Nitrogen Retention in Mesocosm Sediments Received Rural Wastewater Associated with Microbial Community Response to Plant Species



 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Species
2.2. Mesocosms Plastic Drum Sediment (MPDS) Experiment
2.3. Sediment Sampling and Chemical Analyses
2.4. DNA Extraction and MiSeq Sequencing of 16S rRNA Gene Amplicons
2.5. Pyrosequence Data Analysis and Putative Function Prediction
2.6. Statistical Analyses
3. Results
3.1. Nutrient Retention in Sediment
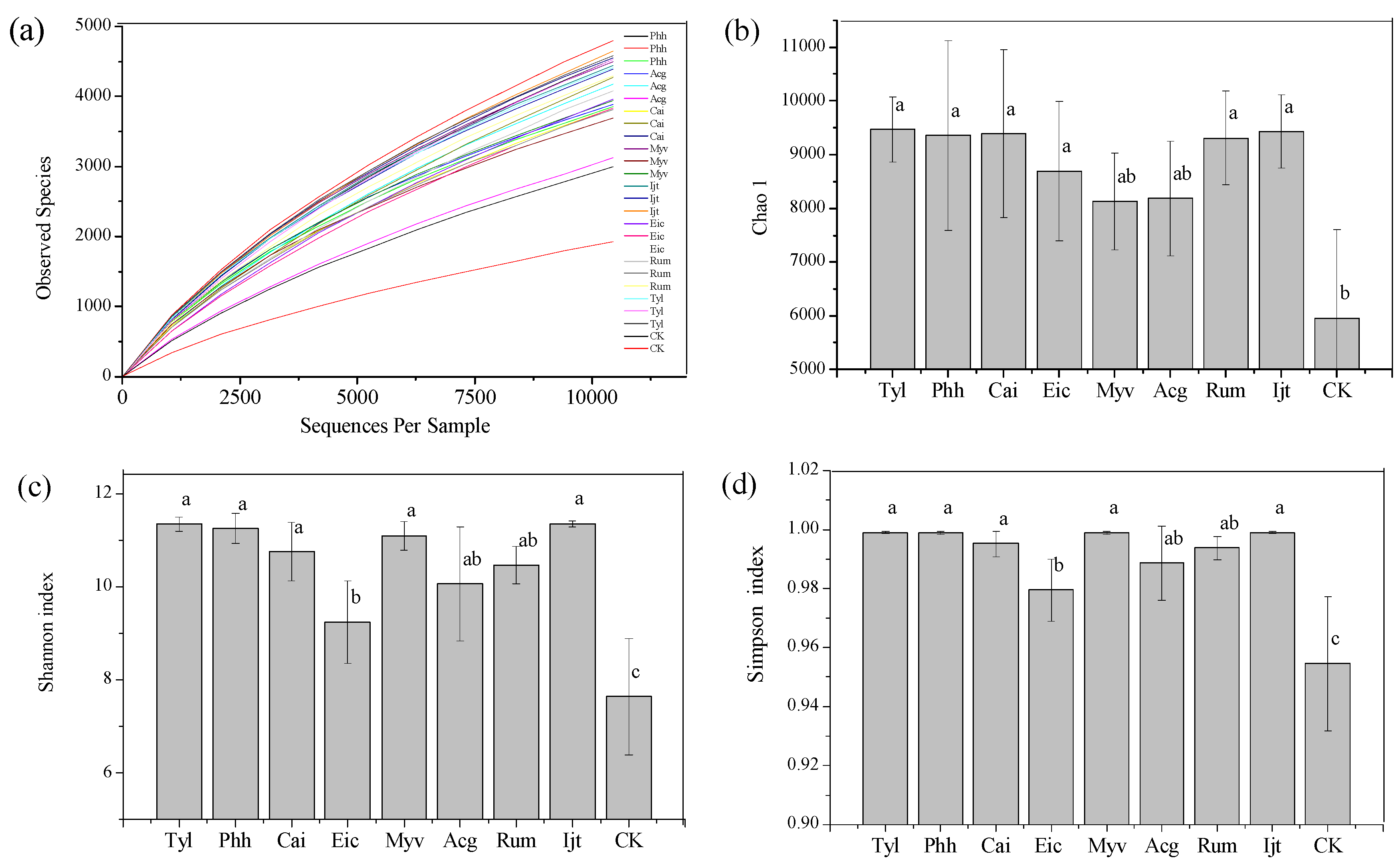
3.2. Taxon Richness and Diversity Coverage Assessment
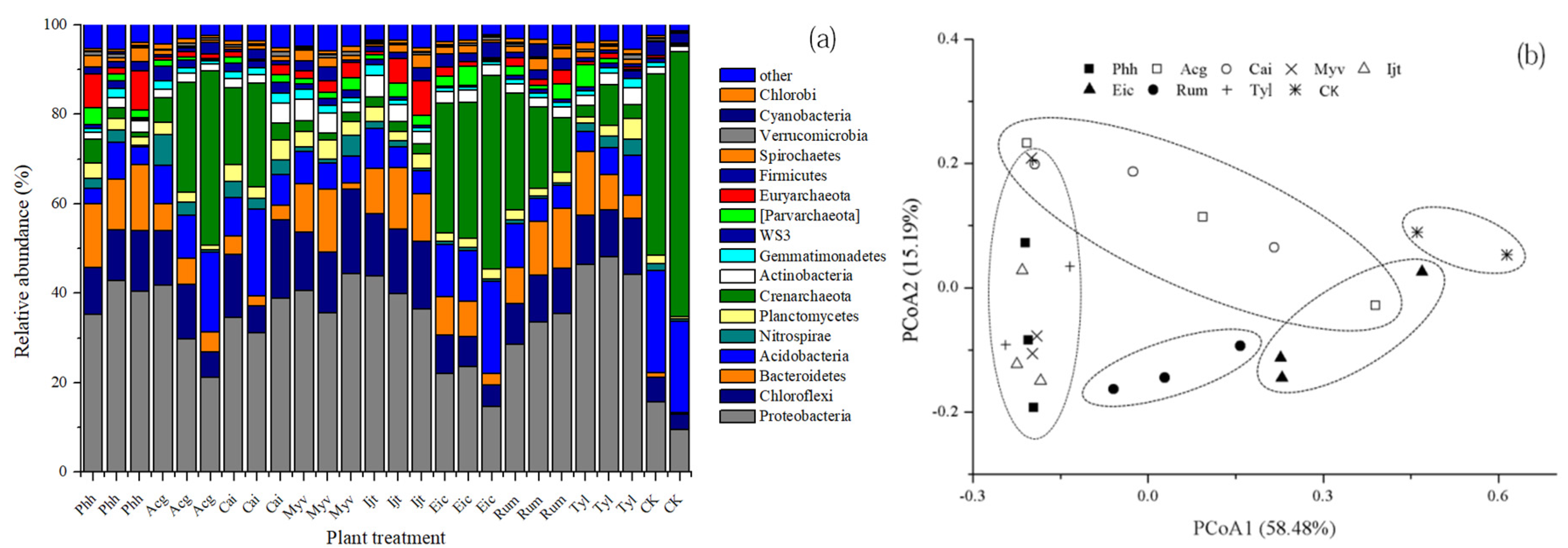
3.3. Prokaryotic Community Composition
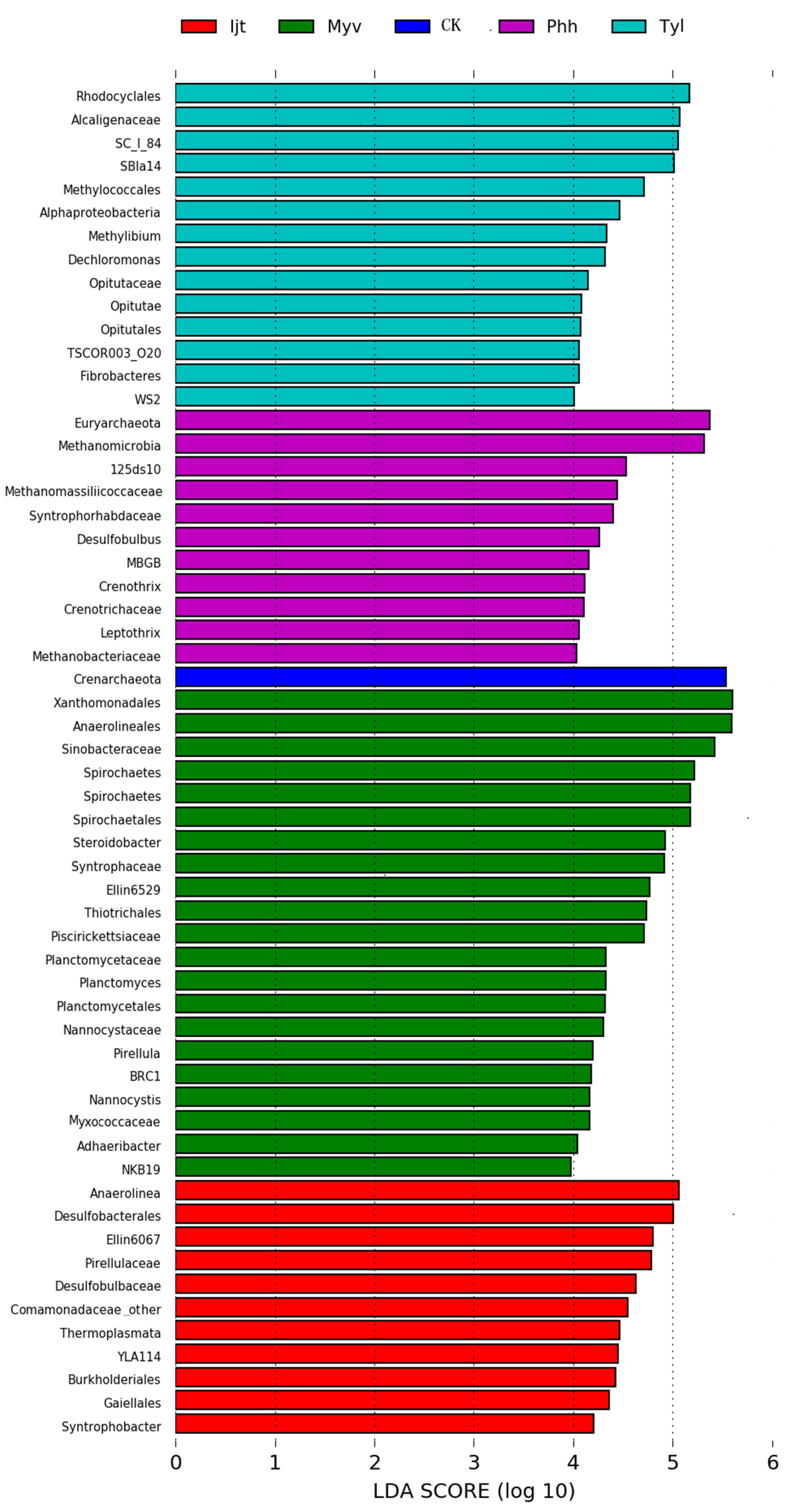
3.4. Abundance Differences Among Sediments Treated with Different Plant Species
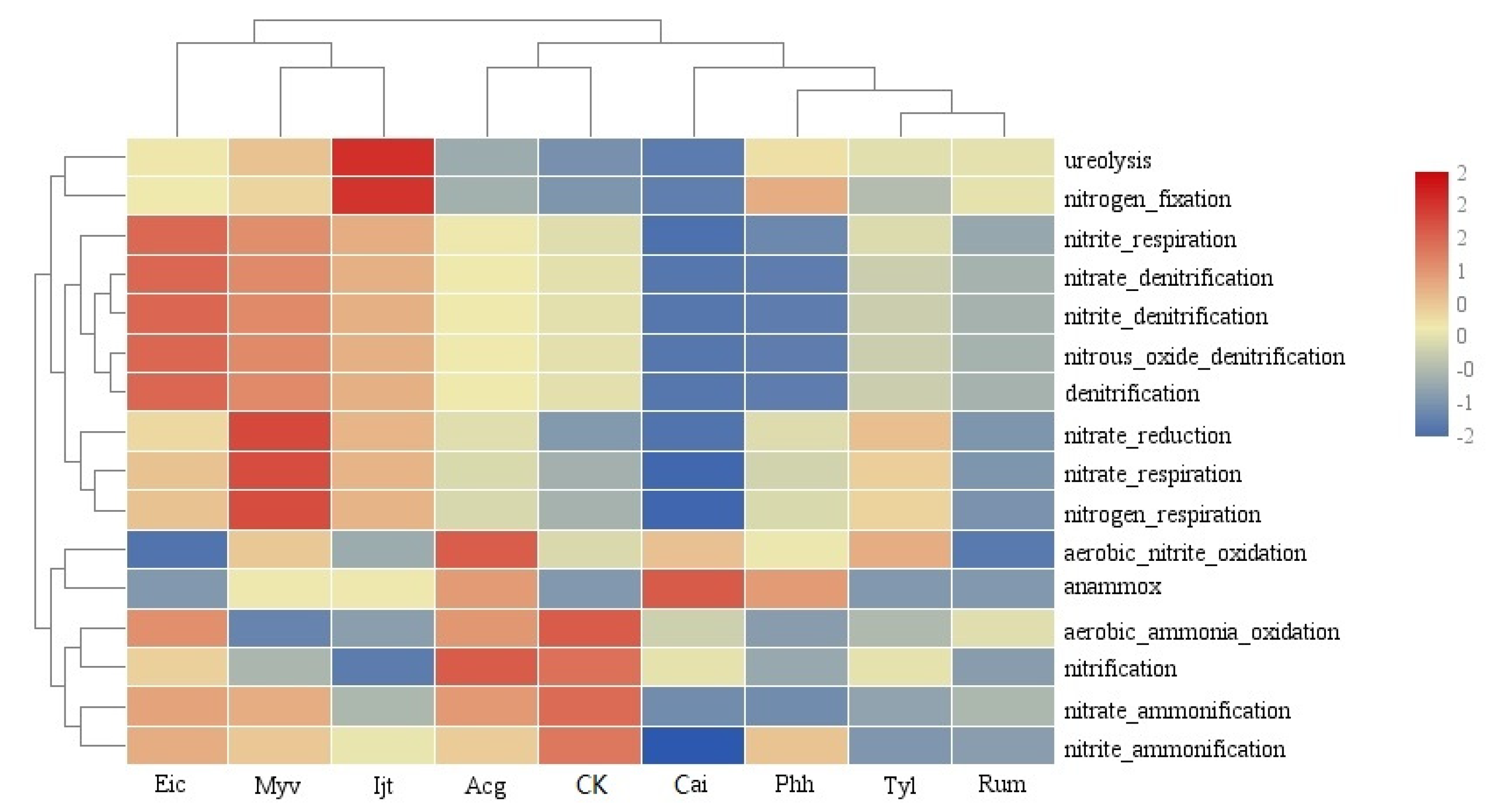
3.5. Effect of Plant Species on Putative Functions of Sediment Prokaryotic Communities
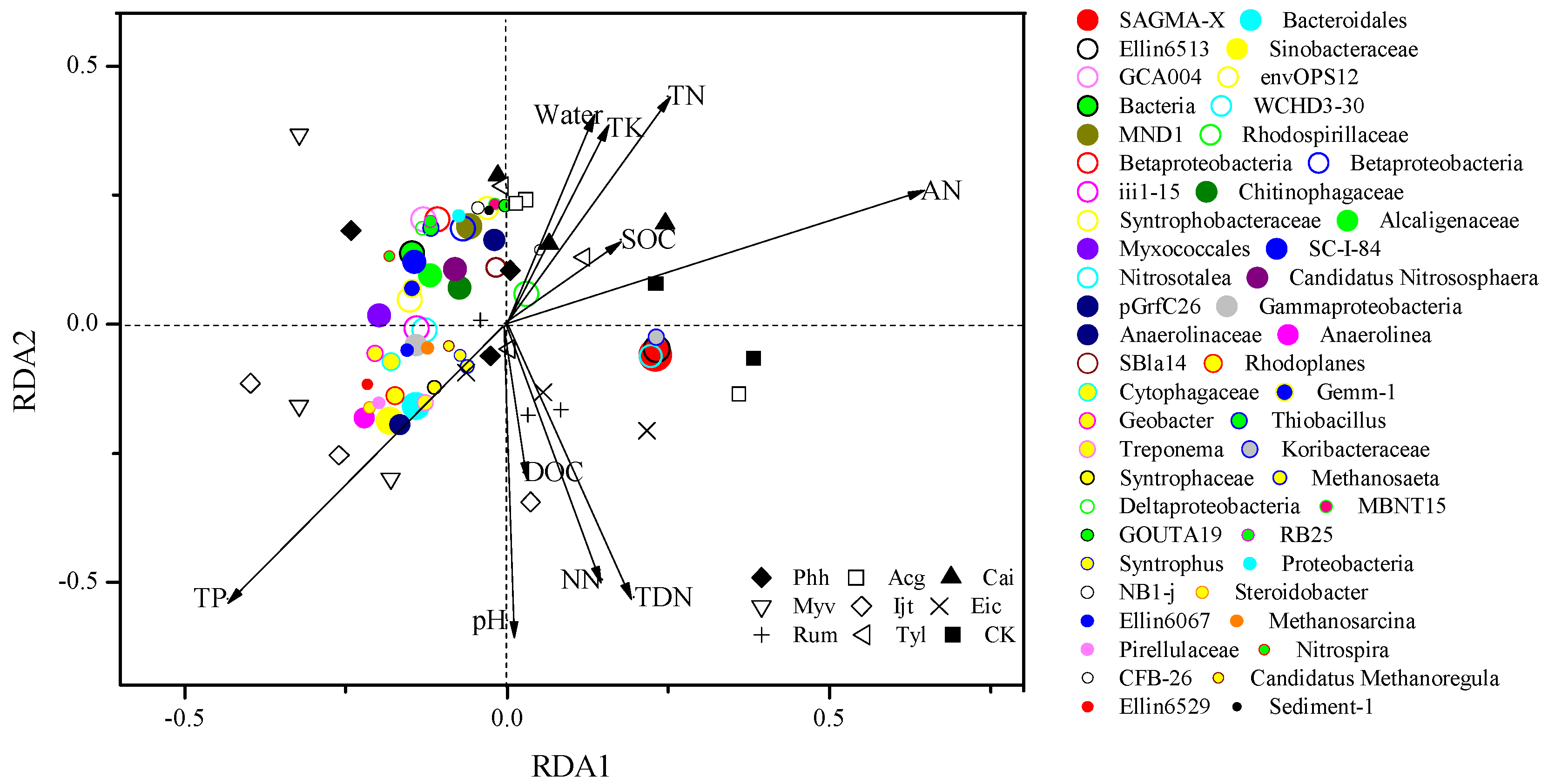
3.6. Correlations between Environmental Variables and Microbial Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Akpor, O.B.; Muchie, M. Environmental and public health implications of wastewater quality. Afr. J. Biotechnol. 2011, 10, 2379–2387. [Google Scholar]
- Carty, A.; Scholz, M.; Hea, K.; Gouriveau, F.; Mustafa, A. The universal design, operation and maintenance guidelines for farm constructed wetlands (FCW) in temperate climates. Bioresour. Technol. 2008, 99, 6780–6792. [Google Scholar] [CrossRef] [PubMed]
- Rai, U.N.; Tripathi, R.D.; Singh, N.K.; Upadhyay, A.K.; Dwivedi, S.K.; Shukla, M.K.; Mallick, S.; Singh, S.; Nautiyal, C.S. Constructed wetland as an ecotechnological tool for pollution treatment for conservation of Ganga river. Bioresour. Technol. 2013, 148, 535–541. [Google Scholar] [CrossRef] [PubMed]
- Vymazal, J. Constructed wetlands for wastewater treatment: Five decades of experience. Environ. Sci. Technol. 2011, 45, 61–69. [Google Scholar] [CrossRef]
- Wu, H.M.; Zhang, J.; HaoNgo, H.; Hu, Z.; Liang, S.; Fan, J.L.; Liu, H. A review on the sustainability of constructed wetlands for wastewater treatment: Design and operation. Bioresour. Technol. 2015, 175, 594–601. [Google Scholar] [CrossRef]
- Kröger, R.; Holland, M.M.; Moore, M.T.; Cooper, C.M. Hydrological variability and agricultural drainage ditch inorganic nitrogen reduction capacity. J. Environ. Qual. 2007, 36, 1646–1652. [Google Scholar] [CrossRef] [Green Version]
- Silvan, N.; Vasander, H.; Laine, J. Vegetation is the main factor in nutrient retention in a constructed wetland buffer. Plant Soil 2004, 258, 179–187. [Google Scholar] [CrossRef]
- Moore, M.; Kröger, R.; Locke, M.A.; Cullum, R.F.; Steinriede, R.W.; Testa, S.; Lizotte, R.E., Jr.; Bryan, C.T.; Cooper, C.M. Nutrient mitigation capacity in Mississippi Delta, USA drainage ditches. Environ. Pollut. 2010, 158, 75–184. [Google Scholar] [CrossRef]
- Kumwimba, M.N.; Zhu, B. Effectiveness of Vegetated Drainage Ditches for Domestic Sewage Effluent Mitigation. B. Environ. Contam. Tox. 2017, 98, 682–689. [Google Scholar] [CrossRef]
- Chen, L.; Liu, F.; Wang, Y.; Li, X.; Zhang, S.L.; Li, Y.; Wu, J.S. Nitrogen removal in an ecological ditch receiving agricultural drainage in subtropical central China. Ecol. Eng. 2015, 82, 487–492. [Google Scholar] [CrossRef]
- Hu, H.X.; Zhu, X.H.; Huang, J.Y.; Ma, Y.H.; Yan, P. Research of ditch ecological interception of nitrogen and phosphorus. J. Soil Water Conserv. 2010, 24, 141–145. [Google Scholar]
- Brix, H. Functions of macrophytes in constructed wetlands. Water Sci. Technol. 1994, 29, 71–78. [Google Scholar] [CrossRef]
- Surrency, D. Evaluation of aquatic plants for constructed wetlands. In Constructed Wetlands for Water Quality Improvement; Moshiri, G.A., Ed.; Lewis Publishers, Inc.: Boca Raton, FL, USA, 1993; pp. 349–357. [Google Scholar]
- Bai, Y.H.; Huo, Y.; Liao, K.L.; Qu, J.H. Influence of microbial community diversity and function on pollutant removal in ecological wastewater treatment. Appl. Microbiol. Biotechnol. 2017, 101, 7293–7302. [Google Scholar] [PubMed]
- Somers, E.; Vanderleyden, J.; Srinivisam, M. Rhizosphere bacterial signaling: A love parade beneath our feet. Crit. Rev. Microbiol. 2004, 30, 205–240. [Google Scholar]
- Briones, A.M.; Okabe, S.; Umemiya, Y.; Ramsing, N.; Reichardt, W.; Okuyama, H. Influence of different cultivars on populations of ammonia-oxidizing bacteria in the root environment of rice. Appl. Environ. Microbiol. 2002, 68, 3067–3075. [Google Scholar] [PubMed] [Green Version]
- Bremer, C.; Braker, G.; Matthies, D.; Reuter, A.; Engels, C.; Conrad, R. Impact of plant functional group, plant species, and sampling time on the composition of nirK-type denitrifier communities in soil. Appl. Environ. Microbiol. 2007, 73, 6876–6884. [Google Scholar]
- Ahn, C.; Gillevet, P.M.; Sikaroodi, M. Molecular characterization of microbial communities in treatment microcosm wetlands as influenced by macrophytes and phosphorus loading. Ecol. Indic. 2007, 7, 852–863. [Google Scholar] [CrossRef]
- Ansola, G.; Arroyo, P.; de Miera, L.E.S. Characterisation of the soil bacterial community structure and composition of natural and constructed wetlands. Sci. Total. Environ. 2014, 473–474, 63–71. [Google Scholar] [CrossRef]
- Zhao, Y.J.; Liu, B.; Zhang, W.G.; Hu, C.W.; An, S.Q. Effects of plant and influent C: N: P ratio on microbial diversity in pilot-scale constructed wetlands. Ecol. Eng. 2010, 36, 441–449. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16SrRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of highthroughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louca, S.; Parfrey, L.W.; Doebeli, M. Decoupling function and taxonomy in the global ocean microbiome. Science 2016, 353, 1272–1277. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, E.R.; Moore, M.T.; Cooper, C.M.; Smith, S., Jr.; Shields, F.D., Jr.; Drouillard, K.G.; Schulz, R. Vegetated agricultural drainage ditches for the mitigation of pyrethroid-associated runoff. Environ. Toxicol. Chem. 2005, 24, 2121–2127. [Google Scholar] [CrossRef] [Green Version]
- Iseyemi, O.O.; Farris, J.L.; Moore, M.T.; Choi, S. Nutrient mitigation efficiency in agricultural drainage ditches: An influence of landscape management. Bull. Environ. Contam. Toxicol. 2016, 96, 750–756. [Google Scholar] [CrossRef]
- Berg, G.; Smalla, K. Plant species and soil type cooperatively shape the structure and function of microbial communities in the rhizosphere. FEMS Microbiol. Ecol. 2009, 68, 1–13. [Google Scholar]
- Monteiro, M.; Séneca, J.; Magalhães, C. The history of aerobic ammonia oxidizers: From the first discoveries to today. J. Microbiol. 2014, 52, 537–547. [Google Scholar] [CrossRef]
- Liu, F.; Xiao, R.L.; Wang, Y.; Li, Y.; Zhang, S.L.; Luo, Q.; Wu, J.S. Effect of a novel constructed drainage ditch on the phosphorus sorption capacity of ditch soils in an agricultural headwater catchment in subtropical central China. Eng. Ecol. 2013, 58, 69–76. [Google Scholar] [CrossRef]
- Zhang, J.P.; Liu, B.; Zhou, X.H.; Chu, J.Y.; Li, Y.M.; Wang, M.Y. Effect of emergent aquatic plants on abundance and community structure of ammonia-oxidising microorganisms. Ecol. Eng. 2015, 81, 504–513. [Google Scholar] [CrossRef]
- Tourna, M.; Stieglmeier, M.; Spang, A.; Könneke, M.; Schintlmeister, A.; Urich, T.; Engel, M.; Schloter, M.; Wagner, M.; Richter, A.; et al. Nitrososphaera viennensis, an ammonia oxidizing archaeon from soil. Proc. Natl. Acad. Sci. USA 2011, 108, 8420–8425. [Google Scholar] [PubMed] [Green Version]
- Seitzinger, S. Nitrogen cycle: Out of reach. Nature 2008, 452, 152–163. [Google Scholar]
- Ma, Q.; Qu, Y.Y.; Shen, W.L.; Zhang, Z.J.; Wang, J.W.; Liu, Z.Y.; Li, D.K.; Li, H.J.; Zhou, J.T. Bacterial community compositions of coking wastewater treatment plants in steel industry revealed by Illumina high-throughput sequencing. Bioresour. Technol. 2015, 179, 436–443. [Google Scholar] [PubMed]
- Khan, T.; Horiba, Y.; Yamamoto, M.; Hiraishi, A. Members of the family Comamonadaceae asprimary poly(3-hydroxybutyrate-co-3hydroxyvalerate)-degrading denitrifiers in activated sludge as revealed by a polyphasic approach. Appl. Environ. Microbiol. 2002, 68, 3206–3214. [Google Scholar]
- Willems, A. The family Comamonadaceae. In The Prokaryote; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: New York, NY, USA, 2014; pp. 777–851. [Google Scholar]
- Hwang, C.; Wu, W.M.; Gentry, T.J.; Carley, J.; Carroll, S.L.; Schadt, C.; Watson, D.; Jardine, P.M.; Zhou, J.; Hickey, R.F.; et al. Changes in bacterial community structure correlate with initial operating conditions of a field-scale denitrifying fluidized bed reactor. Appl. Microbiol. Biotechnol. 2006, 71, 748–760. [Google Scholar]
- Lu, H.J.; Chandran, K.; Stensel, D. Microbial ecology of denitrification in biological wastewater treatment. Water Res. 2014, 64, 237–254. [Google Scholar]
- Carl-Eric, W.; Werner, L. Unexpected Dominance of Elusive Acidobacteria in Early Industrial Soft Coal Slags. Front. Microbiol. 2017, 8, 1023. [Google Scholar] [CrossRef]
- Nedashkovskaya, O.I.; Balabanova, L.A.; Zhukova, N.V.; Kim, S.J.; Bakunina, I.Y.; Rhee, S.K. Flavobacterium ahnfeltiae sp. nov., a new marine polysaccharide-degrading bacterium isolated from a Pacific red alga. Arch. Microbiol. 2014, 196, 745–752. [Google Scholar]
- Sack, E.L.W.; van der Wielen, P.W.J.J.; van der Kooij, D. Flavobacterium johnsoniae as a model organism for characterizing biopolymer utilization in oligotrophic freshwater environments. Appl. Environ. Microbiol. 2011, 77, 6931–6938. [Google Scholar]
- Sun, B.; Ko, K.; Ramsay, J.A. Biodegradation of 1,4-dioxane by a Flavobacterium. Biodegradation 2011, 22, 651–659. [Google Scholar]
- Repert, D.A.; Underwood, J.; Smith, R.L.; Song, B. Nitrogen cycling processes and microbial community composition in bed sediments in the Yukon River at Pilot Station. J. Geophys. Res. Biogeosci. 2014, 119, 2328–2344. [Google Scholar] [CrossRef]
- Cheremisinoff, N.P. Nitrification and denitrification in the activated sludge process. Biotechnol. Waste Wastewater Treat. 1997, 151–188. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Pond Water | Rural Wastewater | Sediment (mg∙kg−1) |
|---|---|---|---|
| pH | 7.85 | 8.01 | - |
| DO | 7.6 | 0.35 | - |
| TDS | 266 | 885 | - |
| EC | 384 | 1452 | - |
| TN (mg∙L−1) | 8.86 | 58.64 ± 2.21–65.42 ± 3.52 | 575.65 ± 25 |
| TP (mg∙L−1) | 0.58 | 4.87 ± 0.25–5.92 ± 0.58 | 498.47 ± 25 |
| NH4-N (mg∙L−1) | 0.38 | 22.73 ± 1.75–28.42 ± 2.07 | 56.24 ± 1.25 |
| NO3-N (mg∙L−1) | 0.92 | 1.97 ± 0.20–2.84 ± 0.29 | 15.24 ± 1.42 |
| pH | Water Content (%) | NH4+-N (mg∙kg−1) | NO3−-N (mg∙kg−1) | TDN (mg∙kg−1) | TN (g∙kg−1) | TP (g∙kg−1) | TK (g∙kg−1) | DOC (mg∙kg−1) | Organic C (g∙kg−1) | |
|---|---|---|---|---|---|---|---|---|---|---|
| Tyl | 8.47 ± 0.16 a | 50.92 ± 8.91 a | 0.02 ± 0.02 ab | 0.04 ± 0.06 a | 2.25 ± 0.57 a | 0.89 ± 0.19 ab | 0.64 ± 0.04 ab | 18.11 ± 1.01 ab | 10.62 ± 1.27 a | 13.20 ± 3.37 a |
| Phh | 8.28 ± 0.03 a | 51.25 ± 3.35 | 0.04 ± 0.02 b | 8.64 ± 2.59 c | 11.66 ± 3.00 c | 0.91 ± 0.02 ab | 0.63 ± 0.02 ab | 16.99 ± 0.76 ab | 9.72 ± 1.44 a | 13.83 ± 0.76 a |
| Cai | 8.44 ± 0.07 a | 44.65 ± 2.30 a | 0.05 ± 0.03 b | 1.13 ± 0.49 ab | 3.37 ± 1.17 ab | 0.93 ± 0.10 ab | 0.61 ± 0.04 ab | 18.58 ± 1.60 ab | 9.85 ± 1.11 a | 14.25 ±1 53 a |
| Eic | 8.55 ± 0.13 a | 42.58 ± 4.23 a | 0.02 ± 0.00 ab | 5.70 ± 1.56 bc | 9.27 ± 1.96 bc | 0.89 ± 0.13 ab | 0.67 ± 0.01 b | 17.61 ± 2.23 ab | 10.97 ± 1.08 a | 14.06 ± 3.10 a |
| Myv | 8.52 ± 0.22 a | 46.01 ± 12.72 a | 0.00 ± 0.00 a | 2.48 ± 2.70 ab | 4.67 ± 3.70 ab | 0.90 ± 0.10 ab | 0.68 ± 0.05 b | 17.34 ± 1.27 a | 10.68 ± 2.32 a | 14.08 ± 0.66 a |
| Acg | 8.33 ± 0.06 a | 44.21 ± 4.79 a | 0.05 ± 0.04 b | 6.09 ± 5.68 bc | 9.62 ± 7.10 bc | 1.18 ± 0.09 b | 0.61 ± 0.04 ab | 20.24 ± 1.75 b | 14.15 ± 10.92 a | 17.14 ± 2.53 a |
| Rum | 8.48 ± 0.22 a | 46.95 ± 16.55 a | 0.01 ± 0.01 a | 4.89 ± 0.84 abc | 8.39 ± 1.51 abc | 0.97 ± 0.31 ab | 0.65 ± 0.06 ab | 16.99 ± 0.76 a | 12.48 ± 4.37 a | 15.70 ± 6.53 a |
| Ijt | 8.57 ± 0.04 a | 39.19 ± 5.78 a | 0.00 ± 0.00 a | 4.83 ± 2.64 abc | 8.03 ± 3.17 abc | 0.87 ± 0.08 a | 0.68 ± 0.05 b | 15.97 ± 0.04 a | 13.77 ± 1.02 a | 14.91 ± 2.00 a |
| CK | 8.54 ± 0.06 a | 48.55 ± 5.50 a | 0.03 ± 0.01 ab | 1.41 ± 1.99 ab | 3.83 ± 2.41 ab | 1.09 ± 0.09 ab | 0.60 ± 0.01 a | 18.21 ± 0.87 ab | 8.84 ± 0.69 a | 17.78 ± 2.68 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Z.; Hu, L.; Li, J.; Kumwimba, M.N.; Tang, J.; Zhu, B. Nitrogen Retention in Mesocosm Sediments Received Rural Wastewater Associated with Microbial Community Response to Plant Species. Water 2020, 12, 3035. https://doi.org/10.3390/w12113035
Dong Z, Hu L, Li J, Kumwimba MN, Tang J, Zhu B. Nitrogen Retention in Mesocosm Sediments Received Rural Wastewater Associated with Microbial Community Response to Plant Species. Water. 2020; 12(11):3035. https://doi.org/10.3390/w12113035
Chicago/Turabian StyleDong, Zhixin, Lei Hu, Jianmei Li, Mathieu Nsenga Kumwimba, Jialiang Tang, and Bo Zhu. 2020. "Nitrogen Retention in Mesocosm Sediments Received Rural Wastewater Associated with Microbial Community Response to Plant Species" Water 12, no. 11: 3035. https://doi.org/10.3390/w12113035
APA StyleDong, Z., Hu, L., Li, J., Kumwimba, M. N., Tang, J., & Zhu, B. (2020). Nitrogen Retention in Mesocosm Sediments Received Rural Wastewater Associated with Microbial Community Response to Plant Species. Water, 12(11), 3035. https://doi.org/10.3390/w12113035