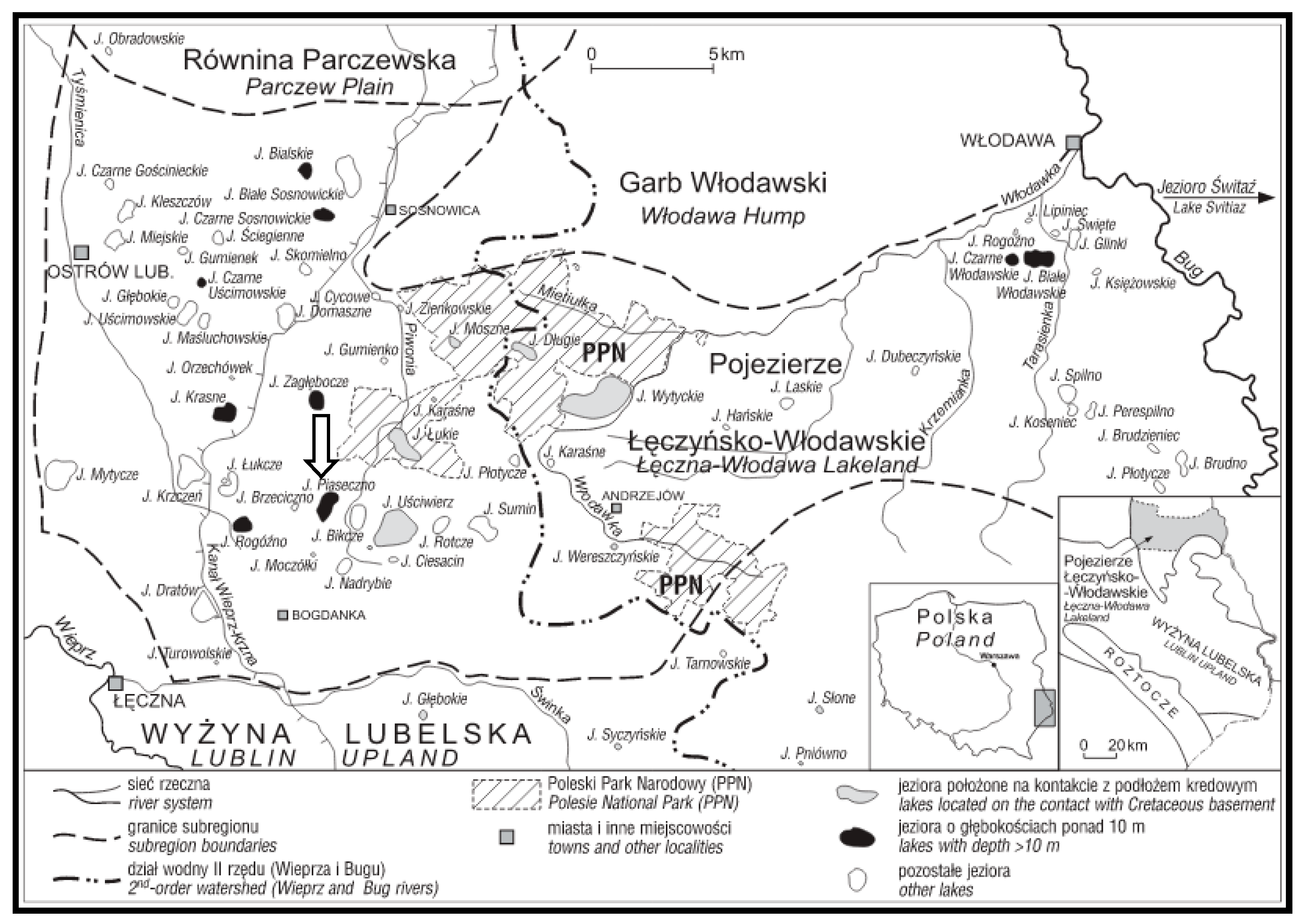
2. Study Area
In the administrative division of Poland, Lake Piaseczno (51°23′03″ N; 23°01′46″ E) is located in the centre of the Lublin Voivodeship, in the northern part of the Łęczna Poviat, in the Ludwin Commune, within the physiographic unit of the Łęczna-Włodawa Lake District included in the macroregion of Polesie Podlaskie (East Poland) (
Figure 1).
In the hydrographic division, Lake Piaseczno is included in the catchment of the Wieprz River, in the upper part of the catchment of the Piwonia Południowa River. The river originates in the vicinity of the lake, and has no surface connection with it. Therefore, its catchment is described as a surface-closed-drainage catchment [
33].
Lake Piaseczno is classified as a very deep lake, with a maximum depth of 38.8 m and a high water volume (10.67 million m
3). It is a dimictic lake in the bradymictic type, and, in terms of trophic status, it is mesotrophic (
Table 1).
Catchment morphology is one of the elements that determines a lake’s hydrological conditions and land use structure. The catchment of Lake Piaseczno is not physiographically differentiated from the remaining area of the Lake District.
The area is dominated by very gentle undulating plains of a higher accumulation horizon, with an indistinctly marked watershed, evidently contrasting with the lake basin landform with steep slopes. This determines rarely occurring surface runoff, which is related to the presence of very permeable formations and the ecological structure of the catchment [
33].
The lake has no inflows or outflows. It is primarily fed by precipitation waters supplying allochthonic substances from the catchment with surface runoff. In the summer period, it is subject to complete thermal stratification.
Part of the area around the lake constitutes a sandy beach that alternates with shore vegetation and shrubs dominated by
Salix cinerea and
Betula pubescens. To the north and south, the lake is adjacent to a mixed forest, and in its northwestern fragment it neighbours on formerly active swamps and peatlands separated by a relatively narrow transitional zone [
34,
35,
36].
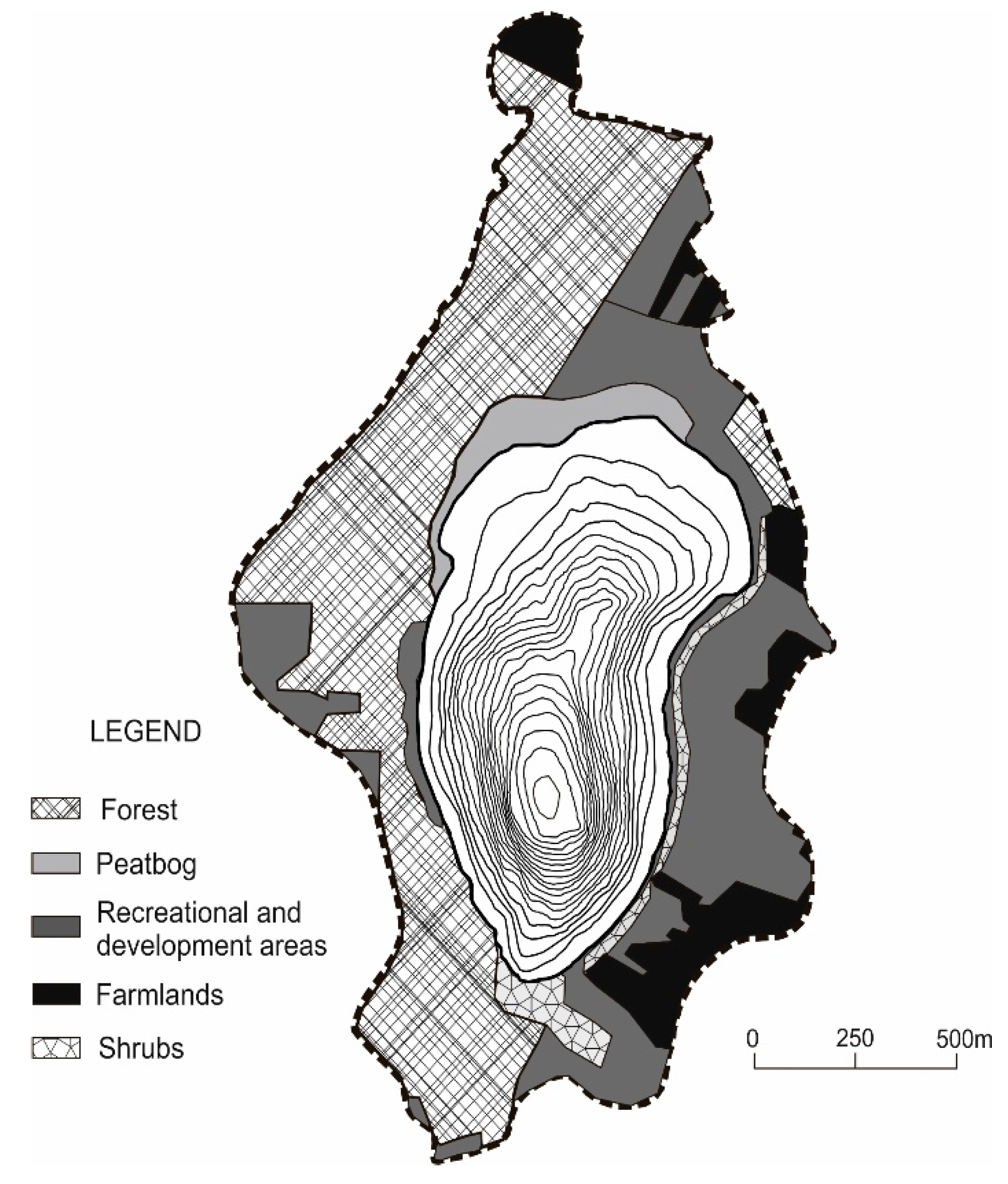
Morphological, landscape and habitat properties, as well as anthropogenic activity, determine the diversity of land use in the catchment of Lake Piaseczno (
Figure 2,
Table 2).
This largely affects the state of its surroundings, resulting in the current recreation–agricultural–forest character of the lake catchment (natural forms, extensively cultivated arable land and recreational building development (numerous cottages and two holiday resorts)). Based on the above, the lake catchment was divided into sectors with a growing share of investment land related to stationary recreation (
Table 2).
Pursuant to general guidelines for the use, protection and reclamation of lakes, and based on the categories of susceptibility to degradation and classes of water cleanliness [
37], Lake Piaseczno is included as a lake with good natural conditions, where the use and preservation of the current state of the lake should be in accordance with the rules for the protection of particularly valuable water resources [
13].
3. Material and Methods
The research on the shore vegetation of Lake Piaseczno was conducted in 2015 and 2018, in spring, summer and autumn, several times in each season. The plant species composition was determined in the shore belt of the lake, depending on the topographic conditions, with a width of approximately 10 m and along the entire shoreline of approximately 3800 m (38,000 m
2). The research was conducted in stages by designating sections with a length of 50 m corresponding to the range of particular sectors of the catchment (length of the shoreline in sectors (m): peatbog–forest–recreational (PFR): 799; recreational (R): 607; recreational–agricultural (RA): 1098; and forest–recreational (FR): 1288). Botanical analyses were focused on a multidimensional analysis of the species composition of the flora in the study area. Species were identified by means of the Rutkowski’s key [
39], and the botanical nomenclature was adopted after Mirek et al. [
40]. The identified species were ascribed to different life forms according to Raunkieaer [
41,
42], syntaxonomic groups [
41] and historical-geographic groups. This permitted the calculation of the following indicators of anthropogenic transformations in the flora [
43,
44]:
Total anthropophytisation
Total apophytisation
where Ap denotes apophytes (taxa of synanthropic plants of local origin that constitute part of the native flora and occur in anthropogenic habitats developed as a result of human activity); Sp denotes spontaneophytes (taxa developed in or imported into a given area that are able to function with no human interference and exist in nature) plus apophytes; and A denotes anthropophytes (taxa developed in a given area as a result of human pressure, taxa imported into a given area by man, and species surviving at sites under constant human pressure).
The Jaccard’s species similarity coefficient of the shore flora of Lake Piaseczno was also calculated in 2015 and 2018 [
45]. The species composition was also referred to index numbers of higher plants (moisture, acidity, trophic status, content of organic matter and granulometric composition of the soil [
42]) for the purpose of ascribing species with preferences for particular habitat properties. Shore vegetation was treated as a complex phytocoenotic system that interacts with the catchment and lake and is often transformed anthropogenically [
46]. Therefore, no analyses concerning the syntaxonomic classification of plant communities were performed. The identified species were only described as characteristic of a given phytosociological unit.
Statistical analyses were carried out in the R environment for statistical computing v. 3.5.1 [
47] with the vegan v. 2.5–3 library [
48]. Based on the Jaccard’s species similarity coefficient of the phytocoenoses, a classification analysis was performed as a measure of distance with the application of the average linkage classification algorithm (UPGMA). The obtained classification results are presented graphically in the form of dendrograms, and in a more complex form as a combination of dendrograms and a heatmap.
The research on macrophytes of Lake Piaseczno was conducted in 2007 and 2018 in July in four transects that correspond to sectors of the dominant land use in the catchment: PRF, R, RA and FR. The littoral zone was analysed from the shoreline to the maximum range of occurrence of macrophytes. The research covered all groups of macrophytes: emerging (EM), submerged (SM), floating (FM) and pleuston (PM). Their species composition was determined—with nomenclature after Matuszkiewicz [
49]—as well as the density of emerging macrophytes and their range of occurrence, biomass and dominance in biomass in particular groups [
38,
50].
The productivity of the littoral of Lake Piaseczno was analysed based on the biological lake water trophy indicators gross primary production of phytoplankton, chlorophyll
a concentration and calculated Carlson’s trophic state indices (TSIChl-
a and TSISD). The analyses were implemented in the summer season of 2015 and 2018 (two terms one month apart in each season) in the littoral zone adjacent to particular sectors of the catchment. Study sites were designated in the centre of the shoreline of each sector at a distance of approximately −10 m from the shore, depending on the bottom slope inclination, to obtain sample immersion to a depth of approximately 0.75 m. For sectors 2 and 3, due to their homogeneity, one study site was designated on their arbitrary boundary. The limnological status of Lake Piaseczno was determined based on the same research conducted in the deepest place in the lake (38.8 m) in the peatogenic zone (depths of 0.75 m and 2.5 m). The study results were averaged for the analysed profile and study season. Gross primary production of phytoplankton was measured by means of the aerobic method [
51]. The concentration of dissolved oxygen in water was determined by means of the Winkler iodometric titration method [
52]. The amount of released oxygen for subsequent study terms was converted to amounts of carbon assimilated from 1 m
2 per hour (mg C
ass·m
−2·h
−1), assuming that 1 g of released oxygen corresponds to 0.312 g of assimilated carbon [
53]. The obtained results after conversion to mgC
ass·m
−2·d
−1 (assuming that d = 12 h) described the level of trophy of lake waters (according to [
2] and [
54]). Concentration of chlorophyll
a, the most important photosynthetic pigment, was analysed by means of the spectrophotometric method [
55] and expressed in mg Chl.
a·m
−2. Based on chlorophyll
a concentration values expressed in μg Chl.
a·dm
−3, TSI CHLa was calculated [
56]:
The research on biological trophic status indicators was supplemented by measurements of the basic physical-chemical water properties, i.e., electrolytic conductivity (μS·cm
−1), reaction (pH), and oxygen concentration in mg O
2·dm
−3, by means of a certified field meter (HI 9811-5) or titration methods (oxygen concentration: control samples for gross primary production of phytoplankton). Based on measurements of Secchi visibility (m), which determines water transparency in the pelagial zone, the TSI SD was calculated [
56]:
The obtained study results were analysed with unitary coefficients of export of surface loads of biogenic substances from catchment areas under different land uses according to Soszka [
10]: 1 ha of arable fields in the lake’s catchment generates 9 kg N and 0.3 kg P; 1 ha of meadows and pastures 3 kg N and 0.2 kg P; 1 ha of forests 1.5 kg N and 0.1 kg P; and loose building development (the equivalent of recreational plots) 9 kg N and 0.3 kg P (kg·ha
−1·year
−1). The above nutrient values, generated by different types of catchment management, were calculated in relation to the surface area and parameters of the Piaseczno Lake catchment. The potential amount of biogens from beach and bathing area recreation was also determined, with the assumption that each tourist in the shore zone of the lake generates 1 g N·day
−1 [
57] and 0.457 g P·day
−1 [
58] and the calculated index of physical carrying capacity (PCC), which describes the maximum number of persons in the shore zone of a lake participating in recreation with no harm to its ecosystem, is 5683.2 person per day [
21].
4. Results
A total of 235 species of vascular plants (233 species in 2015 and 225 species in 2018) from 56 families, representing four botanic classes, were indentified in plant communities of shores of Lake Piaseczno in 2015 and 2018. In class MAGNOLIOPSIDA, the highest species richness concerned families Asteraceae (25 species); Rosaceae (17 species); Plantaginaceae (11 species); Lamiaceae (11 species); and Cariophyllaceae and Salicaceae (10 species each). In class LILIOPSIDA, the highest species richness concerned Poaceae (26 species) and Cyperaceae (19 species). The remaining families in all classes were represented much less abundantly (compare
Table 3).
The tree, shrub and small tree vegetation included 35 species from 10 families belonging to two botanical classes. The most abundant representation concerned families Salicaceae (10 species) and Rosaceae (8 species), and the least abundant representation concerned families Adoxaceae, Oleaceae, Sapindaceae and Pinaceae (1 species each).
For both study years, the sector with the highest number of species was the recreational–agricultural sector (RA, 138 species in 2015 and 135 species in 2018). In 2015, the lowest number of species was identified in the recreational sector (R, 115 species), and in 2018 in the forest–recreational sector (FR, 109 species), where an almost 7% decrease in the number of species was recorded in comparison to the previous study year (
Table 3). The occurrence of 29 species permanently present in all sectors for both study seasons was determined:
Achillea millefolium,
Bidens tripartita,
Eupatorium cannabinum,
Hieracium pilosella,
Hieracium murorum,
Tussilago farfara,
Myosotis palustris,
Stellaria graminea,
Calluna vulgaris,
Lotus corniculatus,
Mentha arvensis,
Prunella vulgaris,
Rumex acetosa, Rumex acetosella, Ranunculus flammula, Potentilla erecta, Galium plaustre, Urtica dioica,
Alisma plantago-aquatica,
Carex flava,
Juncus conglomeratus,
Agrostis capillaris and
Holcus lanatus, and among tree and shrub vegetation:
Quercus robur,
Populus tremula,
Salix aurita,
Salix cinerea,
Salix pentandra and
Pinus sylvestris. A total of 59 species of plants occurring in only one study sector was also identified. The highest number of such species occurred in the peatbog–forest–recreational sector (PFR, 27 species) and forest–recreational sector (FR, 24 species). The remaining study sectors showed less varied character: recreational–agricultural sector (RA, four species); recreational sector (R, zero species). Part of a species occurring in only one sector was found in only one study year, namely 2015 (FR:
Aegopodium podagraria,
Mycelis muralis,
Knautia arvensis,
Scirpus sylvaticus,
Bromus hordeaceus,
Festuca altissima; PFR:
Vicia angustifolia,
Polygonum lapathifolium,
Rumex crispus,
Carex riparia,
Carex rostrata; RA:
Chelidonium majus); and in 2018, only
Capsella bursa-pastoris in FR and
Cerastium holostenoides in RA (
Table 3).
In terms of life forms according to Raunkieaer, for both study years, plant communities of shore sectors of Lake Piaseczno were dominated by hemicryptophytes (H), reaching almost a 60% contribution. Geophytes (G) and terophytes (T) showed more than a 14% share in phytocoenoses. Hydrophytes and helophytes (Hy) reached 11%, megaphanerophytes (M) and nanophanerophytes (N) 7–8%, and green chamaephytes (C) and woody chamaephytes (Ch) showed a maximum of several percent participation in the shore communities of Piaseczno. Low representation also concerned lianas (li, two species) and semi-parasites (pp, one species) (
Table 4). In the phytosociological context, sector PFR was dominated by plant species from the classes Molinio-Arrhenatheretea, Oxycocco-Sphagnetea, Phragmitetea and Scheuzerio-Caricetea nigrae; sectors R and RA were dominated by plant species from the classes Phragmitetea and Molinio-Arrhenatheretea; and sector FR was dominated by plant species from the classes Phragmitetea, Querco roboris-Pinetum and Nardo-Callunetea. Species typical of the designated classes co-occurred, with a considerable number of accompanying species supplementing the composition of the plant communities.
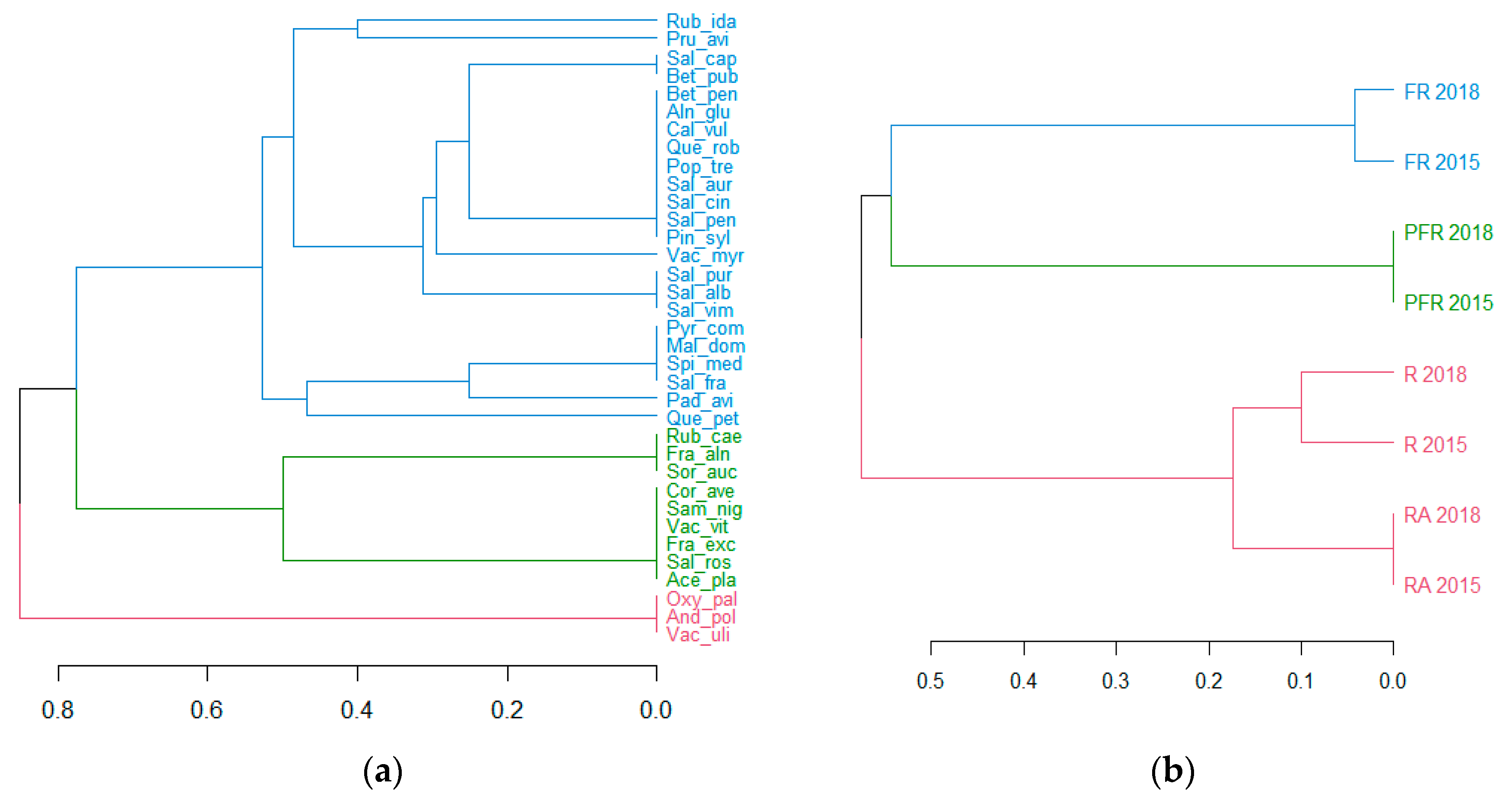
The Jaccard’s species similarity coefficient for the studied phytocoenoses in shore sectors of Lake Piaseczno in 2015 varied from 0.26 (R–FR) to 0.84 (R–RA), and in 2018 from 0.26 (R–FR) to 0.76 (R–RA), for both study seasons, which usually oscillated around 0.3 (
Table 5). This suggests diversity of flora within the study sectors in spite of the typological similarity of habitats. In the comparison of the study sectors in the 2015 and 2018 summer seasons, the Jaccard’s similarity coefficient oscillated between 0.81 (FR–FR) and 0.91 (PFR–PFR), suggesting insignificant changes in the species composition of plants (
Table 5,
Figure 3b).
In the case of tree and shrub vegetation, somewhat more complete information was provided by a hierarchical classification analysis with the Jaccard’s coefficient as the similarity measure, which was performed both for plant species and for shore sectors of Lake Piaseczno (
Figure 3a,b). At the first stage, it permitted the designation of three more or less-diverse groups of plant species, characterised by a higher measure of co-occurrence in the phytosociological context, belonging to five groups: Oxycocco-Sphagnetea, Scheuzcherio-Caricetea fuscae, Betuletum pubescentis, Salicetum pentandro-cinereae and Querco-Piceetum.
Group I includes Rubus idaeus,
Prunus avium,
Salix caprea,
Betula pubescens,
Betula pendula,
Alnus glutinosa,
Calluna vulgaris,
Quercus robur,
Populus tremula,
Salix aurita,
Salix cinerea,
Salix pentandra,
Pinus sylvestris,
Vaccinium myrtillus,
Salix purpurea,
Salix alba,
Salix viminalis,
Pyrus communis,
Malus domestica,
Spiraea media,
Salix fragilis,
Padus avium and
Quercus petraea (
Figure 3a). In phytosociological terms, Group I shows considerable diversity, including species from peatland communities: Oxycocco-Sphagnetea (five species) and Scheuzcherio-Caricetea fuscae (three species); from shrub communities: Betuletum pubescentis and Salicetum pentandro-cinereae (three species each); and from the forest community: Querco-Piceetum (one species). The group included a high number of species occurring in different shore sectors of Lake Piaseczno in low numbers (often single individuals) with variable and other phytosociological affinity (13 species).
Group II includes
Rubus caesius,
Frangula alnus,
Sorbus aucuparia,
Corylus avellana,
Sambucus nigra,
Vaccinium vitis-idaea,
Fraxinus excelsior,
Salix rosmarinifolia and
Acer platanoides (
Figure 3a). The species show similarly diverse phytosociological affinity (Oxycocco-Sphagnetea, one species; Scheuzcherio-Caricetea fuscae, one species; Salicetum pentandro-cinereae, two species; Querco-Piceetum, three species; other, four species).
The small group III includes
Oxycoccus palustris,
Andromeda polifonia and
Vaccinium uliginosum (
Figure 3a), and was particularly related to the communities Oxycocco-Sphagnetea (two species) and Scheuzcherio-Caricetea fuscae (one species).
The hierarchical classification of shore sectors of Lake Piaseczno in terms of species also permitted the determination of a lack of considerable difference in tree and shrub communities for subsequent study years and the determination of the typological similarity for sectors R and RA (
Figure 3b).
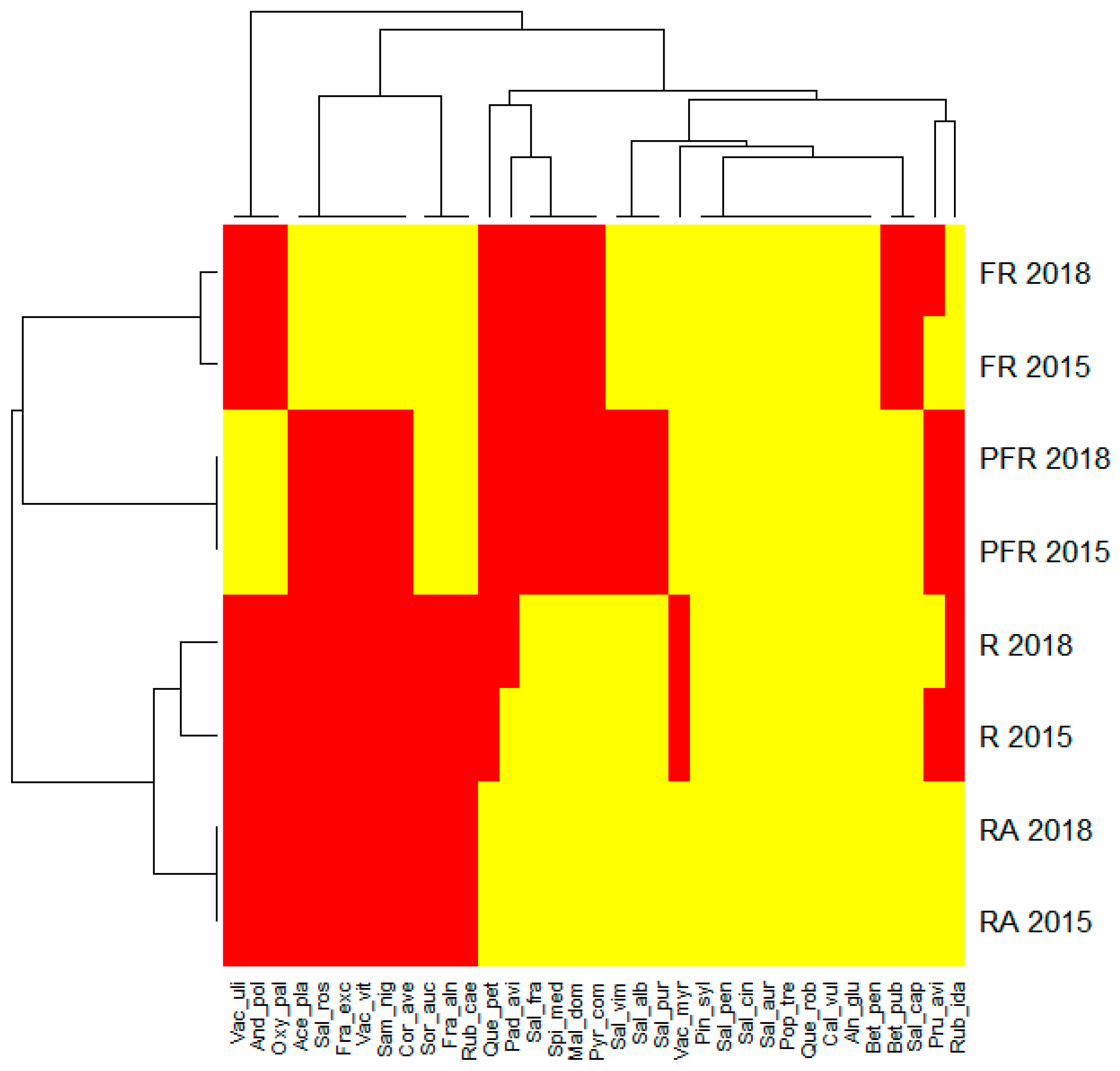
An analysis of the heatmap that ascribed the frequency of co-occurrence of the identified tree and shrub species to the phytocoenoses of the study sectors in subsequent years revealed several other patterns (
Figure 4). For example, more than 54% of tree and shrub species were found in the sectors RA and R, and slightly more than 25% of these species occurred in all sectors in both years of research. Three species (
Andromeda polifolia,
Oxycoccus palustris and
Vaccinium uliginosum) occurred only in sector PFR, two species (
Betula pubescens and
Salix caprea) were absent only in sector FR in both study seasons, and one species (
Vaccinium myrtillus) was absent only in sector R in the same period (compare
Figure 4).
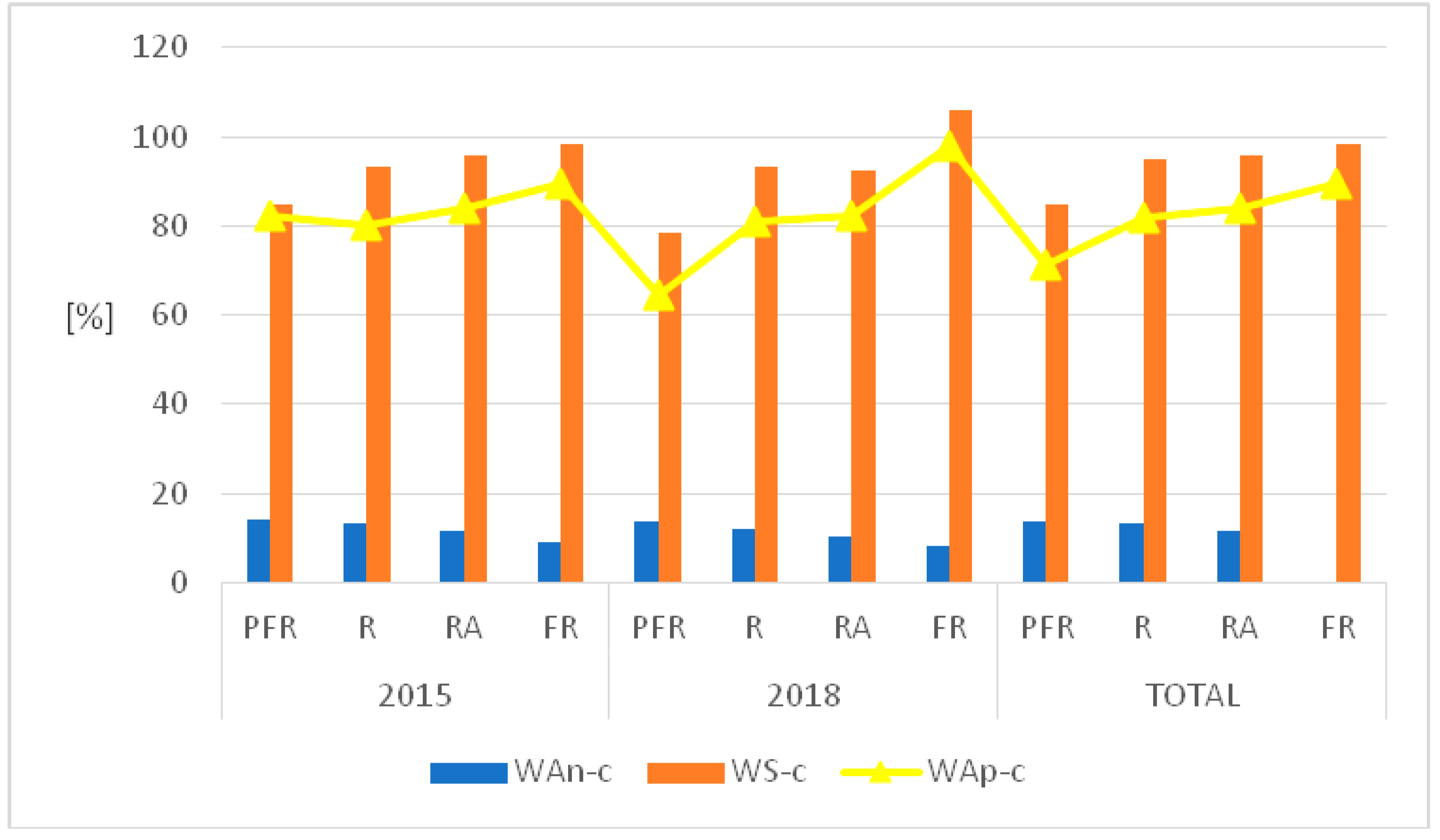
The classification of shore species of Lake Piaseczno that were identified in 2015 and 2018 in terms of historical-geographic range showed that spontaneophytes (Sp) constituted the majority in the studied phytocoenoses (45.9%, 108 species). Apophytes (Ap) reached a 38.7% contribution (91 species). The remaining groups, classified as antropophytes (A) (archeophytes (Arch, 10 species), kenophytes (Ken, 6 species) and diaphytes (0 species)), were represented in a very scarce degree or not at all. It is worth mentioning that 20 of the identified species showed unidentified historical-geographic affinity. The above classification permitted the calculation of indicators of antropogenic transformations in the flora of the entire shore of Lake Piaseczno in 2015 and 2018, which were analysed both together and separately (changes concerned only 2018): total synanthropisation (WS-c) equalled 86.29% (2018, 85.71%); total apophytisation (WAp-c) equalled 73.38% (2018, 72.26%); and total anthropophytisation (WAn-c) reached 12.9% (2018, 13.44%).
The values of indicators of anthropogenic transformations in the flora of the shore of Lake Piaseczno, when divided into sectors, were similar (
Figure 5). The lowest values of WS-c in both study seasons were recorded for sector PFR (2015, 84.61%; 2018, 78.46%; total, 84.84%), and the highest for sector FR (2015, 98.21%; 2018, 106%; total, 98.21%). The values of WAn-c showed the opposite pattern. The lowest values of the indicator were determined for both study years in the case of sector FR (2015, 8.92%; 2018, 8.0%; total, 8.92%), and the highest in sector PFR (2015, 14.03%; 2018, 13.84%; total, 13.63%). For WAp-c, the extreme values were variable: in 2015 for sector R (80.0%); in 2018 for sector PFR (64.61%); and, in the case of the joint analysis of both years (total), for sector PFR (71.21%) (
Figure 5).
The classification of the identified plant species to selected index numbers of higher plants, describing their habitat preferences, shows that the primary component of the shores of the analysed lake was comprised of plants that are characteristic of fresh and moist soils. In terms of trophic status, they prefer habitats that are moderately abundant in nutrients and eutrophic components and occur on moderately acidic and neutral soils, loamy–sandy or sandy, with higher than average permeability. In terms of content of organic matter, habitats of the shore flora of Piaseczno Lake had mineral-humus and organogenic soils that were related to the peatbog areas (compare
Table 6). Based on the selected parameters in the analysed sectors of the lake shore, a high contribution (in majority exceeding 50%) was reached by stenobionts with a relatively narrow range of ecological tolerance (
Table 6).
A total of 37 plant species was identified in the plant communities of the littoral (macrophytes) in all sectors of Lake Piaseczno in 2007 and 2018 (23 species in 2007 and 35 species in 2018). They belonged to 24 families from 5 botanical classes. The highest representation concerned class LILIOPSIDA (24 species, 12 families), and the lowest representation concerned classes EQISETOPSIDA, POLIPODIOPSIDA and BRYOPSIDA (one species and one family each). In class MAGNOLIOPSIDA, the family with the highest number of species was Lamiaceae (three species), and in class LILIOPSIDA, the families with the highest number of species were Characeae (five species), Cyperaceae (five species) and Potamogetonaceae (three species). The remaining families in both classes had scarce representation (one species each) (
Table 7).
The majority of the identified plant species belonged to the group of emerging macrophytes (EM, 22 species). Twelve species were included in the submerged macrophytes (SM) group, two species (
Lemna minor,
Utricularia vulgaris) belonged to the group of pleuston macrophytes (PM), and only one species (
Nymphea alba) represented the group of floating macrophytes (FM) (
Table 7).
For both study seasons, the sector with the highest number of species was the peatbog–forest–recreational sector (PFR, 19 species (2007) and 26 species (2018)). The lowest number of species was found in the recreational–agricultural (RA) sector and the forest–recreational (FR) sector (9 species (2007) and 14 species (2018), respectively). The occurrence of four species (Utricularia vulgaris, Ceratophyllum demersum, Myriophyllum alternifolium and Phragmites australis) permanently occurring in all sectors of the littoral of Lake Piaseczno was determined for both study seasons.
In 2007 and 2018, part of a species occurred in only one study sector: PFR (
Myosotis palustis,
Scutelaria galericulata,
Lythrum salicaria,
Nymphea alba,
Rumex hydrolapathum,
Lysymachia thyrsiflora,
Potamogetom praelongus and
Thelypteris palustris) and FR (
Nitella flexilis). Part of a species was found in one sector in one study year (2018): PFR (
Calla palustris,
Carex pseudocyperus,
Carex vesicaria,
Iris pseudacorus and
Equisetum fluviatile); R (
Galium palustre); and FR (
Fontinalis antypyretica) (
Table 7).
The calculated Jaccard’s species similarity coefficient for the analysed littoral phytocoenoses in sectors of Lake Piaseczno in 2007 varied from 0.36 (PFR–RA) to 0.63 (RA–FR) and in 2018 from 0.26 (PFR–FR) to 0.64 (RA–FR), suggesting diversity of flora within the study sectors.
In the case of the mutual comparison of the species composition of flora of the lake’s littoral in 2007 and 2018, the Jaccard’s coefficient values were in a range from 0.53 (RA–RA) to 0.88 (FR–FR), suggesting more considerable transformations in the species composition of plants (
Table 8).
The distribution and biomass of macrophytes occurring in the phytolittoral of Lake Piaseczno were evidently variable in particular study sectors. The highest species diversity occurred in sector PFR, particularly among emerging macrophytes. They reached the greatest range (1.4 m of depth), density (52 ind·m
−2) and biomass values here (
Table 9). The lowest values of the analysed properties of macrophytes occurred in sector RA, with the exception of the biomass of submerged macrophytes, which varied from 216.9 g·m
−2 to 270.9 g·m
−2 in the sector. The general pattern observed in all of the analysed sectors involved high values of one group of macrophytes, and lower values of the other. If emerging macrophytes reached higher biomass values, submerged macrophytes automatically showed a lower value. A well-developed reed belt may limit the supply of biogenic substances to the depths of the lake and, therefore, the development of the occurring macrophytes. The width of the reed belt is, on the one hand, determined by the morphometric parameters of the lake, and, on the other hand, by the amount of supplied biogenic substances from the buffer zone. The widest reed belt—of more than 100 m—occurred in sector PFR and was approximately 40 m in sector R. The biomass of this group of macrophytes was high (
Table 9). Therefore, a higher than average external supply of biogens presumably occurs here from this form of land use in the zone.
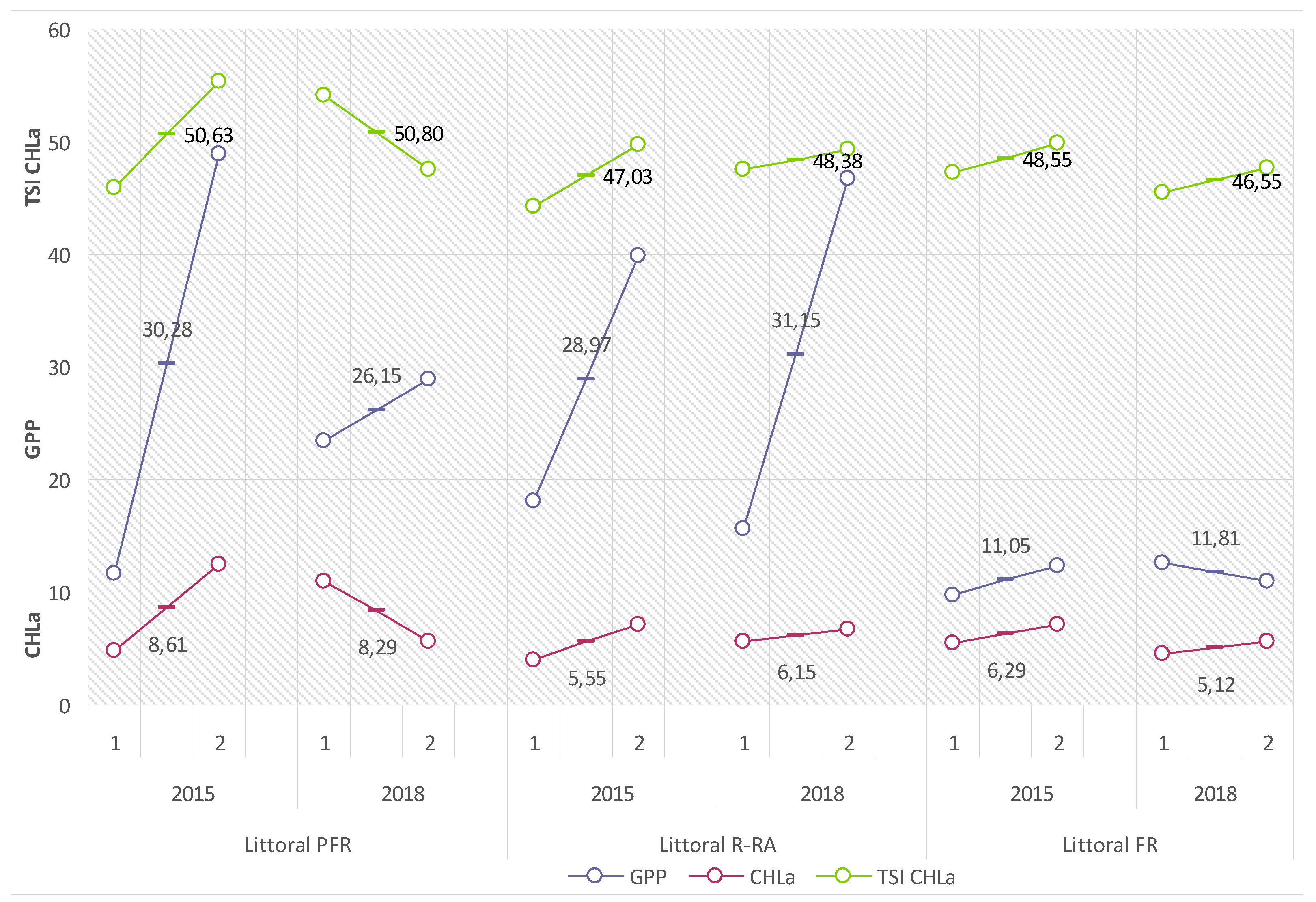
The productivity of the littoral of Lake Piaseczno in the 2015 and 2018 summer seasons, measured with values of biological indicators of water trophy, suggested the intensification of its eutrophication in different ways. This could be related to the diverse dynamics of the internal metabolism of the lake, and—in the modifying effect on biogeochemical buffer zones of the shores and littoral of the lake—variable amounts and rates of supply of biogenic substances from the catchment.
Gross primary production of phytoplankton (GPP) in the 2015 summer season adopted a range of average values from 11.05 (FR) to 30.28 (PFR) mg C
ass·m
−2·h
−1 (132.6–363.4 mg C
ass·m
−2·d
−1), and in 2018 from 11.88 (FR) to 31.15 (R-RA) mg C
ass·m
−2·h
−1 (142.6–373.8 mg C
ass·m
−2·d
−1) (
Figure 6).
Chlorophyll
a concentrations (CHLa) showed a somewhat lower variability. In the 2015 summer season, the mean values of the factor varied from 5.55 (R–RA) to 8.61 (PFR) mg Chl.
a·m
−2, and in 2018 from 5.12 (FR) to 8.29 (PFR) mg Chl.
a·m
−2. They contributed to the adequate variability of values of the calculated Carlson’s Trophic State Index (TSI CHLa): 47.03 (R–RA) and 50.63 (PFR) for the 2015 summer season and 46.55 (FR) and 50.8 (PFR) for the 2018 summer season (compare
Figure 6).
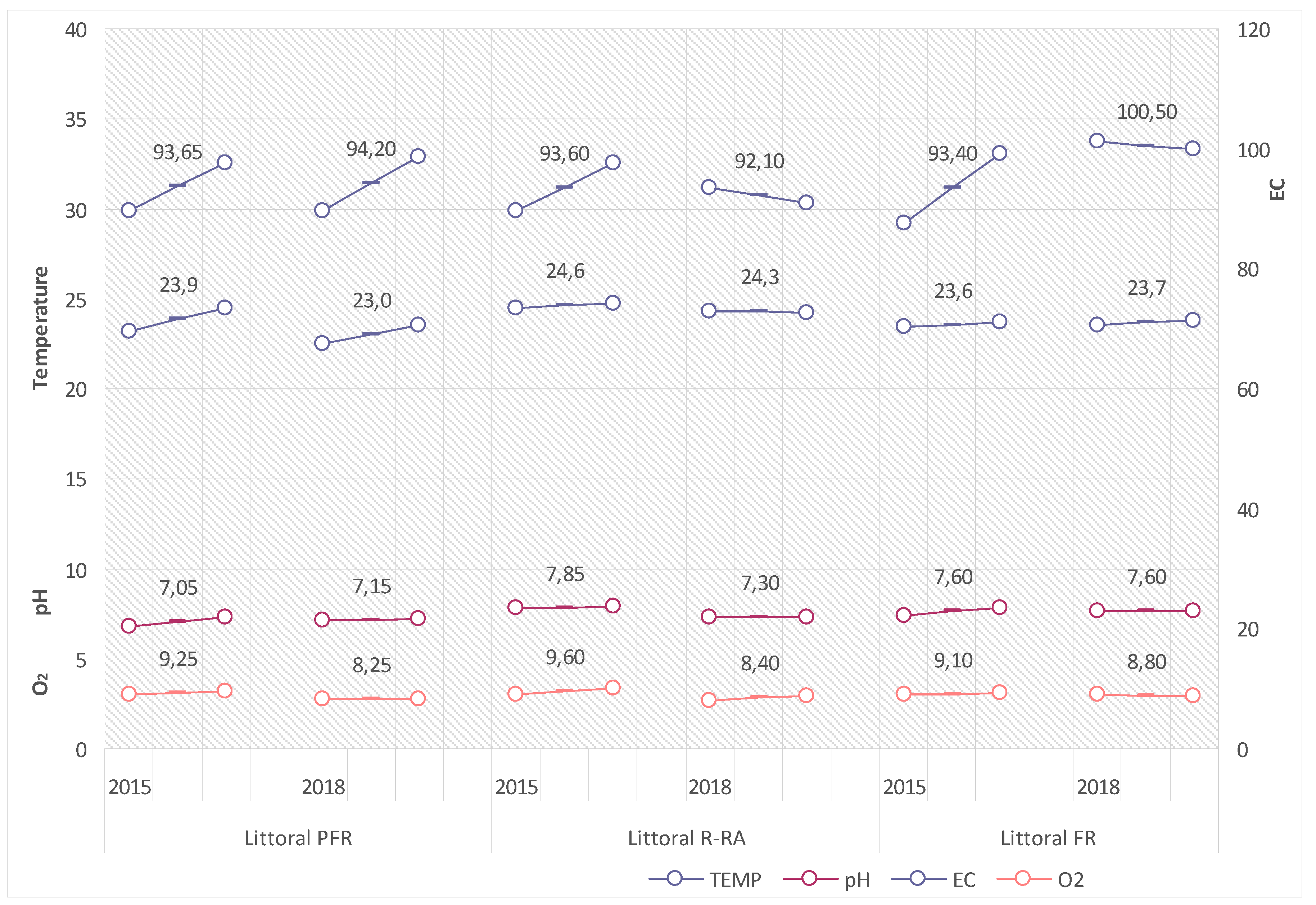
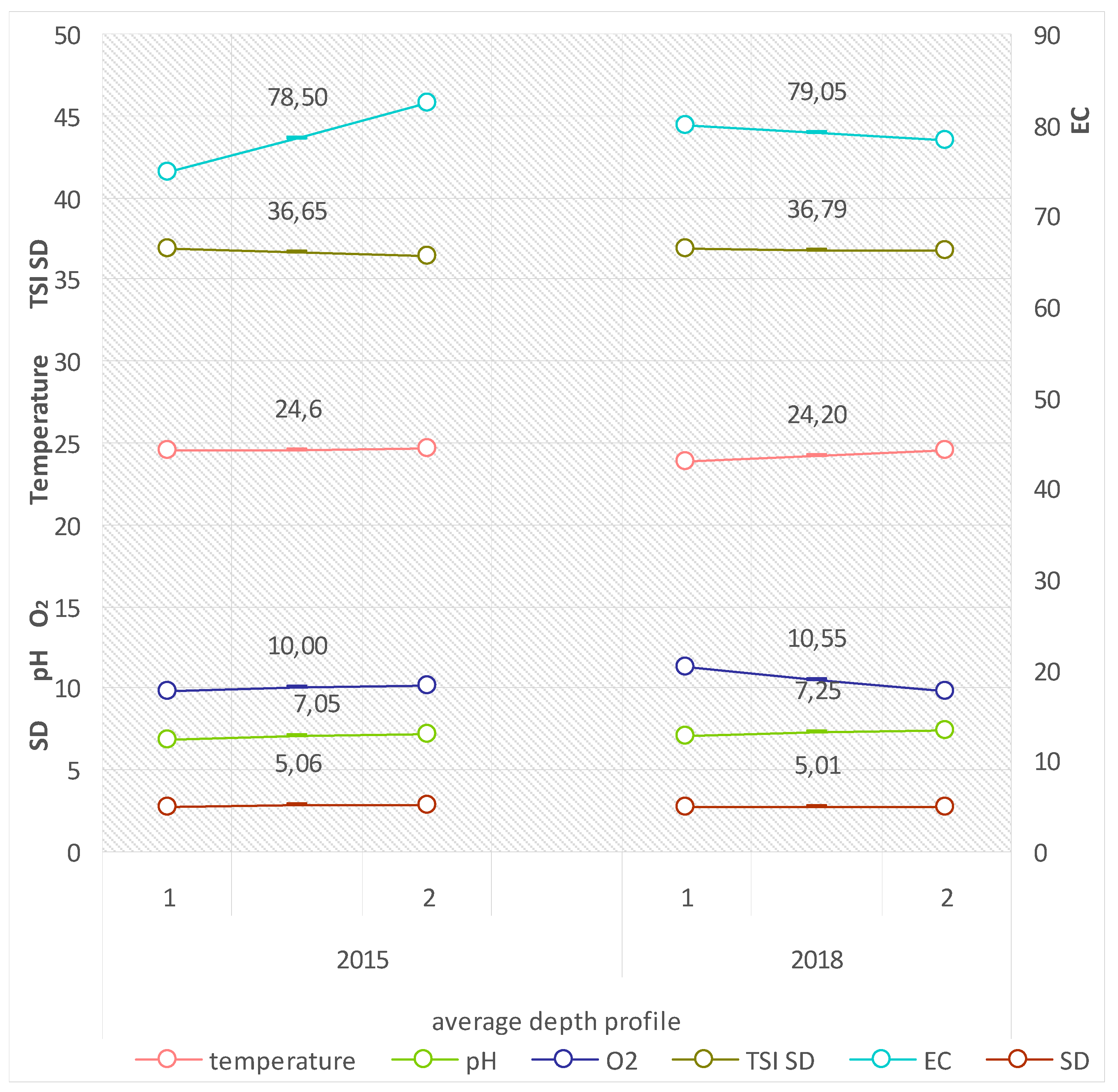
The values of the analysed physical-chemical properties of littoral waters in the period showed low variability, adopting ranges that are typical of waters with low intensity in the eutrophication process. Reaction (pH) in the 2015 and 2018 summer seasons varied from 7.05 to 7.85, the electrolytic conductivity (EC) from 92.1 to 100.5 µS·cm
−1, the oxygen concentration (O
2) from 8.25 to 9.60 mg O
2·dm
−3, and water temperature from 23.0 to 24.6 °C (compare
Figure 7).
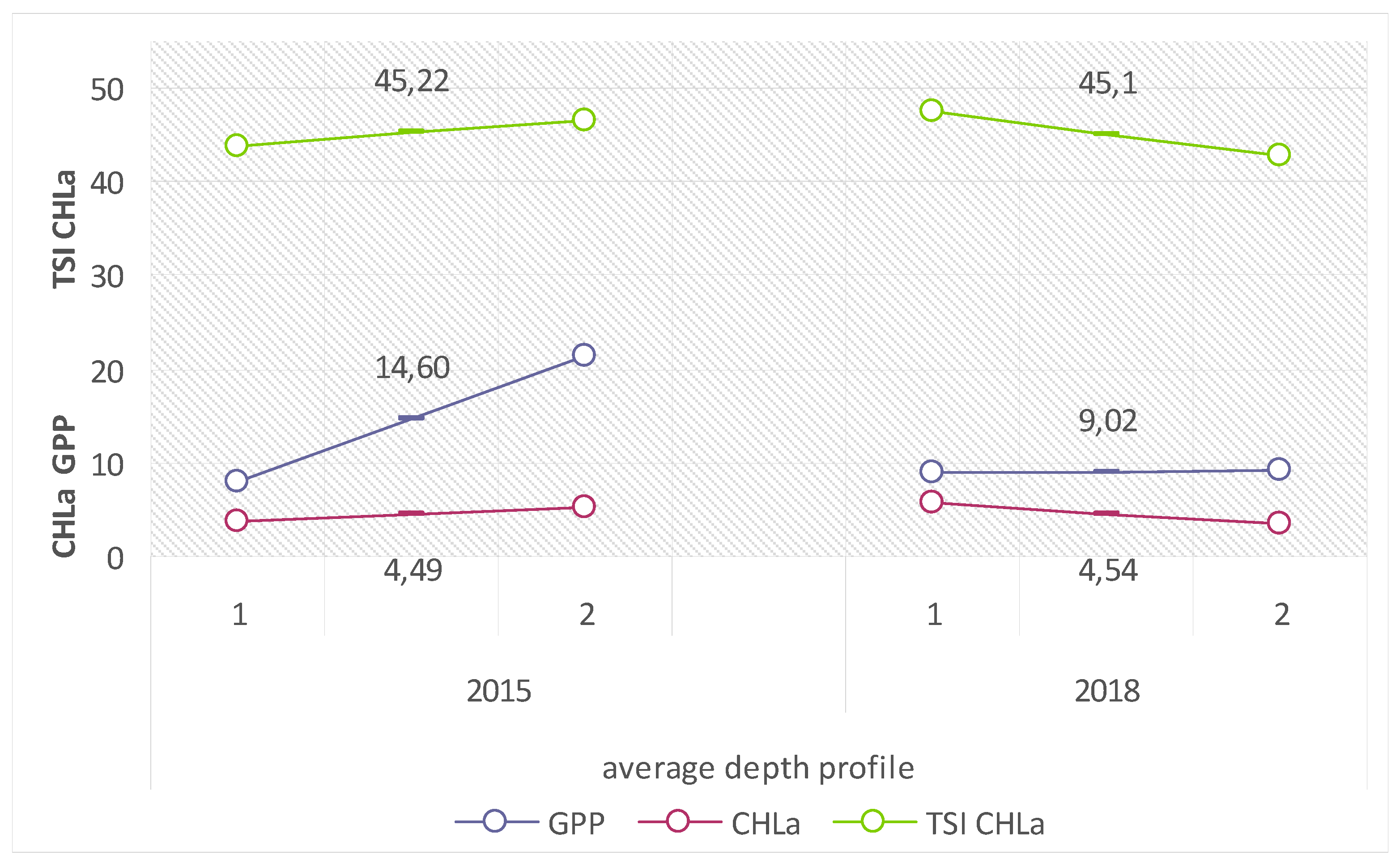
Due to the intensification of metabolic processes related to the direct contact with the catchment, the lake’s littoral is not a competent location for the analysis of the limnological status of the entire lake. The appropriate place for such analyses is the trophogenic zone of the pelagial of Lake Piaseczno, for which the mean values of biological trophic indices in the 2015 and 2018 summer seasons were as follows: GPP: from 9.02 (2018) to 14.6 (2015) mg C
ass·m
−2·h
−1 (translating into: 108.24–175.2 mg C
ass·m
−2·d
−1); CHLa: from 4.49 (2015) to 4.54 (2018) mg Chl.
a·m
−2 (which translated into similar values of the calculated indicator); and TSI CHLa: from 45.1 (2018) to 45.22 (2015) (
Figure 8).
Research supplementing the values of the physical-chemical properties of the pelagial waters in Lake Piaseczno also showed no considerable differences for both study seasons, and were in ranges that are typical of water bodies with a poor trophic status, as additionally suggested by the value of the indicator TSI SD being <40 (compare
Figure 9).
The limnological status of a water body is determined by the quantity and quality of allochthonic substances that are supplied from the catchment, depending on the character of the occurring ecosystems and their anthropogenic transformations. An analysis of the unitary coefficients of the export of surface loads of biogenic substances from the catchment area of Lake Piaseczno with variable forms of land use showed that the load of biogenic substances generated annually to the lake is 1151.1 kg N and 42.49 kg P. The highest contribution in the balance of biogens is reached by recreational areas and loose rural building development: 689.4 kg N and 22.98 kg P·year−1. Due to their phytogeochemical property of the bioretention of biogenic substances, forests, tree stands and shrubs generate relatively small amounts of N and P (123.57 and 8.24 kg·year−1, respectively). Currently, extensively used arable land introduces approximately 338.13 kg N and 11.27 kg P·year−1 to the lake waters annually.
Tourism and unorganised recreation related to beaches and bathing areas generate variable amounts of biogenic substances, depending on the number of persons participating in recreation in the shore zone, which in turn depends on appropriate weather conditions and the day of the week (weekday or weekend, [
20,
21]). Based on the adopted methodical assumptions, it was determined that the maximum amount of biogenic substances from this type of human pressure could potentially reach approximately 170.49 kg N·year
−1 and 77.91 kg P·year
−1 (arbitrarily assuming 30 days in a year with the maximum pressure of beach tourism, which is currently doubtful due to the decreasing number of tourists).












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

