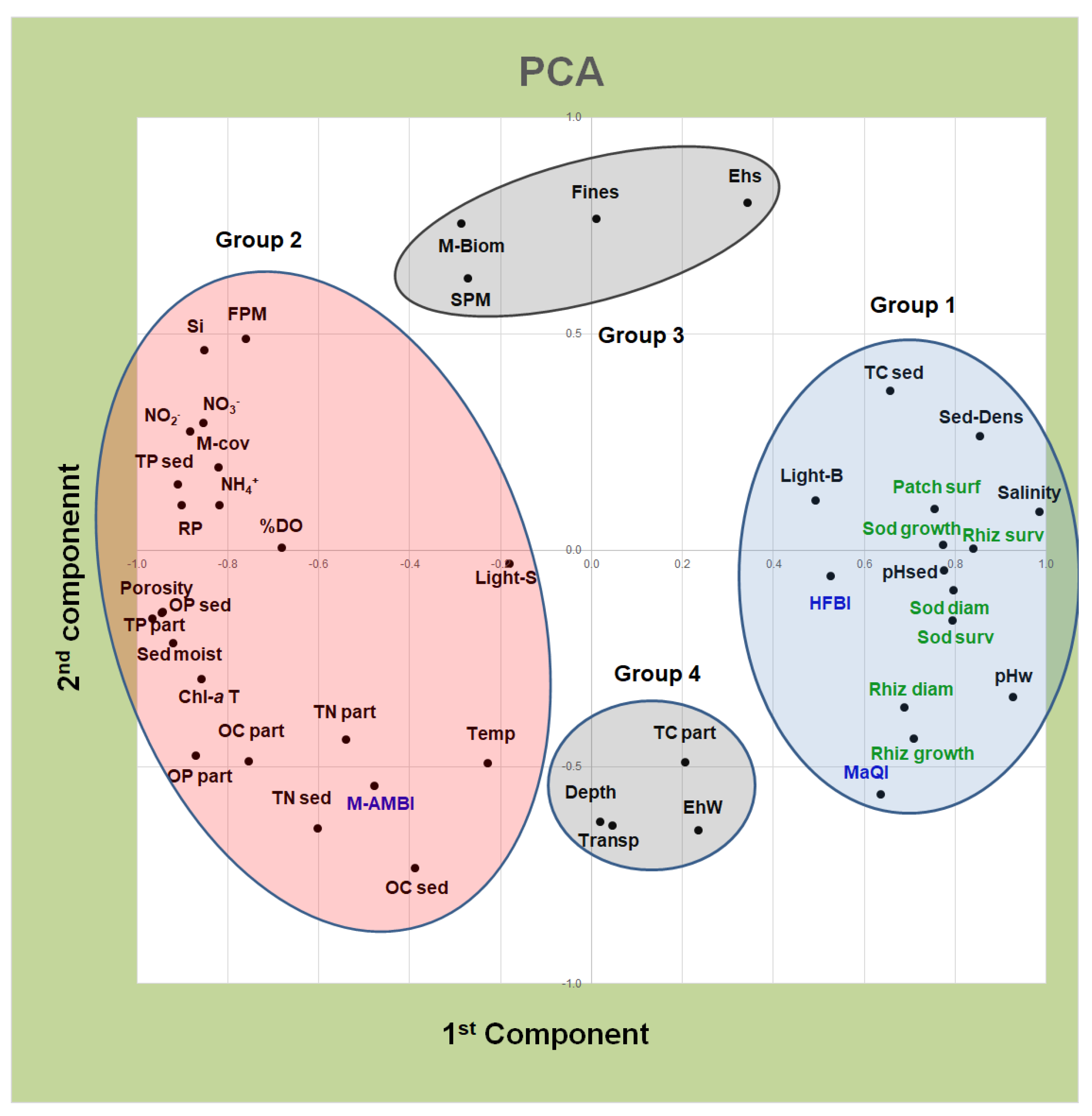{kind=link}
| Spearman’s Non Parametric Coefficients | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temp | Depth | Transp | Salin | %DO | pHw | Ehw | pHs | Ehs | Light-S | Light-B | FPM | |
| Survived sods | 0.03 | −0.21 | −0.09 | 0.63 | −0.33 | 0.43 | 0.18 | 0.30 | 0.39 | −0.34 | 0.48 | −0.31 |
| Sod diameter | −0.29 | −0.27 | −0.17 | 0.73 | −0.54 | 0.44 | 0.00 | 0.41 | 0.46 | −0.56 | 0.54 | −0.29 |
| Sod growth | −0.29 | −0.27 | −0.17 | 0.73 | −0.54 | 0.44 | 0.00 | 0.41 | 0.46 | −0.56 | 0.54 | −0.29 |
| Survived rhizomes | 0.07 | −0.39 | −0.29 | 0.66 | −0.32 | 0.42 | 0.44 | 0.51 | 0.59 | 0.00 | 0.71 | −0.27 |
| Rhizome diameter | −0.14 | −0.06 | 0.11 | 0.75 | −0.71 | 0.69 | 0.02 | 0.34 | 0.20 | −0.62 | 0.24 | −0.67 |
| Rhizome growth | −0.07 | −0.12 | 0.05 | 0.67 | −0.57 | 0.69 | 0.19 | 0.36 | 0.21 | −0.50 | 0.40 | −0.60 |
| Patch surface | 0.00 | −0.32 | −0.22 | 0.63 | −0.34 | 0.39 | 0.29 | 0.37 | 0.51 | −0.22 | 0.63 | −0.20 |
| MaQI | 0.34 | 0.40 | 0.56 | 0.27 | −0.38 | 0.87 | 0.39 | 0.46 | −0.38 | −0.08 | −0.04 | −0.95 |
| M-AMBI | 0.02 | 0.14 | 0.17 | −0.71 | 0.26 | 0.10 | 0.24 | −0.31 | −0.67 | 0.21 | −0.19 | 0.07 |
| HFBI | 0.33 | 0.10 | 0.19 | 0.26 | 0.00 | 0.19 | 0.02 | 0.00 | 0.00 | −0.05 | −0.17 | −0.43 |
| Si | RP | NH4+ | NO2− | NO3− | DIN | M-biom | M-cover | Chl-a tot | ||||
| Survived sods | −0.65 | −0.65 | −0.74 | −0.30 | −0.50 | −0.56 | −0.33 | −0.72 | −0.33 | |||
| Sod diameter | −0.61 | −0.61 | −0.61 | −0.17 | −0.27 | −0.34 | −0.27 | −0.66 | −0.41 | |||
| Sod growth | −0.61 | −0.61 | −0.61 | −0.17 | −0.27 | −0.34 | −0.27 | −0.66 | −0.41 | |||
| Survived rhizomes | −0.68 | 0.71 | 0.56 | −0.41 | −0.56 | −0.56 | −0.39 | -0.68 | −0.51 | |||
| Rhizome diameter | −0.57 | −0.60 | −0.83 | −0.42 | −0.61 | −0.68 | −0.24 | −0.66 | −0.54 | |||
| Rhizome growth | −0.71 | −0.74 | −0.79 | −0.45 | −0.60 | −0.64 | −0.41 | −0.81 | −0.40 | |||
| Patch surface | −0.66 | −0.66 | −0.56 | −0.22 | −0.41 | −0.44 | −0.47 | −0.71 | −0.37 | |||
| MaQI | −0.69 | −0.69 | −0.70 | −0.95 | −0.88 | −0.84 | −0.37 | −0.61 | −0.36 | |||
| M-AMBI | 0.12 | 0.05 | 0.48 | −0.10 | 0.21 | 0.31 | −0.18 | −0.02 | 0.62 | |||
| HFBI | −0.17 | −0.19 | −0.76 | −0.48 | −0.67 | −0.76 | 0.26 | −0.19 | −0.24 | |||
| Fines | Density | Moisture | Porosity | TP sed | IP sed | OP sed | TN sed | TC sed | OC sed | |||
| Survived sods | −0.20 | 0.39 | −0.39 | −0.39 | −0.73 | −0.37 | −0.86 | −0.65 | 0.78 | −0.05 | ||
| Sod diameter | −0.15 | 0.49 | −0.46 | −0.37 | −0.81 | −0.49 | −0.90 | −0.65 | 0.75 | −0.10 | ||
| Sod growth | −0.15 | 0.49 | −0.46 | −0.37 | −0.81 | −0.49 | −0.90 | −0.65 | 0.75 | −0.10 | ||
| Survived rhizomes | −0.07 | 0.44 | −0.51 | −0.49 | −0.56 | −0.20 | −0.83 | −0.63 | 0.68 | −0.20 | ||
| Rhizome diameter | −0.18 | 0.74 | −0.66 | −0.57 | −0.92 | −0.61 | −0.84 | −0.61 | 0.66 | −0.22 | ||
| Rhizome growth | −0.26 | 0.57 | −0.50 | −0.43 | −0.93 | −0.64 | −0.83 | −0.54 | 0.66 | −0.02 | ||
| Patch surface | −0.24 | 0.39 | −0.41 | −0.32 | −0.66 | −0.29 | −0.85 | −0.65 | 0.75 | −0.10 | ||
| MaQI | −0.35 | 0.48 | −0.40 | −0.47 | −0.64 | −0.63 | −0.25 | −0.01 | −0.07 | 0.12 | ||
| M-AMBI | −0.07 | −0.52 | 0.64 | 0.64 | 0.17 | −0.31 | 0.74 | 0.93 | −0.71 | 0.68 | ||
| HFBI | 0.21 | 0.26 | −0.29 | −0.60 | −0.24 | 0.00 | −0.33 | −0.37 | 0.37 | −0.20 | ||
| SPM | TP part | IP part | OP part | N part | TC part | OC part | ||||||
| Survived sods | −0.01 | −0.51 | −0.31 | −0.74 | −0.26 | 0.03 | −0.16 | |||||
| Sod diameter | 0.00 | −0.56 | −0.27 | −0.81 | −0.15 | 0.17 | −0.17 | |||||
| Sod growth | 0.00 | −0.56 | −0.27 | −0.81 | −0.15 | 0.17 | −0.17 | |||||
| Survived rhizomes | 0.05 | −0.68 | −0.37 | −0.78 | −0.32 | −0.15 | −0.34 | |||||
| Rhizome diameter | −0.30 | −0.68 | −0.63 | −0.75 | −0.30 | 0.30 | −0.29 | |||||
| Rhizome growth | −0.31 | −0.57 | −0.57 | −0.71 | −0.24 | 0.29 | −0.14 | |||||
| Patch surface | 0.10 | −0.61 | −0.32 | −0.76 | −0.05 | 0.12 | −0.12 | |||||
| MaQI | −0.64 | −0.32 | −0.91 | −0.22 | −0.56 | 0.21 | −0.16 | |||||
| M-AMBI | −0.67 | 0.74 | 0.07 | 0.79 | 0.29 | 0.29 | 0.55 | |||||
| HFBI | −0.19 | −0.17 | −0.21 | −0.31 | −0.86 | −0.55 | −0.45 | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sfriso, A.; Buosi, A.; Tomio, Y.; Juhmani, A.-S.; Facca, C.; Sfriso, A.A.; Franzoi, P.; Scapin, L.; Bonometto, A.; Ponis, E.; et al. Aquatic Angiosperm Transplantation: A Tool for Environmental Management and Restoring in Transitional Water Systems. Water 2019, 11, 2135. https://doi.org/10.3390/w11102135
Sfriso A, Buosi A, Tomio Y, Juhmani A-S, Facca C, Sfriso AA, Franzoi P, Scapin L, Bonometto A, Ponis E, et al. Aquatic Angiosperm Transplantation: A Tool for Environmental Management and Restoring in Transitional Water Systems. Water. 2019; 11(10):2135. https://doi.org/10.3390/w11102135
Chicago/Turabian StyleSfriso, Adriano, Alessandro Buosi, Yari Tomio, Abdul-Salam Juhmani, Chiara Facca, Andrea Augusto Sfriso, Piero Franzoi, Luca Scapin, Andrea Bonometto, Emanuele Ponis, and et al. 2019. "Aquatic Angiosperm Transplantation: A Tool for Environmental Management and Restoring in Transitional Water Systems" Water 11, no. 10: 2135. https://doi.org/10.3390/w11102135
APA StyleSfriso, A., Buosi, A., Tomio, Y., Juhmani, A.-S., Facca, C., Sfriso, A. A., Franzoi, P., Scapin, L., Bonometto, A., Ponis, E., Rampazzo, F., Berto, D., Gion, C., Oselladore, F., Cacciatore, F., & Boscolo Brusà, R. (2019). Aquatic Angiosperm Transplantation: A Tool for Environmental Management and Restoring in Transitional Water Systems. Water, 11(10), 2135. https://doi.org/10.3390/w11102135