Effect of Water Column Stability on Surface Chlorophyll and Time Lags under Different Nutrient Backgrounds in a Deep Reservoir





Abstract
:1. Introduction
2. Materials and Methods
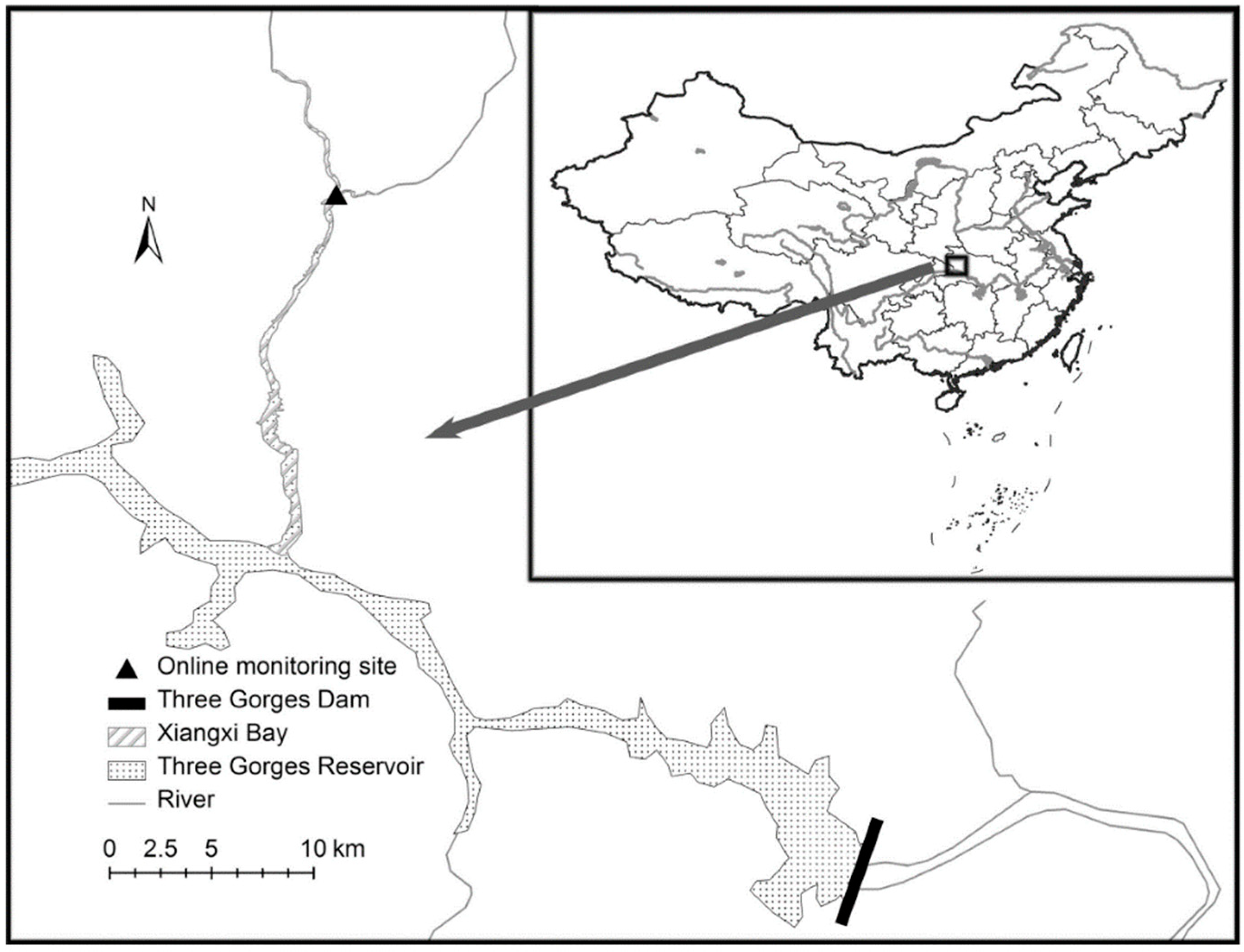
2.1. Study Site
2.2. Field Sampling and Data Analysis
3. Results
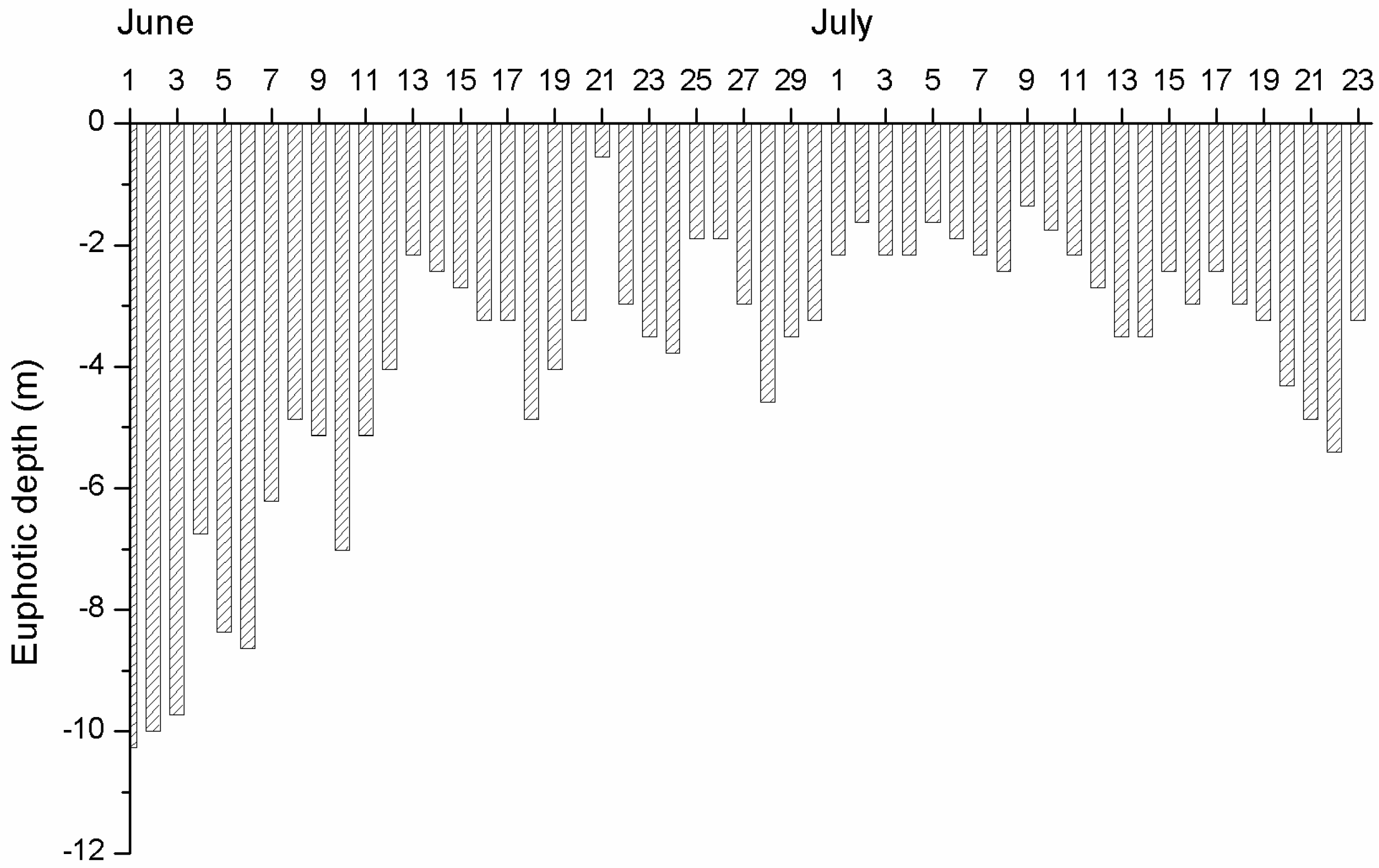
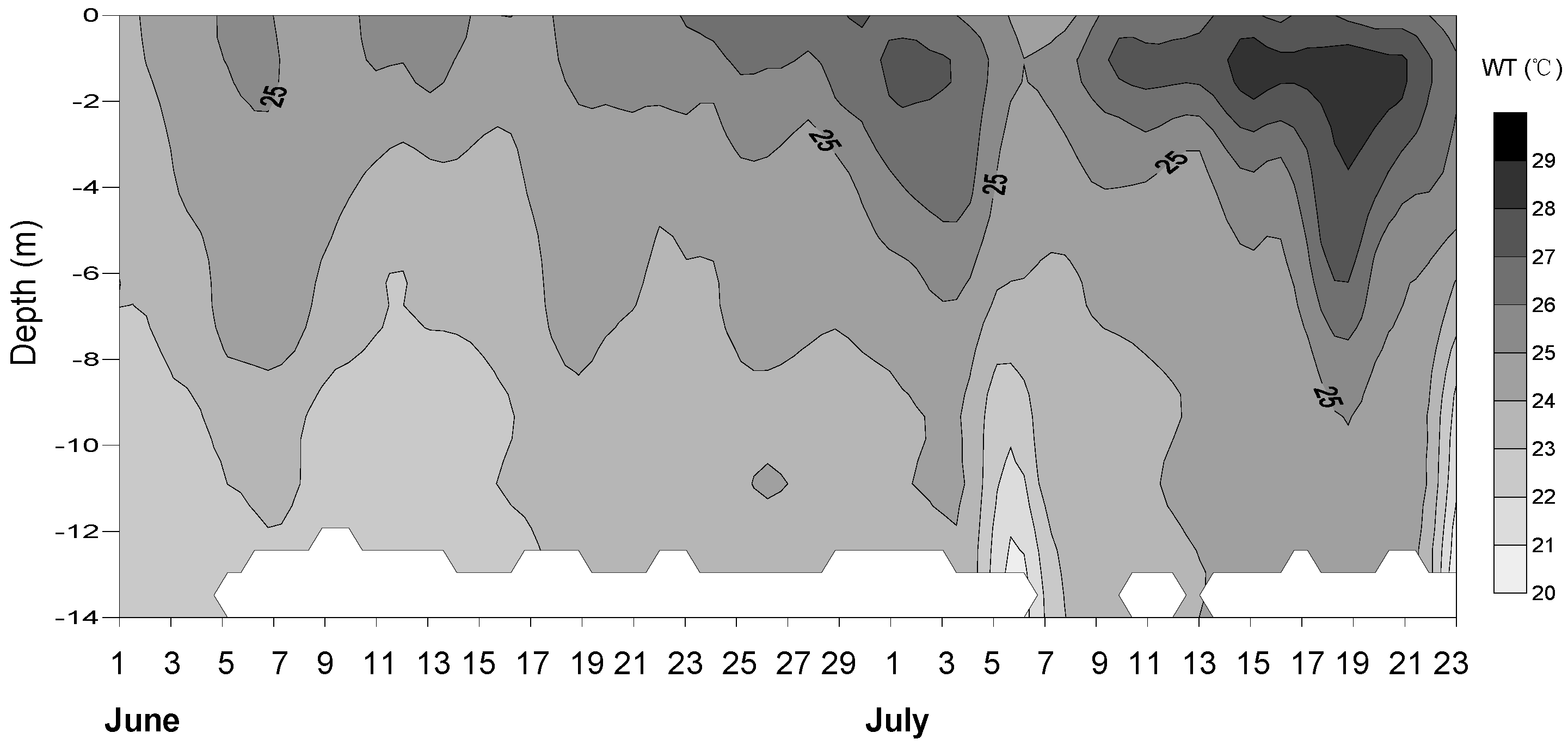
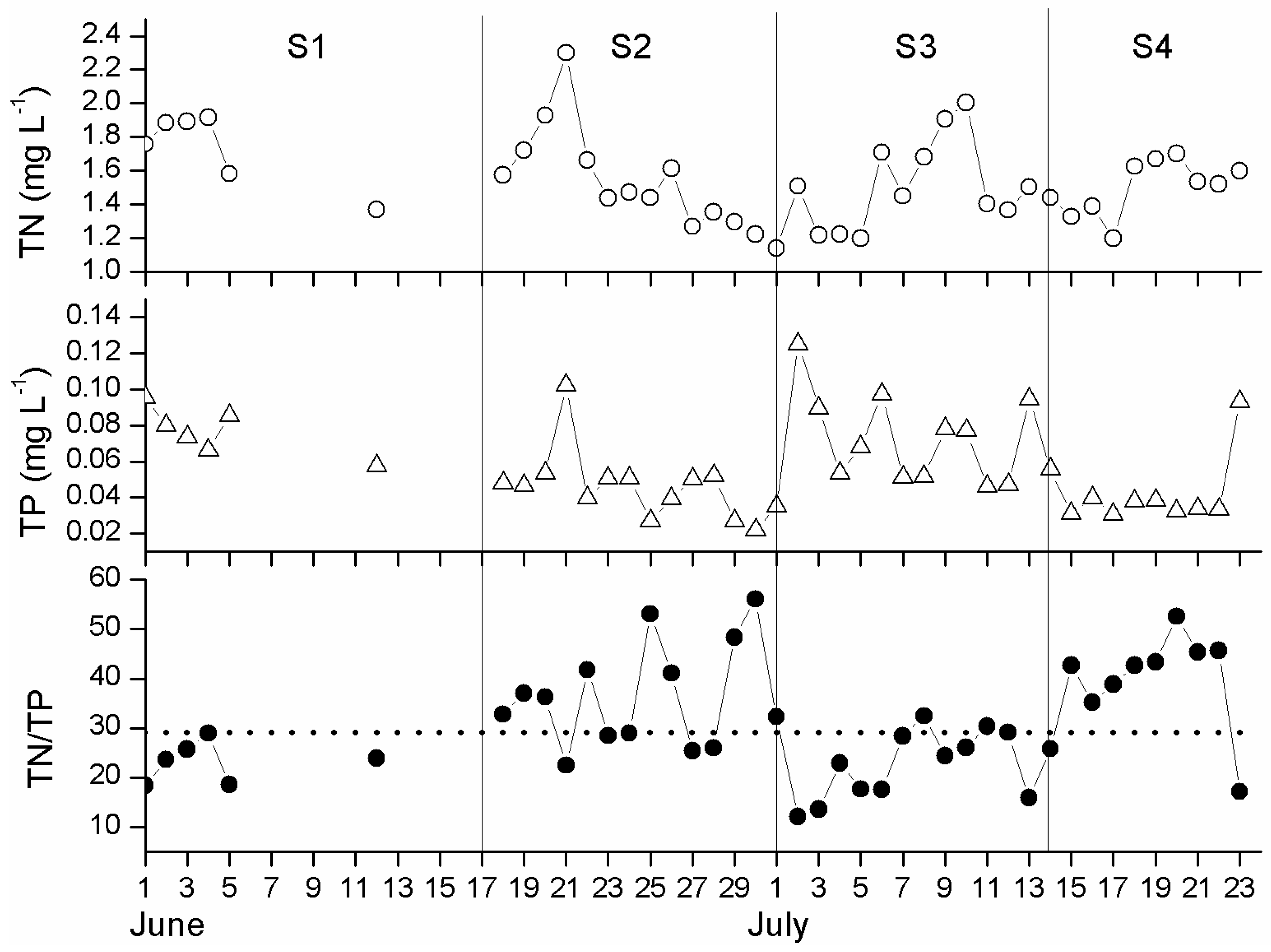
3.1. Physical and Chemical Conditions
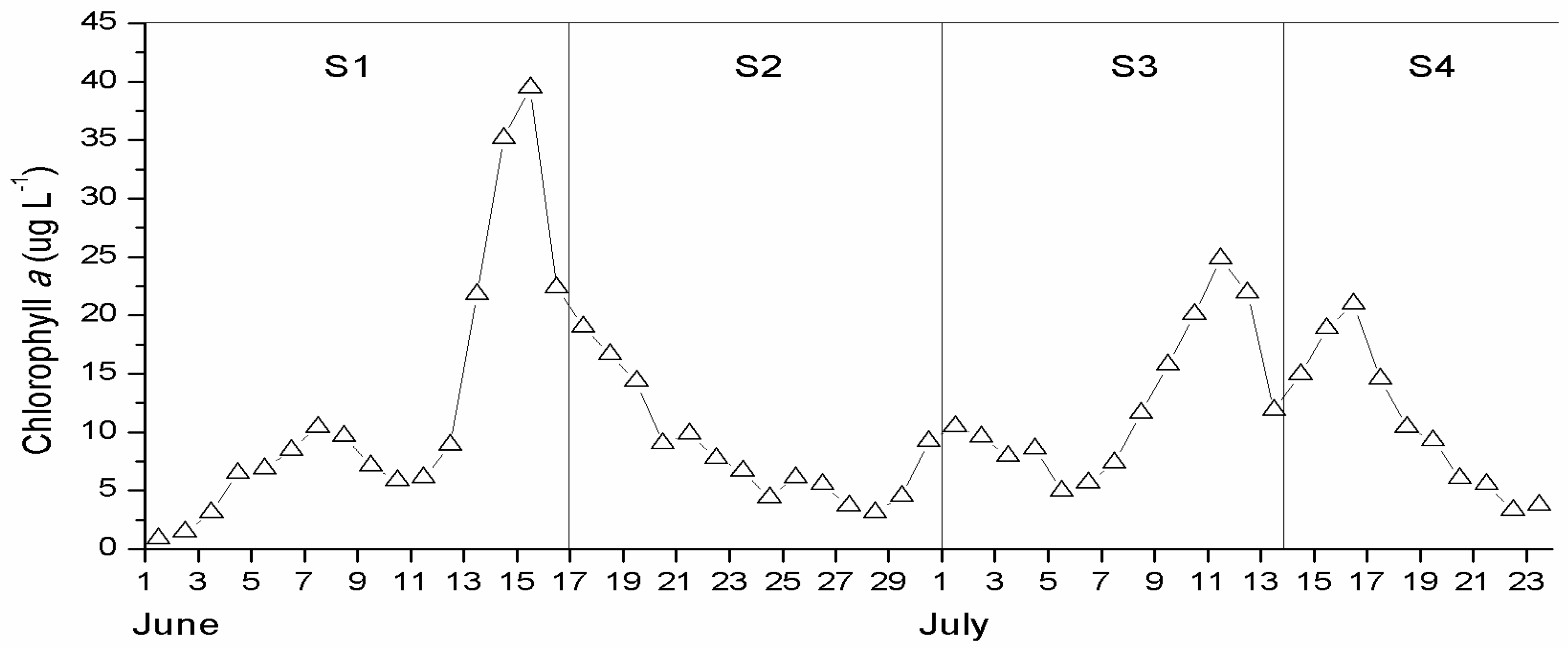
3.2. Surface Chlorophyll a
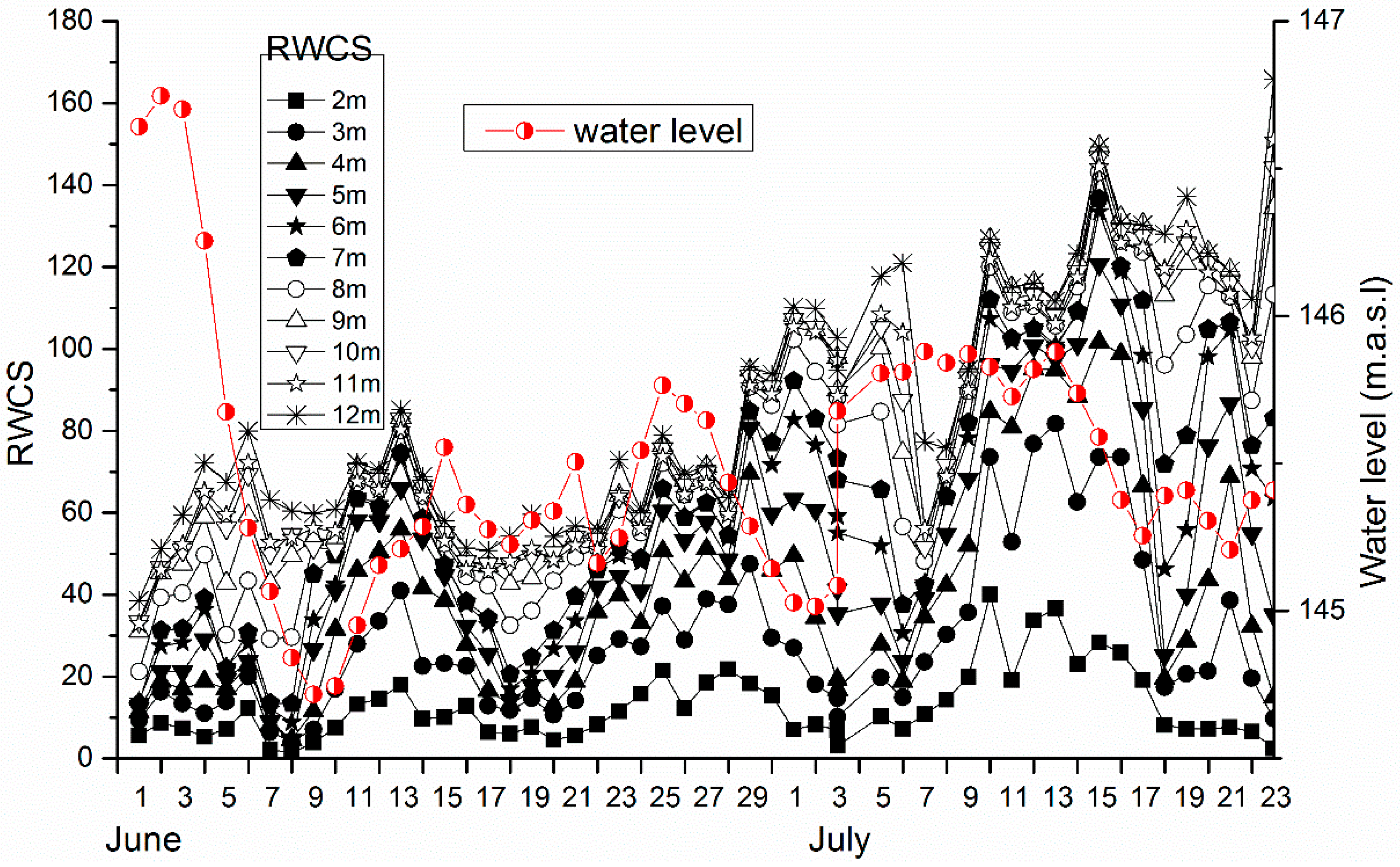
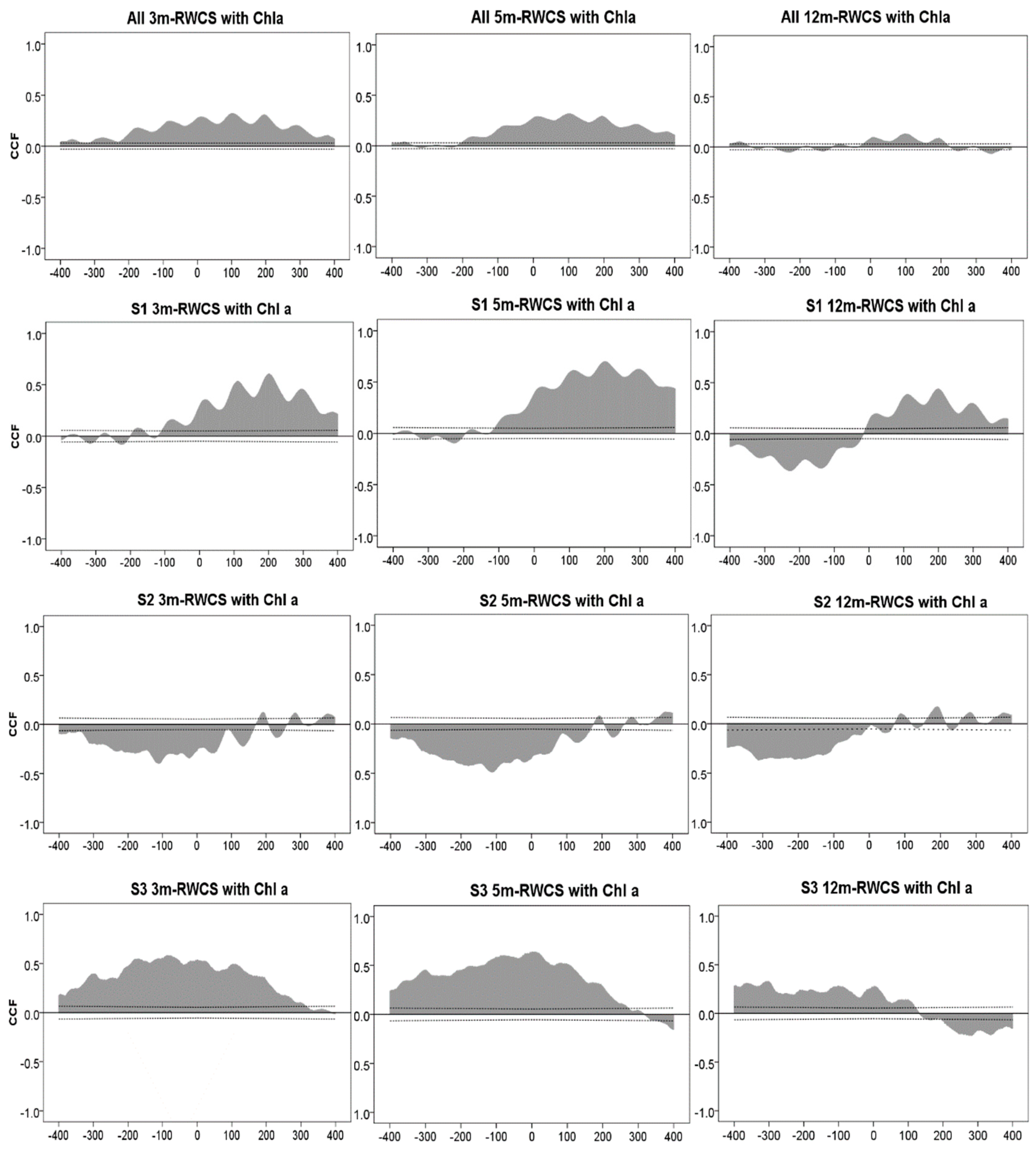
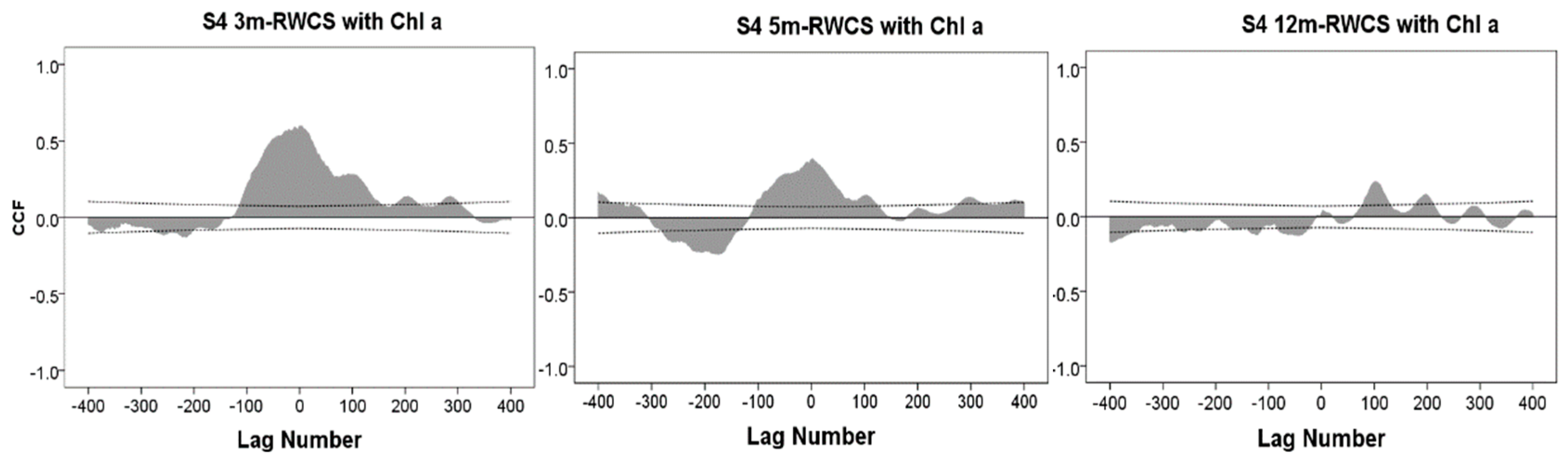
3.3. Cross-Correlation Analysis between Water Column Stability and Chlorophyll a
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pan, Y.; Guo, S.; Li, Y.; Yin, W.; Qi, P.; Shi, J.; Hu, L.; Li, B.; Zhu, J.; Yangdong, P.; et al. Effects of Water Level Increase on Phytoplankton Assemblages in a Drinking Water Reservoi. Water 2018, 10, 256. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Bouman, H.A.; Ulloa, O.; Barlow, R.; Li, W.K.W.; Platt, T.; Zwirglmaier, K.; Scanlan, D.J.; Sathyendranath, S. Water-column stratification governs the community structure of subtropical marine picophytoplankton. Environ. Microbiol. 2011, 3, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Ait Hammou, H.; Latour, D.; Sabart, M.; Samoudi, S.; Mouhri, K.; Robin, J.; Loudiki, M. Temporal evolution and vertical stratification of Microcystis toxic potential during a first bloom event. Aquat. Ecol. 2014, 48, 219–228. [Google Scholar] [CrossRef]
- Visser, P.M.; Ibelings, B.W.; Bormans, M.; Huisman, J. Artificial mixing to control cyanobacterial blooms: A review. Aquat. Ecol. 2016, 50, 423–441. [Google Scholar] [CrossRef]
- Leigh, C.; Burford, M.A.; Roberts, D.T.; Udy, J.W. Predicting the vulnerability of reservoirs to poor water quality and cyanobacterial blooms. Water Res. 2010, 44, 4487–4496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bleiker, W.; Schanz, F. Light climate as the key factor controlling the spring dynamics of phytoplankton in Lake Zürich. Aquat. Sci. 1997, 59, 135–157. [Google Scholar] [CrossRef]
- Carrias, J.F.; Thouvenot, A.; Amblard, C.; Sime-Ngando, T. Dynamics and growth estimates of planktonic protists during early spring in Lake Pavin, France. Aquat. Microb. Ecol. 2001, 24, 163–174. [Google Scholar] [CrossRef]
- Romo, S.; Soria, J.; Fernández, F.; Ou hid, Y.; Barón-Solá, A. Water residence time and the dynamics of toxic cyanobacteria. Freshw. Biol. 2012, 58, 513–522. [Google Scholar] [CrossRef]
- Elliott, J.; Jones, I.; Thackeray, S. Testing the sensitivity of phytoplankton communities to changes in water temperature and nutrient load, in a temperate lake. Hydrobiologia 2006, 559, 401–411. [Google Scholar] [CrossRef]
- Rigosi, A.C.; Carey, C.C.; Ibeling, B.W.; Brookes, J.D. The interaction between climate warming and eutrophication to promote cyanobacteria is dependent on trophic state and varies among taxa. Limnol. Oceanogr. 2014, 59, 99–114. [Google Scholar] [CrossRef] [Green Version]
- Isles, P.D.F.; Giles, C.D.T.; Gearhart, A.; Xu, Y.; Druschel, G.K.; Schroth, A.W. Dynamic internal drivers of a historically severe cyanobacteria bloom in Lake Champlain revealed through comprehensive monitoring. J. Gt. Lakes Res. 2005, 41, 818–829. [Google Scholar] [CrossRef]
- Buranapratheprat, A.; Yanagi, T.; Olaf Niemann, K.; Matsumura, S.; Sojisuporn, P. Surface chlorophyll-a dynamics in the upper Gulf of Thailand revealed by a coupled hydrodynamic-ecosystem model. J. Oceanogr. 2008, 64, 639–656. [Google Scholar] [CrossRef]
- Stainsby, E.A.; Winter, J.G.; Jarjanazi, H.; Paterson, A.M.; Evans, D.O.; Young, J.D. Changes in the thermal stability of Lake Simcoe from 1980 to 2008. J. Gt. Lakes Res. 2011, 37, 55–62. [Google Scholar] [CrossRef]
- Becker, V.; Caputo, L.; Ordóñez, J.; Marcé, R.; Armengol, J.; Crossetti, L.O.; Huszar, V.L.M. Driving factors of the phytoplankton functional groups in a deep Mediterranean reservoir. Water Res. 2010, 44, 3345–3354. [Google Scholar] [CrossRef] [PubMed]
- Jones, I.; Elliott, J. Modelling the effects of changing retention time on abundance and composition of phytoplankton species in a small lake. Freshw. Biol. 2007, 52, 988–997. [Google Scholar] [CrossRef]
- Špoljar, M.; Dražina, T.; Lajtner, J.; Kovačević, G.; Pestić, A.; Matijašec, D.; Tomljanović, T. Impact of water level fluctuation in the shaping of zooplankton assemblage in a shallow lake. Croat. J. Fish. 2018, 76, 27–34. [Google Scholar] [CrossRef]
- Valdespino-Castillo, P.M.; Merino-Ibarra, M.; Jiménez-Contreras, J.; Castillo-Sandoval, F.S.; Ramírez-Zierold, J.A. Community metabolism in a deep (stratified) tropical reservoir during a period of high water-level fluctuations. Environ. Monit. Assess. 2014, 186, 6505–6520. [Google Scholar] [CrossRef] [Green Version]
- Coloso, J.J.; Cole, J.J.; Hanson, P.C.; Pace, M.L. Depth-integrated, continuous estimates of metabolism in a clear-water lake. Can. J. Fish. Aquat. Sci. 2008, 65, 712–722. [Google Scholar] [CrossRef]
- Andersen, M.R.; Jensen, K.S.; Woolway, R.I.; Jones, I.D. Profound daily vertical stratification and mixing in a small shallow, wind-exposed lake with submerged macrophytes. Aquat. Sci. 2016, 79, 395–406. [Google Scholar] [CrossRef]
- Millet, B.; Cecchi, P. Wind-induced hydrodynamic control of the phytoplankton biomass in a lagoon ecosystem. Limnol. Oceanogr. 1992, 37, 140–146. [Google Scholar] [CrossRef]
- Lee, T.A.; Rollwagen-Bollens, G.; Bollens, S.M. The influence of water quality variables on cyanobacterial blooms and phytoplankton community composition in a shallow temperate lake. Environ. Monit. Assess. 2015, 187, 315. [Google Scholar] [CrossRef] [PubMed]
- Staehr, P.A.; Sand-Jensen, K.; Raun, A.L.; Nilsson, B.; Kidmose, J. Drivers of metabolism and net heterotrophy in contrasting lakes. Limnol. Oceanogr. 2010, 55, 817–830. [Google Scholar] [CrossRef]
- Coloso, J.J.; Cole, J.J.; Pace, M.L. Short-term variation in thermal stratification complicates estimation of lake metabolism. Aquat. Sci. 2011, 73, 305–315. [Google Scholar] [CrossRef]
- De Gregorio, S.; Camarda, M.; Longo, M.; Cappuzzo, S.; Sergio Gurrieri, G.G. Long-term continuous monitoring of the dissolved CO2 performed by using a new device in groundwater of the Mt. Etna (southern Italy). Water Res. 2011, 45, 3005–3011. [Google Scholar] [CrossRef] [PubMed]
- Coyle, K.O.; Pinchuk, A.I.; Eisner, L.B.; Napp, J.M. Zooplankton species composition, abundance and biomass on the eastern Bering Sea shelf during summer: The potential role of water-column stability and nutrients in structuring the zooplankton community. Deep Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 1775–1791. [Google Scholar] [CrossRef]
- Ye, L.; Han, X.; Xu, Y.; Cai, Q. Spatial analysis for spring bloom and nutrient limitation in Xiangxi bay of three Gorges Reservoir. Environ. Monit. Assess. 2007, 127, 135–145. [Google Scholar] [CrossRef]
- Zhang, M.; Cai, Q.; Wang, L.; Xu, Y.; Kong, L.; Tan, L. Preliminary study on cyanobacterial bloom in Xiangxi Bay, Three Gorges Reservoir. Wetl. Sci. 2009, 7, 230–236, (In Chinese with English Abstract). [Google Scholar]
- Straskraba, M.; Tundisi, J. Guidelines of Lake Management (Volume 9): Reservoir Water Quality Management; International Lake Environment Committee of UNEP: Shiga, Japan, 1999. [Google Scholar]
- Wang, L.; Cai, Q.; Zhang, M.; Xu, Y.; Kong, L.; Tan, L. Vertical distribution patterns of phytoplankton in summer Microcystis bloom period of Xiangxi Bay, Three Gorges reservoir, China. Fresenius Environ. Bull. 2011, 20, 553–560. [Google Scholar]
- Cole, G.A. Textbook of Limnology; Waveland Press Inc.: Long Grove, IL, USA, 1994. [Google Scholar]
- Padisák, J.; Barbosa, F.; Koschel, R.; Krienitz, L. Deep layer cyanoprokaryota maxima are constitutional features of lakes: Examples from temperate and tropical regions. Arch. für Hydrobiol. Spec. Issues Adv. Limnol. 2003, 58, 175–199. [Google Scholar]
- Jöhnk, K.D.; Huisman, J.E.F.; Sharples, J.; Sommeijer, B.E.N.; Visser, P.M.; Stroom, J.M. Summer heatwaves promote blooms of harmful cyanobacteria. Glob. Chang. Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef]
- Brookes, J.D.; Carey, C.C. Resilience to blooms. Science 2011, 334, 46–47. [Google Scholar] [CrossRef]
- Smith, V.H. Low nitrogen to phosphorus ratios favor dominance by blue-green algae in lake phytoplankton. Science 1983, 221, 669–671. [Google Scholar] [CrossRef]
- Mitchell, B.G.; Brody, E.A.; Holm-Hansen, O.; McClain, C.; Bishop, J. Light limitation of phytoplankton biomass and macronutrient utilization in the Southern Ocean. Limnol. Oceanogr. 1991, 36, 1662–1677. [Google Scholar] [CrossRef]
- Znachor, P.; Zapomělová, E.; Řeháková, K.; Nedoma, J.; Šimek, K. The effect of extreme rainfall on summer succession and vertical distribution of phytoplankton in a lacustrine part of a eutrophic reservoir. Aquat. Sci. 2008, 70, 77–86. [Google Scholar] [CrossRef]
- Scofield, A.E.; Watkins, J.M.; Weidel, B.C.; Luckey, F.J.; Rudstam, L.G. The deep chlorophyll layer in Lake Ontario: Extent, mechanisms of formation, and abiotic predictors. J. Gt. Lakes Res. 2017, 43, 782–794. [Google Scholar] [CrossRef]
- Schindler, D.W.; Hecky, R.E.; Findlay, D.L.; Stainton, M.P.; Parker, B.R.; Paterson, M.J.; Beaty, K.G.; Lyng, M.; Kasian, S.E. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment. Proc. Natl. Acad. Sci. USA 2008, 105, 11254–11258. [Google Scholar] [CrossRef] [Green Version]
- Paerl, H.W.; Xu, H.; McCarthy, M.J.; Zhu, G.; Qin, B.; Li, Y.; Gardner, W.S. Controlling harmful cyanobacterial blooms in a hyper-eutrophic lake (Lake Taihu, China): The need for a dual nutrient (N & P) management strategy. Water Res. 2011, 45, 1973–1983. [Google Scholar]
- Yu, C.; Li, C.; Wang, T.; Zhang, M.; Xu, J. Combined Effects of Experimental Warming and Eutrophication on Phytoplankton Dynamics and Nitrogen Uptake. Water 2018, 10, 1057. [Google Scholar] [CrossRef]
- Dokulil, M.T.; Teubner, K. Cyanobacterial dominance in lakes. Hydrobiologia 2000, 438, 1–12. [Google Scholar] [CrossRef]
- Alpine, A.E.; Cloern, J.E. Trophic interactions and direct physical effects control phytoplankton biomass and production in an estuary. Limnol. Oceanogr. 1992, 37, 946–955. [Google Scholar] [CrossRef]
- Peeters, F.; Kerimoglu, O.; Straile, D. Implications of seasonal mixing for phytoplankton production and bloom development. Theor. Ecol. 2013, 6, 115–129. [Google Scholar] [CrossRef]
- Volpe, G.; Nardelli, B.B.; Cipollini, P.; Santoleri, R.; Robinson, I.S. Seasonal to interannual phytoplankton response to physical processes in the Mediterranean Sea from satellite observations. Remote Sens. Environ. 2011, 117, 223–235. [Google Scholar] [CrossRef]
- Nezlin, N.P.; Li, B.L. Time-series analysis of remote-sensed chlorophyll and environmental factors in the Santa Monica–San Pedro Basin off Southern California. J. Mar. Syst. 2003, 39, 185–202. [Google Scholar] [CrossRef]
- Sarkar, R.R.; Mukhopadhyay, B.; Bhattacharyya, R.; Banerjee, S. Time lags can control algal bloom in two harmful phytoplankton–zooplankton system. Appl. Math. Comput. 2007, 186, 445–459. [Google Scholar] [CrossRef]
- Weisse, T.; Muller, H.; Pinto-Coelho, R.M.; Schweizer, A.; Springmann, D.; Baldringer, G. Response of the microbial loop to the phytoplankton spring bloom in a large prealpine lake. Limnol. Oceanogr. 1990, 35, 781–794. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RWCS | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 m | 3 m | 4 m | 5 m | 6 m | 7 m | 8 m | 9 m | 10 m | 11 m | 12 m | ||
| A | CCF | 0.28 | 0.32 | 0.34 | 0.32 | 0.28 | 0.25 | 0.21 | 0.18 | 0.17 | 0.14 | 0.13 |
| lag | 195 | 101 | 103 | 104 | 97 | 97 | 97 | 98 | 97 | 97 | 97 | |
| S1 | CCF | 0.49 | 0.61 | 0.69 | 0.70 | 0.69 | 0.66 | 0.64 | 0.60 | 0.54 | 0.47 | 0.44 |
| lag | 202 | 202 | 202 | 200 | 200 | 200 | 199 | 199 | 200 | 200 | 200 | |
| S2 | CCF | −0.31 | −0.40 | −0.48 | −0.49 | −0.46 | −0.41 | −0.38 | −0.37 | −0.37 | −0.37 | −0.37 |
| lag | −109 | −112 | −112 | −112 | −112 | −111 | −125 | −207 | −207 | −207 | −312 | |
| S3 | CCF | 0.43 | 0.54 | 0.62 | 0.64 | 0.63 | 0.58 | 0.51 | 0.46 | 0.40 | 0.34 | 0.30 |
| lag | 0 | 0 | 3 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | 2 | |
| S4 | CCF | 0.42 | 0.60 | 0.51 | 0.39 | 0.30 | 0.34 | 0.32 | 0.27 | 0.19 | 0.11 | 0.04 |
| lag | 0 | 2 | 3 | 2 | 2 | 2 | 2 | 2 | 2 | 3 | 3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Niu, Z.; Cai, Q.; Xu, Y.; Qu, X. Effect of Water Column Stability on Surface Chlorophyll and Time Lags under Different Nutrient Backgrounds in a Deep Reservoir. Water 2019, 11, 1504. https://doi.org/10.3390/w11071504
Zhang M, Niu Z, Cai Q, Xu Y, Qu X. Effect of Water Column Stability on Surface Chlorophyll and Time Lags under Different Nutrient Backgrounds in a Deep Reservoir. Water. 2019; 11(7):1504. https://doi.org/10.3390/w11071504
Chicago/Turabian StyleZhang, Min, Zhipan Niu, Qinghua Cai, Yaoyang Xu, and Xiaodong Qu. 2019. "Effect of Water Column Stability on Surface Chlorophyll and Time Lags under Different Nutrient Backgrounds in a Deep Reservoir" Water 11, no. 7: 1504. https://doi.org/10.3390/w11071504
APA StyleZhang, M., Niu, Z., Cai, Q., Xu, Y., & Qu, X. (2019). Effect of Water Column Stability on Surface Chlorophyll and Time Lags under Different Nutrient Backgrounds in a Deep Reservoir. Water, 11(7), 1504. https://doi.org/10.3390/w11071504