The Impact of Climate Change on Hydroecological Response in Chalk Streams



Abstract
:1. Introduction
- Increased diversity fosters greater productivity of ecosystem functions;
- The diversity-stability hypothesis [5] states that biodiversity introduces redundancy in the system, thereby introducing both resistance and resilience to environmental change;
- The loss of certain species may have keystone effects which cascade through the ecosystem [6]; for example, Woodward, et al. [7] observed that the presence and absence of freshwater shrimp (Gammarus pulex), a dominant predator in chalk streams, exerted a strong influence on detrital processing rates.
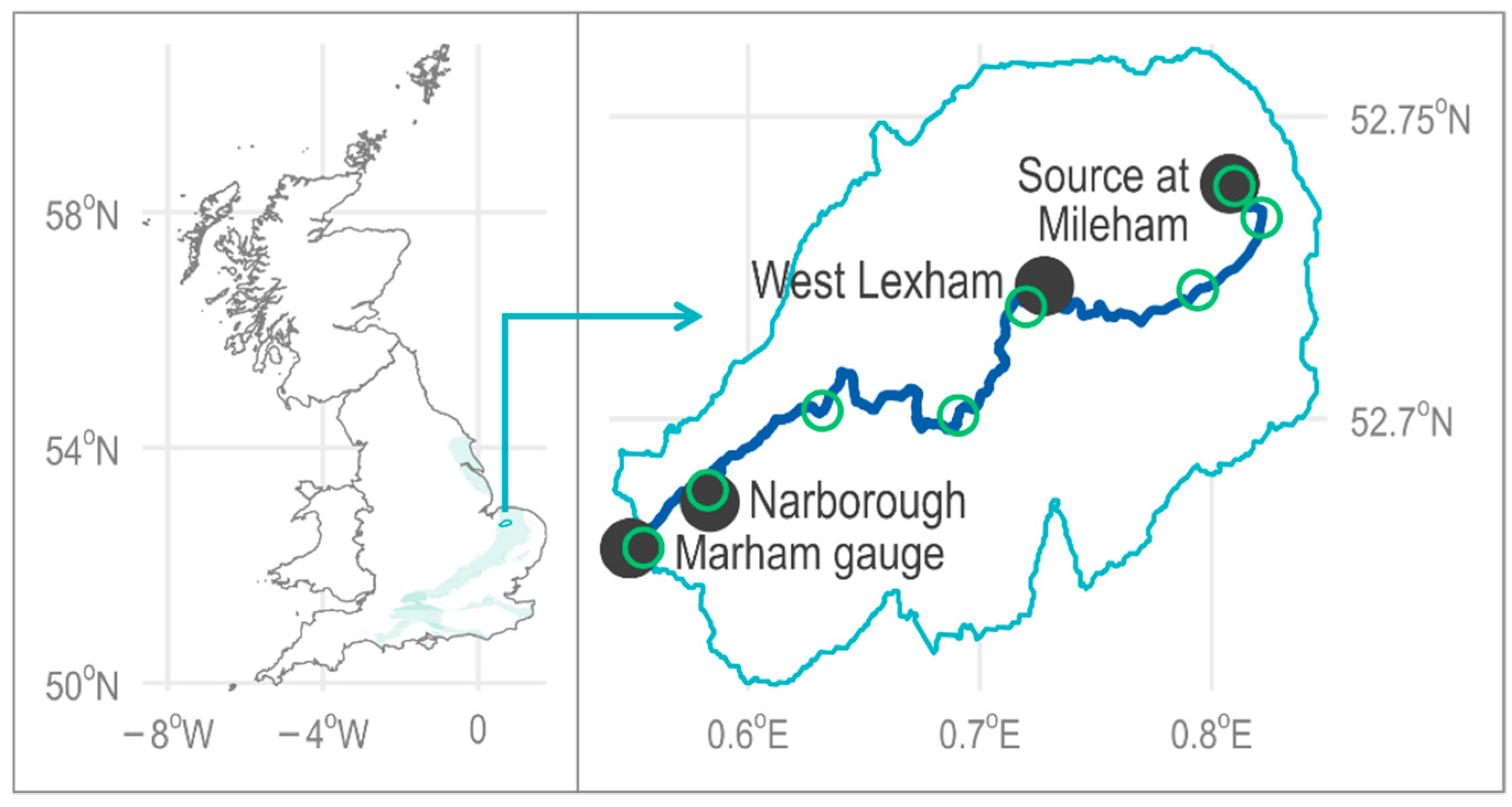
2. Case Study Catchment—River Nar
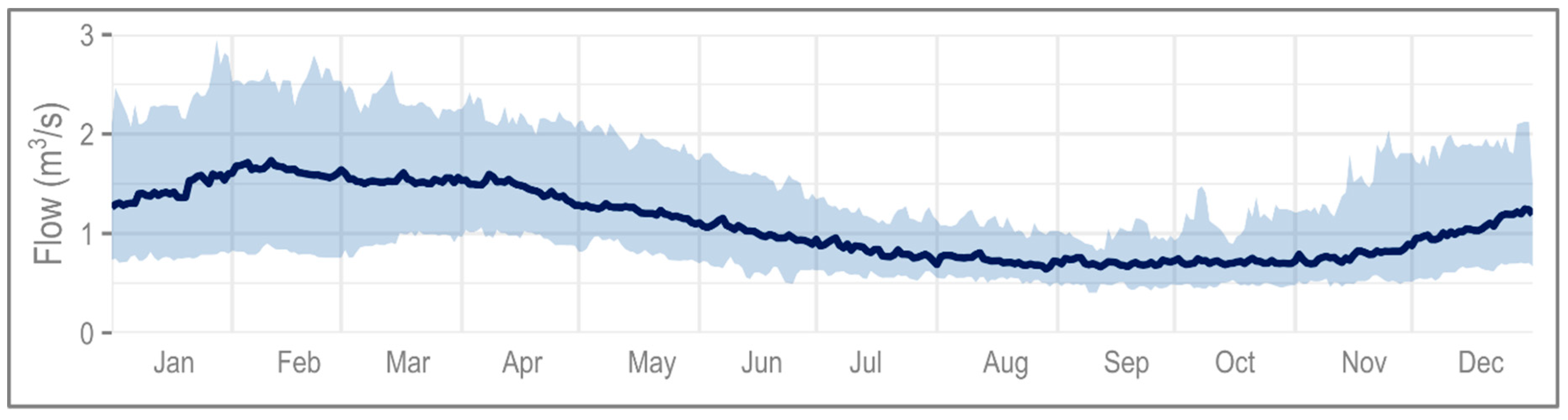
2.1. Hydrology
2.2. Hydrogeomorphological Pressures
2.3. Biodiversity
3. Methods
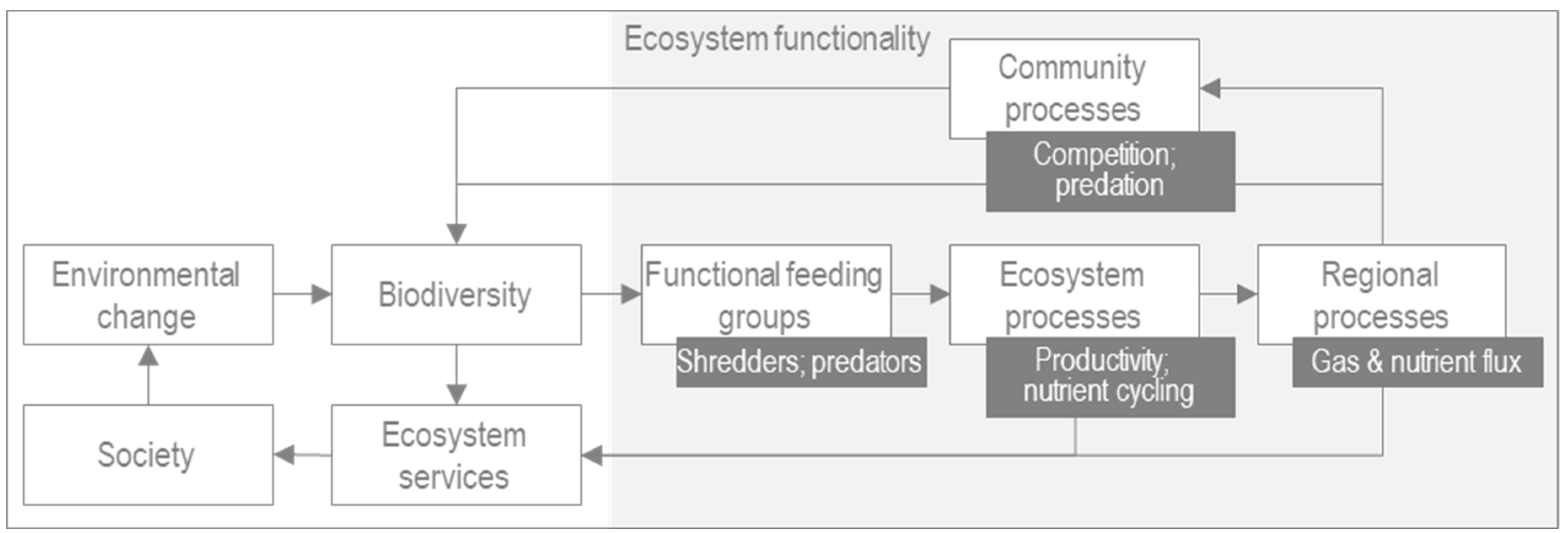
3.1. River Health and Ecosystem Functionality
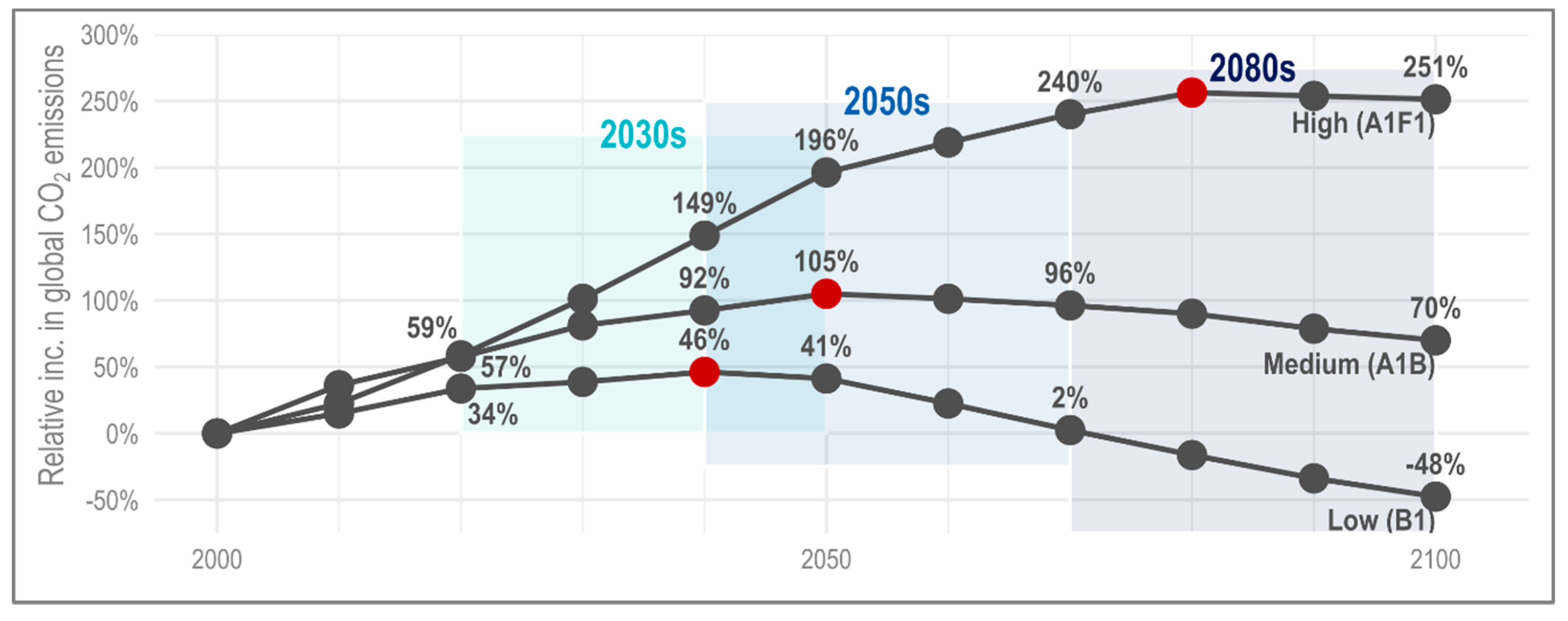
3.2. Climate Projections
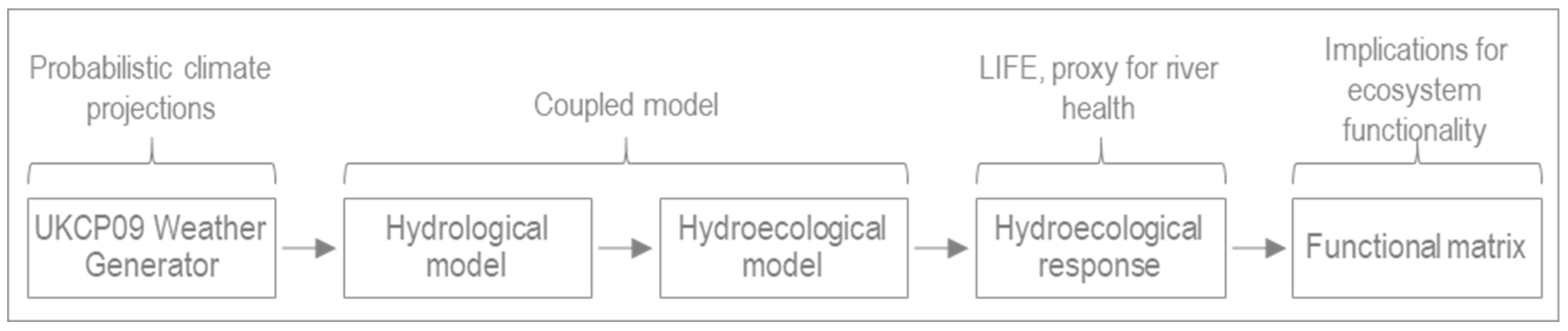
3.3. Coupled Hydrological-Hydroecological Modelling Framework
3.3.1. Hydrological Model
3.3.2. Hydroecological Model
3.3.3. Analysis
4. Results
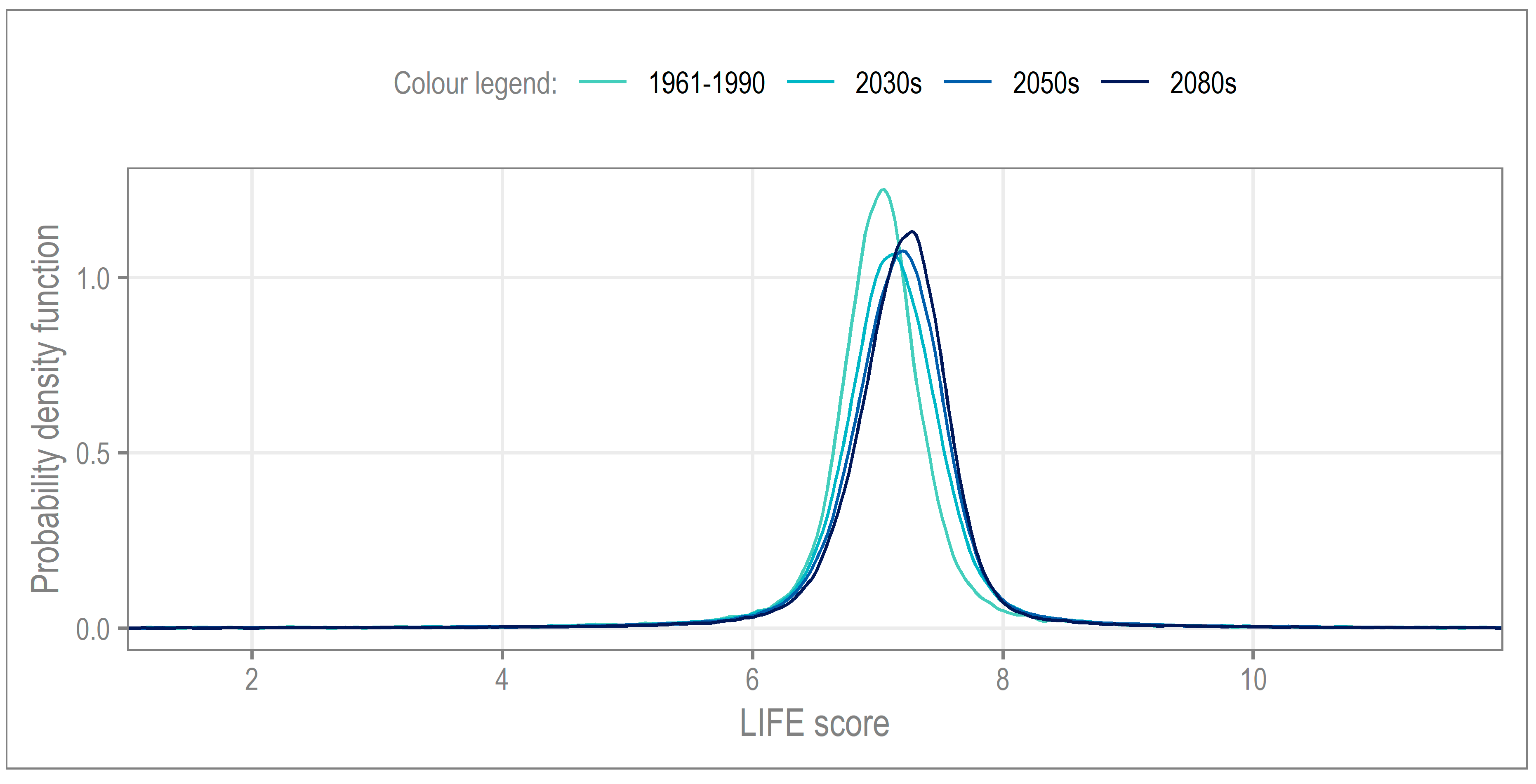
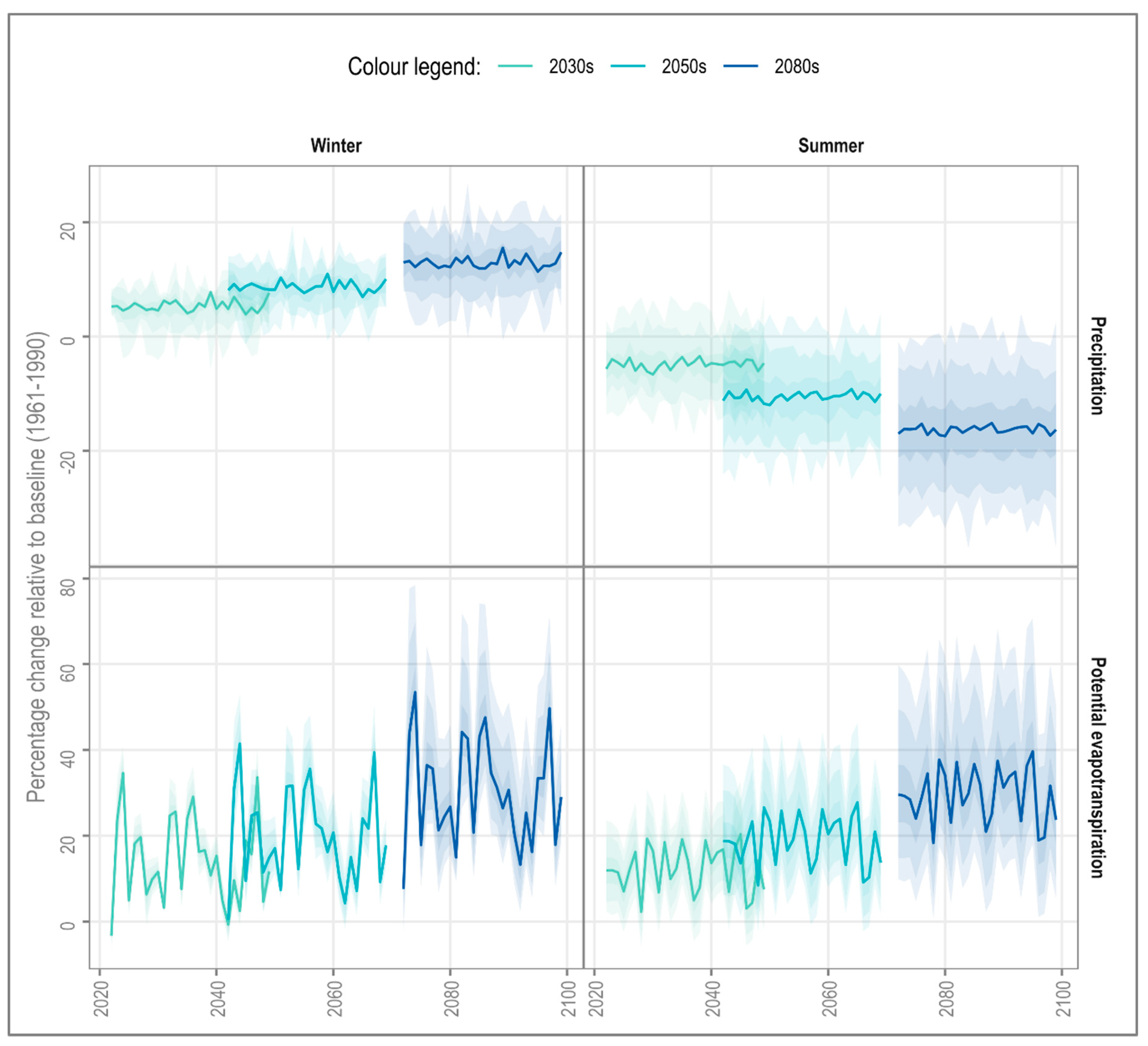
4.1. Long-Term Change
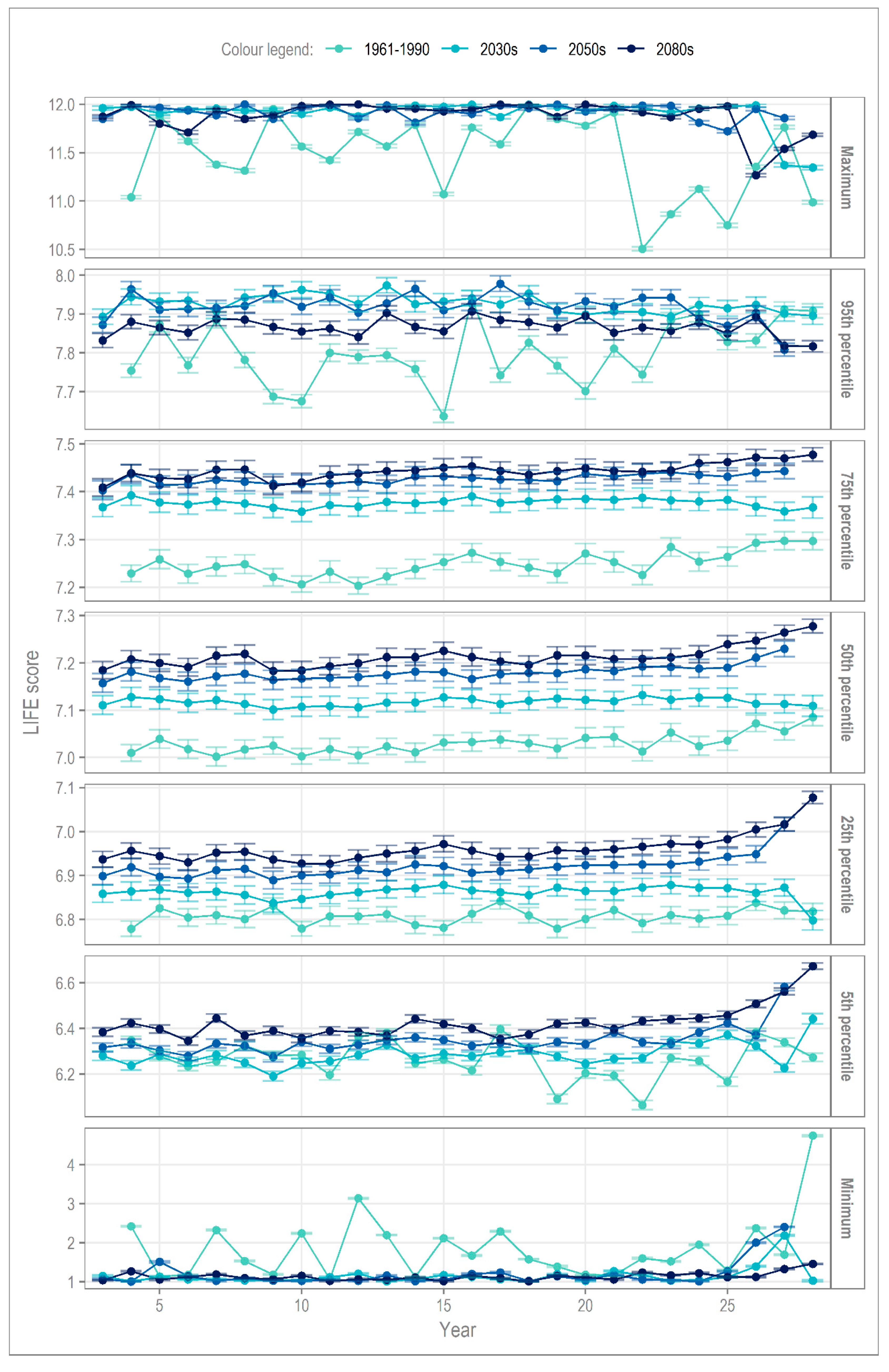
4.2. Interannual Variability
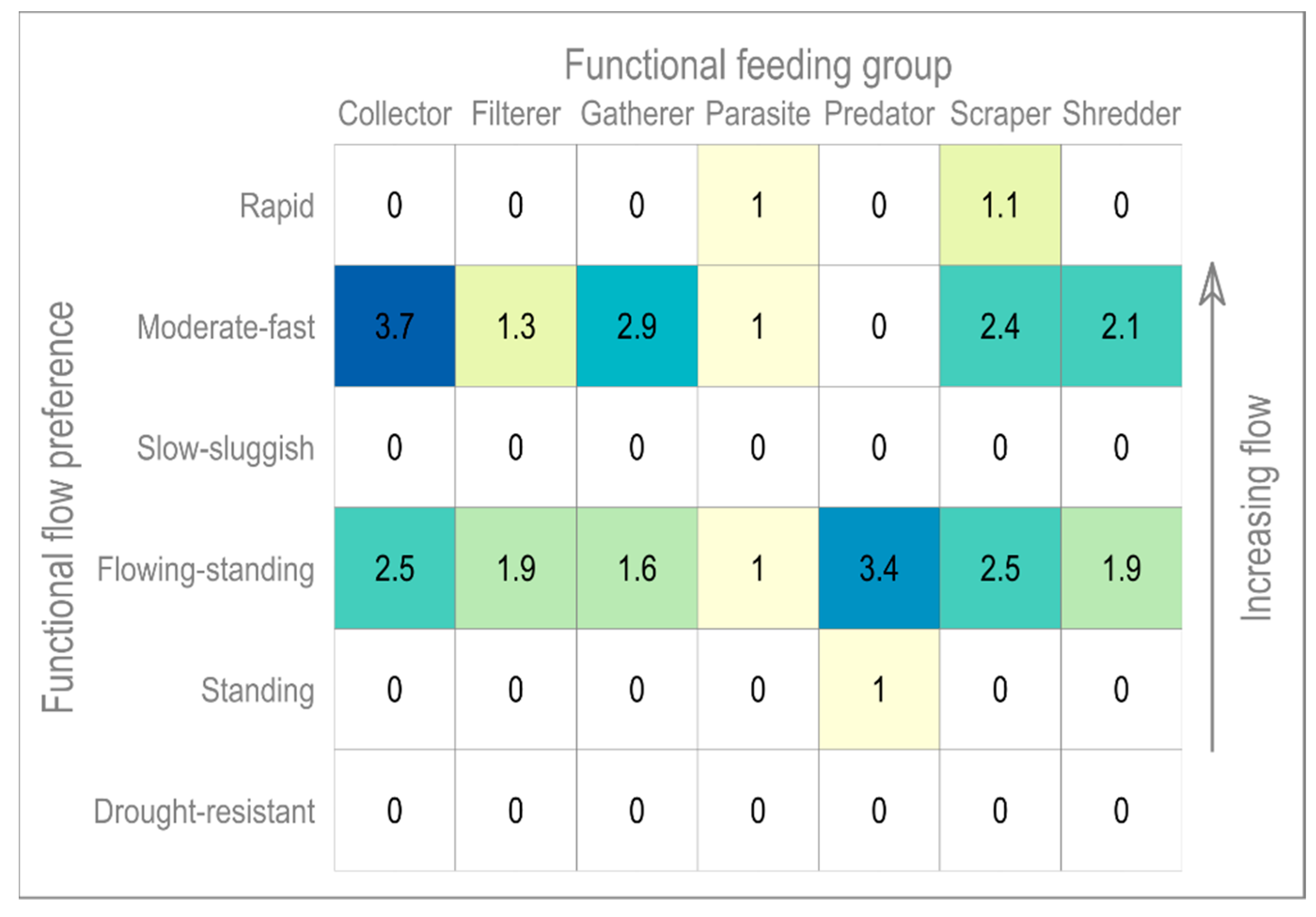
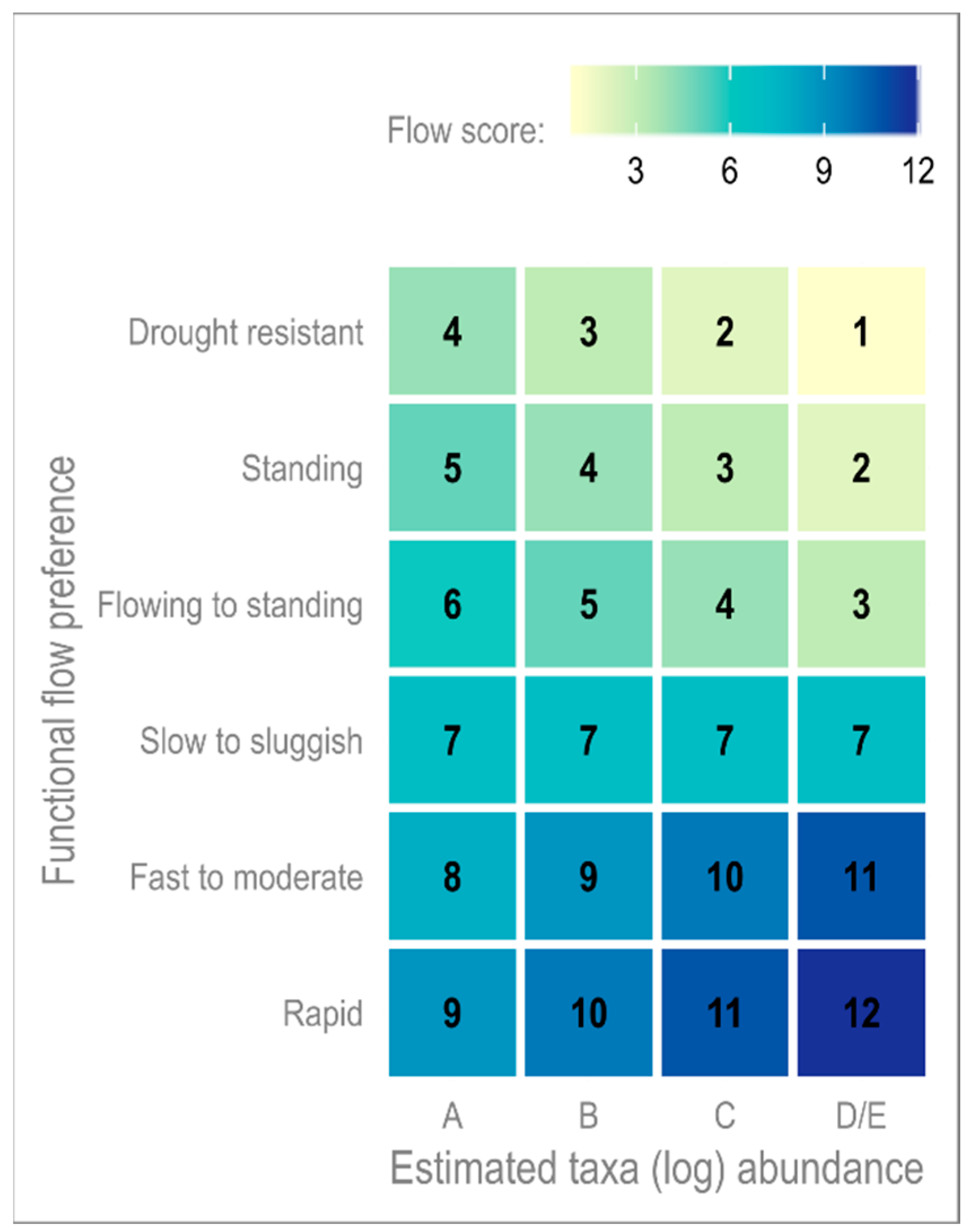
4.3. Functional Matrix
5. Discussion
5.1. Impact of Climate Change
5.2. Uncertainty
5.3. Enhancing and Encouraging Ecological Resilience in Chalk Streams
6. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Definition |
|---|---|
| CEDA | Centre for Environmental Data Analysis (UK) |
| CPOM | Coarse particulate organic matter |
| CMIP | Coupled Model Intercomparison Project |
| EA | Environment Agency (UK) |
| FPOM | Fine particulate organic matter |
| GR4J | Genie Rural a 4 parametres Journalier |
| IPCC | Intergovernmental Panel on Climate Change |
| IQR | Interquartile range |
| LIFE | Lotic-invertebrate index for flow evaluation |
| Probability density function | |
| SSSI | Site of Special Scientific Interest (UK) |
| SRES | Special Report on Emissions Scenarios |
| UKCP09 | UK Climate Projections 2009 |
| WFD | Water Framework Directive (EU) |
| WWF | World Wildlife Fund |

| Functional Feeding Group | Description |
|---|---|
| Collector | A broad grouping generally capturing both filterers and gatherers. |
| Filterer | Filter suspended FPOM from the water column. |
| Gatherer | Gather FPOM settled on the substrate. |
| Parasite | Taxa which do not fit into other groups. |
| Predator | Carnivorous macroinvertebrates which prey on smaller invertebrates. |
| Scraper | Consumers of food sources attached to the substrate; e.g., algae and biofilm. |
| Shredder | Shred and consume plant material such as leaf litter and wood. |


| Index Name | Hydrological Season | Time-Offset | Unit | Description |
|---|---|---|---|---|
| 10R90Logw,t-0 | Winter | t-0 | - | Ratio of log-transformed low to high flows: log(P10)/log(P90). Log-transformation represents the log-normal distribution of flow. |
| revPoss,t-1 | Summer | t-1 | days | Number of days when flow is increasing (positive reversals). |
| Q80Q50s,t-0 | Summer | t-0 | - | Characterisation of moderate low flows; Q80 relative to the median. |
| logQVars,t-1 | Summer | t-1 | m3s−1 | Variance in log flows. |
| Q90Q50s,t-1 | Summer | t-1 | - | Characterisation of low flows; Q90 relative to the median. |
| Q70Q50s,t-1 | Summer | t-1 | - | Characterisation of moderate low flows; Q70 relative to the median. |
| riseMnw,t-0 | Winter | t-0 | m3s−1 | Mean rise rate in flow. |
References
- United Nations. Convention on Biological Diversity—Article 2. Use of Terms. Available online: https://www.cbd.int/doc/legal/cbd-en.pdf (accessed on 18 December 2018).
- Balvanera, P.; Siddique, I.; Dee, L.; Paquette, A.; Isbell, F.; Gonzalez, A.; Byrnes, J.; O’Connor, M.I.; Hungate, B.A.; Griffin, J.N. Linking Biodiversity and Ecosystem Services: Current Uncertainties and the Necessary Next Steps. BioScience 2014, 64, 49–57. [Google Scholar] [CrossRef]
- Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Grime, J.P.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.; Schmid, B.; et al. Biodiversity and Ecosystem Functioning: Current Knowledge and Future Challenges. Science 2001, 294, 804. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59. [Google Scholar] [CrossRef] [PubMed]
- Chapin Iii, F.S.; Zavaleta, E.S.; Eviner, V.T.; Naylor, R.L.; Vitousek, P.M.; Reynolds, H.L.; Hooper, D.U.; Lavorel, S.; Sala, O.E.; Hobbie, S.E.; et al. Consequences of changing biodiversity. Nature 2000, 405, 234. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F.S.; Walker, B.H.; Hobbs, R.J.; Hooper, D.U.; Lawton, J.H.; Sala, O.E.; Tilman, D. Biotic Control over the Functioning of Ecosystems. Science 1997, 277, 500–504. [Google Scholar] [CrossRef] [Green Version]
- Woodward, G.; Papantoniou, G.; Edwards, F.; Lauridsen, R.B. Trophic Trickles and Cascades in a Complex Food Web: Impacts of a Keystone Predator on Stream Community Structure and Ecosystem Processes. Oikos 2008, 117, 683–692. [Google Scholar] [CrossRef]
- Martens, K. The International Year of Biodiversity. Hydrobiologia 2009, 637, 1. [Google Scholar] [CrossRef]
- Allen, G.H.; Pavelsky, T.M. Global extent of rivers and streams. Science 2018. [Google Scholar] [CrossRef]
- Vié, J.-C.; Hilton-Taylor, C.; Stuart, S.N.E. Wildlife in a Changing World—An Analysis of the 2008 IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2009; p. 180.
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.-I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.-H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2007, 81, 163–182. [Google Scholar] [CrossRef]
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.R.; et al. Global threats to human water security and river biodiversity. Nature 2010, 467, 555. [Google Scholar] [CrossRef]
- Gilvear, D.J.; Beevers, L.C.; O’Keeffe, J.; Acreman, M. Environmental Water Regimes and Natural Capital: Free-Flowing Ecosystem Services. In Water for the Environment; Academic Press: Cambridge, MA, USA, 2017; Chapter 8; pp. 151–171. [Google Scholar] [CrossRef]
- WWF. Living Planet Report 2016. Risk and Resilience in a New era; WWF International: Gland, Switzerland, 2016. [Google Scholar]
- Minshall, G.W.; Petersen, R.C.; Cummins, K.W.; Bott, T.L.; Sedell, J.R.; Cushing, C.E.; Vannote, R.L. Interbiome Comparison of Stream Ecosystem Dynamics. Ecol. Monogr. 1983, 53, 1–25. [Google Scholar] [CrossRef]
- Wallace, J.B.; Webster, J.R. The role of macroinvertebrates in stream ecosystem function. Annu. Rev. Entomol. 1996, 41, 115–139. [Google Scholar] [CrossRef] [PubMed]
- Mendel, R.J. Benthic Macroinvertebrates. Available online: http://enviroscienceinc.com/benthic-macroinvertebrates/ (accessed on 24 October 2013).
- EA. Water Framework Directive—Method Statement for the Classification of Surface Water Bodies v3 (2012 Classification Release); Environment Agency: Bristol, UK, 2013.
- Acreman, M.; Dunbar, M.; Hannaford, J.; Mountford, O.; Wood, P.; Holmes, N.; Wx, I.C.; Noble, R.; Extence, C.; Aldrick, J.; et al. Developing environmental standards for abstractions from UK rivers to implement the EU Water Framework Directive / Développement de standards environnementaux sur les prélèvements d’eau en rivière au Royaume Uni pour la mise en œuvre de la directive cadre sur l’eau de l’Union Européenne. Hydrol. Sci. J. 2008, 53, 1105–1120. [Google Scholar] [CrossRef]
- Cummins, K.W. Invertebrates. In The Rivers Handbook; Calow, P., Petts, G.E., Eds.; Blackwell Scientific: Oxford, UK, 1995; pp. 234–250. [Google Scholar]
- Nock, C.A.; Vogt, R.J.; Beisner, B.E. Functional Traits. eLS 2016. [Google Scholar] [CrossRef]
- White, J.C.; Hannah, D.M.; House, A.; Beatson, S.J.V.; Martin, A.; Wood, P.J. Macroinvertebrate responses to flow and stream temperature variability across regulated and non-regulated rivers. Ecohydrology 2017, 10, e1773. [Google Scholar] [CrossRef]
- Acreman, M.C.; Dunbar, M.J. Defining environmental river flow requirements ? A review. Hydrol. Earth Syst. Sci. Discuss. 2004, 8, 861–876. [Google Scholar] [CrossRef]
- Lake, P.S. Resistance, Resilience and Restoration. Ecol. Manag. Restor. 2013, 14, 20–24. [Google Scholar] [CrossRef]
- Lytle, D.A.; Poff, N.L. Adaptation to natural flow regimes. Trends Ecol. Evol. 2004, 19, 94–100. [Google Scholar] [CrossRef]
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The Natural Flow Regime. BioScience 1997, 47, 769–784. [Google Scholar] [CrossRef] [Green Version]
- Poff, N.L.; Zimmerman, J.K.H. Ecological responses to altered flow regimes: A literature review to inform the science and management of environmental flows. Freshw. Biol. 2010, 55, 194–205. [Google Scholar] [CrossRef]
- Visser, A.; Beevers, L.; Patidar, S. Macro-invertebrate Community Response to Multi-annual Hydrological Indicators. River Res. Appl. 2017, 33, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Marshall, J.C.; Negus, P.M. Application of a Multistressor Risk Framework to the Monitoring, Assessment, and Diagnosis of River Health. In Multiple Stressors in River Ecosystems; Sabater, S., Elosegi, A., Ludwig, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; Chapter 15; pp. 255–280. [Google Scholar]
- O’Neill, R.; Hughes, K. The State of England’s Chalk Streams; WWF-UK: Surrey, UK, 2014. [Google Scholar]
- Mann, R.H.K.; Blackburn, J.H.; Beaumont, W.R.C. The ecology of brown trout Salmo trutta in English chalk streams. Freshw. Biol. 1989, 21, 57–70. [Google Scholar] [CrossRef]
- Berrie, A.D. The chalk-stream environment. Hydrobiologia 1992, 248, 3–9. [Google Scholar] [CrossRef]
- Norfolk Rivers Trust. The River Nar—A Water Framework Directive Local Catchment Plan; Norfolk Rivers Trust: Norfolk, UK, 2014. [Google Scholar]
- Environment Agency; English Nature. The State of England’s Chalk Rivers: A Report by the UK Biodiversity Action Plan Steering Group for Chalk Rivers; Environment Agency: Bristol, UK, 2004.
- Collet, L.; Harrigan, S.; Prudhomme, C.; Formetta, G.; Beevers, L. Future hot-spots for hydro-hazards in Great Britain: A probabilistic assessment. Hydrol. Earth Syst. Sci. 2018, 22, 5387–5401. [Google Scholar] [CrossRef]
- Visser, A.; Beevers, L.; Collet, L.; Formetta, G.; Smith, K.; Wanders, N.; Thober, S.; Pan, M.; Kumar, R. Spatio-temporal analysis of compound hydro-hazard extremes across the UK. Adv. Water Resour. 2019. under review. [Google Scholar]
- Rounsevell, M.; Fischer, M.; Boeraeve, F.; Jacobs, S.; Liekens, I.; Marques, A.; Molnár, Z.; Osuchova, J.; Shkaruba, A.; Whittingham, M.; et al. Setting the scene. In The IPBES Regional Assessment Report on Biodiversity and Ecosystem Services for Europe and Central Asia; Rounsevell, M., Fischer, M., Torre-Marin Rando, A., Mader, A., Eds.; Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services, Bonn: Bonn, Germany, 2018; Chapter 8. [Google Scholar]
- Klaar, M.J.; Dunbar, M.J.; Warren, M.; Soley, R. Developing hydroecological models to inform environmental flow standards: A case study from England. Wiley Interdiscip. Rev. Water 2014, 1, 207–217. [Google Scholar] [CrossRef]
- Gleick, P.H. Water strategies for the next administration. Science 2016, 354, 555–556. [Google Scholar] [CrossRef]
- Gleick, P.H. Water in crisis: Paths to sustainable water use. Ecol. Appl. 1998, 8, 571–579. [Google Scholar] [CrossRef]
- Davis, J.; O’Grady, A.P.; Dale, A.; Arthington, A.H.; Gell, P.A.; Driver, P.D.; Bond, N.; Casanova, M.; Finlayson, M.; Watts, R.J.; et al. When trends intersect: The challenge of protecting freshwater ecosystems under multiple land use and hydrological intensification scenarios. Sci. Total Environ. 2015, 534, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Visser, A.; Beevers, L.; Patidar, S. A coupled modelling framework to assess the hydroecological impact of climate change. Environ. Model. Softw. 2019, 114, 12–28. [Google Scholar] [CrossRef]
- Mastrandrea, M.D.; Field, C.B.; Stocker, T.F.; Edenhofer, O.; Ebi, K.L.; Frame, D.J.; Held, H.; Kriegler, E.; Mach, K.J.; Matschoss, P.R. Guidance Note for Lead Authors of the IPCC Fifth Assessment Report on Consistent Treatment of Uncertainties; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2010. [Google Scholar]
- Sear, D.A.; Newson, M.; Old, J.C.; Hill, C. Geomorphological Appraisal of the River Nar Site of Special Scientific Interest; N684; English Nature: Peterborough, UK, 2005.
- Natural England. Designation 1006323—River Nar, West Norfolk, Norfolk. Available online: https://designatedsites.naturalengland.org.uk/PDFsForWeb/Citation/1006323.pdf (accessed on 9 December 2018).
- NRFA. 33007—Nar at Marham—Gauged Daily Flow (1953–2017). Available online: http://nrfa.ceh.ac.uk/data/station/meanflow/33007 (accessed on 15 December 2018).
- Arnell, N.W. Factors controlling the effects of climate change on river flow regimes in a humid temperate environment. J. Hydrol. 1992, 132, 321–342. [Google Scholar] [CrossRef]
- Rangeley-Wilson, C. The River Nar—A Water Framework Directive Local Catchment Plan; Norfolk Rivers Trust: Norfolk, UK, 2012. [Google Scholar]
- Bertholdt, N. River Nar SSSI. Available online: https://designatedsites.naturalengland.org.uk/SiteDetail.aspx?SiteCode=S1006323&SiteName=river (accessed on 15 December 2018).
- Garbe, J.; Beevers, L.; Pender, G. The interaction of low flow conditions and spawning brown trout (Salmo trutta) habitat availability. Ecol. Eng. 2016, 88, 53–63. [Google Scholar] [CrossRef]
- Extence, C.A.; Balbi, D.M.; Chadd, R.P. River flow indexing using British benthic macroinvertebrates: A framework for setting hydroecological objectives. Regul. Rivers Res. Manag. 1999, 15, 545–574. [Google Scholar] [CrossRef]
- Durance, I.; Bruford, M.W.; Chalmers, R.; Chappell, N.A.; Christie, M.; Cosby, B.J.; Noble, D.; Ormerod, S.J.; Prosser, H.; Weightman, A.; et al. The Challenges of Linking Ecosystem Services to Biodiversity: Lessons from a Large-Scale Freshwater Study. In Advances in Ecological Research; Woodward, G., Bohan, D.A., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 54, Chapter Three; pp. 87–134. [Google Scholar] [CrossRef]
- Ncube, S.; Beevers, L.; Adeloye, A.J.; Visser, A. Assessment of freshwater ecosystem services in the Beas River Basin, Himalayas region, India. Proc. IAHS 2018, 379, 67–72. [Google Scholar] [CrossRef]
- EA. River Nar Macroinvertebrate Monitoring Data. Available upon request from the Environment Agency.
- Kay, A.L.; Jones, R.G. Comparison of the use of alternative UKCP09 products for modelling the impacts of climate change on flood frequency. Clim. Chang. 2012, 114, 211–230. [Google Scholar] [CrossRef] [Green Version]
- Murphy, J.M.; Sexton, D.M.H.; Jenkins, G.J.; Booth, B.B.B.; Brown, C.C.; Clark, R.T.; Collins, M.; Harris, G.R.; Kendon, E.J.; Betts, R.A.; et al. UK Climate Projections Science Report: Climate Change Projections; Met Office Hadley Centre: Exeter, UK, 2009.
- Wigley, T.M.L. Climatology: Impact of extreme events. Nature 1985, 316, 106–107. [Google Scholar] [CrossRef]
- Schlabing, D.; Frassl, M.A.; Eder, M.M.; Rinke, K.; Bárdossy, A. Use of a weather generator for simulating climate change effects on ecosystems: A case study on Lake Constance. Environ. Model. Softw. 2014, 61, 326–338. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Murphy, J.; Sexton, D. Improvements to the UKCP09 Land Projection Data; Met Office Hadley Centre: Exeter, UK, 2013. [Google Scholar]
- UKCP09. Validation of Weather Generator Outputs; UK Climate Projections 2009: Exeter, UK, 2011.
- Perrin, C.; Michel, C.; Andréassian, V. Improvement of a parsimonious model for streamflow simulation. J. Hydrol. 2003, 279, 275–289. [Google Scholar] [CrossRef]
- Coron, L.; Thirel, G.; Delaigue, O.; Perrin, C.; Andréassian, V. The suite of lumped GR hydrological models in an R package. Environ. Model. Softw. 2017, 94, 166–171. [Google Scholar] [CrossRef]
- Visser, A.; Beevers, L.; Patidar, S. Replication of ecologically relevant hydrological indicators following a covariance approach to hydrological model parameterisation. Hydrol. Earth Syst. Sci. Discuss. 2018, 2018, 1–24. [Google Scholar] [CrossRef]
- Seibert, J. Multi-criteria calibration of a conceptual runoff model using a genetic algorithm. Hydrol. Earth Syst. Sci. 2000, 4, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Euser, T.; Winsemius, H.C.; Hrachowitz, M.; Fenicia, F.; Uhlenbrook, S.; Savenije, H.H.G. A framework to assess the realism of model structures using hydrological signatures. Hydrol. Earth Syst. Sci. 2013, 17, 1893–1912. [Google Scholar] [CrossRef] [Green Version]
- Richter, B.D.; Baumgartner, J.V.; Powell, J.; Braun, D.P. A Method for Assessing Hydrologic Alteration within Ecosystems. Conserv. Biol. 1996, 10, 1163–1174. [Google Scholar] [CrossRef]
- Visser, A.; Beevers, L.; Patidar, S. Complexity in hydroecological modelling, a comparison of stepwise selection and information theory. River Res. Appl. 2018. [Google Scholar] [CrossRef]
- Jones, P.; Harpham, C.; Kilsby, C.; Glenis, V.; Burton, A. UK Climate Projections Science Report: Projections of Future Daily Climate for the UK from the Weather Generator; DEFRA: London, UK, 2010.
- Westfall, P.H. Kurtosis as Peakedness, 1905–2014. R.I.P. Am. Stat. 2014, 68, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Met Office. Met Office: Regional Climates: Eastern England. Available online: http://www.metoffice.gov.uk/climate/uk/ee/ (accessed on 30 December 2013).
- Wood, P.J.; Petts, G.E. Low flows and recovery of macroinvertebrates in a small regulated chalk stream. Regul. Rivers: Res. Manag. 1994, 9, 303–316. [Google Scholar] [CrossRef]
- Lake, P.S. Ecological effects of perturbation by drought in flowing waters. Freshw. Biol. 2003, 48, 1161–1172. [Google Scholar] [CrossRef] [Green Version]
- Everard, M. The importance of periodic droughts for maintaining diversity in the freshwater environment. Freshw. Forum 1996, 7, 33–50. [Google Scholar]
- Garbe, J.; Beevers, L. Modelling the impacts of a water trading scheme on freshwater habitats. Ecol. Eng. 2017, 105, 284–295. [Google Scholar] [CrossRef]
- Erfani, T.; Binions, O.; Harou, J.J. Protecting environmental flows through enhanced water licensing and water markets. Hydrol. Earth Syst. Sci. 2015, 19, 675–689. [Google Scholar] [CrossRef] [Green Version]







| Order, Latin Name (Common Name) | No. Species per Order |
|---|---|
| Coleoptera (Beetles) | 35 |
| Diptera (True flies) | 3 |
| Ephemeroptera (Mayfly) | 17 |
| Gastropoda c. (Snails and slugs) | 19 |
| Hemiptera (True bugs) | 14 |
| Odonata (Dragonfly and damselfly) | 8 |
| Trichoptera (Caddisfly) | 52 |
| Other (13 orders) | 40 |
| Total | 188 |
| 1961–1990 | 2030s | 2050s | 2080s | |
|---|---|---|---|---|
| Standard deviation | 0.68 | 0.72 | 0.7 | 0.65 |
| Excess kurtosis | 12.43 | 9.75 | 10.57 | 12.9 |
| Skewness | −0.86 | −0.76 | −0.83 | −1 |
| 2030s | 2050s | 2080s | |
|---|---|---|---|
| 95th percentile | −92 | −81 | −91 |
| 5th percentile | −65 | −52 | −31 |
| Maximum | −84 | −97 | −83 |
| Minimum | −92 | −84 | −98 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Visser, A.; Beevers, L.; Patidar, S. The Impact of Climate Change on Hydroecological Response in Chalk Streams. Water 2019, 11, 596. https://doi.org/10.3390/w11030596
Visser A, Beevers L, Patidar S. The Impact of Climate Change on Hydroecological Response in Chalk Streams. Water. 2019; 11(3):596. https://doi.org/10.3390/w11030596
Chicago/Turabian StyleVisser, Annie, Lindsay Beevers, and Sandhya Patidar. 2019. "The Impact of Climate Change on Hydroecological Response in Chalk Streams" Water 11, no. 3: 596. https://doi.org/10.3390/w11030596
APA StyleVisser, A., Beevers, L., & Patidar, S. (2019). The Impact of Climate Change on Hydroecological Response in Chalk Streams. Water, 11(3), 596. https://doi.org/10.3390/w11030596