Isotopic Signatures as an Indicator of Long-Term Water-Use Efficiency of Haloxylon Plantations on the Dried Aral Sea Bed


Abstract
:1. Introduction
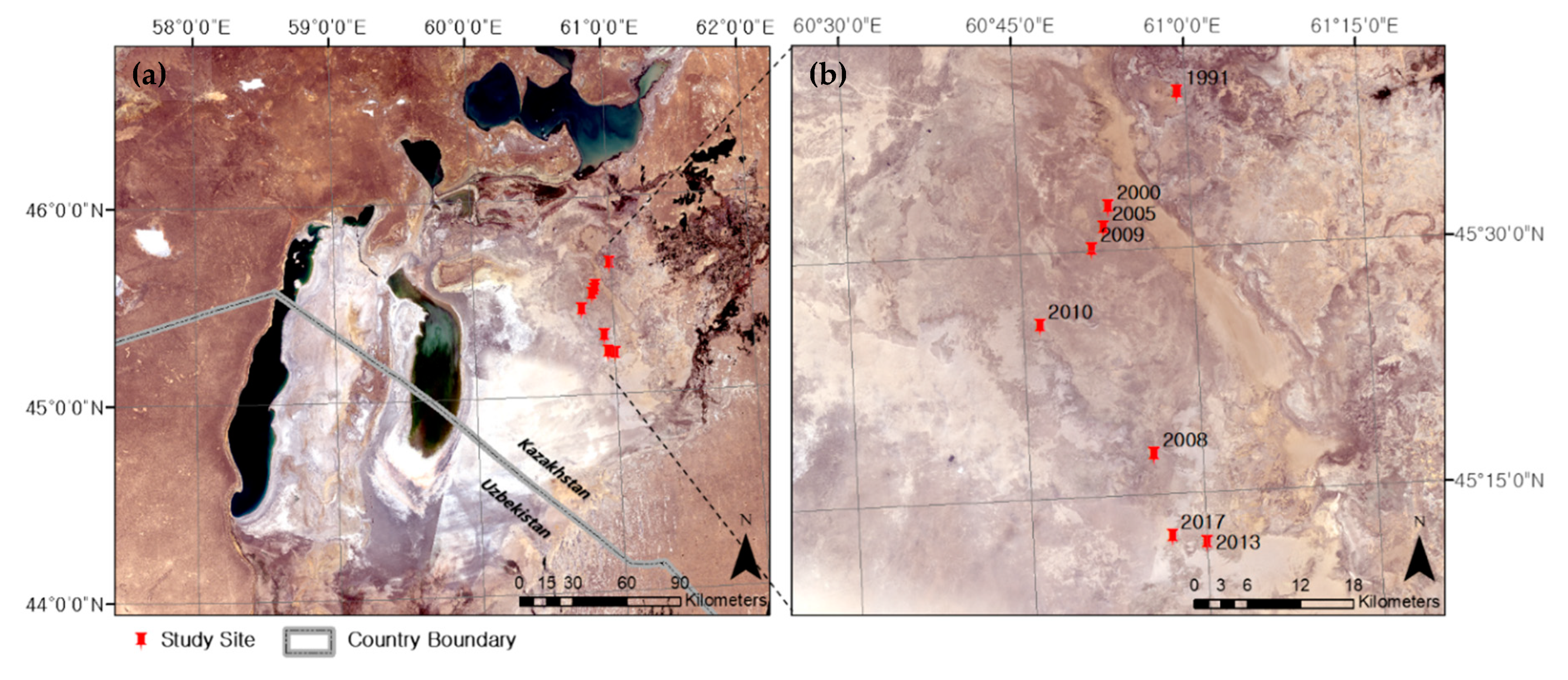
2. Materials and Methods
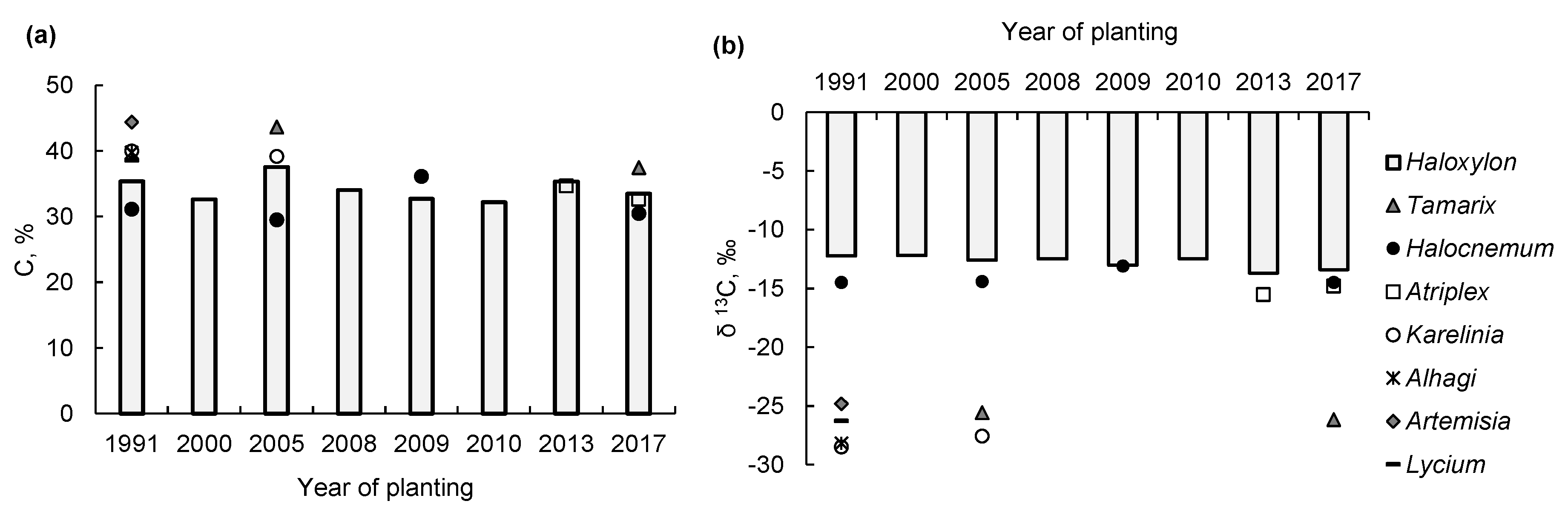
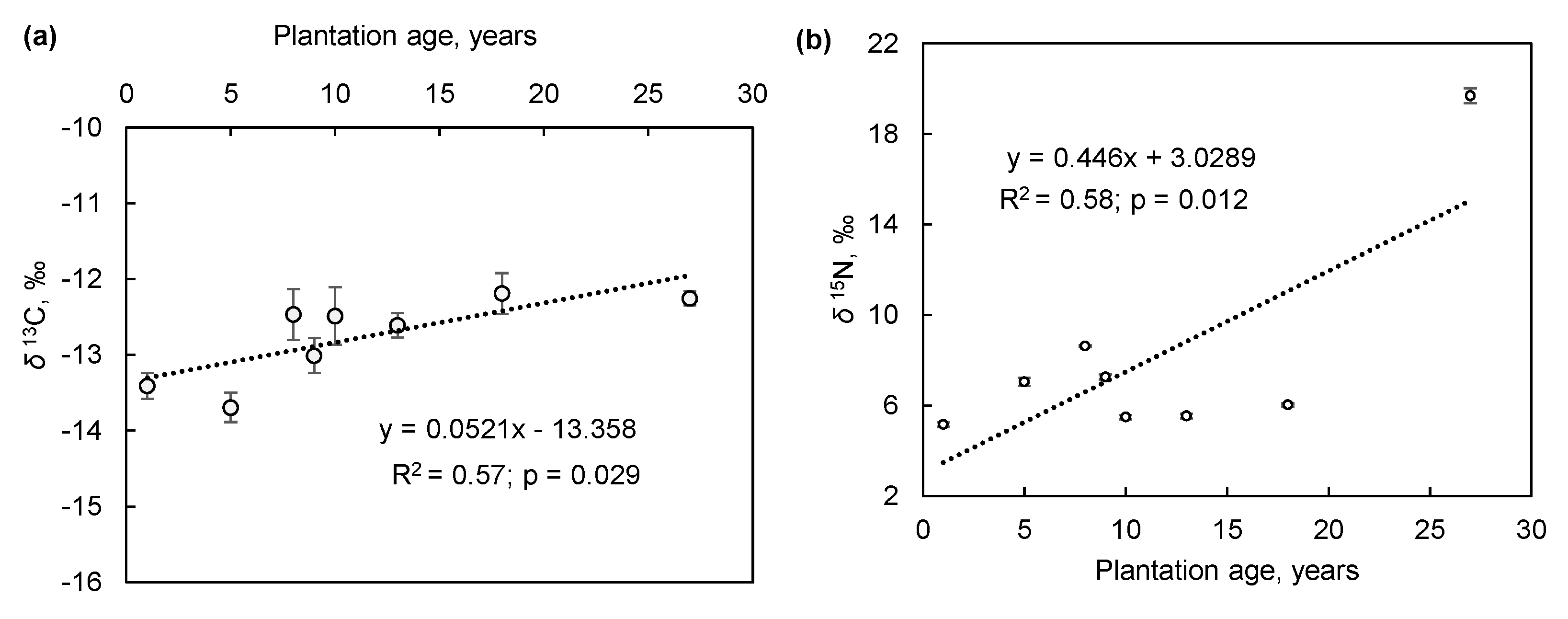
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Breckle, S.W.; Wucherer, W.; Dimeyeva, L.; Ogar, N. Aralkum—A Man-Made Desert. The Desiccated Floor of the Aral Sea (Central Asia); Springer: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Roy, S.B.; Smith, M.; Morris, L.; Orlovsky, N.; Khalilov, A. Impact of the desiccation of the Aral Sea on summertime surface air temperatures. J. Arid Environ. 2014, 110, 79–85. [Google Scholar]
- Ayzel, G.; Izhitskiy, A. Climate change impact assessment on freshwater inflow into the Small Aral Sea. Water 2019, 11, 2377. [Google Scholar] [CrossRef] [Green Version]
- Orlovsky, N.; Birnbaum, E.H. The role of Haloxylon species for combating desertification in Central Asia. Plant Biosyst. 2002, 36, 233–240. [Google Scholar] [CrossRef]
- Novitskiy, Z. Phytomelioration in the Southern Aralkum. In Aralkum—A Man-Made Desert. The Desiccated Floor of the Aral Sea (Central Asia); Breckle, S.-W., Dimeyeva, L., Wucherer, W., Ogar, N.P., Eds.; Springer: Berlin, Germany, 2012; pp. 387–406. [Google Scholar]
- Wucherer, W.; Breckle, S.-W.; Kaverin, V.S.; Dimeyeva, L.A.; Zhamntikov, K. Phytoremediation in the Northern Aralkum. In Aralkum—A Man-Made Desert. The Desiccated Floor of the Aral Sea (Central Asia); Breckle, S.W., Wucherer, W., Dimeyeva, L., Ogar, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 343–386. [Google Scholar]
- Breckle, S.-W. From Aral Sea to Aralkum: An ecological disaster or halophytes’ paradise. In Progress in Botany; Lüttge, U., Beyschlag, W., Francis, D., Cushman, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 74, pp. 351–398. [Google Scholar]
- Löw, F.; Navratil, P.; Kotte, K.; Schöler, H.F.; Bubenzer, O. Remote-sensing-based analysis of landscape change in the desiccated seabed of the Aral Sea—A potential tool for assessing the hazard degree of dust and salt storms. Environ. Monit. Assess. 2013, 185, 8303–8319. [Google Scholar] [CrossRef] [PubMed]
- Pate, J.S. Carbon isotope discrimination and plant water-use efficiency. In Stable Isotope Techniques in the Study of Biological Processes and Functioning of Ecosystems; Unkovich, M., Pate, J.S., McNeill, A., Gibbs, D.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 19–36. [Google Scholar]
- Dawson, T.E.; Mambelli, S.; Plamboeck, A.H.; Templer, P.H.; Tu, K.P. Stable isotopes in plant ecology. Annu. Rev. Ecol. Evol. Syst. 2002, 33, 507–559. [Google Scholar] [CrossRef]
- Swap, R.J.; Aranibar, J.N.; Dowty, P.R.; Gilhooly, W.P.; Macko, S.A. Natural abundance of 13C and 15N in C3 and C4 vegetation of Southern Africa: Patterns and implications. Glob. Chang. Biol. 2004, 10, 350–358. [Google Scholar] [CrossRef]
- Hartman, G.; Fanin, A. Isotopic values of plants in relation to water availability in the Eastern Mediterranean Region. Oecologia 2010, 162, 837–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munns, R. Plant adaptations to salt and water stress: Differences and commonalities. In Advances in Botanical Research; Kader, J.-C., Delseny, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; Volume 57, pp. 2–32. [Google Scholar]
- Ma, J.-Y.; Sun, W.; Liu, X.-N.; Chen, F.-H. Variation in the stable carbon and nitrogen isotope composition of plants and soil along a precipitation gradient in northern China. PLoS ONE 2012, 7, e51894. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, Y.; Xu, G.; Zou, T. Ecophysiological response and morphological adjustment of two Central Asian desert shrubs towards variation in summer precipitation. Plant Cell Environ. 2007, 30, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Cheng, J.; Liu, Y.; Liu, L.; Li, R.; Sheng, J. Relationship of stable carbon isotope composition with environmental factors in the desert plant, Haloxylon ammodendron. Acta Ecol. Sin. 2017, 37, 2743–2752. [Google Scholar]
- Su, P.-X.; Liu, X.-M.; Zhang, L.-X.; Zhao, A.-F.; Li, W.-R.; Chen, H.-S. Comparison of d13C values and gas exchange of assimilating shoots of desert plants Haloxylon ammodendron and Calligonum mongolicum with other plants. Isr. J. Plant Sci. 2013, 52, 87–97. [Google Scholar] [CrossRef]
- Matsuo, N.; Ojika, K.; Shuyskaya, E.; Radjabov, T.; Toderich, K.; Yamanaka, N. Responses of the carbon and oxygen isotope compositions of desert plants to spatial variation in soil salinity in Central Asia. Ecol. Res. 2013, 28, 717–723. [Google Scholar] [CrossRef]
- Gintzburger, G.; Toderich, K.N.; Mardonov, B.K.; Mahmudov, M.M. Rangelands of the Arid and Semi-Arid Zones in Uzbekistan; CIRAD ICARDA: Montpellier, France, 2003; p. 431. [Google Scholar]
- Poss, J.A.; Gratten, S.R.; Suarez, D.L.; Grieve, C.M. Stable carbon isotope discrimination: An indicator of cumulative salinity and boron stress in Eucalyptus camaldulensis. Tree Physiol. 2000, 20, 1121–1127. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Number | Family | Species | Common or Local Name | Year of Afforestation |
|---|---|---|---|---|
| 1. | Amaranthaceae/Chenopodiaceae | Haloxylon aphyllum (Minkw.) Iljin. (Synonym: Haloxylon ammodendron (C.A.Mey.) | Black saksaul | 1991, 2000, 2005, 2008, 2009, 2010, 2013, 2017 |
| 2. | Amaranthaceae | Halocnemum strobilaceum (Pall.) Bieb | Sarsazan | 1991, 2005, 2009, 2017 |
| 3. | Amaranthaceae | Atriplex fominii Iljin. | Lebeda, Olabuta | 2013, 2017 |
| 4. | Asteraceae | Karelinia caspia (Pall.) Less. | Ak-bash | 1991, 2005 |
| 5. | Asteraceae | Artemisia diffusa H. Krasch. | Common wormwood | 1991 |
| 6. | Tamaricaceae | Tamarix hispida Willd. | Russian tamarisk | 2005, 2017 |
| 7. | Fabaceae | Alhagi maurorum Medik. | Camelthorn | 1991 |
| 8. | Solanaceae | Lycium ruthenicum Murray | Karamik | 1991 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khamzina, A.; An, J.; Chang, H.; Son, Y. Isotopic Signatures as an Indicator of Long-Term Water-Use Efficiency of Haloxylon Plantations on the Dried Aral Sea Bed. Water 2020, 12, 99. https://doi.org/10.3390/w12010099
Khamzina A, An J, Chang H, Son Y. Isotopic Signatures as an Indicator of Long-Term Water-Use Efficiency of Haloxylon Plantations on the Dried Aral Sea Bed. Water. 2020; 12(1):99. https://doi.org/10.3390/w12010099
Chicago/Turabian StyleKhamzina, Asia, Jiae An, Hanna Chang, and Yowhan Son. 2020. "Isotopic Signatures as an Indicator of Long-Term Water-Use Efficiency of Haloxylon Plantations on the Dried Aral Sea Bed" Water 12, no. 1: 99. https://doi.org/10.3390/w12010099
APA StyleKhamzina, A., An, J., Chang, H., & Son, Y. (2020). Isotopic Signatures as an Indicator of Long-Term Water-Use Efficiency of Haloxylon Plantations on the Dried Aral Sea Bed. Water, 12(1), 99. https://doi.org/10.3390/w12010099