When Green Infrastructure Turns Grey: Plant Water Stress as a Consequence of Overdesign in a Tree Trench System


Abstract
:1. Introduction
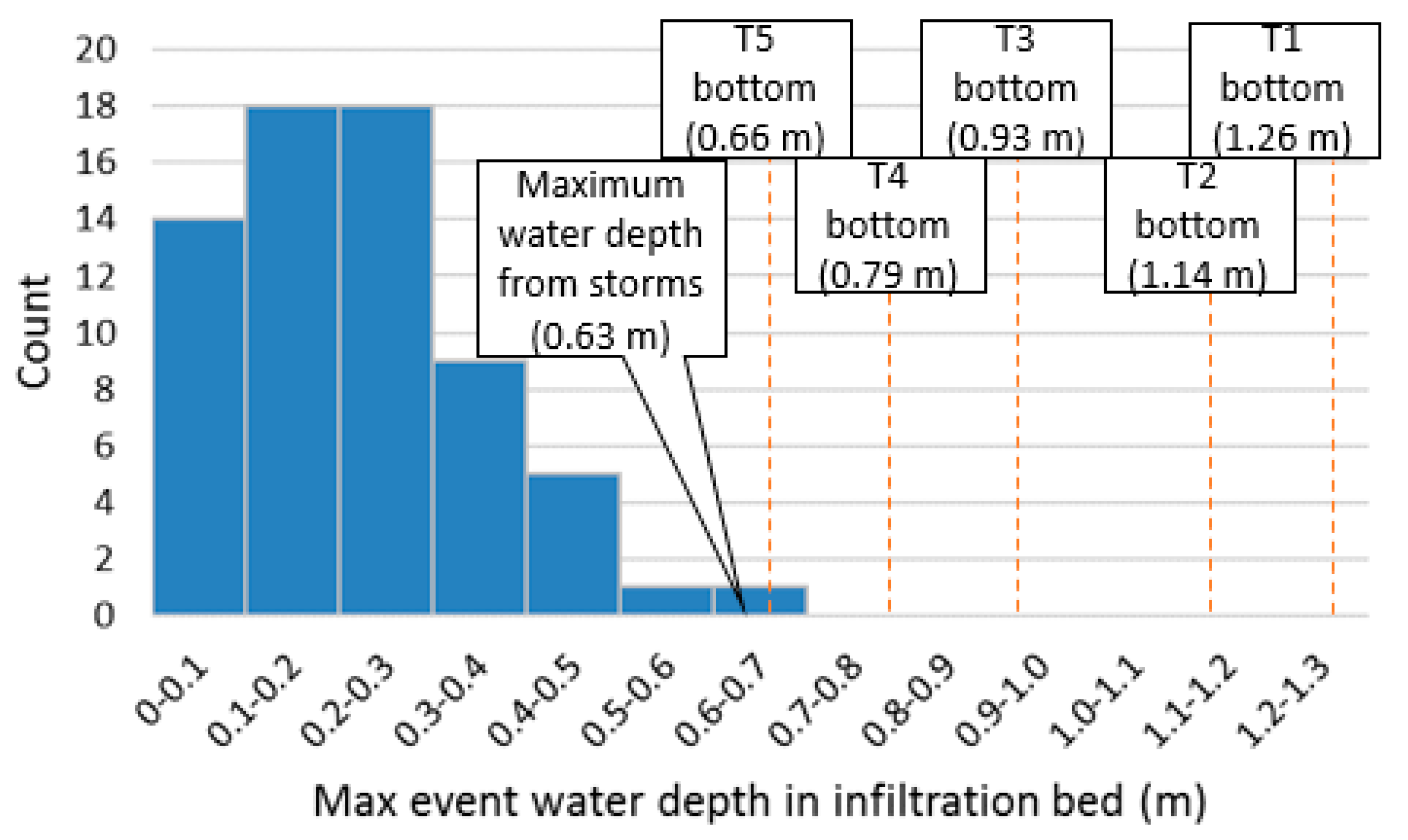
- Confirm with hydrological data that the tree trench was overdesigned. This was done by determining the frequency at which stormwater in the trench intercepted soil pits.
- Characterize the response of tree water relations to varying soil moisture conditions and determine how this response differed between species. This was done by evaluating how leaf water potential and stomatal conductance changed for two species in response to variation in soil moisture conditions throughout a growing season.
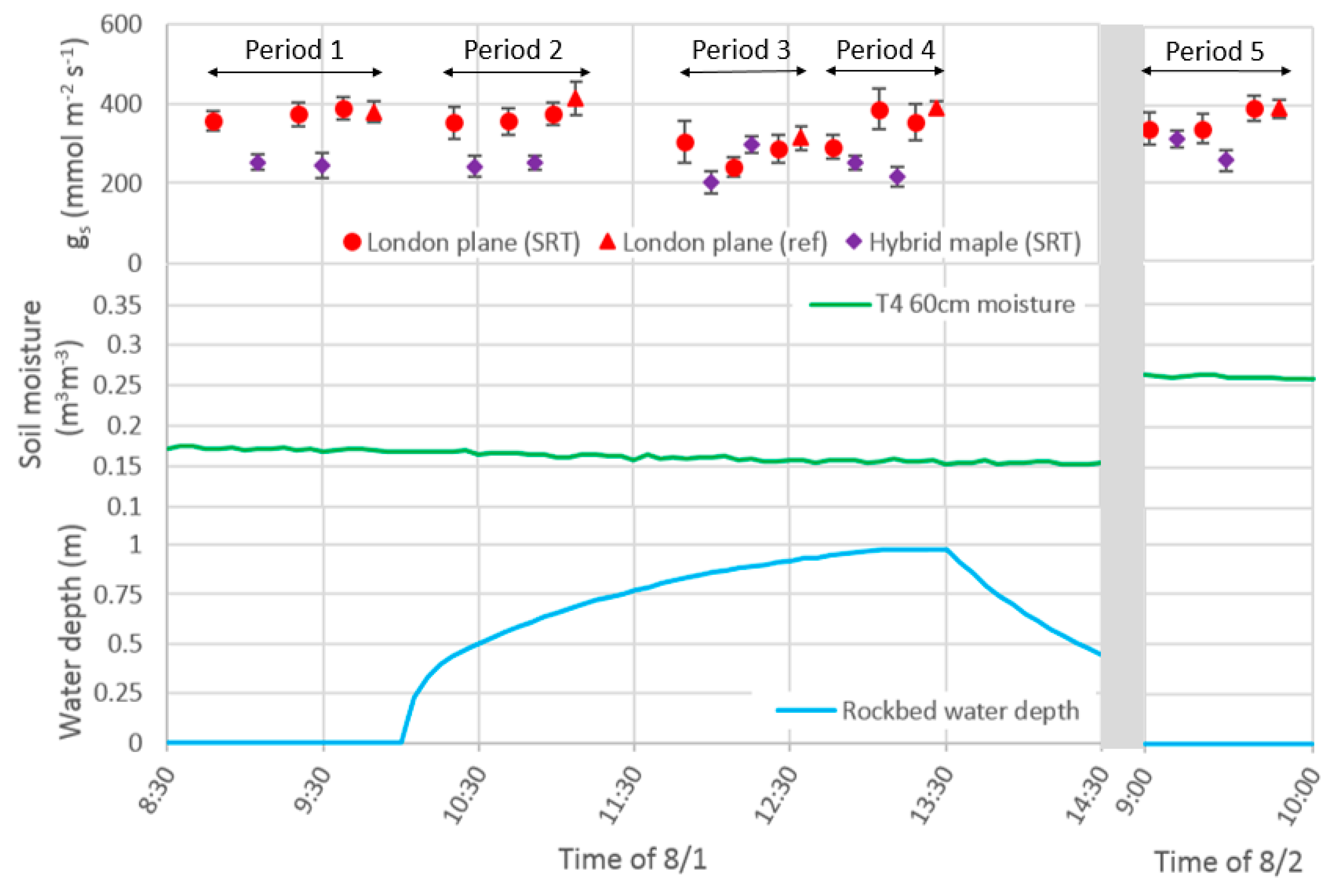
- Determine how soil moisture levels and tree water use respond to an extremely large storm event (i.e., the best-case scenario with respect to stormwater delivery to soil pits if the system is overdesigned). To achieve this, a simulated runoff test was conducted in the focal system simultaneous with repeated measurements of stomatal conductance.
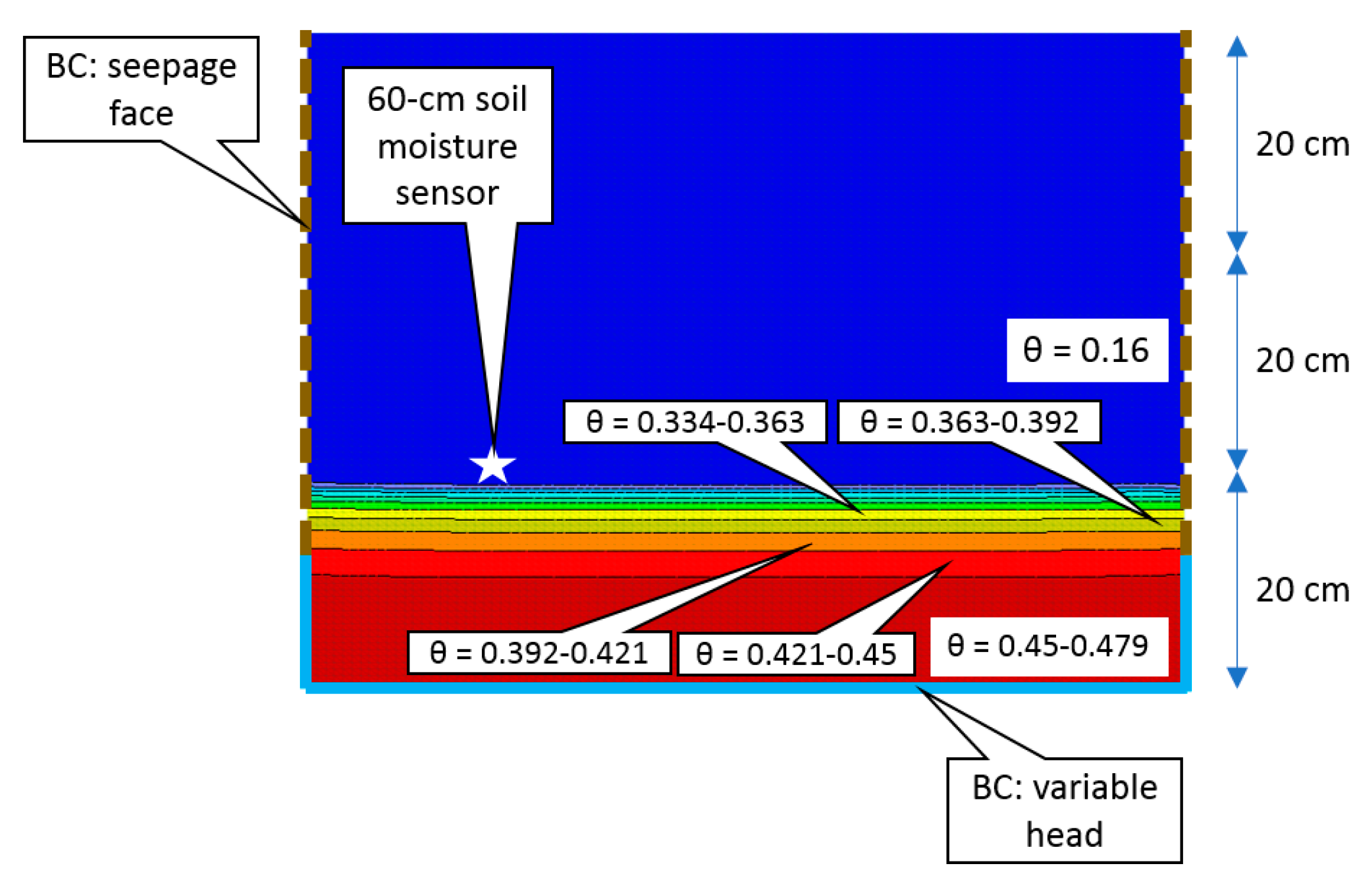
- Evaluate how the composition of media in the soil pits influences the soil moisture response to wetting by the infiltration bed. This was done using a 2D unsaturated zone hydrologic model of a soil pit that was parameterized with empirical measurements of the soil’s hydraulic properties and the time series of water inputs and water levels in the infiltration bed during the simulated runoff test.
2. Materials and Methods
2.1. Site Description
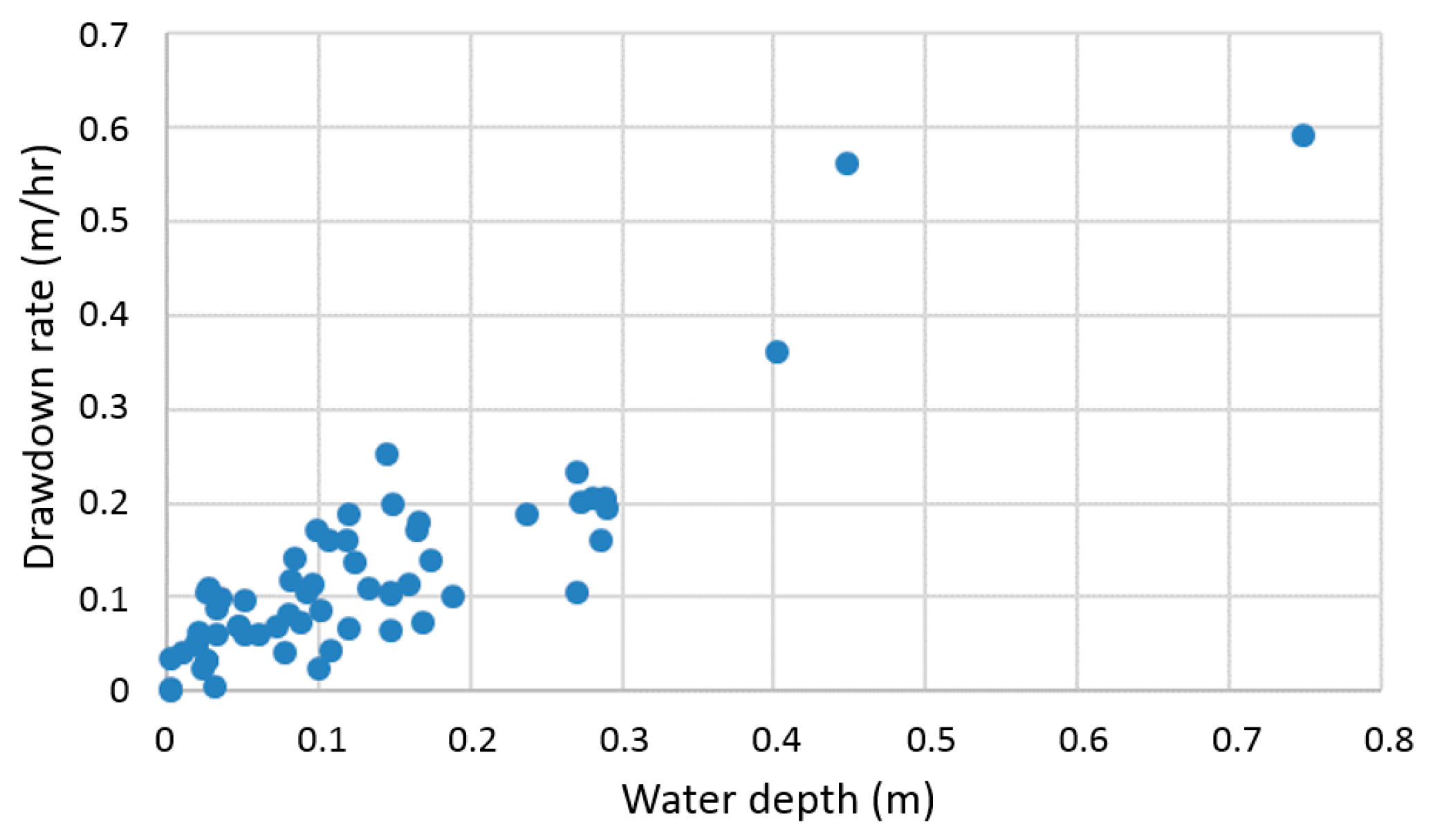
2.2. Hydrological Characterization
2.3. Plant Water Stress
2.4. Simulated Runoff Test
2.5. Hydrological Modeling
3. Results
3.1. Hydrological Characterization
3.2. Plant Water Stress
3.3. Simulated Runoff Test
3.4. Hydrological Modeling
4. Discussion
4.1. Overdesign & Plant Water Stress
4.2. Implications for Design
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Botting, J.; Bellette, K. Stormwater Pollution Prevention—Code of Practice for Local, State, and Federal Government; Environment Protection Authority: Adelaide, Australia, 1998. [Google Scholar]
- Roe, M.; Mell, I. Negotiating Value and Priorities: Evaluating the Demands of Green Infrastructure Development. J. Environ. Plan. Manag. 2012, 56, 650–673. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency. National Pollutant Discharge Elimination System (NPDES). Available online: https://www.epa.gov/npdes (accessed on 17 May 2019).
- Tu, M.-C.; Smith, P. Modeling Pollutant Buildup and Washoff Parameters for SWMM Based on Land Use in a Semiarid Urban Watershed. Water Air Soil Pollut. 2018, 229, 121. [Google Scholar] [CrossRef]
- Ahiablame, L.M.; Engel, B.A.; Chaubey, I. Effectiveness of Low Impact Development Practices: Literature Review and Suggestions for Future Research. Water Air Soil Pollut. 2012, 223, 4253–4273. [Google Scholar] [CrossRef]
- Buccola, N.; Spolek, G. A Pilot-scale Evaluation of Greenroof Runoff Retention, Detention, and Quality. Water Air Soil Pollut. 2011, 216, 83–92. [Google Scholar] [CrossRef]
- Lucas, W.C.; Greenway, M. Hydraulic Response and Nitrogen Retention in Bioretention Mesocosms with Regulated Outlets: Part I—Hydraulic Response. Water Environ. Res. 2011, 83, 692–702. [Google Scholar] [CrossRef]
- Yang, B.; Li, S. Green Infrastructure Design for Stormwater Runoff and Water Quality: Empirical Evidence from Large Watershed-scale Community Developments. Water 2013, 5, 2038–2057. [Google Scholar] [CrossRef]
- Tu, M.-C.; Traver, R.G. Water Table Fluctuation from Green Infrastructure Sidewalk Planters in Philadelphia, Pennsylvania. J. Irrig. Drain. Eng. 2019, 145, 05018008. [Google Scholar] [CrossRef]
- Rejskova, A.; Cizkova, H.; Brom, J. Transpiration, Evapotranspiration and Energy Fluxes in a Temperate Wetland Dominated by Phalaris arundinacea under Hot Summer Conditions. Ecohydrology 2012, 5, 19–27. [Google Scholar] [CrossRef]
- DiGiovanni, K.; Montalto, F.; Gaffin, S. Applicability of Classical Predictive Equations for the Estimation of Evapotranspiration from Urban Green Spaces: Green Roof Results. J. Hydrol. Eng. 2013, 18, 99–107. [Google Scholar] [CrossRef]
- Wadzuk, B.M.; Schneider, D.; Feller, M. Evapotranspiration from a Green-roof Storm-water Control Measure. J. Irrig. Drain. Eng. 2013, 139, 995–1003. [Google Scholar] [CrossRef]
- Wadzuk, B.M.; Hickman, J.M.; Traver, R.G. Understanding the Role of Evapotranspiration in Bioretention: Mesocosm Study. J. Sustain. Water Built Environ. 2015, 1, 04014002. [Google Scholar] [CrossRef]
- Zaremba, G.J.; Traver, R.G.; Wadzuk, B.M. Impact of Drainage on Green Roof Evapotranspiration. J. Irrig. Drain Eng. 2016, 142, 04016022. [Google Scholar] [CrossRef]
- Hess, A.; Wadzuk, B.; Welker, A. Evapotranspiration in Rain Gardens Using Weighing Lysimeters. J. Irrig. Drain Eng. 2017, 143, 04017004. [Google Scholar] [CrossRef]
- Wang, Y.; Bakker, F.; de Groot, R. Effect of Ecosystem Services Provided by Urban Green Infrastructure on Indoor Environment: A Literature Review. Build. Environ. 2014, 77, 88–100. [Google Scholar] [CrossRef]
- Vrban, S.; Wang, Y.; McBean, E.A. Evaluation of Stormwater Infrastructure Design Storms Developed Using Partial Duration and Annual Maximum Series Models. J. Hydrol. Eng. 2018, 23, 04018051. [Google Scholar] [CrossRef]
- Brown, R.A.; O’Connor, T.P.; Borst, M. Divergent Vegetation Growth Patterns Relative to Bioinfiltration Unit Size and Plant Placement. J. Sustain. Water Built Environ. 2015, 1, 04015001. [Google Scholar] [CrossRef]
- Baldocchi, D.; Xu, L.; Kiang, N. How Plant Functional-type, Weather, Seasonal Drought, and Soil Physical Properties Alter Water and Energy Fluxes of an Oak–grass Savanna and an Annual Grassland. Agric. For. Meteorol. 2004, 123, 13–39. [Google Scholar] [CrossRef] [Green Version]
- Reece, C.F.; Riha, S.J. Role of Root Systems of Eastern Larch and White Spruce in Response to Flooding. Plant Cell Environ. 1991, 14, 229–234. [Google Scholar] [CrossRef]
- Bouda, M.; Brodersen, C.; Saiers, J. Whole Root System Water Conductance Responds to Both Axial and Radial Traits and Network Topology over Natural Range of Trait Variation. J. Theor. Biol. 2018, 456, 49–61. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin III, F.S.; Pons, T.L. Plant Physiological Ecology; Springer: New York, NY, USA, 1998. [Google Scholar]
- Flexas, J.; Medrano, H. Drought-inhibition of Photosynthesis in C3 Plants: Stomatal and Non-stomatal Limitations Revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Tu, M.-C.; Traver, R.G. Clogging Impacts on Distribution Pipe Delivery of Street Runoff to an Infiltration Bed. Water 2018, 10, 1045. [Google Scholar] [CrossRef] [Green Version]
- Caplan, J.S.; Galanti, R.C.; Olshevski, S.; Eisenman, S.W. Water Relations of Street Trees in Green Infrastructure Tree Trench Systems. Urban For. Urban Green. 2019, 41, 170–178. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B. LME4: Linear Mixed-effects Models Using Eigen and S4. Available online: https://cran.r-project.org/web/packages/lme4/index.html (accessed on 8 January 2019).
- R Foundation for Statistical Computing. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 10 January 2020).
- Gelman, A. Scaling Regression Inputs by Dividing by Two Standard Deviations. Stat. Med. 2008, 27, 2865–2873. [Google Scholar] [CrossRef] [PubMed]
- Grueber, C.E.; Nakagawa, S.; Laws, R.J. Multimodel Inference in Ecology and Evolution: Challenges and Solutions. J. Evol. Biol. 2011, 24, 699–711. [Google Scholar] [CrossRef] [PubMed]
- PC-PROGRESS. HYDRUS 2D/3D for Windows. Available online: https://www.pc-progress.com/en/Default.aspx?hydrus-3d (accessed on 10 January 2020).
- Ebrahimian, H.; Noory, H. Modeling Paddy Field Subsurface Drainage Using HYDRUS-2D. Paddy Water Environ. 2014, 13, 477–485. [Google Scholar] [CrossRef]
- van Genuchten, M.T. A Closed-form Equation for Predicting the Hydraulic Conductivity of Unsaturated Soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef] [Green Version]
- Veylon, G.; Stoltz, G.; Meriaux, P. Performance of Geotextile Filters after 18 Years’ Service in Drainage Trenches. Geotext. Geomembr. 2016, 44, 515–533. [Google Scholar] [CrossRef]
- Campbell, R.J.; Marini, R.P.; Birch, J.B. Canopy Position Affects Light Response Curves for Gas Exchange Characteristics of Apple Spur Leaves. J. Am. Soc. Hort. Sci. 1992, 117, 467–472. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-J.; Meinzer, F.C.; Qi, J.-H. Midday Stomatal Conductance Is More Related to Stem rather than Leaf Water Status in Subtropical Deciduous and Evergreen Broadleaf Trees. Plant Cell Environ. 2013, 36, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Tolk, J.A. Plant Available Soil Water. Encyclopedia of Water Science; Marcel-Dekker: New York, NY, USA, 2003; pp. 669–672. [Google Scholar]
- Varone, L.; Ribas-carbo, M.; Cardona, C. Stomatal and Non-stomatal Limitations to Photosynthesis in Seedlings and Saplings of Mediterranean Species Pre-conditioned and Aged in Nurseries: Different Response to Water Stress. Environ. Exp. Bot. 2012, 75, 235–247. [Google Scholar] [CrossRef]
- Biswell, H.H. Effects of Environment upon the Root Habits of Certain Deciduous Forest Trees. Bot. Gaz. 1935, 96, 676–708. [Google Scholar] [CrossRef]
- Sprackling, J.A.; Read, R.A. Tree Root Systems in Eastern Nebraska. Neb. Conserv. Bull. 1979, 37, 1–73. [Google Scholar]
- Schenk, H.J.; Jackson, R.B. Rooting Depths, Lateral Root Spreads and Below-ground/Above-ground Allometries of Plants in Water-limited Ecosystems. J. Ecol. 2002, 90, 480–494. [Google Scholar] [CrossRef] [Green Version]
- Bühler, O.; Kristoffersen, P.; Larsen, S.U. Growth of Street Trees in Copenhagen with Emphasis on the Effect of Different Establishment Concepts. Arboric. Urban For. 2007, 33, 330–337. [Google Scholar]
- Todorova, A.; Asakawa, S.; Aikoh, T. Preferences for and Attitudes towards Street Flowers and Trees in Sapporo, Japan. Landsc. Urban Plan. 2004, 69, 403–416. [Google Scholar] [CrossRef]
- Santiago, L.S.; Wright, S.J. Leaf Functional Traits of Tropical Forest Plants in Relation to Growth Form. Funct. Ecol. 2007, 21, 19–27. [Google Scholar] [CrossRef]
- Wadzuk, B.M.; Lewellyn, C.; Lee, R. Green Infrastructure Recovery: Analysis of the Influence of Back-to-back Rainfall Events. J. Sustain. Water Built Environ. 2017, 3, 04017001. [Google Scholar] [CrossRef]
- Emerson, C.H.; Wadzuk, B.M.; Traver, R.G. Hydraulic Evolution and Total Suspended Solids Capture of an Infiltration Trench. Hydrol. Process. 2010, 24, 1008–1014. [Google Scholar] [CrossRef]
- Winston, R.J.; Dorsey, J.D.; Hunt, W.F. Quantifying Volume Reduction and Peak Flow Mitigation for Three Bioretention Cells in Clay Soils in Northeast Ohio. Sci. Total Environ. 2016, 553, 83–95. [Google Scholar] [CrossRef]
- De Paola, F.; Giugni, M.; Pugliese, F.; Romano, P. Optimal Design of LIDs in Urban Stormwater Systems Using a Harmony-Search Decision Support System. Water Resour. Manag. 2018, 32, 4933–4951. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Unit | Meaning | Value |
|---|---|---|---|
| θr | m3 m−3 | Residual water content | 0.029 |
| θs | m3 m−3 | Saturated water content | 0.479 |
| α | cm−1 | Air entry suction parameter | 0.040 |
| n | dimensionless | Pore size distribution parameter | 1.344 |
| Ks | cm min−1 | Saturated hydraulic conductivity | 0.296 |
| Storm Set | Count | Length (hr) | Depth (mm) | Intensity (mm hr−1) | Peak Intensity (mm 5-min−1) |
|---|---|---|---|---|---|
| All storms | 119 | 11.9 | 20.0 | 3.9 | 2.6 |
| Detectable in infiltration bed | 66 | 11.5 | 23.2 | 5.5 | 3.4 |
| Model Term | SE | 95% CI | |
|---|---|---|---|
| Period 2 | 0.019 | 0.069 | (−0.116, 0.155) |
| Period 3 | −0.176 | 0.075 | (−0.324, -0.029) |
| Period 4 | −0.038 | 0.070 | (−0.176, 0.099) |
| Period 5 | 0.098 | 0.070 | (−0.040, 0.236) |
| Canopy position | −0.375 | 0.055 | (−0.483, −0.268) |
| Species | 0.427 | 0.100 | (0.232, 0.622) |
| GI identity | −0.144 | 0.075 | (−0.290, 0.003) |
| Period 2 × Species | 0.022 | 0.119 | (−0.211, 0.255) |
| Period 3 × Species | −0.251 | 0.200 | (−0.643, 0.140) |
| Period 4 × Species | 0.002 | 0.119 | (−0.232, 0.236) |
| Period 5 × Species | −0.135 | 0.146 | (−0.421, 0.151) |
| Canopy position × Species | −0.205 | 0.110 | (−0.420, 0.010) |
| Canopy position × GI identity | −0.016 | 0.068 | (−0.149, 0.118) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tu, M.-c.; Caplan, J.S.; Eisenman, S.W.; Wadzuk, B.M. When Green Infrastructure Turns Grey: Plant Water Stress as a Consequence of Overdesign in a Tree Trench System. Water 2020, 12, 573. https://doi.org/10.3390/w12020573
Tu M-c, Caplan JS, Eisenman SW, Wadzuk BM. When Green Infrastructure Turns Grey: Plant Water Stress as a Consequence of Overdesign in a Tree Trench System. Water. 2020; 12(2):573. https://doi.org/10.3390/w12020573
Chicago/Turabian StyleTu, Min-cheng, Joshua S. Caplan, Sasha W. Eisenman, and Bridget M. Wadzuk. 2020. "When Green Infrastructure Turns Grey: Plant Water Stress as a Consequence of Overdesign in a Tree Trench System" Water 12, no. 2: 573. https://doi.org/10.3390/w12020573
APA StyleTu, M.-c., Caplan, J. S., Eisenman, S. W., & Wadzuk, B. M. (2020). When Green Infrastructure Turns Grey: Plant Water Stress as a Consequence of Overdesign in a Tree Trench System. Water, 12(2), 573. https://doi.org/10.3390/w12020573