Habitat Suitability Curves for Freshwater Macroinvertebrates of Tropical Andean Rivers


Abstract
:1. Introduction
2. Materials and Methods
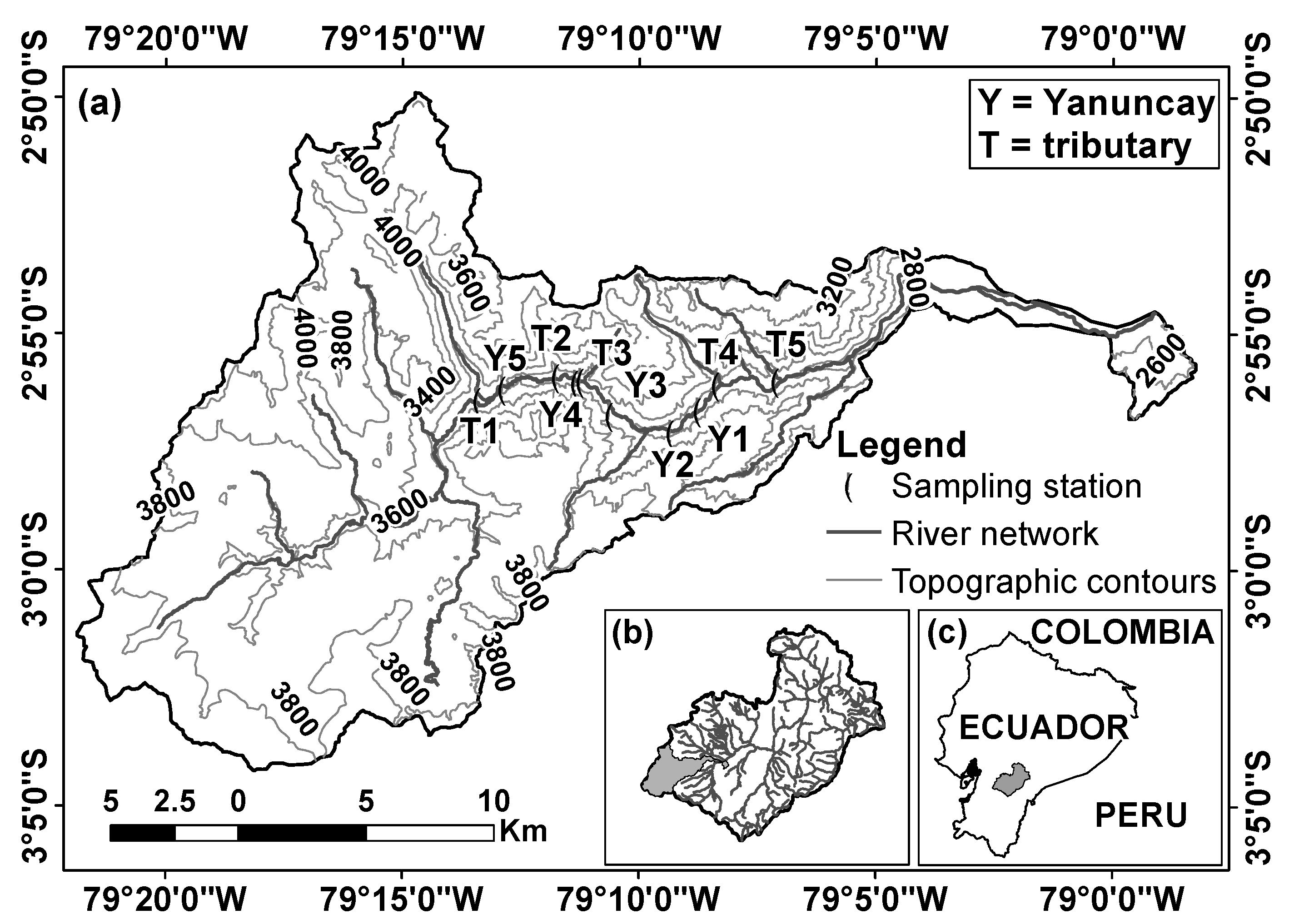
2.1. Study Area
2.2. Abiotic Monitoring
2.3. Biotic Monitoring
2.4. Data Processing
2.5. Statistical Analysis
Development of Habitat Suitability Curves
3. Results
3.1. Environmental Characteristics and Macroinvertebrate Taxa
3.2. Variables Influencing the Macroinvertebrate Communities at Microhabitat Scale
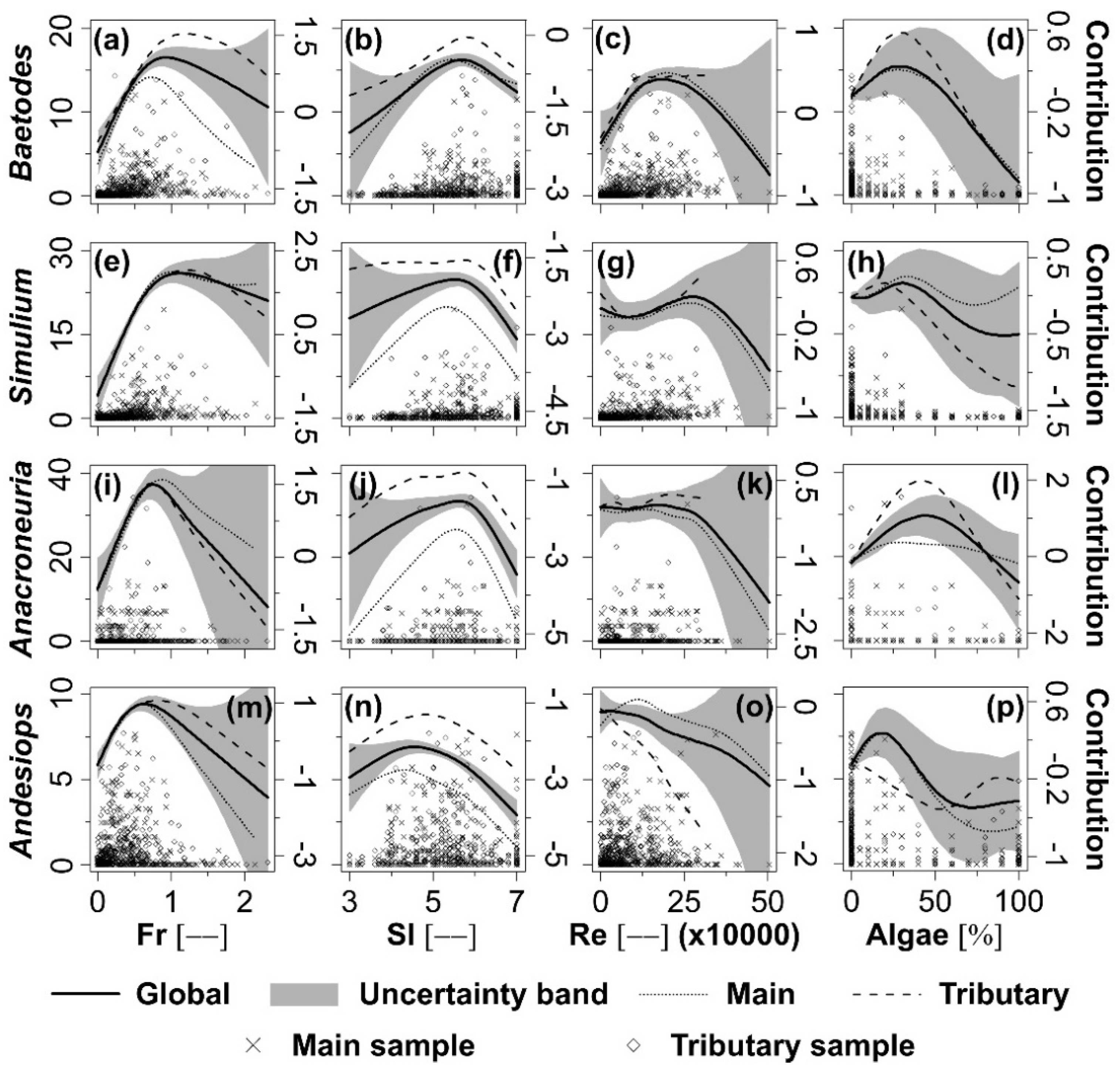
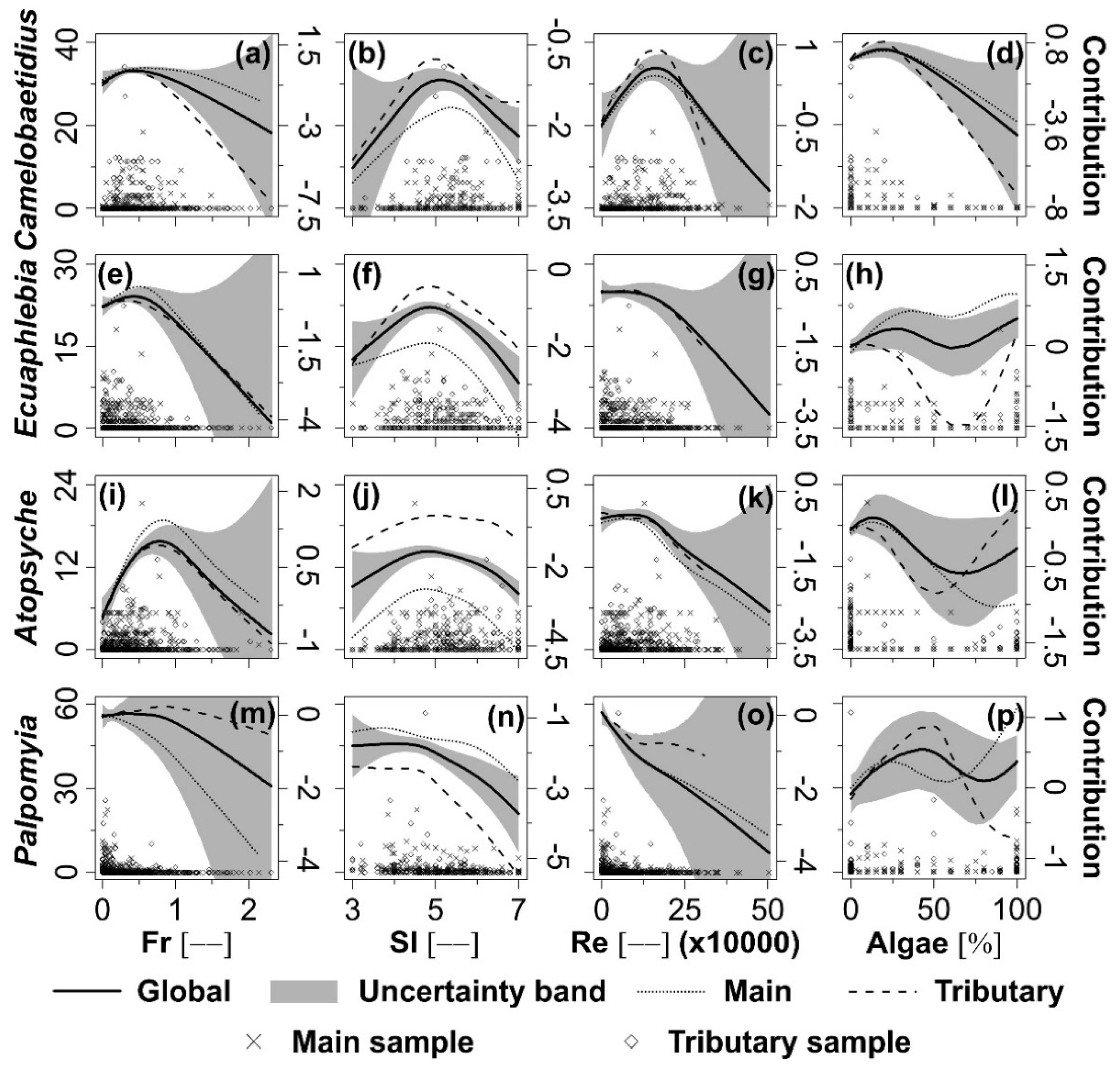
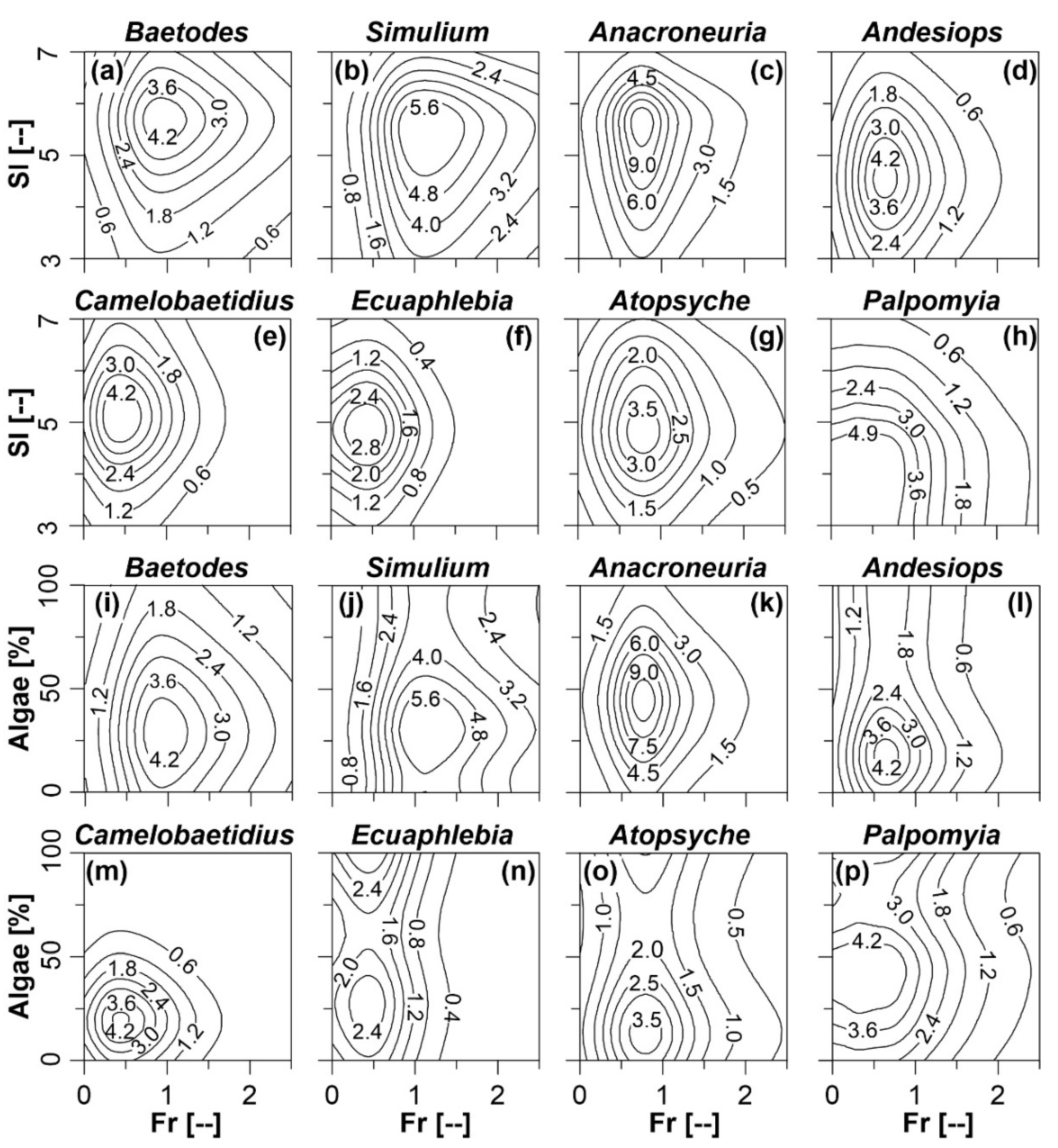
3.3. Individual Taxa Response to Selected Hydraulic and Habitat Variables
4. Discussion
4.1. Variables Influencing Macroinvertebrate Distribution at the Microhabitat Scale
4.2. Individual Taxa Response to Selected Hydraulic and Habitat Variables
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Uribe, N.; Srinivasan, R.; Corzo, G.; Arango, D.; Solomatine, D. Spatio-temporal critical source area patterns of runoff pollution from agricultural practices in the Colombian Andes. Ecol. Eng. 2020, 149, 105810. [Google Scholar] [CrossRef]
- Miserendino, M.L.; Pizzolon, L.A. Interactive effects of basin features and land-use change on macroinvertebrate communities of headwater streams in the Patagonian Andes. River Res. Appl. 2004, 20, 967–983. [Google Scholar] [CrossRef]
- Durance, I.; Ormerod, S.J. Climate change effects on upland stream macroinvertebrates over a 25-year period. Glob. Chang. Biol. 2007, 13, 942–957. [Google Scholar] [CrossRef]
- Bo, T.; Piano, E.; Doretto, A.; Bona, F.; Fenoglio, S. Microhabitat preference of sympatric Hydraena Kugelann, 1794 species (Coleoptera: Hydraenidae) in a low-order forest stream. Aquat. Insects 2017, 37, 287–292. [Google Scholar] [CrossRef]
- Li, F.; Cai, Q.; Fu, X.; Liu, J. Construction of habitat suitability models (HSMs) for benthic macroinvertebrate and their applications to instream environmental flows: A case study in Xiangxi River of Three Gorges Reservior region, China. Prog. Nat. Sci. 2009, 19, 359–367. [Google Scholar] [CrossRef]
- Bedoya, D.; Manolakos, E.S.; Novotny, V. Characterization of biological responses under different environmental conditions: A hierarchical modeling approach. Ecol. Model. 2011, 222, 532–545. [Google Scholar] [CrossRef]
- Schröder, M.; Kiesel, J.; Schattmann, A.; Jähnig, S.C.; Lorenz, A.W.; Kramm, S.; Keizer-Vlek, H.; Rolauffs, P.; Graf, W.; Leitner, P.; et al. Substratum associations of benthic invertebrates in lowland and mountain streams. Ecol. Indic. 2013, 30, 178–189. [Google Scholar] [CrossRef]
- Serpa, K.V.; Kiffer, W.P.; Borelli, M.F.; Ferraz, M.A.; Moretti, M.S. Niche breadth of invertebrate shredders in tropical forest streams: Which taxa have restricted habitat preferences? Hydrobiologia 2019, 847, 1739–1752. [Google Scholar] [CrossRef]
- Parsons, M.; Norris, R. The effect of habitat-specific sampling on biological assessment of water quality using a predictive model. Freshw. Biol. 1996, 36, 419–434. [Google Scholar] [CrossRef]
- Vimos-Lojano, D.; Martínez-Capel, F.; Hampel, H. Riparian and microhabitat factors determine the structure of the EPT community in Andean headwater rivers of Ecuador. Ecohydrology 2017, 10, e1894. [Google Scholar] [CrossRef] [Green Version]
- Collier, K.J. Flow preferences of larval Chironomidae (Diptera) in Tongariro River, New Zealand. N. Z. J. Mar. Freshw. Res. 1993, 27, 219–226. [Google Scholar] [CrossRef]
- Degani, G.; Herbst, G.N.; Ortal, R.; Bromley, H.J.; Levanon, D.; Netzer, Y.; Harari, N.; Glazman, H. Relationships between current velocity, depth and the invertebrate community in a stable river system. Hydrobiologia 1993, 263, 163–172. [Google Scholar] [CrossRef]
- Guevara-Mora, M.; Pedreros, P.; Urrutia, R.; Figueroa, R. Efectos de la extracción agrícola del agua en el hábitat fluvial de macroinvertebrados bentónicos en Chile. Hidrobiológica 2016, 26, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Mérigoux, S.; Dolédec, S. Hydraulic requirements of stream communities: A case study on invertebrates. Freshw. Biol. 2004, 49, 600–613. [Google Scholar] [CrossRef]
- Statzner, B.; Gore, J.A.; Resh, V.H. Hydraulic stream ecology: Observed patterns and potential applications. J. N. Am. Benthol. Soc. 1988, 7, 307–360. [Google Scholar] [CrossRef]
- Shearer, K.; Hayes, J.; Jowett, I.; Olsen, D. Habitat suitability curves for benthic macroinvertebrates from a small New Zealand river. N. Z. J. Mar. Freshw. Res. 2015, 49, 178–191. [Google Scholar] [CrossRef]
- Kim, S.K.; Choi, S.-U. Ecological evaluation of weir removal based on physical habitat simulations for macroinvertebrate community. Ecol. Eng. 2019, 138, 362–373. [Google Scholar] [CrossRef]
- Mesa, L.M. Hydraulic parameters and longitudinal distribution of macroinvertebrates in a subtropical Andean basin. Interciencia 2010, 35, 759–764. [Google Scholar]
- Jerves-Cobo, R.; Everaert, G.; Iñiguez-Vela, X.; Córdova-Vela, G.; Díaz-Granda, C.; Cisneros, F.; Nopens, I.; Goethals, P.L. A methodology to model environmental preferences of EPT taxa in the Machangara River Basin (Ecuador). Water 2017, 9, 195. [Google Scholar] [CrossRef] [Green Version]
- Masikini, R.; Kaaya, L.T.; Chicharo, L. Evaluation of ecohydrological variables in relation to spatial and temporal variability of macroinvertebrate assemblages along the Zigi River–Tanzania. Ecohydrol. Hydrobiol. 2018, 18, 130–141. [Google Scholar] [CrossRef]
- Holzapfel, P.; Leitner, P.; Habersack, H.; Graf, W.; Hauer, C. Evaluation of hydropeaking impacts on the food web in alpine streams based on modelling of fish- and macroinvertebrate habitats. Sci. Total Environ. 2017, 575, 1489–1502. [Google Scholar] [CrossRef] [PubMed]
- Monk, W.A.; Compson, Z.G.; Armanini, D.G.; Orlofske, J.M.; Curry, C.J.; Peters, D.L.; Crocker, J.B.; Baird, D.J. Flow velocity–ecology thresholds in Canadian rivers: A comparison of trait and taxonomy-based approaches. Freshw. Biol. 2017, 63, 891–905. [Google Scholar] [CrossRef]
- Asamblea-Nacional. Ley Orgánica de Recursos Hídricos, Usos y Aprovechamiento del Agua; Asamblea-Nacional: Quito, Ecuador, 2014. [Google Scholar]
- MINAMBIENTE IDEAM. Guía Metodológica para la Estimación del Caudal Ambiental; Ministerio de Ambiente y Desarrollo Sostenible (MINAMBIENTE), Meteorología y Estudios Ambientales Instituto de Hidrología, Eds.; MINAMBIENTE IDEAM: Bogotá, Colombia, 2017. [Google Scholar]
- Anderson, E.P.; Jenkins, C.N.; Heilpern, S.; Maldonado-Ocampo, J.A.; Carvajal-Vallejos, F.M.; Encalada, A.C.; Rivadeneira, J.F.; Hidalgo, M.; Cañas, C.M.; Ortega, H.; et al. Fragmentation of Andes-to-Amazon connectivity by hydropower dams. Sci. Adv. 2018, 4, eaao1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borja, P.; Molina, A.; Govers, G.; Vanacker, V. Check dams and afforestation reducing sediment mobilization in active gully systems in the Andean mountains. Catena 2018, 165, 42–53. [Google Scholar] [CrossRef]
- Gore, J.A.; Judy, R.D. Predictive models of benthic macroinvertebrate density for use in instream flow studies and regulated flow management. Can. J. Fish. Aquat. Sci. 1981, 38, 1363–1370. [Google Scholar] [CrossRef]
- Theodoropoulos, C.; Vourka, A.; Skoulikidis, N.; Rutschmann, P.; Stamou, A. Evaluating the performance of habitat models for predicting the environmental flow requirements of benthic macroinvertebrates. J. Ecohydraulics 2018, 3, 30–44. [Google Scholar] [CrossRef]
- Kelly, D.J.; Hayes, J.W.; Allen, C.; West, D.; Hudson, H. Evaluating habitat suitability curves for predicting variation in macroinvertebrate biomass with weighted usable area in braided rivers in New Zealand. N. Z. J. Mar. Freshw. Res. 2015, 49, 398–418. [Google Scholar] [CrossRef]
- Muñoz-Mas, R.; Papadaki, C.; Martínez-Capel, F.; Zogaris, S.; Ntoanidis, L.; Dimitriou, E. Generalized additive and fuzzy models in environmental flow assessment: A comparison employing the West Balkan trout (Salmo farioides; Karaman, 1938). Ecol. Eng. 2016, 91, 365–377. [Google Scholar] [CrossRef]
- Ahmadi-Nedushan, B.; St-Hilaire, A.; Bérubé, M.; Robichaud, É.; Thiémonge, N.; Bobée, B. A review of statistical methods for the evaluation of aquatic habitat suitability for instream flow assessment. River Res. Appl. 2006, 22, 503–523. [Google Scholar] [CrossRef]
- Payne, T.R.; Jowett, I. SEFA—Computer Software System for Environmental Flow Analysis Based on the Instream Flow Incremental Methodology. In Proceedings of the 2013 Georgia Water Resources Conference, Athens, Georgia, 10–11 April 2013. [Google Scholar]
- Lamouroux, N.; Jowett, I.G. Generalized instream habitat models. Can. J. Fish. Aquat. Sci. 2005, 62, 7–14. [Google Scholar] [CrossRef]
- Jowett, I.G. Flow management. In New Zealand Stream Invertebrates: Ecology and Implications for Management; Collier, K., Winterbourn, M., Eds.; New Zealand Limnological Society: Christchurch, New Zealand, 2000; pp. 289–312. ISBN 0473066793. [Google Scholar]
- Jowett, I.G. Hydraulic constraints on habitat suitability for benthic invertebrates in gravel-bed rivers. River Res. Appl. 2003, 19, 495–507. [Google Scholar] [CrossRef]
- Lancaster, J.; Downes, B.J. Linking the hydraulic world of individual organisms to ecological processes: Putting ecology into ecohydraulics. River Res. Appl. 2010, 26, 385–403. [Google Scholar] [CrossRef]
- Lamouroux, N.; Olivier, J.M.; Persat, H.; PouilLy, M.; Souchon, Y.; Statzner, B. Predicting community characteristics from habitat conditions: Fluvial fish and hydraulics. Freshw. Biol. 1999, 42, 275–299. [Google Scholar] [CrossRef]
- Vázquez, R.F.; Célleri, R.; Samaniego, E.; Vanegas, P.; Orellana, J.; Campozano, L.; Vázquez, A.; Avilés, A. Informe Final del Proyecto: “Gestión de Datos y Modelación Hidrológica Para Soporte al Pronóstico de Alerta Temprana del Sistema Paute Integral”; Universidad de Cuenca: Cuenca, Ecuador, 2012; p. 187. [Google Scholar]
- Studholme, A.M.; Hampel, H.; Finn, D.S.; Vázquez, R.F. Secondary production of caddisflies reflects environmental heterogeneity among tropical Andean streams. Hydrobiologia 2017, 797, 231–246. [Google Scholar] [CrossRef]
- Sotomayor, G.; Hampel, H.; Vázquez, R.F.; Goethals, P.L.M. Multivariate-statistics based selection of a benthic macroinvertebrate index for assessing water quality in the Paute River basin (Ecuador). Ecol. Indic. 2020, 111, 106037. [Google Scholar] [CrossRef] [Green Version]
- Sierra, R. Propuesta Preliminar de un Sistema de Clasificación de Vegetación Para el Ecuador Continental; Proyecto INEFAN/GEF-BIRF y Ecociencia: Quito, Ecuador, 1999; p. 174. [Google Scholar]
- Tenesaca, C.; Quinde, T.; Delgado, G.; Toledo, E.; Delgado, O. Generación del mapa de cobertura y uso del suelo de la provincia del Azuay. Univ. Verdad 2017, 73, 23–37. [Google Scholar]
- Boiten, W. Hydrometry: IHE Delft Lecture Note Series; CRC Press: Boca Raton, FL, USA, 2008; p. 244. ISBN 0415467632. [Google Scholar]
- Blair, T.C.; McPherson, J.G. Grain-size and textural classification of coarse sedimentary particles. J. Sediment. Res. 1999, 69, 6–19. [Google Scholar] [CrossRef]
- Domínguez, E.; Fernández, H.R.; Lillo, F.M. Macroinvertebrados Bentónicos Sudamericanos: Sistemática y Biología; Fundación Miguel Lillo Tucumán: Tucumán, Argentina, 2009; p. 656. ISBN 9506680159. [Google Scholar]
- Steinman, A.D.; Lamberti, G.A.; Leavitt, P.R.; Uzarski, D.G. Biomass and pigments of benthic algae. In Methods in Stream Ecology, 3rd ed.; Richard Hauer, F., Lamberti, G.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 223–241. ISBN 978-0-12-416558-8. [Google Scholar]
- Kennen, J.G.; Riva-Murray, K.; Beaulieu, K.M. Determining hydrologic factors that influence stream macroinvertebrate assemblages in the northeastern US. Ecohydrology 2010, 3, 88–106. [Google Scholar] [CrossRef]
- Jowett, I.G. A method for objectively identifying pool, run, and riffle habitats from physical measurements. N. Z. J. Mar. Freshw. Res. 1993, 27, 241–248. [Google Scholar] [CrossRef]
- Rempel, L.L.; Richardson, J.S.; Healey, M.C. Macroinvertebrate community structure along gradients of hydraulic and sedimentary conditions in a large gravel-bed river. Freshw. Biol. 2000, 45, 57–73. [Google Scholar] [CrossRef]
- Masad, J.A. On the Roughness Reynolds Number Transition Criterion. J. Fluids Eng. 1995, 117, 727–729. [Google Scholar] [CrossRef]
- Wilkes, M.A.; Maddock, I.; Visser, F.; Acreman, M.C. Incorporating hydrodynamics into ecohydraulics: The role of turbulence in the swimming performance and habitat selection of stream-dwelling fish. In Ecohydraulics: An Integrated Approach; Maddock, I., Harby, A., Kemp, P., Wood, P.J., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 9–30. [Google Scholar]
- Brooks, A.J.; Haeusler, T.; Reinfelds, I.; Williams, S. Hydraulic microhabitats and the distribution of macroinvertebrate assemblages in riffles. Freshw. Biol. 2005, 50, 331–344. [Google Scholar] [CrossRef]
- Boyero, L. The quantification of local substrate heterogeneity in streams and its significance for macroinvertebrate assemblages. Hidrobiológica 2003, 499, 161–168. [Google Scholar] [CrossRef]
- Jowett, I.G.; Richardson, J.; Biggs, B.J.; Hickey, C.W.; Quinn, J.M. Microhabitat preferences of benthic invertebrates and the development of generalised Deleatidium spp. habitat suitability curves, applied to four New Zealand rivers. N. Z. J. Mar. Freshw. Res. 1991, 25, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003; p. 110. ISBN 0521891086. [Google Scholar]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data Using CANOCO 5; Cambridge University Press: Cambridge, UK, 2014; p. 362. ISBN 110769440X. [Google Scholar]
- Ter-Braak, C.J.; Šmilauer, P. Canoco Reference Manual and User’s Guide: Software for Ordination (Vol. Version 5.0); Microcomputer Power: Ithaca, NY, USA, 2012. [Google Scholar]
- Acosta, R.; Ríos, B.; Rieradevall, M.; Prat, N. Propuesta de un protocolo de evaluación de la calidad ecológica de ríos andinos (CERA) y su aplicación a dos cuencas en Ecuador y Perú. Limnetica 2009, 28, 35–64. [Google Scholar]
- Jowett, I.G.; Davey, A.J. A comparison of composite habitat suitability indices and generalized additive models of invertebrate abundance and fish presence–habitat availability. Trans. Am. Fish. Soc. 2007, 136, 428–444. [Google Scholar] [CrossRef]
- Statzner, B. How views about flow adaptations of benthic stream invertebrates changed over the last century. Int. Rev. Hydrobiol. 2008, 93, 593–605. [Google Scholar] [CrossRef]
- Eveleens, R.A.; McIntosh, A.R.; Warburton, H.J. Interactive community responses to disturbance in streams: Disturbance history moderates the influence of disturbance types. Oikos 2019, 128, 1170–1181. [Google Scholar] [CrossRef]
- Waringer, J.; Vitecek, S.; Martini, J.; Zittra, C.; Handschuh, S.; Vieira, A.; Kuhlmann, H.C. Hydraulic stress parameters of a cased caddis larva (Drusus biguttatus) using spatio-temporally filtered velocity measurements. Hidrobiológica 2020, 847, 3437–3451. [Google Scholar] [CrossRef]
- Crosskey, R.W. The identification of the larvae of African Simulium. Bull. Word Health Organ. 1962, 27, 483–489. [Google Scholar]
- Ríos-Pulgarín, M.I.; Barletta, M.; Arango-Jaramillo, M.C.; Mancera-Rodríguez, N.J. The role of the hydrological cycle on the temporal patterns of macroinvertebrate assemblages in an Andean foothill stream in Colombia. J. Limnol. 2016, 75 (Suppl. 1), 107–120. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Fonseca, P.E.; Springer, M. A new species of Anacroneuria Klapálek 1909 (Plecoptera: Perlidae) and notes on the altitudinal distribution of the genus in Costa Rica. Zootaxa 2015, 4058, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Forero-Céspedes, A.M.; Gutiérrez, C.; Reinoso-Flórez, G. Composición y estructura de la familia Baetidae (Insecta: Ephemeroptera) en una cuenca andina colombiana. Hidrobiológica 2016, 26, 459–474. [Google Scholar] [CrossRef] [Green Version]
- Fritz, K.M.; Feminella, J.W. Invertebrate colonization of leaves and roots within sediments of intermittent Coastal Plain streams across hydrologic phases. Aquat. Sci. 2011, 73, 459–469. [Google Scholar] [CrossRef]
- González-Trujillo, J.D.; Donato-Rondon, J.C. Changes in invertebrate assemblage structure as affected by the flow regulation of a páramo river. Ann. Limnol.-Int. J. Limnol. 2016, 52, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Chester, E.; Robson, B. Drought refuges, spatial scale and recolonisation by invertebrates in non-perennial streams. Freshw. Biol. 2011, 56, 2094–2104. [Google Scholar] [CrossRef]
- Stubbington, R.; Wood, P.J. Benthic and interstitial habitats of a lentic spring as invertebrate refuges during supra-seasonal drought. Fundam. Appl. Limnol. 2013, 182, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Dostine, P.; Humphrey, C.; Paltridge, R.; Boulton, A. Macroinvertebrate recolonization after re-wetting of a tropical seasonally-flowing stream (Magela Creek, Northern Territory, Australia). Mar. Freshw. Res. 1997, 48, 633–645. [Google Scholar] [CrossRef]
- Cobb, D.; Galloway, T.; Flannagan, J. Effects of discharge and substrate stability on density and species composition of stream insects. Can. J. Fish. Aquat. Sci. 1992, 49, 1788–1795. [Google Scholar] [CrossRef]
- Palmer, C.G.; Maart, B.; Palmer, A.R.; O’Keeffe, J.H. An assessment of macroinvertebrate functional feeding groups as water quality indicators in the Buffalo River, eastern Cape Province, South Africa. Hidrobiológica 1996, 318, 153–164. [Google Scholar] [CrossRef]
- Hemphill, N.; Cooper, S.D. The effect of physical disturbance on the relative abundances of two filter-feeding insects in a small stream. Oecologia 1983, 58, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Gamboa, M.; Chacón, M.M.; Segnini, S. Diet composition of the mature larvae of four Anacroneuria species (Plecoptera: Perlidae) from the Venezuelan Andes. Aquat. Insects 2009, 31 (Suppl. 1), 409–417. [Google Scholar] [CrossRef]
- Granados-Martínez, C.; Zúñiga-Céspedes, B.; Acuña-Vargas, J. Diets and trophic guilds of aquatic insects in Molino River, La Guajira, Colombia. J. Limnology 2016, 75 (Suppl. 1). [Google Scholar] [CrossRef] [Green Version]
- Tomanova, S.; Goitia, E.; Helešic, J. Trophic levels and functional feeding groups of macroinvertebrates in neotropical streams. Hidrobiológica 2006, 556, 251–264. [Google Scholar] [CrossRef]
- Vimos-Lojano, D.; Martínez-Capel, F.; Hampel, H.; Vázquez, R.F. Hydrological influences on aquatic communities at the mesohabitat scale in high Andean streams of southern Ecuador. Ecohydrology 2018, 12, e2033. [Google Scholar] [CrossRef] [Green Version]
- Finn, D.S.; Encalada, A.C.; Hampel, H. Genetic isolation among mountains but not between stream types in a tropical high-altitude mayfly. Freshw. Biol. 2016, 61, 702–714. [Google Scholar] [CrossRef] [Green Version]
- Nieto, M.C. El género Camelobaetidius (Ephemeroptera: Baetidae) en la Argentina. Acta Zool Mexi 2003, 88, 233–255. [Google Scholar]
- Mérigoux, S.; Lamouroux, N.; Olivier, J.M.; Doledec, S. Invertebrate hydraulic preferences and predicted impacts of changes in discharge in a large river. Freshw. Biol. 2009, 54, 1343–1356. [Google Scholar] [CrossRef]
- Finer, M.; Jenkins, C.N. Proliferation of hydroelectric dams in the Andean Amazon and implications for Andes-Amazon connectivity. PLoS ONE 2012, 7, e35126. [Google Scholar] [CrossRef]
- Vuille, M.; Carey, M.; Huggel, C.; Buytaert, W.; Rabatel, A.; Jacobsen, D.; Soruco, A.; Villacis, M.; Yarleque, C.; Elison Timm, O.; et al. Rapid decline of snow and ice in the tropical Andes—Impacts, uncertainties and challenges ahead. Earth Sci. Rev. 2018, 176, 195–213. [Google Scholar] [CrossRef] [Green Version]
- Mora, D.E.; Campozano, L.; Cisneros, F.; Wyseure, G.; Willems, P. Climate changes of hydrometeorological and hydrological extremes in the Paute basin, Ecuadorean Andes. Hydrol. Earth Syst. Sci. 2014, 18, 631–648. [Google Scholar] [CrossRef] [Green Version]
- Davie, A.W.; Mitrovic, S.M. Benthic algal biomass and assemblage changes following environmental flow releases and unregulated tributary flows downstream of a major storage. Mar. Freshw Res. 2014, 65, 1059–1071. [Google Scholar] [CrossRef]
- Spaulding, S.A.; Elwell, E. Increase in Nuisance Blooms and Geographic Expansion of the Freshwater Diatom Didymosphenia Geminata; US Geological Survey Open-File Report 2007–1425; US Geological Survey: Reston, VA, USA, 2007; p. 38. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Unit of Measure | n | Main River | Tributaries | ||||
|---|---|---|---|---|---|---|---|---|
| Average | STD | Range | Average | STD | Range | |||
| Physical-Chemical | ||||||||
| Water temperature | °C | 20 | 11.7 | 1.9 | 9.3–14.0 | 10.1 | 1.6 | 8.2–12.0 |
| pH | -- | 20 | 7.3 | 0.5 | 6.6–8.1 | 7.5 | 0.3 | 6.8–7.8 |
| Oxidation reduction potential | mV | 20 | 304.9 | 66.4 | 243.0–467.0 | 328.8 | 87.4 | 258.0–492.0 |
| Turbidity | NTU | 20 | 1.2 | 2.8 | 0.0–8.5 | 0.1 | 0.2 | 0.0–0.7 |
| Total dissolved solids | g·L−1 | 20 | 0.1 | 0.0 | 0.045–0.101 | 0.1 | 0.0 | 0.046–0.082 |
| Dissolved oxygen | mg·L−1 | 20 | 8.1 | 1.6 | 6.6–10.5 | 8.2 | 1.5 | 6.8–10.2 |
| Saturation oxygen | % | 20 | 77.2 | 12.0 | 64.3–96.1 | 76.6 | 10.4 | 66.2–91.2 |
| Electric conductivity | μS·cm−1 | 20 | 73.2 | 15.4 | 50.2–95.3 | 86.9 | 20.5 | 62.0–129.0 |
| Slope of the reach | % | 10 | 2.9 | 0.4 | 2.4–3.5 | 6.7 | 2.3 | 3.5–9.1 |
| Elevation | m | 10 | 3035.2 | 105.0 | 2919–3194 | 3023.4 | 158.7 | 2826–3236 |
| Hydraulic | ||||||||
| Discharge | m3·s−1 | 20 | 2.12 | 1.16 | 0.78–3.32 | 0.24 | 0.41 | 0.02–1.39 |
| Water depth | cm | 597 | 19.2 | 8.2 | 3.0–39.0 | 18.1 | 8.0 | 3.0–45.0 |
| Water velocity | cm·s−1 | 597 | 55.3 | 45.2 | 0.0–198.6 | 44.8 | 37.5 | 0.0–205.0 |
| Froude number | -- | 597 | 0.434 | 0.398 | 0.0–2.139 | 0.381 | 0.373 | 0.0–2.314 |
| Reynolds number | -- | 597 | 107,204 | 97,175 | 0–504,637 | 75,833 | 68,163 | 0–308,207 |
| Roughness Reynolds number | -- | 597 | 2.4 × 106 | 1.8 × 106 | 0–7.3 × 106 | 2.1 × 106 | 1.7 × 106 | 0–8.7 × 106 |
| Habitat | ||||||||
| Substrate index | -- | 597 | 5.6 | 1.1 | 3.0–7.0 | 5.4 | 1.1 | 3.0–7.0 |
| Shannon-Wiener substrate diversity | -- | 597 | 0.8 | 0.6 | 0.0–2.0 | 0.9 | 0.5 | 0.0–1.8 |
| Boulder (>256 mm) | % | 597 | 32.6 | 45.2 | 0–100 | 21.0 | 37.2 | 0–100 |
| Coarse cobble (256–128 mm) | % | 597 | 12.4 | 23.7 | 0–100 | 14.1 | 23.4 | 0–100 |
| Fine cobble (128–64 mm) | % | 597 | 15.2 | 19.9 | 0–80 | 17.0 | 20.1 | 0–80 |
| Very coarse pebble (64–32 mm) | % | 597 | 14.9 | 18.6 | 0–80 | 17.4 | 18.0 | 0–70 |
| Coarse pebble (32–16 mm) | % | 597 | 9.3 | 12.3 | 0–70 | 10.9 | 14.6 | 0–80 |
| Medium pebble (16–8 mm) | % | 597 | 6.6 | 11.2 | 0–80 | 6.9 | 13.7 | 0–80 |
| Fine pebble (8–4 mm) | % | 597 | 3.3 | 7.0 | 0–40 | 3.3 | 8.5 | 0–60 |
| Granule gravel (4–2 mm) | % | 597 | 2.2 | 7.6 | 0–60 | 4.0 | 12.6 | 0–100 |
| Sand (0.06–2 mm) | % | 597 | 3.4 | 12.4 | 0–100 | 5.0 | 17.0 | 0–100 |
| Silt (0.004–0.06 mm) | % | 597 | 0.2 | 0.9 | 0–10 | 0.3 | 2.1 | 0–20 |
| Clay (<0.004 mm) | % | 597 | 0.02 | 0.3 | 0–5 | 0.03 | 0.6 | 0–10 |
| Organic matter | g m−2 | 597 | 9.9 | 27.7 | 0.06–229.7 | 37.9 | 190.9 | 0.02–755.5 |
| Algae coverage | % | 597 | 20.3 | 32.6 | 0.0–100 | 16.1 | 34.0 | 0.0–100 |
| Bryophytes coverage | % | 597 | 0.0 | 0.0 | 0.0–0.0 | 1.4 | 9.3 | 0.0–90.0 |
| Taxa | Y | T | Relative Abundance | Taxa | Y | T | Relative Abundance |
|---|---|---|---|---|---|---|---|
| Chironomidae | X | X | 40.28 | Austrelmis | X | X | 0.015 |
| Oligochaeta | X | X | 28.32 | Maruina sp.2 | X | X | 0.015 |
| Baetodes | X | X | 12.21 | Thraulodes | X | X | 0.011 |
| Metrichia | X | X | 8.65 | Pseudosuccinea | X | X | 0.011 |
| Andesiops | X | X | 2.21 | Claudioper.la | X | X | 0.011 |
| Simulium | X | X | 2.17 | Limonicola | X | 0.009 | |
| Hydracarina | X | X | 1.15 | Hexatomini | X | X | 0.009 |
| Palpomyia | X | X | 0.98 | Nectopsyche | X | X | 0.009 |
| Austrolimnius | X | X | 0.60 | Cyphon | X | X | 0.009 |
| Leptohyphes | X | X | 0.49 | Sphaeriidae | X | X | 0.008 |
| Neoplasta | X | X | 0.47 | Limonia | X | X | 0.008 |
| Paltostoma | X | X | 0.38 | Neoelmis | X | X | 0.008 |
| Collembola | X | X | 0.36 | Xiphocentronidae | X | X | 0.007 |
| Molophilus | X | X | 0.27 | Girardia | X | X | 0.007 |
| Atopsyche | X | X | 0.27 | Hexanchorus | X | X | 0.006 |
| Camelobaetidius | X | X | 0.20 | Cyrnellus | X | X | 0.005 |
| Heterelmis | X | X | 0.13 | Onychelmis | X | X | 0.005 |
| Ecuaphlebia | X | X | 0.10 | Alluaudomyia | X | X | 0.005 |
| Huleechius | X | X | 0.08 | Pericoma | X | X | 0.005 |
| Gigantodax | X | X | 0.08 | Polycentropus | X | X | 0.004 |
| Hydra | X | X | 0.07 | Pheneps | X | X | 0.004 |
| Anacroneuria | X | X | 0.07 | Haplohyphes | X | X | 0.004 |
| Contulma | X | X | 0.07 | Farrodes | X | X | 0.003 |
| Phylloicus | X | X | 0.06 | Anomalocosmoecus | X | X | 0.003 |
| Cailloma | X | X | 0.04 | Tricorythodes | X | X | 0.003 |
| Hatia | X | X | 0.03 | Helicopsyche | X | X | 0.003 |
| Hyalella | X | X | 0.02 | Bezzia | X | X | 0.001 |
| Neotrichia | X | X | 0.02 | Forcipomyia | X | X | 0.001 |
| Limnophora | X | X | 0.02 | Smicridea | X | X | 0.001 |
| Ostracoda | X | X | 0.02 | Triplectides | X | X | 0.001 |
| Maruina sp.1 | X | X | 0.02 |
| (a) Main River | ||||
| Statistical Parameter | Axis | |||
| 1 | 2 | 3 | 4 | |
| Eigenvalue | 0.104 | 0.039 | 0.008 | 0.005 |
| Explained variation (cumulative) | 10.4 | 14.3 | 15 | 15.5 |
| Pseudo-canonical correlation | 0.68 | 0.44 | 0.40 | 0.28 |
| Explained fitted variation (cumulative) | 66.2 | 90.8 | 95.9 | 98.8 |
| Variable (%) | Explanation (%) | Contribution | pseudo-F | p |
| Froude number (Fr) | 7.9 | 39.0 | 25.5 | 0.002 |
| Shannon-Wiener substrate diversity (SuD) | 2.4 | 11.8 | 7.9 | 0.002 |
| Algae coverage (Algae) | 2.3 | 11.3 | 8.0 | 0.002 |
| Organic matter | 1.6 | 8.1 | 5.5 | 0.002 |
| Reynolds number (Re) | 1.5 | 7.4 | 5.1 | 0.002 |
| (b) Tributaries | ||||
| Statistical parameter | Axis | |||
| 1 | 2 | 3 | 4 | |
| Eigenvalue | 0.066 | 0.041 | 0.012 | 0.008 |
| Explained variation (cumulative) | 6.6 | 10.7 | 11.9 | 12.7 |
| Pseudo-canonical correlation | 0.71 | 0.46 | 0.40 | 0.37 |
| Explained fitted variation (cumulative) | 48.8 | 79.0 | 88.1 | 93.7 |
| Variable | Explanation (%) | Contribution (%) | pseudo-F | p |
| Froude number (Fr) | 4.5 | 23.9 | 13.9 | 0.002 |
| Fine cobble substrate | 3.0 | 15.3 | 9.4 | 0.002 |
| Substrate index (SI) | 1.5 | 7.6 | 4.8 | 0.002 |
| Reynolds number (Re) | 1.3 | 6.8 | 4.2 | 0.002 |
| Shannon-Wiener substrate diversity (SuD) | 1.2 | 6.0 | 3.8 | 0.002 |
| Algae coverage (Algae) | 1.1 | 5.4 | 3.4 | 0.002 |
| Bryophytes coverage (Bryo) | 1.0 | 5.1 | 3.3 | 0.002 |
| Test for Significance | Test for Non-Linearity | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Taxa | Model | Res. dev. | %TD Explained | Res. df | F Ratio | p | Model | Res. dev. | Res. df | F Ratio | p |
| Baetodes | Null | 1192 | 0.0 | 596.0 | |||||||
| Fr | 970 | 18.6 | 594.2 | 58.8 | <0.001 | Fr | 881 | 592.9 | 78.1 | <0.001 | |
| Fr + SI | 815 | 31.6 | 591.0 | 51.8 | <0.001 | SI | 1043 | 593.1 | 39.3 | <0.001 | |
| Fr + SI + Re | 790 | 33.7 | 587.9 | 36.8 | <0.001 | Re | 937 | 592.9 | 61.1 | <0.001 | |
| Fr + SI + Re + Algae | 781 | 34.5 | 585.0 | 28.7 | <0.001 | Algae | 1105 | 593.0 | 24.8 | <0.001 | |
| Simulium | Null | 1336 | 0.0 | 590.0 | |||||||
| Fr | 1113 | 16.7 | 588.3 | 54.7 | <0.001 | Fr | 1001 | 587.0 | 76.8 | <0.001 | |
| Fr + SI | 920 | 31.1 | 585.1 | 50.6 | <0.001 | SI | 1235 | 587.1 | 25.7 | <0.001 | |
| Fr + SI + Re | 912 | 31.7 | 581.8 | 33.4 | <0.001 | Re | 1163 | 586.8 | 36.1 | <0.001 | |
| Fr + SI + Re + Algae | 903 | 32.4 | 578.8 | 25.7 | <0.001 | Algae | 1274 | 587.0 | 18.1 | <0.001 | |
| Anacroneuria | Null | 2443 | 0.0 | 595.0 | |||||||
| Fr | 2329 | 4.7 | 593.3 | 20.0 | <0.001 | Fr | 2292 | 592.1 | 22.0 | <0.001 | |
| Fr + SI | 2126 | 13.0 | 590.0 | 19.3 | <0.001 | SI | 2246 | 592.0 | 25.9 | <0.001 | |
| Fr + SI + Re | 2106 | 13.8 | 586.6 | 13.0 | <0.001 | Re | 2391 | 591.8 | 11.4 | <0.001 | |
| Fr + SI + Re + Algae | 1994 | 18.4 | 583.8 | 13.1 | <0.001 | Algae | 2388 | 592.1 | 12.9 | <0.001 | |
| Andesiops | Null | 1146 | 0.0 | 593.0 | |||||||
| Fr | 1012 | 11.7 | 591.2 | 29.1 | <0.001 | Fr | 1012 | 590.1 | 29.0 | <0.001 | |
| Fr + SI | 983 | 14.2 | 587.9 | 16.7 | <0.001 | SI | 1007 | 589.9 | 28.1 | <0.001 | |
| Fr + SI + Re | 980 | 14.5 | 584.6 | 11.0 | <0.001 | Re | 1105 | 589.9 | 8.4 | <0.001 | |
| Fr + SI + Re + Algae | 966 | 15.7 | 581.5 | 9.1 | <0.001 | Algae | 1108 | 590.0 | 8.1 | <0.001 | |
| Camelobaetidius | Null | 2249 | 0.0 | 594.0 | |||||||
| Fr | 2041 | 9.2 | 592.2 | 26.4 | <0.001 | Fr | 2039 | 591.2 | 25.9 | <0.001 | |
| Fr + SI | 1971 | 12.4 | 589.2 | 16.6 | <0.001 | SI | 2102 | 591.1 | 18.4 | <0.001 | |
| Fr + SI + Re | 1909 | 15.1 | 586.3 | 13.5 | <0.001 | Re | 2070 | 591.1 | 21.6 | <0.001 | |
| Fr + SI + Re + Algae | 1825 | 18.9 | 583.7 | 13.2 | <0.001 | Algae | 2083 | 591.4 | 22.7 | <0.001 | |
| Ecuaphlebia | Null | 1756 | 0.0 | 595.0 | |||||||
| Fr | 1679 | 4.4 | 592.6 | 10.1 | <0.001 | Fr | 1627 | 592.7 | 23.7 | <0.001 | |
| Fr + SI | 1577 | 10.2 | 590.6 | 13.9 | <0.001 | SI | 1492 | 592.1 | 38.9 | <0.001 | |
| Fr + SI + Re | 1562 | 11.0 | 587.7 | 9.8 | <0.001 | Re | 1699 | 592.3 | 9.9 | <0.001 | |
| Fr + SI + Re + Algae | 1498 | 14.7 | 584.7 | 9.6 | <0.001 | Algae | 1736 | 592.0 | 4.7 | <0.001 | |
| Atopsyche | Null | 1439 | 0.0 | 593.0 | |||||||
| Fr | 1354 | 5.9 | 591.2 | 16.4 | <0.001 | Fr | 1352 | 590.2 | 16.3 | <0.001 | |
| Fr + SI | 1302 | 9.5 | 588.1 | 11.8 | <0.001 | SI | 1341 | 590.0 | 17.1 | <0.001 | |
| Fr + SI + Re | 1262 | 12.3 | 585.0 | 10.1 | <0.001 | Re | 1368 | 590.0 | 13.1 | <0.001 | |
| Fr + SI + Re + Algae | 1252 | 13.0 | 581.9 | 7.9 | <0.001 | Algae | 1415 | 590.0 | 5.8 | <0.001 | |
| Palpomyia | Null | 1950 | 0.0 | 594.0 | |||||||
| Fr | 1686 | 13.5 | 591.2 | 33.6 | <0.001 | Fr | 1623 | 591.7 | 65.1 | <0.001 | |
| Fr + SI | 1677 | 14.0 | 589.4 | 21.8 | <0.001 | SI | 1605 | 591.0 | 53.6 | <0.001 | |
| Fr + SI + Re | 1643 | 15.7 | 586.5 | 16.5 | <0.001 | Re | 1621 | 591.5 | 60.5 | <0.001 | |
| Fr + SI + Re + Algae | 1562 | 19.9 | 583.4 | 15.3 | <0.001 | Algae | 1791 | 590.9 | 25.9 | <0.001 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez, R.F.; Vimos-Lojano, D.; Hampel, H. Habitat Suitability Curves for Freshwater Macroinvertebrates of Tropical Andean Rivers. Water 2020, 12, 2703. https://doi.org/10.3390/w12102703
Vázquez RF, Vimos-Lojano D, Hampel H. Habitat Suitability Curves for Freshwater Macroinvertebrates of Tropical Andean Rivers. Water. 2020; 12(10):2703. https://doi.org/10.3390/w12102703
Chicago/Turabian StyleVázquez, Raúl F., Diego Vimos-Lojano, and Henrietta Hampel. 2020. "Habitat Suitability Curves for Freshwater Macroinvertebrates of Tropical Andean Rivers" Water 12, no. 10: 2703. https://doi.org/10.3390/w12102703
APA StyleVázquez, R. F., Vimos-Lojano, D., & Hampel, H. (2020). Habitat Suitability Curves for Freshwater Macroinvertebrates of Tropical Andean Rivers. Water, 12(10), 2703. https://doi.org/10.3390/w12102703