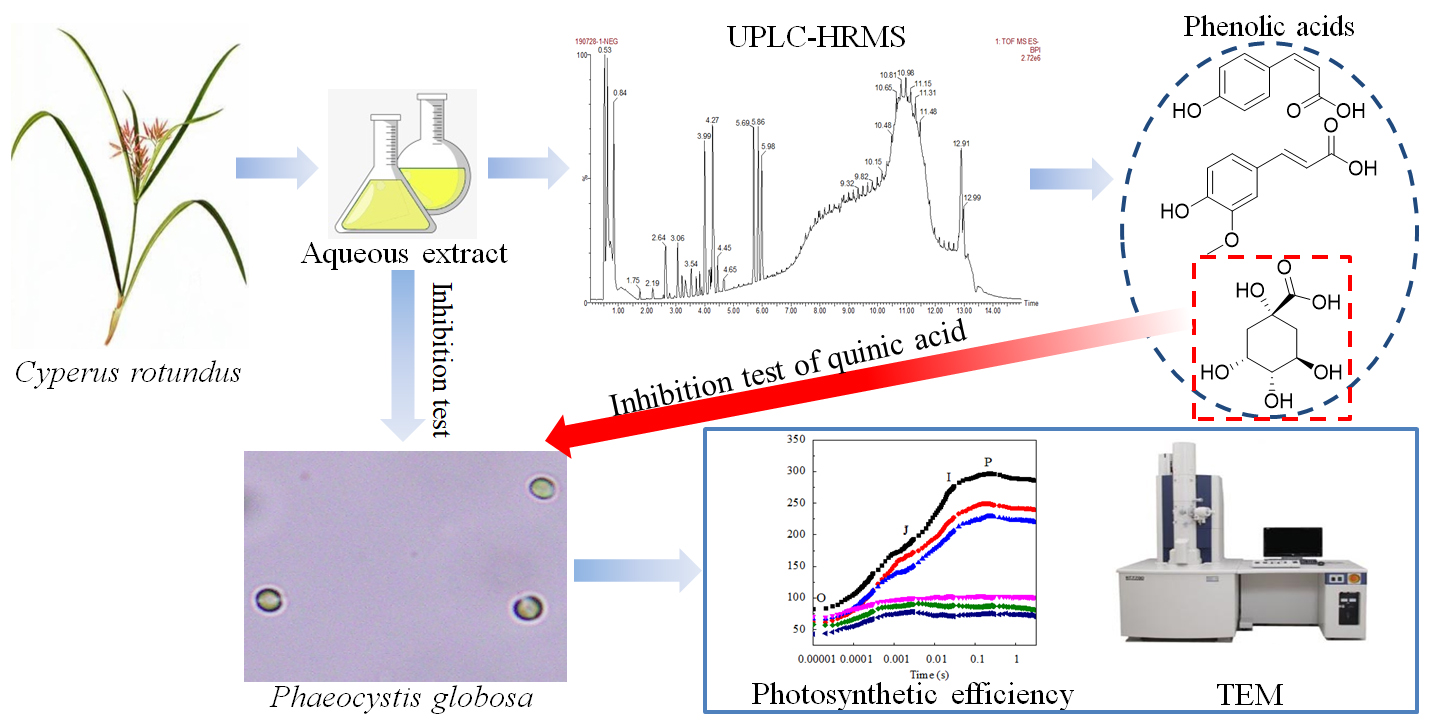
Algicidal Activity of Cyperus rotundus Aqueous Extracts Reflected by Photosynthetic Efficiency and Cell Integrity of Harmful Algae Phaeocystis globosa

 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Phaeocystis Globosa Cultures
2.2. Cyperus Rotundus Plant Sampling and Treatments
2.3. Co-Cultures of C. rotundus Extracts and P. globosa
2.4. Measurement of Photosynthetic Efficiency and Chlorophyll-a Content
2.5. TEM Analysis of the Cell Ultrastructure
2.6. Analysis of the Main Compounds of C. rotundus Extracts
2.7. Reagents and Algal Cultures
2.8. Statistical Analyses
3. Results
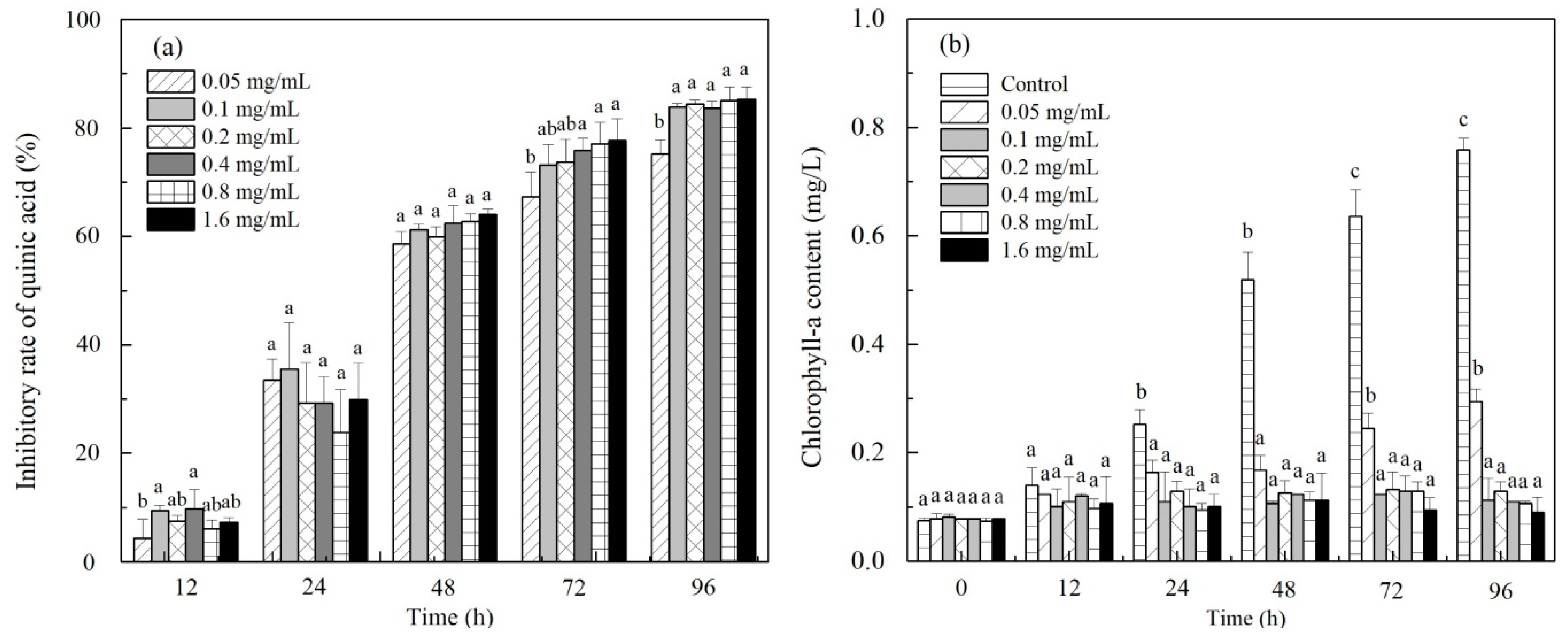
3.1. Effect of C. rotundus Extracts on Algal Growth and Chlorophyll-a Content
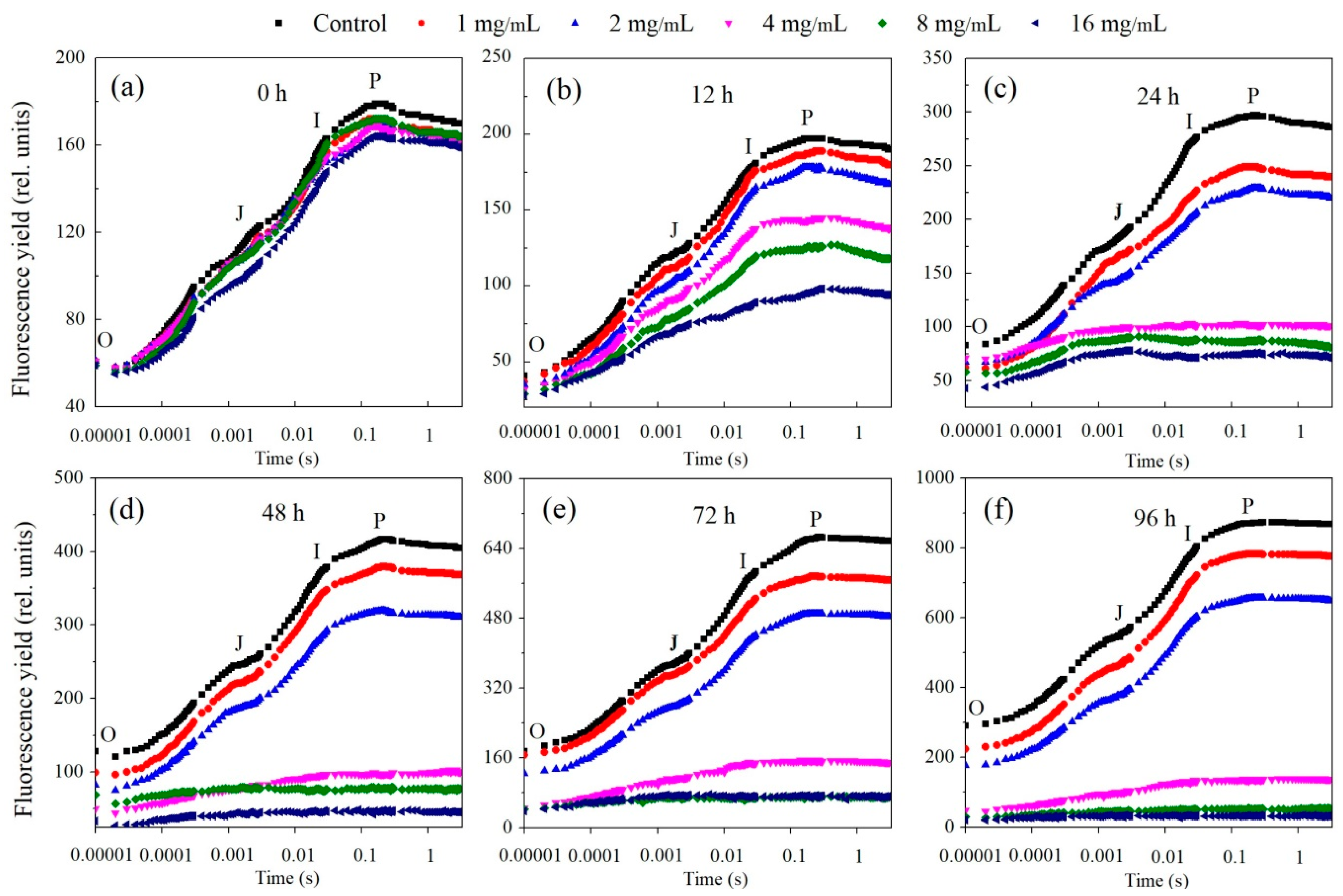
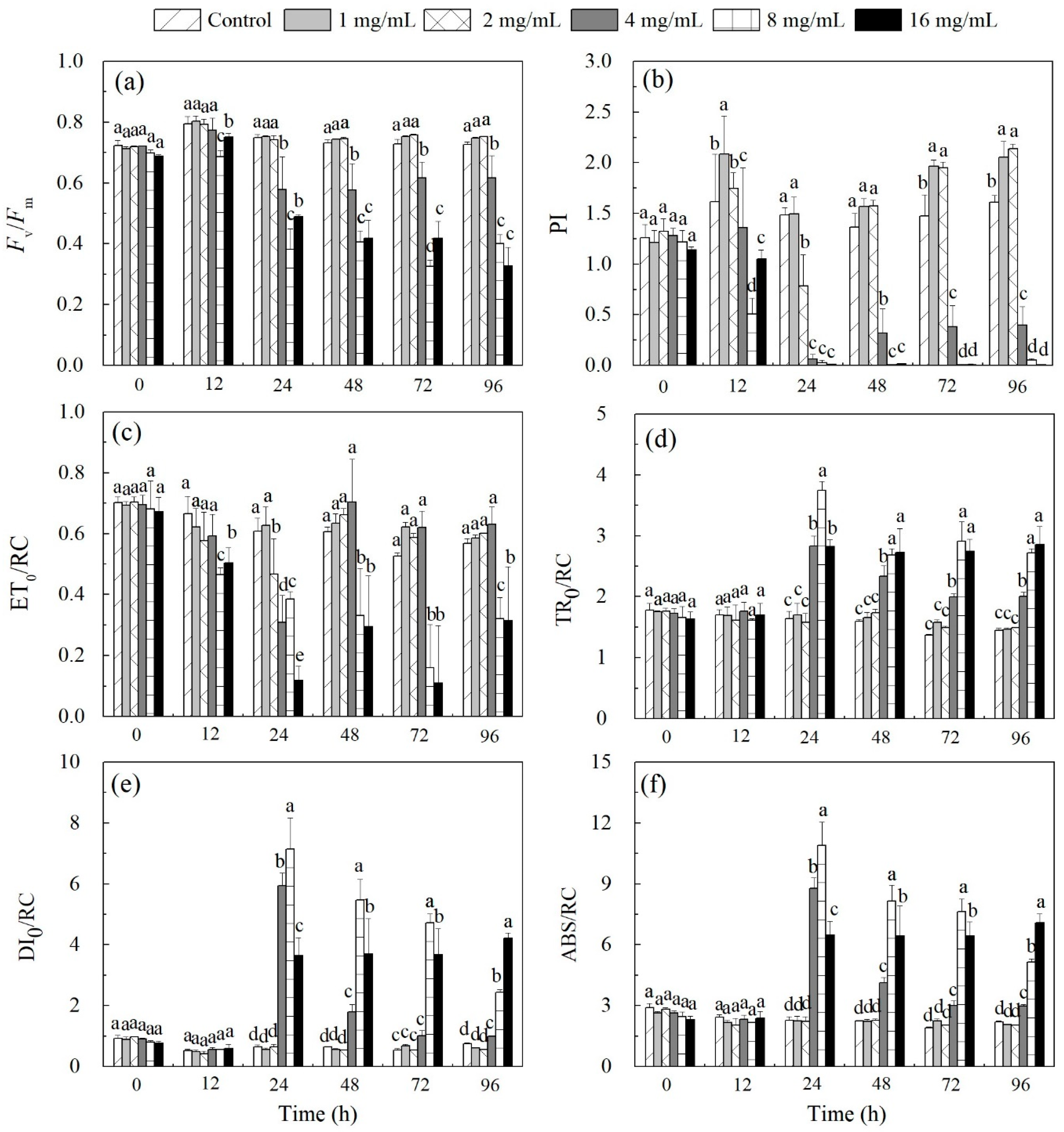
3.2. Inhibition of Algal Photosynthetic Efficiency
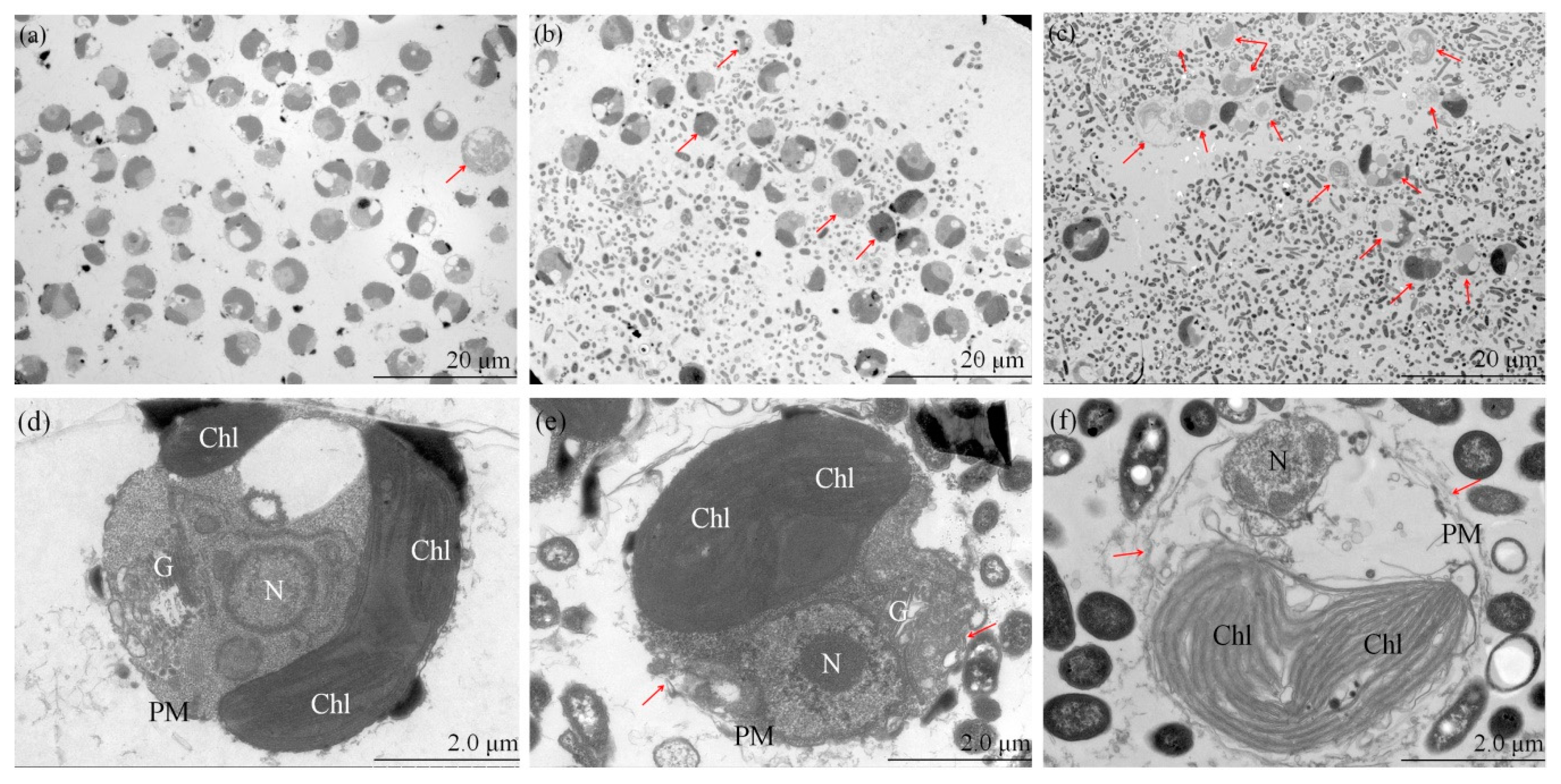
3.3. Ultrastructure of the Test Algae
3.4. The Main Compounds in C. rotundus Aqueous Extracts
3.5. Quinic Acid Test for the Algal Growth Assessment
4. Discussion
4.1. Modes of Physiological Progression
4.2. Algicidal Potential of Phenolic Acids
4.3. Bioavailability of Phenolic Compounds
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Heisler, J.; Glibert, P.; Burkholder, J.; Anderson, D.; Cochlan, W.; Dennison, W.; Gobler, C.; Dortch, Q.; Heil, C.; Humphries, E.; et al. Eutrophication and Harmful Algal Blooms: A Scientific Consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, P.J.; Piontkovski, S.; Al-Hashmi, K. Understanding how physical-biological coupling influences harmful algal blooms, low oxygen and fish kills in the Sea of Oman and the Western Arabian Sea. Mar. Pollut. Bull. 2017, 114, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Furuya, K.; Iwataki, M.; Lim, P.T.; Lu, S.; Leaw, C.-P.; Azanza, R.V.; Kim, H.-G.; Fukuyo, Y. Overview of harmful algal blooms in Asia. In Global Ecology and Oceanography of Harmful Algal Blooms; Springer: Berlin/Heidelberg, Germany, 2018; pp. 289–308. [Google Scholar]
- Schoemann, V.; Becquevort, S.; Stefels, J.; Rousseau, V.; Lancelot, C. Phaeocystis blooms in the global ocean and their controlling mechanisms: A review. J. Sea Res. 2005, 53, 43–66. [Google Scholar] [CrossRef]
- Qi, Y.; Chen, J.; Wang, Z.; Xu, N.; Wang, Y.; Shen, P.; Lu, S.; Hodgkiss, I.J. Some observations on harmful algal bloom (HAB) events along the coast of Guangdong, southern China in 1998. Hydrobiologia 2004, 512, 209–214. [Google Scholar] [CrossRef]
- Smith, W.O., Jr.; Liu, X.; Tang, K.W.; DeLizo, L.M.; Doan, N.H.; Nguyen, N.L.; Wang, X. Giantism and its role in the harmful algal bloom species Phaeocystis globosa. Deep Sea Res. Part II Top. Stud. Oceanogr. 2014, 101, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Doan, H.; Nguyen, N.L.; Nguyen, C.; Ho, V.T.; Nguyen, M.A. Plankton assemblages during the late bloom of haptophyte algae in Binh Thuan province, Southern central Vietnam, in July 2002. Collect. Mar. Res. Work. 2003, 13, 105–118. [Google Scholar]
- Rousseau, V.; Lantoine, F.; Rodriguez, F.; LeGall, F.; Chretiennot-Dinet, M.J.; Lancelot, C. Characterization of Phaeocystis globosa (Prymnesiophyceae), the blooming species in the Southern North Sea. J. Sea Res. 2013, 76, 105–113. [Google Scholar] [CrossRef]
- Hai, D.-N.; Lam, N.-N.; Dippner, J.W. Development of Phaeocystis globosa blooms in the upwelling waters of the South Central coast of Viet Nam. J. Mar. Syst. 2010, 83, 253–261. [Google Scholar] [CrossRef]
- Mars Brisbin, M.; Mitarai, S. Differential gene expression supports a resource-intensive, defensive role for colony production in the bloom-forming haptophyte, Phaeocystis globosa. J. Eukaryot. Microbiol. 2019, 66, 788–801. [Google Scholar] [CrossRef] [Green Version]
- Rauch, M.; Denis, L.; Dauvin, J.-C. The effects of Phaeocystis globosa bloom on the dynamics of the mineralization processes in intertidal permeable sediment in the Eastern English Channel (Wimereux, France). Mar. Pollut. Bull. 2008, 56, 1284–1293. [Google Scholar] [CrossRef]
- Rousseau, V.; Chrétiennot-Dinet, M.-J.; Jacobsen, A.; Verity, P.; Whipple, S. The life cycle of Phaeocystis: State of knowledge and presumptive role in ecology. Biogeochemistry 2007, 83, 29–47. [Google Scholar] [CrossRef]
- Veldhuis, M.J.; Brussaard, C.P.; Noordeloos, A.A. Living in a Phaeocystis colony: A way to be a successful algal species. Harmful Algae 2005, 4, 841–858. [Google Scholar] [CrossRef]
- Rousseau, V.; Vaulot, D.; Casotti, R.; Cariou, V.; Lenz, J.; Gunkel, J.; Baumann, M. The life cycle of Phaeocystis (Prymnesiophycaea): Evidence and hypotheses. J. Mar. Syst. 1994, 5, 23–39. [Google Scholar] [CrossRef]
- Spilmont, N.; Denis, L.; Artigas, L.F.; Caloin, F.; Courcot, L.; Créach, A.; Desroy, N.; Gevaert, F.; Hacquebart, P.; Hubas, C. Impact of the Phaeocystis globosa spring bloom on the intertidal benthic compartment in the eastern English Channel: A synthesis. Mar. Pollut. Bull. 2009, 58, 55–63. [Google Scholar] [CrossRef] [Green Version]
- Blauw, A.; Los, F.; Huisman, J.; Peperzak, L. Nuisance foam events and Phaeocystis globosa blooms in Dutch coastal waters analyzed with fuzzy logic. J. Mar. Syst. 2010, 83, 115–126. [Google Scholar] [CrossRef]
- Seuront, L.; Vincent, D.; Mitchell, J.G. Biologically induced modification of seawater viscosity in the Eastern English Channel during a Phaeocystis globosa spring bloom. J. Mar. Syst. 2006, 61, 118–133. [Google Scholar] [CrossRef]
- Tian, F.; Zhou, J.Y.; Sun, Z.W.; Cai, Z.P.; Xu, N.; An, M.; Duan, S.S. Inhibitory effects of Chinese traditional herbs and herb-modified clays on the growth of harmful algae, Phaeocystis globosa and Prorocentrum donghaiense. Harmful Algae 2014, 37, 153–159. [Google Scholar] [CrossRef]
- Zhao, M.; Xiao, H.; Sun, D.; Duan, S. Investigation of the Inhibitory Effects of Mangrove Leaves and Analysis of Their Active Components on Phaeocystis globosa during Different Stages of Leaf Age. Int. J. Environ. Res. Public Health 2018, 15, 2434. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Ge, Z.; Li, C.; Wan, F.; Xiao, X. Inhibition of harmful algae Phaeocystis globosa and Prorocentrum donghaiense by extracts of coastal invasive plant Spartina alterniflora. Sci. Total Environ. 2019, 696, 133930. [Google Scholar] [CrossRef]
- Brussaard, C.; Mari, X.; Van Bleijswijk, J.; Veldhuis, M. A mesocosm study of Phaeocystis globosa (Prymnesiophyceae) population dynamics: II. Significance for the microbial community. Harmful Algae 2005, 4, 875–893. [Google Scholar] [CrossRef]
- Sun, R.; Sun, P.F.; Zhang, J.H.; Esquivel-Elizondo, S.; Wu, Y.H. Microorganisms-based methods for harmful algal blooms control: A review. Bioresour. Technol. 2018, 248, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Jancula, D.; Marsalek, B. Critical review of actually available chemical compounds for prevention and management of cyanobacterial blooms. Chemosphere 2011, 85, 1415–1422. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Rodríguez, J.J.; Astuya-Villalón, A.; Llanos-Rivera, A.; Avello-Fontalba, V.; Ulloa-Jofré, V. A critical review on control methods for harmful algal blooms. Rev. Aquac. 2018, 11, 661–684. [Google Scholar] [CrossRef]
- Zhu, J.; Xiao, H.; Chen, Q.; Zhao, M.; Sun, D.; Duan, S. Growth Inhibition of Phaeocystis Globosa Induced by Luteolin-7-O-glucuronide from Seagrass Enhalus acoroides. Int. J. Environ. Res. Public Health 2019, 16, 2615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patino, R.; Rashel, R.H.; Rubio, A.; Longing, S. Growth-suppressing and algicidal properties of an extract from Arundo donax, an invasive riparian plant, against Prymnesium parvum, an invasive harmful alga. Harmful Algae 2018, 71, 1–9. [Google Scholar] [CrossRef]
- Chen, S.; Zheng, T.; Ye, C.; Huannixi, W.; Yakefu, Z.; Meng, Y.; Peng, X.; Tian, Z.; Wang, J.; Ma, Y.; et al. Algicidal properties of extracts from Cinnamomum camphora fresh leaves and their main compounds. Ecotoxicol. Environ. Saf. 2018, 163, 594–603. [Google Scholar] [CrossRef]
- Hussain, I.; Singh, N.; Singh, A.; Singh, H. Allelopathic potential of sesame plant leachate against Cyperus rotundus L. Ann. Agrar. Sci. 2017, 15, 141–147. [Google Scholar] [CrossRef]
- Bryson, C.T.; Reddy, K.N.; Molin, W.T. Purple nutsedge (Cyperus rotundus) population dynamics in narrow row transgenic cotton (Gossypium hirsutum) and soybean (Glycine max) rotation. Weed Technol. 2003, 17, 805–810. [Google Scholar] [CrossRef]
- Dhima, K.; Vasilakoglou, I.; Stefanou, S.; Gatsis, T.; Paschalidis, K.; Aggelopoulos, S.; Eleftherohorinos, I. Differential competitive and allelopathic ability of Cyperus rotundus on Solanum lycopersicum, Solanum melongena and Capsicum annuum. Arch. Agron. Soil Sci. 2016, 62, 1250–1263. [Google Scholar] [CrossRef]
- Hu, Q.P.; Cao, X.M.; Hao, D.L.; Zhang, L.L. Chemical Composition, Antioxidant, DNA Damage Protective, Cytotoxic and Antibacterial Activities of Cyperus rotundus Rhizomes Essential Oil against Foodborne Pathogens. Sci. Rep. 2017, 7, 45231. [Google Scholar] [CrossRef] [Green Version]
- Peerzada, A.M.; Ali, H.H.; Naeem, M.; Latif, M.; Bukhari, A.H.; Tanveer, A. Cyperus rotundus L.: Traditional uses, phytochemistry, and pharmacological activities. J. Ethnopharmacol. 2015, 174, 540–560. [Google Scholar] [CrossRef] [PubMed]
- Lawal, O.A.; Oyedeji, A.O. Chemical composition of the essential oils of Cyperus rotundus L. from South Africa. Molecules 2009, 14, 2909–2917. [Google Scholar] [CrossRef] [PubMed]
- Kilani-Jaziri, S.; Bhouri, W.; Skandrani, I.; Limem, I.; Chekir-Ghedira, L.; Ghedira, K. Phytochemical, antimicrobial, antioxidant and antigenotoxic potentials of Cyperus rotundus extracts. S. Afr. J. Bot. 2011, 77, 767–776. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Lu, S.H.; Cen, J.Y. Spatio-temporal variations of Harmful algal blooms along the coast of Guangdong, Southern China during 1980–2016. J. Oceanol. Limnol. 2019, 37, 535–551. [Google Scholar] [CrossRef]
- Wang, S.F.; Tang, D.L.; He, F.L.; Fukuyo, Y.S.; Azanza, R.V. Occurrences of harmful algal blooms (HABs) associated with ocean environments in the South China Sea. Hydrobiologia 2008, 596, 79–93. [Google Scholar] [CrossRef]
- Hoshaw, R. Methods for microscopic algae. In Handbook of Phycological Methods: Culture Methods and Growth Measurements; Cambridge University Press: Cambridge, UK, 1973; pp. 53–68. [Google Scholar]
- Xu, N.; Huang, B.Z.; Hu, Z.X.; Tang, Y.Z.; Duan, S.S.; Zhang, C.W. Effects of temperature, salinity, and irradiance on the growth of harmful algal bloom species Phaeocystis globosa Scherffel (Prymnesiophyceae) isolated from the South China Sea. Chin. J. Oceanol. Limnol. 2017, 35, 557–565. [Google Scholar] [CrossRef]
- Guillard, R.R.; Ryther, J.H. Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt, and Detonula confervacea (cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef]
- Sun, D.; He, N.; Chen, Q.; Duan, S. Effects of Lanthanum on the Photosystem II Energy Fluxes and Antioxidant System of Chlorella vulgaris and Phaeodactylum tricornutum. Int. J. Environ. Res. Public Health 2019, 16, 2242. [Google Scholar] [CrossRef] [Green Version]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, G.C., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. [Google Scholar]
- Strasser, R.J.; Tsimilli-Michael, M.; Qiang, S.; Goltsev, V. Simultaneous in vivo recording of prompt and delayed fluorescence and 820-nm reflection changes during drying and after rehydration of the resurrection plant Haberlea rhodopensis. Biochim. Biophys. Acta (BBA)-Bioenerg. 2010, 1797, 1313–1326. [Google Scholar] [CrossRef] [Green Version]
- Kula, M.; Kalaji, H.; Skoczowski, A. Culture density influence on the photosynthetic efficiency of microalgae growing under different spectral compositions of light. J. Photochem. Photobiol. B Biol. 2017, 167, 290–298. [Google Scholar] [CrossRef]
- Lin, S.J.; He, L.J.; Huang, P.S.; Han, B.P. Comparison and improvement on the extraction method for chlorophyll a in phytoplankton. Ecol. Sci. 2005, 24, 9–11. [Google Scholar]
- Zhang, J.; Li, R.; Zhang, X.; Bai, Y.; Cao, P.; Hua, P. Vehicular contribution of PAHs in size dependent road dust: A source apportionment by PCA-MLR, PMF, and Unmix receptor models. Sci. Total Environ. 2019, 649, 1314–1322. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, A.; Blondeau, D.; Lajeunesse, A.; Bley, J.; Bourdeau, N.; Desgagné-Penix, I. Phytochemical screening of quaking aspen (populus tremuloides) extracts by UPLC-QTOF-MS and evaluation of their antimicrobial activity. Molecules 2018, 23, 1739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, J.-H.; Kuang, Z.; Wang, P.; Park, B.S.; Patidar, S.K.; Han, M.-S. Ecological assessment of an algaecidal naphthoquinone derivate for the mitigation of Stephanodiscus within a mesocosm. Environ. Pollut. 2017, 229, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Joo, J.-H.; Wang, P.; Park, B.S.; Byun, J.-H.; Choi, H.J.; Kim, S.H.; Han, M.-S. Improvement of cyanobacterial-killing biologically derived substances (BDSs) using an ecologically safe and cost-effective naphthoquinone derivative. Ecotoxicol. Environ. Saf. 2017, 141, 188–198. [Google Scholar] [CrossRef]
- Shao, J.H.; Li, R.H.; Lepo, J.E.; Gu, J.D. Potential for control of harmful cyanobacterial blooms using biologically derived substances: Problems and prospects. J. Environ. Manag. 2013, 125, 149–155. [Google Scholar] [CrossRef]
- Liu, H.; Huang, J.; Yang, S.; Li, J.; Zhou, L. Chemical Composition, Algicidal, Antimicrobial, and Antioxidant Activities of the Essential Oils of Taiwania flousiana Gaussen. Molecules 2020, 25, 967. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.M.; Xiao, X.; Ghadouani, A.; Wu, J.P.; Nie, Z.Y.; Peng, C.; Xu, X.H.; Shi, J.Y. Effects of Natural Flavonoids on Photosynthetic Activity and Cell Integrity in Microcystis aeruginosa. Toxins 2015, 7, 66–80. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.Y.; Liu, B.Y.; Wang, J.; Gao, Y.N.; Wu, Z.B. Study on the mechanism of allelopathic influence on cyanobacteria and chlorophytes by submerged macrophyte (Myriophyllum spicatum) and its secretion. Aquat. Toxicol. 2010, 98, 196–203. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanisms, Regulation and Adaptation; Routledge: London, UK, 2000; pp. 445–483. [Google Scholar]
- Gauthier, A.; Joly, D.; Boisvert, S.; Carpentier, R. Period-four Modulation of Photosystem II Primary Quinone Acceptor (QA) Reduction/Oxidation Kinetics in Thylakoid Membranes. Photochem. Photobiol. 2010, 86, 1064–1070. [Google Scholar] [CrossRef]
- Singh-Tomar, R.; Jajoo, A. Alteration in PS II heterogeneity under the influence of polycyclic aromatic hydrocarbon (fluoranthene) in wheat leaves (Triticum aestivum). Plant Sci. 2013, 209, 58–63. [Google Scholar] [PubMed]
- Ye, C.; Liao, H.; Yang, Y. Allelopathic inhibition of photosynthesis in the red tide-causing marine alga, Scrippsiella trochoidea (Pyrrophyta), by the dried macroalga, Gracilaria lemaneiformis (Rhodophyta). J. Sea Res. 2014, 90, 10–15. [Google Scholar]
- Gomes, M.T.G.; da Luz, A.C.; dos Santos, M.R.; Batitucci, M.d.C.P.; Silva, D.M.; Falqueto, A.R. Drought tolerance of passion fruit plants assessed by the OJIP chlorophyll a fluorescence transient. Sci. Hortic. 2012, 142, 49–56. [Google Scholar]
- Van Heerden, P.; Swanepoel, J.; Krüger, G. Modulation of photosynthesis by drought in two desert scrub species exhibiting C3-mode CO2 assimilation. Environ. Exp. Bot. 2007, 61, 124–136. [Google Scholar]
- Oukarroum, A. Change in photosystem II photochemistry during algal growth phases of Chlorella vulgaris and Scenedesmus obliquus. Curr. Microbiol. 2016, 72, 692–699. [Google Scholar] [PubMed]
- Li, X.; Cai, J.; Liu, F.; Zhou, Q.; Dai, T.; Cao, W.; Jiang, D. Wheat plants exposed to winter warming are more susceptible to low temperature stress in the spring. Plant Growth Regul. 2015, 77, 11–19. [Google Scholar]
- Yang, C.-M.; Lee, C.-N.; Chou, C.-H. Effects of three allelopathic phenolics on chlorophyll accumulation of rice (Oryza sativa) seedlings: I. Inhibition of supply-orientation. Bot. Bull. Acad. Sin. 2002, 43, 299–304. [Google Scholar]
- Jin, P.; Wang, H.; Huang, W.; Liu, W.; Fan, Y.; Miao, W. The allelopathic effect and safety evaluation of 3, 4-Dihydroxybenzalacetone on Microcystis aeruginosa. Pestic. Biochem. Physiol. 2018, 147, 145–152. [Google Scholar]
- Campos, F.; Couto, J.; Figueiredo, A.; Tóth, I.; Rangel, A.O.; Hogg, T. Cell membrane damage induced by phenolic acids on wine lactic acid bacteria. Int. J. Food Microbiol. 2009, 135, 144–151. [Google Scholar]
- Wagner, H.; Efferth, T. Introduction: Novel hybrid combinations containing synthetic or antibiotic drugs with plant-derived phenolic or terpenoid compounds. Phytomedicine 2017, 37, 1–3. [Google Scholar]
- Wang, R.; Hua, M.; Yu, Y.; Zhang, M.; Xian, Q.M.; Yin, D.Q. Evaluating the effects of allelochemical ferulic acid on Microcystis aeruginosa by pulse-amplitude-modulated (PAM) fluorometry and flow cytometry. Chemosphere 2016, 147, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.-T.; Zheng, C.-Y.; Hu, W.; Xu, W.-W.; Wang, H.-F. The allelopathy and allelopathic mechanism of phenolic acids on toxic Microcystis aeruginosa. J. Appl. Phycol. 2010, 22, 71–77. [Google Scholar] [CrossRef]
- Gao, Y.-N.; Liu, B.-Y.; Xu, D.; Zhou, Q.-H.; Hu, C.-Y.; Ge, F.-J.; Zhang, L.-P.; Wu, Z.-B. Phenolic Compounds Exuded from Two Submerged Freshwater Macrophytes and Their Allelopathic Effects on Microcystis aeruginosa. Pol. J. Environ. Stud. 2011, 20, 1153–1159. [Google Scholar]
- Hua, Q.; Liu, Y.-G.; Yan, Z.-L.; Zeng, G.-M.; Liu, S.-B.; Wang, W.-J.; Tan, X.-F.; Deng, J.-Q.; Tang, X.; Wang, Q.-P. Allelopathic effect of the rice straw aqueous extract on the growth of Microcystis aeruginosa. Ecotoxicol. Environ. Saf. 2018, 148, 953–959. [Google Scholar] [CrossRef]
- Zhang, C.; Ling, F.; Yi, Y.L.; Zhang, H.Y.; Wang, G.X. Algicidal activity and potential mechanisms of ginkgolic acids isolated from Ginkgo biloba exocarp on Microcystis aeruginosa. J. Appl. Phycol. 2014, 26, 323–332. [Google Scholar] [CrossRef]
- Shao, J.; Wu, Z.; Yu, G.; Peng, X.; Li, R. Allelopathic mechanism of pyrogallol to Microcystis aeruginosa PCC7806 (Cyanobacteria): From views of gene expression and antioxidant system. Chemosphere 2009, 75, 924–928. [Google Scholar] [CrossRef]
- Chen, Q.; Zhu, B.; Sun, D.; Liu, W.; Sun, X.; Duan, S. The effect of protocatechuic acid on the phycosphere in harmful algal bloom species Scrippsiella trochoidea. Aquat. Toxicol. 2020, 227, 105591. [Google Scholar] [CrossRef]
- Ni, L.X.; Acharya, K.; Mao, X.Y.; Li, S.Y. Isolation and identification of an anti-algal compound from Artemisia annua and mechanisms of inhibitory effect on algae. Chemosphere 2012, 88, 1051–1057. [Google Scholar] [CrossRef]
- Zhang, C.; Yi, Y.-L.; Hao, K.; Liu, G.-L.; Wang, G.-X. Algicidal activity of Salvia miltiorrhiza Bung on Microcystis aeruginosa—Towards identification of algicidal substance and determination of inhibition mechanism. Chemosphere 2013, 93, 997–1004. [Google Scholar] [CrossRef]
- Sun, Y.-Y.; Xing, J.-Z.; Zhang, J.-S.; Zhou, W.-J.; Pu, Y.-F. Sesquiterpenoids with antialgal activity against the common red tide microalgae from marine macroalga Porphyra yezoensis. Environ. Sci. Pollut. Res. Int. 2018, 25, 7844–7859. [Google Scholar] [CrossRef]
- Bartwal, A.; Mall, R.; Lohani, P.; Guru, S.K.; Arora, S. Role of Secondary Metabolites and Brassinosteroids in Plant Defense Against Environmental Stresses. J. Plant Growth Regul. 2013, 32, 216–232. [Google Scholar] [CrossRef]
- Macías, F.A.; Galindo, J.L.; García-Díaz, M.D.; Galindo, J.C. Allelopathic agents from aquatic ecosystems: Potential biopesticides models. Phytochem. Rev. 2008, 7, 155–178. [Google Scholar] [CrossRef]
- Turek, C.; Stintzing, F.C. Stability of essential oils: A review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 40–53. [Google Scholar] [CrossRef]
- Xiao, X.; Li, C.; Huang, H.; Lee, Y.P. Inhibition effect of natural flavonoids on red tide alga Phaeocystis globosa and its quantitative structure-activity relationship. Environ. Sci. Pollut. Res. Int. 2019, 26, 23763–23776. [Google Scholar] [CrossRef]
- Falcone Ferreyra, M.L.; Rius, S.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koes, R.; Verweij, W.; Quattrocchio, F. Flavonoids: A colorful model for the regulation and evolution of biochemical pathways. Trends Plant Sci. 2005, 10, 236–242. [Google Scholar] [CrossRef]
- Tommasini, S.; Raneri, D.; Ficarra, R.; Calabrò, M.L.; Stancanelli, R.; Ficarra, P. Improvement in solubility and dissolution rate of flavonoids by complexation with β-cyclodextrin. J. Pharm. Biomed. Anal. 2004, 35, 379–387. [Google Scholar] [CrossRef]
- Laue, P.; Bährs, H.; Chakrabarti, S.; Steinberg, C.E. Natural xenobiotics to prevent cyanobacterial and algal growth in freshwater: Contrasting efficacy of tannic acid, gallic acid, and gramine. Chemosphere 2014, 104, 212–220. [Google Scholar] [CrossRef]
- Chung, K.-T.; Wei, C.-I.; Johnson, M.G. Are tannins a double-edged sword in biology and health? Trends Food Sci. Technol. 1998, 9, 168–175. [Google Scholar] [CrossRef]
- Brglez Mojzer, E.; Knez Hrnčič, M.; Škerget, M.; Knez, Ž.; Bren, U. Polyphenols: Extraction methods, antioxidative action, bioavailability and anticarcinogenic effects. Molecules 2016, 21, 901. [Google Scholar] [CrossRef]
- Khadem, S.; Marles, R.J. Monocyclic phenolic acids; hydroxy-and polyhydroxybenzoic acids: Occurrence and recent bioactivity studies. Molecules 2010, 15, 7985–8005. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-H.; Wang, Q.; Ruan, X.; Pan, C.-D.; Jiang, D.-A. Phenolics and plant allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olofsdotter, M.; Rebulanan, M.; Madrid, A.; Dali, W.; Navarez, D.; Olk, D.C. Why phenolic acids are unlikely primary allelochemicals in rice. J. Chem. Ecol. 2002, 28, 229–242. [Google Scholar] [CrossRef]
- Nakai, S.; Inoue, Y.; Hosomi, M. Algal growth inhibition effects and inducement modes by plant-producing phenols. Water Res. 2001, 35, 1855–1859. [Google Scholar] [CrossRef]
- Yu, X.-B.; Hao, K.; Ling, F.; Wang, G.-X. Aquatic environmental safety assessment and inhibition mechanism of chemicals for targeting Microcystis aeruginosa. Ecotoxicology 2014, 23, 1638–1647. [Google Scholar] [CrossRef] [PubMed]
- Lindner, A.V.; Pleissner, D. Utilization of phenolic compounds by microalgae. Algal Res. 2019, 42, 101602. [Google Scholar] [CrossRef]
- Surkatti, R.; Al-Zuhair, S. Microalgae cultivation for phenolic compounds removal. Environ. Sci. Pollut. Res. Int. 2018, 25, 33936–33956. [Google Scholar] [CrossRef]
- Ghasemi, Y.; Rasoul-Amini, S.; Fotooh-Abadi, E. The biotransformation, biodegradation, and bioremediation of organic compounds by microalgae 1. J. Phycol. 2011, 47, 969–980. [Google Scholar] [CrossRef]
- Maza-Márquez, P.; Martinez-Toledo, M.V.; Fenice, M.; Andrade, L.; Lasserrot, A.; Gonzalez-Lopez, J. Biotreatment of olive washing wastewater by a selected microalgal-bacterial consortium. Int. Biodeterior. Biodegrad. 2014, 88, 69–76. [Google Scholar] [CrossRef]
- Escapa, C.; Coimbra, R.; Paniagua, S.; García, A.; Otero, M. Paracetamol and salicylic acid removal from contaminated water by microalgae. J. Environ. Manag. 2017, 203, 799–806. [Google Scholar] [CrossRef]
- Haritash, A.; Kaushik, C. Biodegradation aspects of polycyclic aromatic hydrocarbons (PAHs): A review. J. Hazard. Mater. 2009, 169, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Xiao, X.; Lin, F.; Grossart, H.-P.; Nie, Z.; Sun, L.; Xu, C.; Shi, J. Continuous-release beads of natural allelochemicals for the long-term control of cyanobacterial growth: Preparation, release dynamics and inhibitory effects. Water Res. 2016, 95, 113–123. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Definition |
|---|---|
| Fv/Fm | Maximum photochemical efficiency of PSII |
| ABS/RC | Number of QA reducing reaction centers per PSII antenna chlorophyll |
| TR0/RC | Maximum trapped exaction flux per PSII |
| ET0/RC | Electron transport from QA to QB per PSII reaction center |
| DI0/RC | Heat dissipation per PSII reaction center |
| PI | Performance index |
| No. | Rt (min) | Main Compounds | Formula | Exact Mass (m/z) | Peak Area | |
|---|---|---|---|---|---|---|
| [M+H]+ | [M−H]− | |||||
| 1 | 0.62 | Quinic acid | C7H12O6 | / | 191.0556 | 138,266 |
| 2 | 2.19 | 2-hydroxy-2-(2-((4-hydroxy-3-(hydroxymethyl)but-2-en-1-yl)oxy)-2-oxoethyl)succinic acid | C11H16O9 | / | 291.0727 | 24,200 |
| 3 | 3.06 | 3-O-trans-coumaroylquinic acid | C16H16O8 | / | 337.0910 | 22,574 |
| 4 | 3.20 | Quinic acid derivative | C13H24O9 | / | 323.1328 | 31,057 |
| 5 | 3.32 | 3-O-feruloylquinic acid | C16H16O8 | / | 367.1045 | 11,735 |
| 6 | 3.54 | 4-O-trans-coumaroylquinic acid | C16H16O8 | / | 337.0910 | 13,202 |
| 7 | 3.70 | 4-(1-carboxy-2-(3,4-dihydroxyphenyl)ethoxy)-2-hydroxy-4-oxobutanoic acid | C13H14O9 | / | 313.0544 | 17,665 |
| 8 | 3.90 | 5-O-trans-coumaroylquinic acid | C16H16O8 | / | 337.0910 | 2142 |
| 9 | 3.99 | trans-p-coumaric acid | C9H8O3 | 165.0543 | 163.0398 | 109,235 |
| 10 | 4.02 | cis-p- coumaric acid | C9H8O3 | 165.0543 | 163.0398 | 2969 |
| 11 | 4.22 | Ferulic Acid | C10H10O4 | 195.0647 | 193.0489 | 37,364 |
| 12 | 4.27 | N-acetyl-5-carboxytryptophan | C14H14N2O5 | 291.0976 | 289.0808 | 2253 |
| 13 | 4.80 | 5,6,7,8-tetrahydroquinazoline-2,4(1H,3H)-dione | C8H10N2O3 | 183.0777 | / | 77,655 |
| 14 | 6.36 | 2-(4-(methoxymethyl)phenyl)propan-2-ol | C11H16O2 | 181.1224 | / | 47,605 |
| 15 | 6.92 | Toddanone | C16H18O5 | 291.1220 | / | 88,422 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lan, Y.; Chen, Q.; Gou, T.; Sun, K.; Zhang, J.; Sun, D.; Duan, S. Algicidal Activity of Cyperus rotundus Aqueous Extracts Reflected by Photosynthetic Efficiency and Cell Integrity of Harmful Algae Phaeocystis globosa. Water 2020, 12, 3256. https://doi.org/10.3390/w12113256
Lan Y, Chen Q, Gou T, Sun K, Zhang J, Sun D, Duan S. Algicidal Activity of Cyperus rotundus Aqueous Extracts Reflected by Photosynthetic Efficiency and Cell Integrity of Harmful Algae Phaeocystis globosa. Water. 2020; 12(11):3256. https://doi.org/10.3390/w12113256
Chicago/Turabian StyleLan, Yu, Qi Chen, Ting Gou, Kaifeng Sun, Jin Zhang, Dong Sun, and Shunshan Duan. 2020. "Algicidal Activity of Cyperus rotundus Aqueous Extracts Reflected by Photosynthetic Efficiency and Cell Integrity of Harmful Algae Phaeocystis globosa" Water 12, no. 11: 3256. https://doi.org/10.3390/w12113256
APA StyleLan, Y., Chen, Q., Gou, T., Sun, K., Zhang, J., Sun, D., & Duan, S. (2020). Algicidal Activity of Cyperus rotundus Aqueous Extracts Reflected by Photosynthetic Efficiency and Cell Integrity of Harmful Algae Phaeocystis globosa. Water, 12(11), 3256. https://doi.org/10.3390/w12113256