1. Introduction
Under the Convention on Biological Diversity, biodiversity is defined as the variability among living organisms, within & between species and ecosystems [
1,
2]. Within the public sphere, reasons for preserving biodiversity are, frequently, purely aesthetic, cultural and economic [
3]. Critically, the societal cost of biodiversity loss, in terms of ecosystem functionality, may be severe. In recent years, significant progress has been made towards understanding this dependency [
2,
3]; if not universal, broad consensus points include [
4]:
Termed the freshwater paradox, freshwaters are disproportionately rich in biodiversity [
8]. Rivers and streams cover approximately 0.58% of the world’s (nonglacial) surface [
9], yet up to 7% of species make freshwaters their home [
10,
11]. For humans, freshwater is considered the most essential natural resource [
12]. In addition to water supply, rivers support prosperity, health, and well-being through the provision of ecosystem services; examples include hydro-hazard regulation, water purification and recreation [
13]. Our need for freshwater has seen a rapid decline in freshwater biodiversity; in a 2016 World Wildlife Fund (WWF; see
Table A1 for definitions of all abbreviations used) report [
14] it was estimated that, between 1970 and 2012, freshwater biodiversity declined by 81%, more than double that of terrestrial and marine combined.
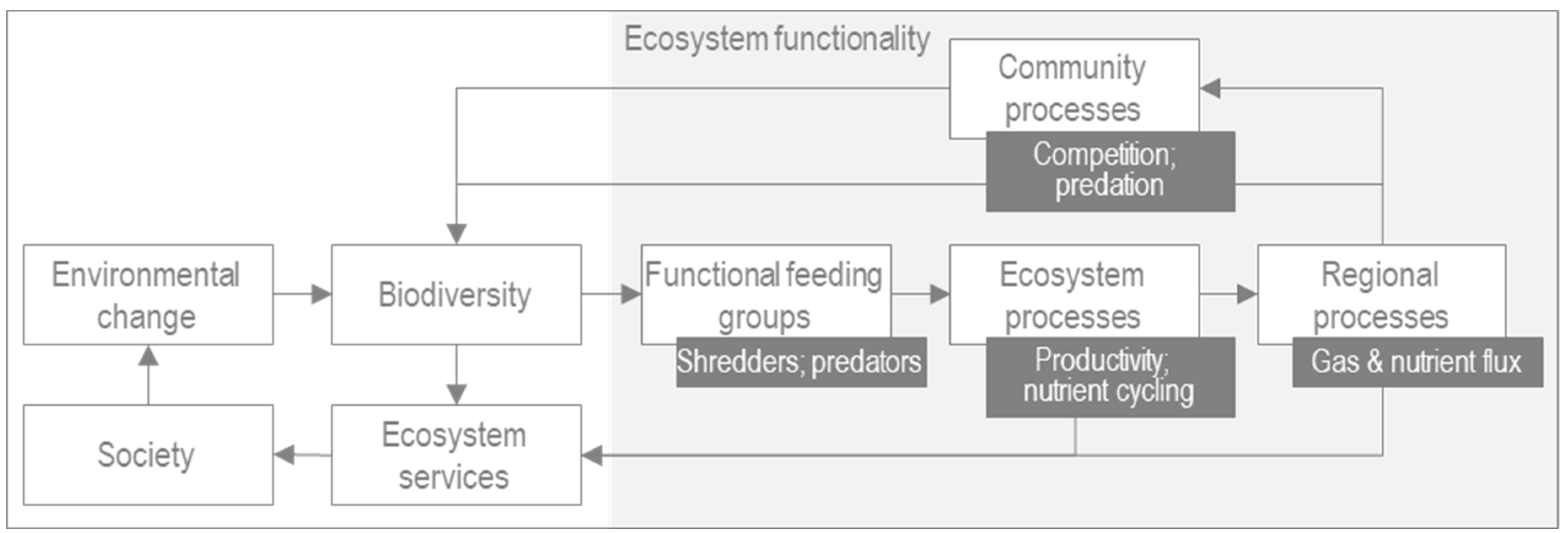
Figure 1 illustrates the impact of environmental change on biodiversity, ecosystem functionality and hence the provision of the vital ecosystem services upon which we depend.
The functional composition of the macroinvertebrate community is a major determinant of ecosystem functionality [
15]. As consumers at intermediate trophic levels, macroinvertebrates exert strong bottom-up and top-down controls [
16]. The above, coupled with their sensitivity to environmental change, makes macroinvertebrates ideal biological and functional indicators [
17,
18,
19].
Macroinvertebrate functional feeding groups describe their consumption of resources [
20], for example, scrapers consume foodstuffs such as algae which are attached to substrate. It is this processing of organic matter which facilitates essential ecosystem processes such as productivity and nutrient cycling [
4,
6], which in turn supports processes at the regional level. Understanding how the composition of the macroinvertebrate community changes helps to understand the ecological processes in a river, thereby aiding understanding for the purposes of conservation and restoration [
21], as well as adaptation to environmental change [
22]; the latter being the focus in this study. Flow is widely acknowledged as a major determinant of the health of the river ecosystem (for example, see [
23,
24,
25,
26,
27]). Data-driven numerical models are used to link flow and hydroecological response in order to understand the instream response to changes in flow [
28]. Arguably, the term river health is more useful for interpretation than hydroecological response [
29]; hereafter, the term river health should be considered interchangeable with hydroecological response.
Chalk streams provide a steady flow of cool, clear and nutrient-rich water whose gravel channels support uniquely “diverse and fecund ecosystems” [
30]. Such streams are famous amongst anglers due to the high levels of fish production that chalk waters are able to support (relative to other river types) [
31]. Charles Rangeley-Wilson [
30] describes the importance of England’s 224 chalk streams as analogous to such biodiversity hotspots as the Great Barrier Reef and equatorial rainforests. Indeed, these streams are (almost entirely) unique to Southern England, with only a handful located in Northern France [
30]. The result of a legacy of historical physical modifications—e.g., for systems of water mills and meadows for irrigation [
32] as well as more recent fisheries management [
33]—75% of English chalk-streams were designated ‘heavily modified water bodies’ under the 2008–2012 River Habitat Surveys [
30]. Following on from their first report on the state of England’s chalk streams a decade prior, the Environment Agency (EA) and WWF-UK concluded that English chalk streams “remain in a shocking state of health” [
30,
34]. With increasing water demand and climatic variability (e.g., increased hydro-hazards [
35,
36]), there are significant questions as to the long-term sustainability of this water resource [
6,
30,
37,
38,
39,
40,
41]. This is of particular concern given the chalk aquifer provides 70% of the public drinking water in south-east England [
30].
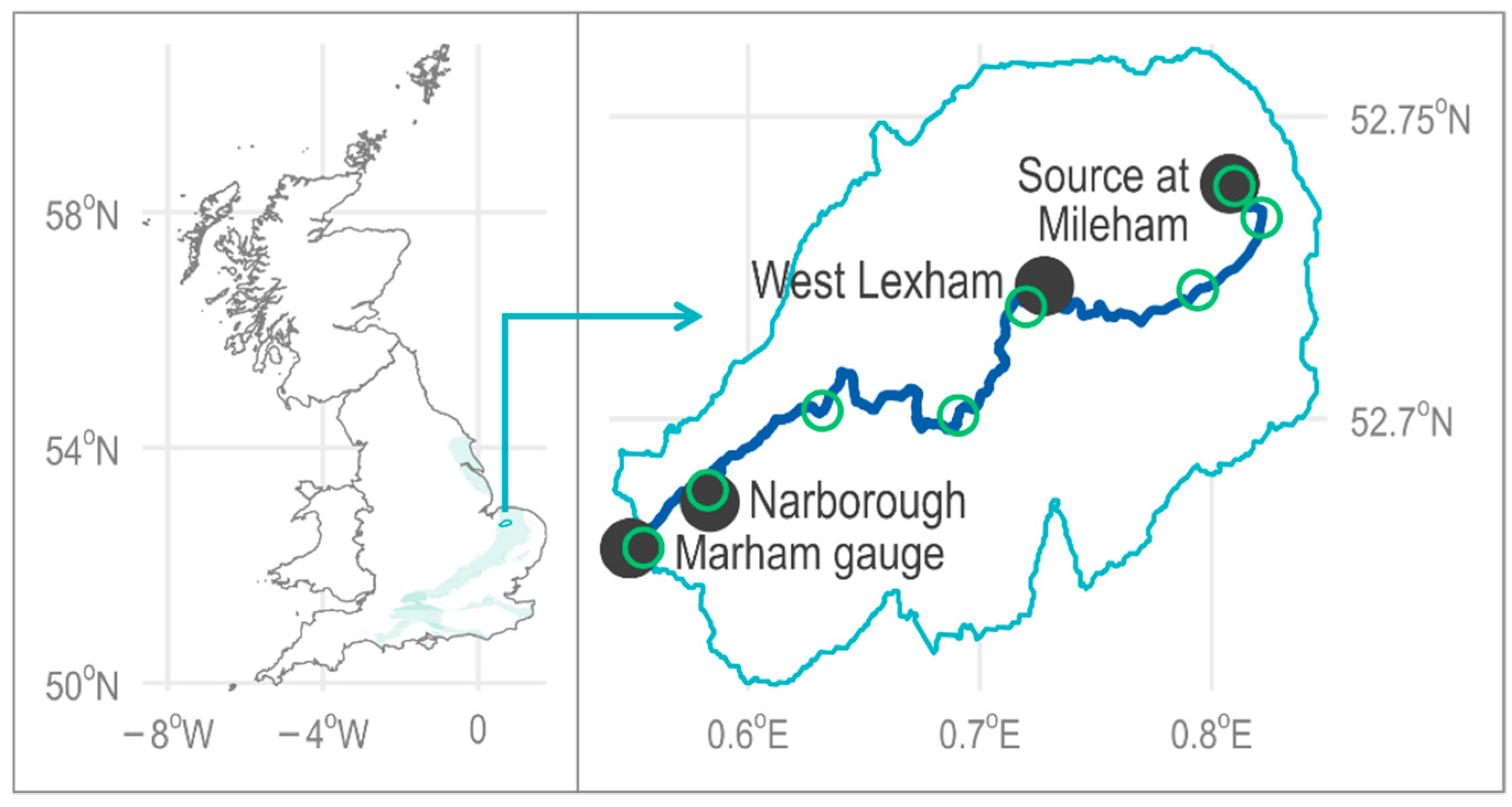
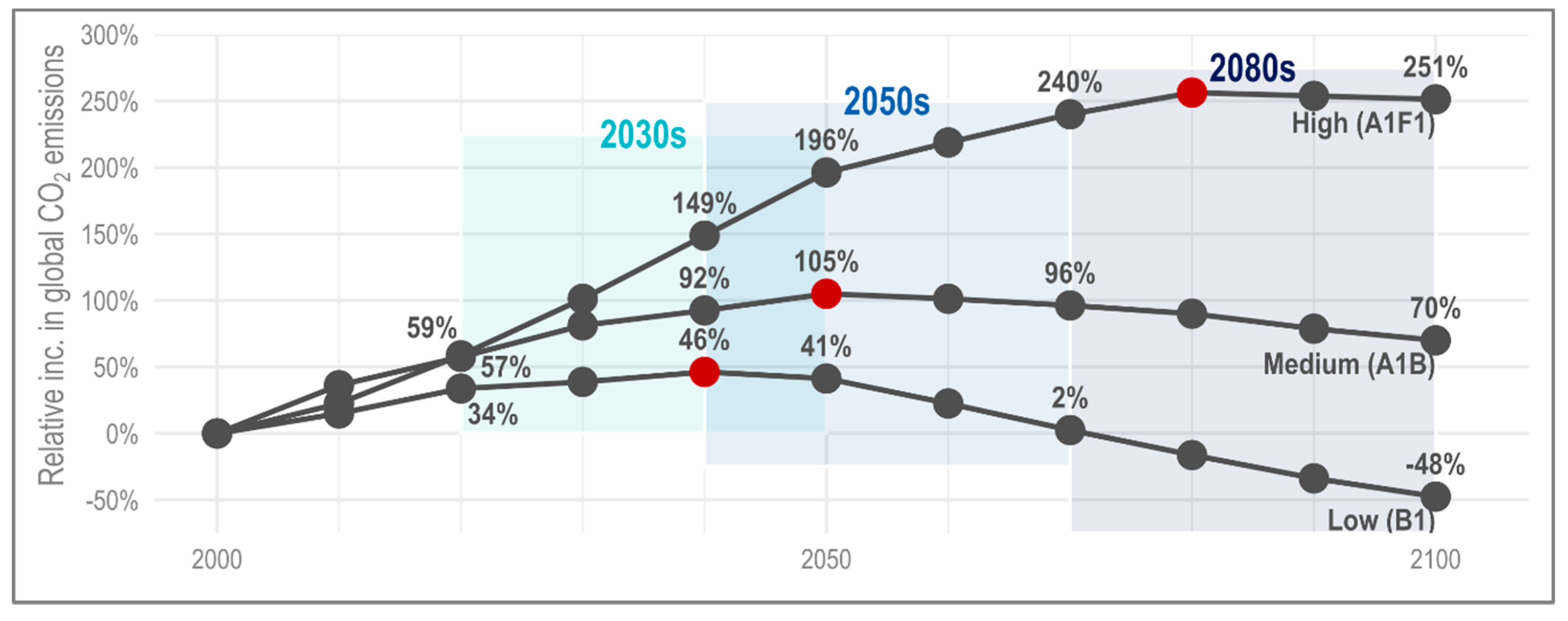
The aim of this paper is to quantify the effect of climate change on the river health of a chalk stream. Methods investigating hydroecological response have, typically, been qualitative in nature or quantitative with limited scope, whilst the effect of uncertainty (e.g., parameter, structural, emissions scenario) is rarely considered [
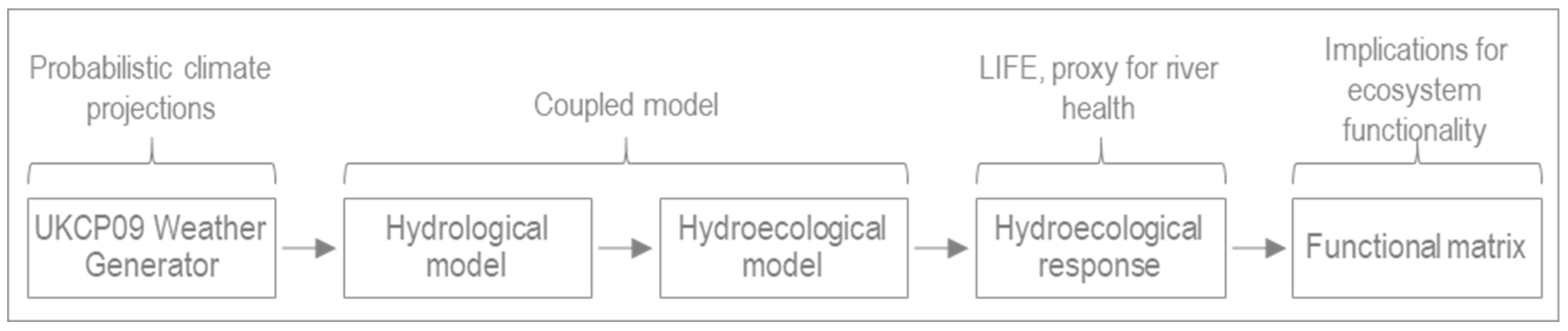
42]. To address this research gap, the author’s proposed a coupled hydrological and hydroecological modelling framework [
42]. The framework was developed using an English chalk stream, the River Nar in Norfolk, where the coupled model was run for a single scenario, CMIP3 SRES A1B high emissions (Coupled Model Intercomparison Project; Special Report on Emissions Scenarios) and 30-year time slice (2041–2070). This paper considers both change in river health over time (from the 2030s to the end of century) as well as the implications for ecosystem functionality. To this end, we consider the same case study river, eliminating the need for model calibration. The UK probabilistic climate projections 2009 (UKCP09) weather generator serves as input to the coupled model; specifically, the high emissions scenario (CMIP3 SRES B1). The results focus on the 99–100% probability, consistent with the Intergovernmental Panel on Climate Change’s (IPCC) definition of a virtually certain outcome [
43]. The wider implications for chalk streams and groundwater-fed rivers more generally are also reflected upon.
4. Results
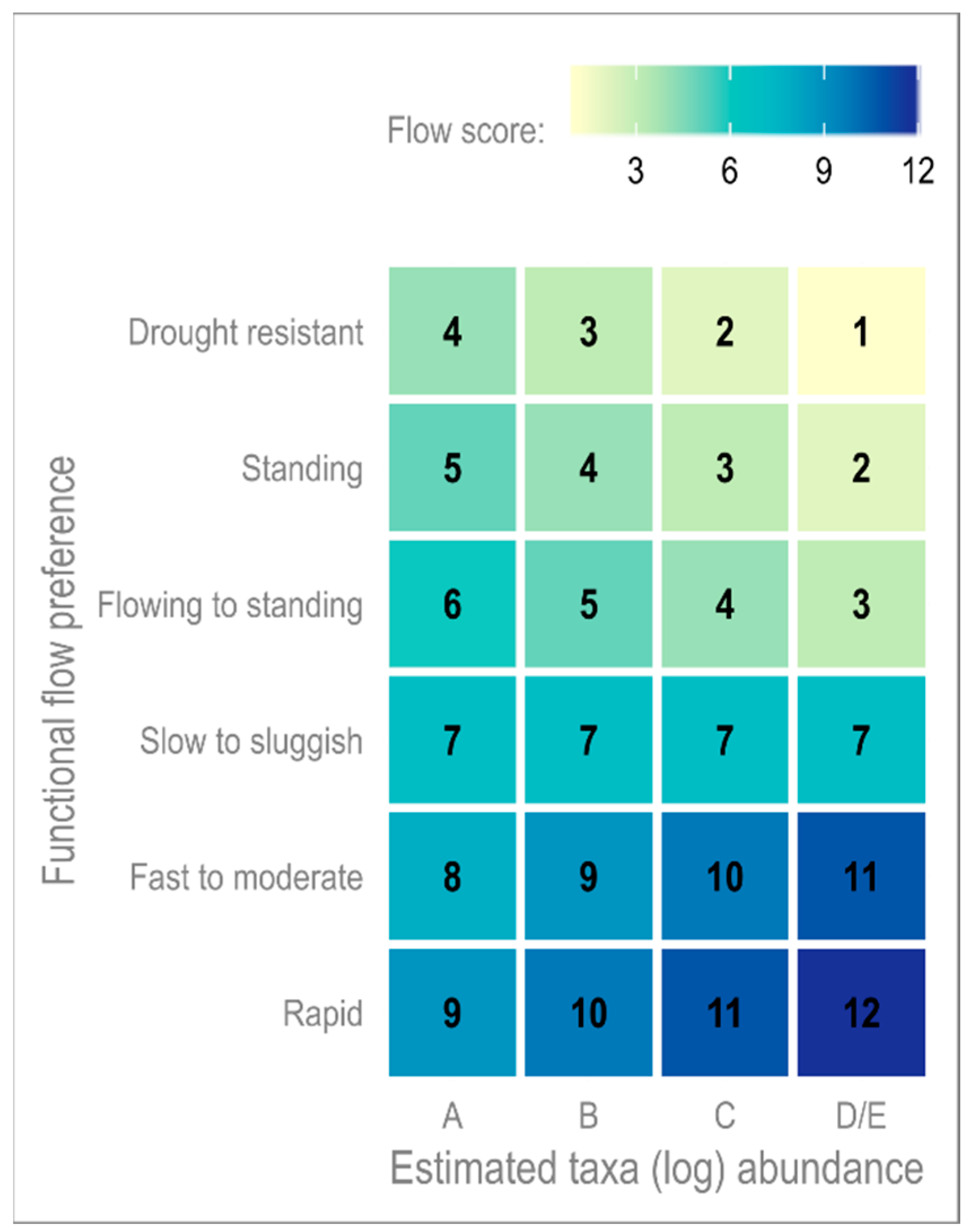
The focus here is on comparison of the distribution of LIFE score, the proxy for river health, over the four time-periods. See
Appendix (
Figure A1) for how to interpret LIFE scores relative to functional flow preference. To provide a general overview of the change over time, the long-term trends (aggregate 30-year time slices;
Section 4.1) are presented first, followed by the interannual change (
Section 4.2) to examine year-on-year variation. Finally, in
Section 4.3, the functional matrix, relating functional flow preferences to feeding groups, is considered in the context of these hydroecological projections.
4.1. Long-Term Change
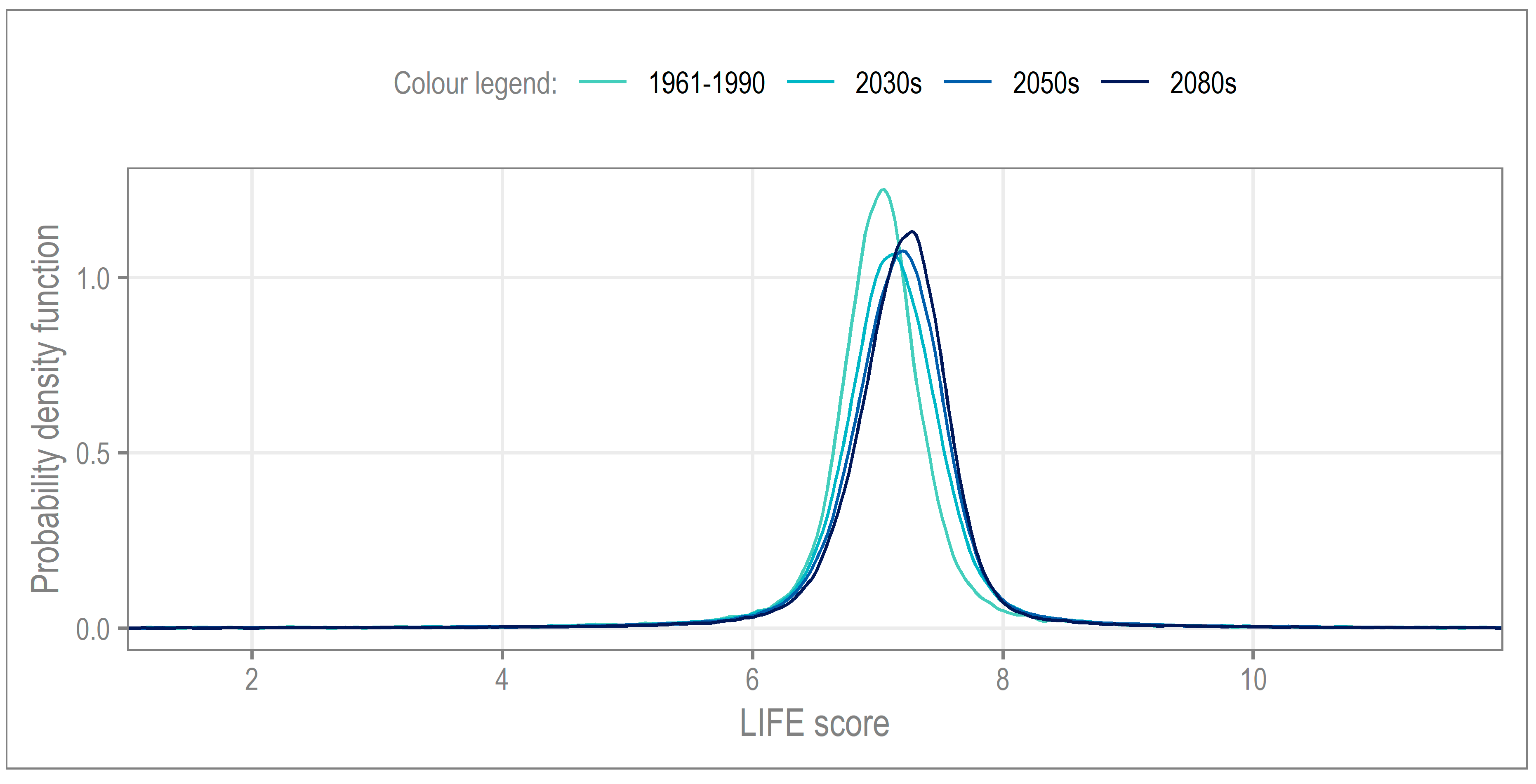
The probability density function (PDF;
Figure 5) provides a visual representation of the LIFE score distribution for each time slice. The baseline distribution, 1961–1990, sees LIFE scores centered on ~7 (functional flow preference slow to sluggish). From the baseline to the 2030s, the reduction in this clustering coincides with an increase in LIFE scores, whilst the change from the 2030s to 2050s is less marked. The trend for increasing LIFE scores continues into the 2080s where the clustering of LIFE scores can be seen to increase again.
To elucidate further, we consider the standard deviation, as well as the measures of distribution excess kurtosis and skewness (
Table 2). The standard deviation reveals an initial increase in variance (2030s), with a subsequent decrease to below the baseline level by the 2080s, suggestive of a slight increase in low probability hydroecological responses by the end of the century. However, the difference across the time slices is relatively small, indicating a limited change in the central distribution of hydroecological response overall.
Note that for excess kurtosis and skewness, comparisons from baseline to future are not possible, due to differences in sample size (n = 1000 on baseline [
69] p. 24). Excess kurtosis is a measure of the combined weight of the tails relative to the normal distribution; for example, a negative value means that more of the dataset is located in the tails than the normal distribution (note that kurtosis is often misinterpreted as a measure of peakedness [
70]).
Table 2 shows that, for all four time periods, the weight is not located in the tails (hence the observed clustering in
Figure 5 previously).
Table 2 shows that the change in kurtosis from the 2030s to 2050s, −0.07, is more than half that of the 2050s to 2080s, −0.17. Skewness, a measure of the symmetry in the distribution, shows that all four time-periods are right-skewed; here, the increase from 2050s to 2080s is almost 3 times that of 2030s to 2050s.
In summary, the aggregated projections indicate a very limited change in the mean hydroecological response under climate change. However,
Table 2 does highlight that, by the end-of-century, there may be a restructuring of the macroinvertebrate community response to low-probability events. Note that, the smaller scale of change observed between the 2030s to 2050s may be explained by the overlap between these two time slices (2041–2050).
4.2. Interannual Variability
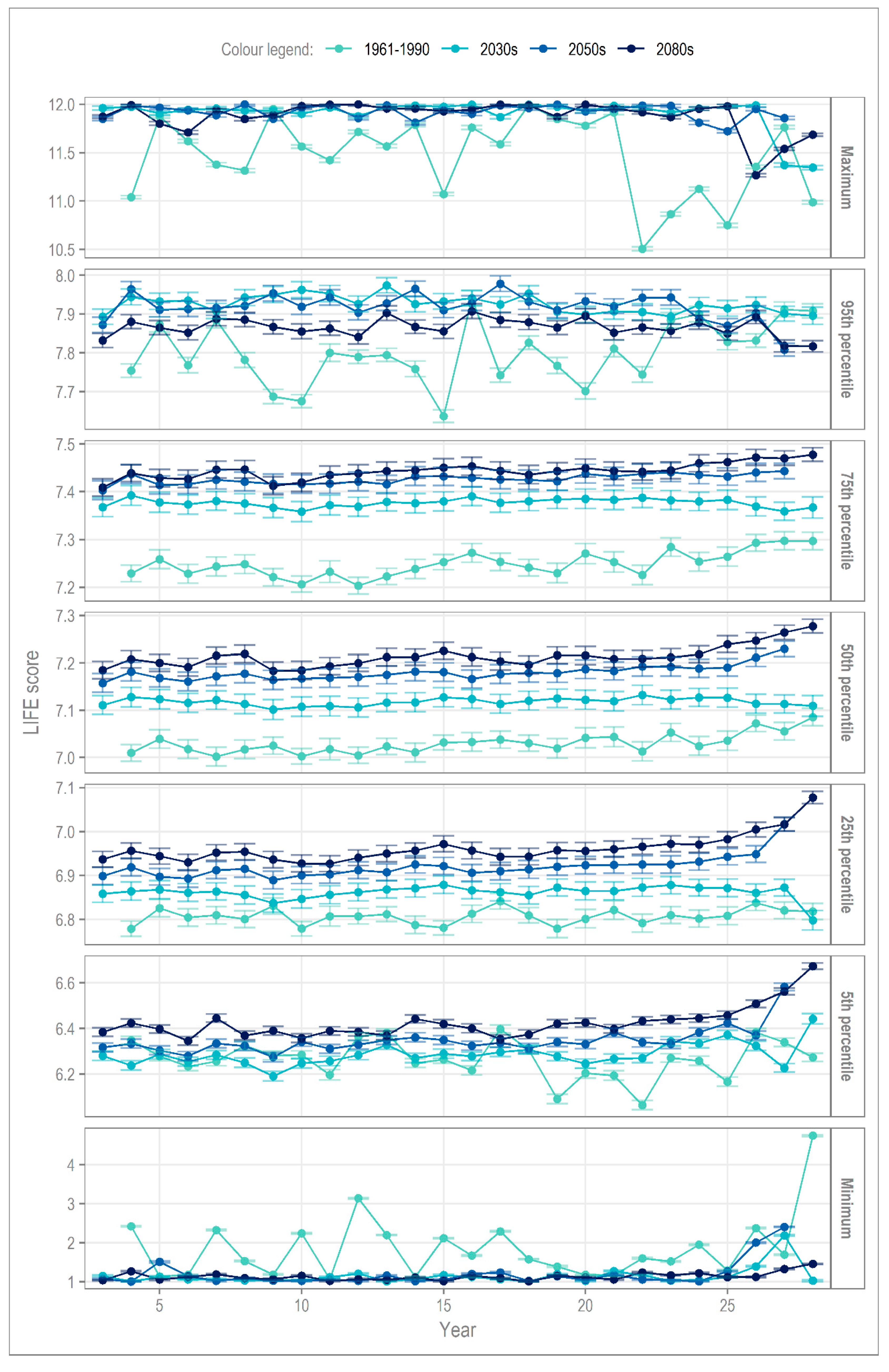
The long-term mean may mask significant changes in the interannual variability of hydroecological response.
Figure 6 describes vertical cross-sections (at specific quantiles) through annual PDFs of LIFE score; the error bars represent the range of values possible for a
virtually certain outcome (99.5% probability, based on the available information). Whilst the y-axis for each quantile does vary, it is clear that, perhaps counter to expectation, that the greatest uncertainty surrounds projections of the median response, and across the 5th to 95th quantiles more generally. Next, we consider the change per cross-section (
Figure 6), starting with the median, interquartile range (IQR) and finally the tails of the distribution.
The small shift in median LIFE scores (by time slice) indicates the increased presence of taxa with higher flow scores (Equation (1)), though the variability of LIFE score remains broadly unchanged. At the 75th percentile, a large change occurs between baseline and the 2030s, while the change from the 2050s to 2080s are almost negligible. At the other end of the IQR (25th percentile), the increase in LIFE scores is approximately linear until the end-of the century. As for the 50th percentile, the variation in interannual LIFE scores, per time slice, remains constant.
We now look to the tails of the distribution, essentially the hydroecological response to lower probability extreme events. At the 95th percentile, the change in variance relative to the baseline stands out (
Figure 6), with
Table 3 revealing that the reduction in variance may reach 92% as early as the 2030s. At the other end of the spectrum, the 5th percentile, the reduction in variance, although reduced, is still high at −65%.
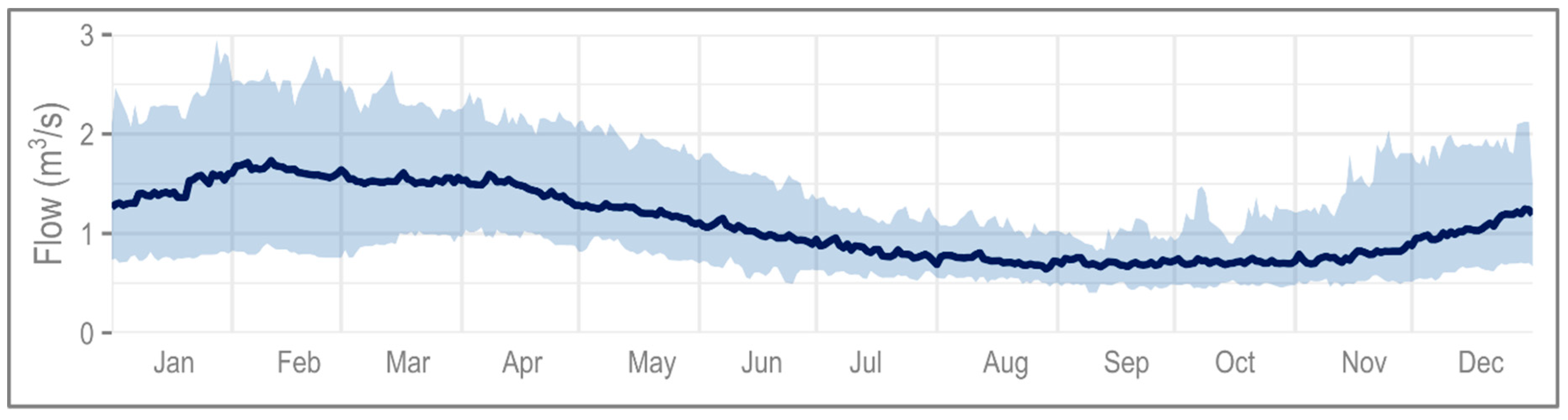
Changes in the maximum and minimum hydroecological responses are marked, affecting not only variance, but also LIFE score. For the maximum, on the baseline, LIFE scores can be seen to vary from 10.5 to the maximum of 12. However, the projections for all three future time slices show a plateau at LIFE scores of 12; a varied response becomes almost impossible. At the minimum, the same phenomenon is observed, with LIFE scores plateauing to a value of 1 with almost no variance. Further, the reduction in range is more notable than for the maximum.
Examination of the year-on-year change in hydroecological response provides further clarification on the subtle changes observed over the long-term (
Section 4.1).
Figure 6 and
Table 3 also highlight the timing of a major change in hydroecological response could occur as early as the 2030s, 2021–2050. This suggests that a major high or low flow event, in the very near future, could result in a hydroecological response very different to the past (baseline period), where there was the probability of a more varied response. By considering the associated uncertainty, we can be
virtually certain of this outcome, based on the available information. Given a potentially highly limited period of preparation, this is of concern for the future health of the River Nar.
4.3. Functional Matrix
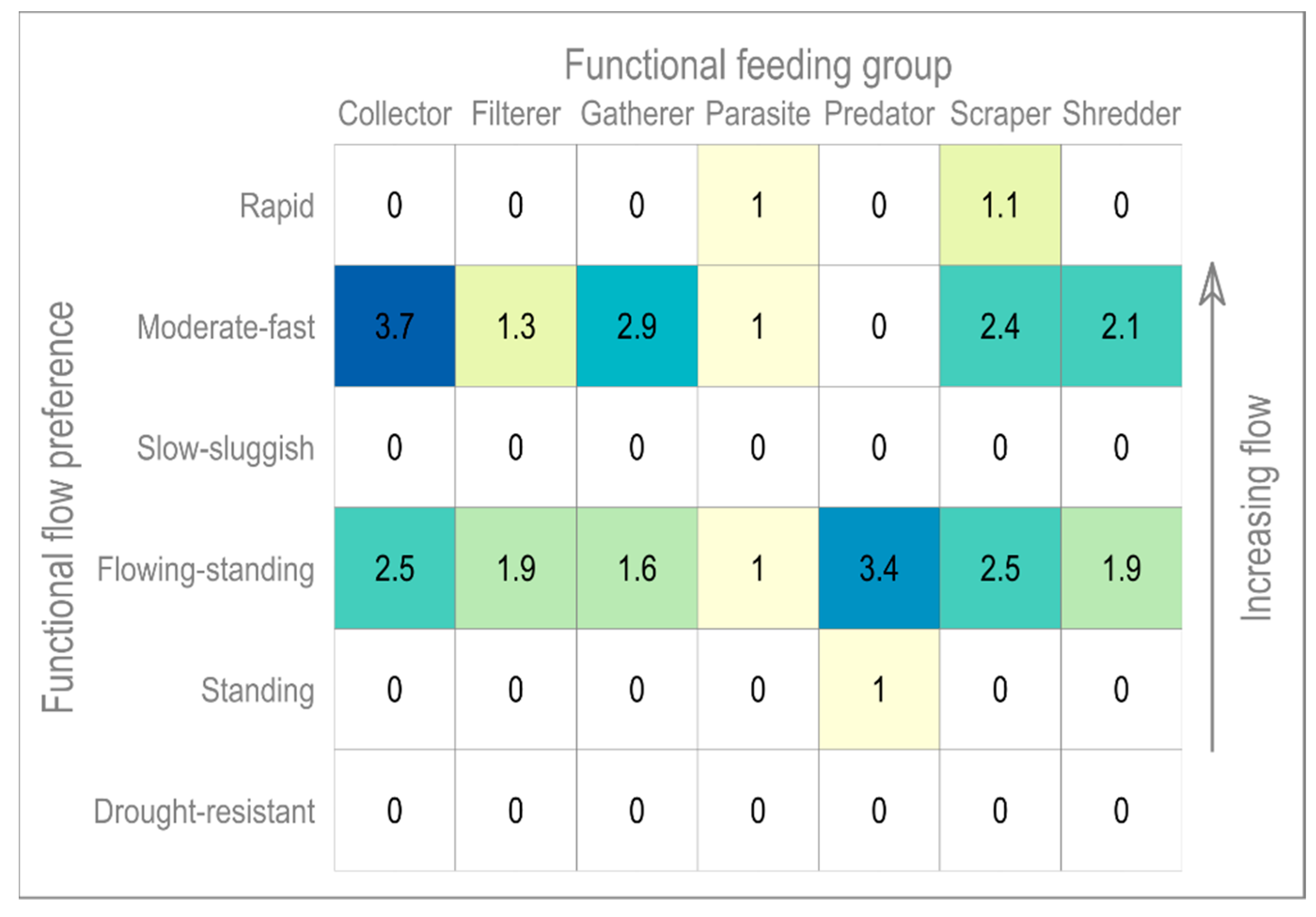
This paper introduces the functional matrix,
Figure 7, relating species-level macroinvertebrate functional flow preferences to functional food groups. See
Appendix A for definitions.
Figure 7 is determined based on observed macroinvertebrate data collected in spring (April-May-June), and thus reflects average conditions between 1993 and 2014. In terms of functional feeding group, only a limited number of species with a range of flow preferences are observed, e.g., scrapers which may tolerate anything from very low to rapid flows. The matrix highlights several functional feeding group traits potentially unrepresented under extreme conditions. The data covers periods of very high and low flows, ensuring that response to extremes are captured. For example, the available time series began in 1993, at the end of the 1989–1992 supra-seasonal drought where groundwater levels fell to their lowest in over 90 years [
71]; inadequate groundwater supplies, coupled with increased water abstraction due to the ongoing drought, saw summer and winter Q95 flow fall below 0.16 and 0.19 m
3/s, respectively [
46].
In the context of the hydroecological projections, we see an increase in the probability of both very high and low LIFE scores. Looking to
Figure A1 in the
Appendix, we can see that LIFE scores below 5/4 are dominated by taxa with preferences for standing waters or drought, and for the highest scores, it is taxa that prefer rapid flows that dominate. Looking then to the functional matrix, it is evident that almost none of the taxa previously observed in the River Nar would be able to perform their functional roles, long-term, under such environmental conditions. In the short-term, the ecosystem has been able to successfully recover, consistent with findings by Wood and Petts [
72]. Wood and Petts found in their 1994 study that the impact of drought on chalk streams was, in part, determined by the health of the river ecosystem prior to the drought event. With the projections indicating a reduction in future biodiversity, the concomitant decrease in macroinvertebrate adaptability may significantly impact the resilience of the riverine system.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









