Effects of Low Concentration of Selected Analgesics and Successive Bioaugmentation of the Activated Sludge on Its Activity and Metabolic Diversity





Abstract
:1. Introduction
2. Materials and Methods
2.1. Characteristics of the Activated Sludge
2.2. Composition of Bacterial Consortium
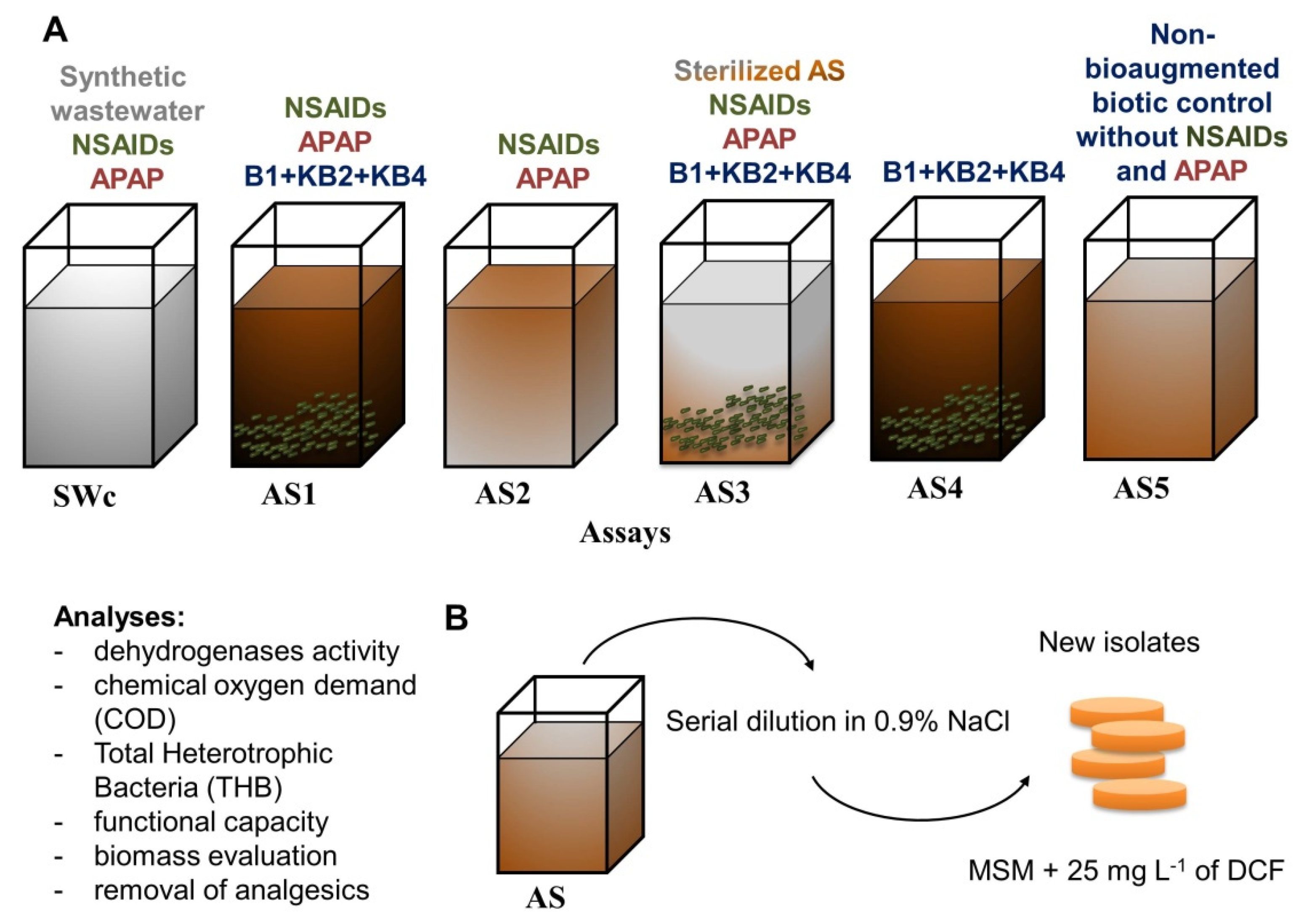
2.3. Experimental Set-Up and Isolating Diclofenac-Degrading Strains
2.4. Evaluating the Activated Sludge Quality and Impact of Bioaugmentation
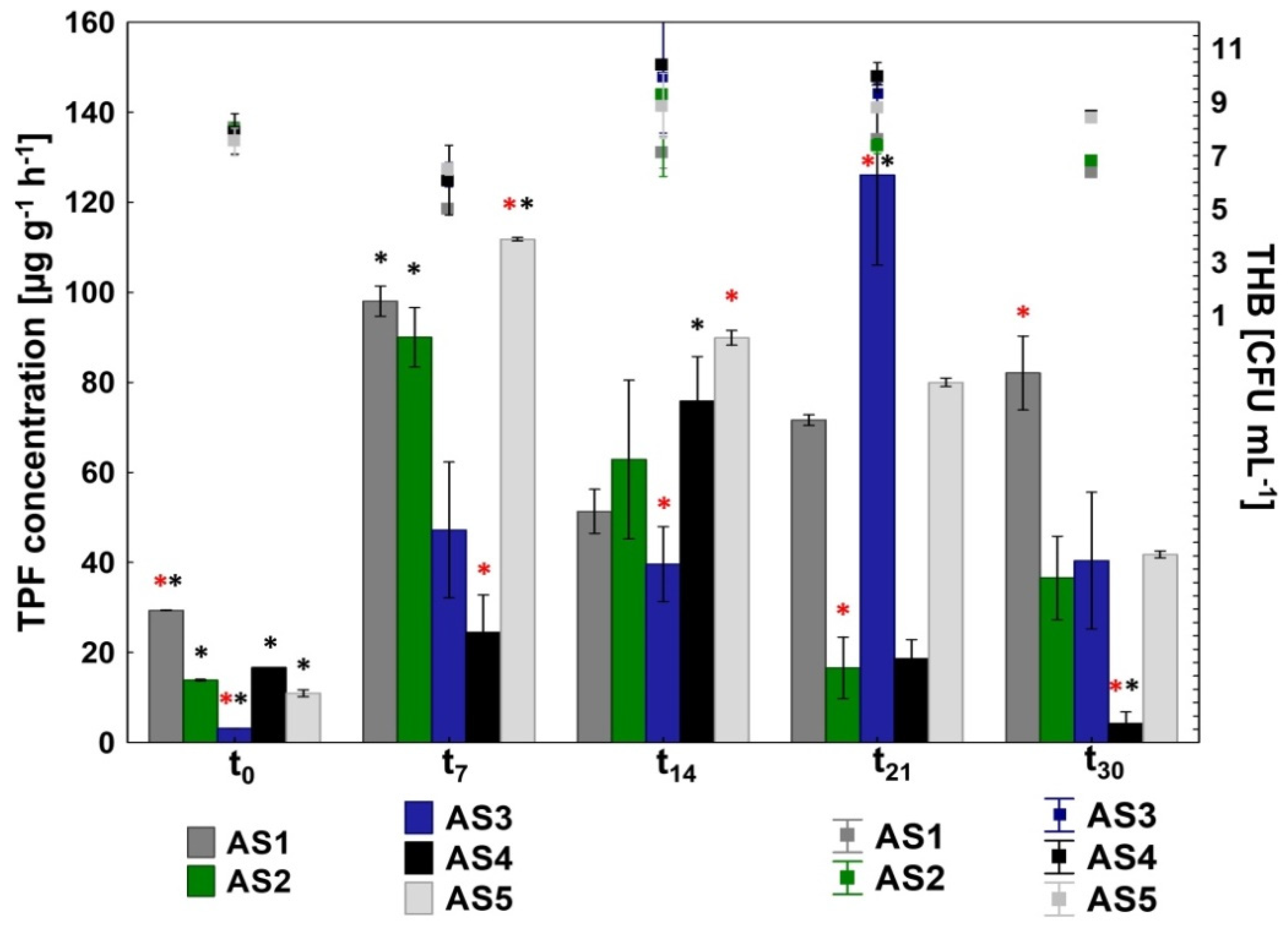
2.4.1. Measuring the Dehydrogenases Activity
2.4.2. Analyzing the Chemical Oxygen Demand
2.4.3. Enumerating the Total Heterotrophic Bacteria
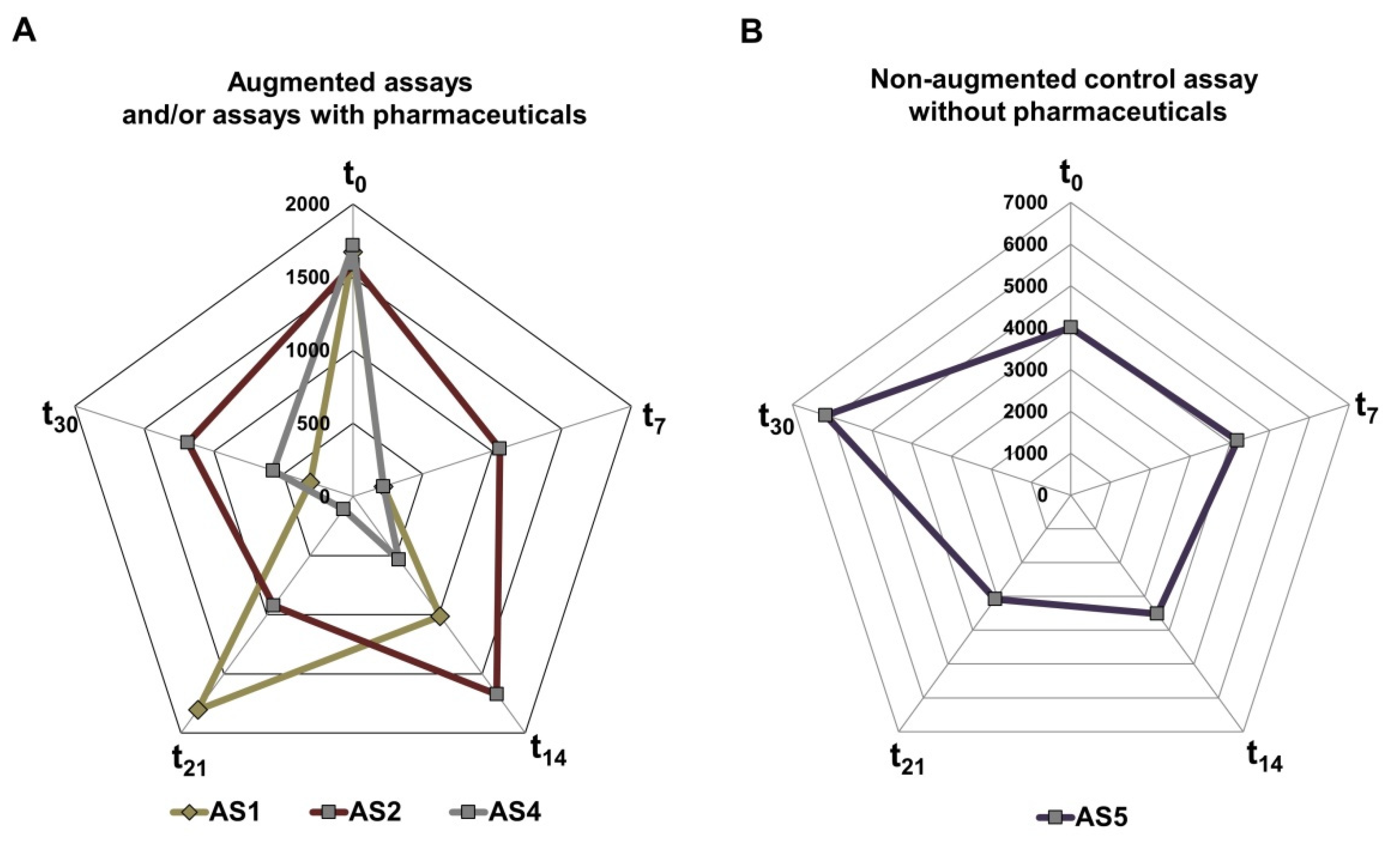
2.4.4. Metabolic Activity of the AS Measuring by BIOLOG
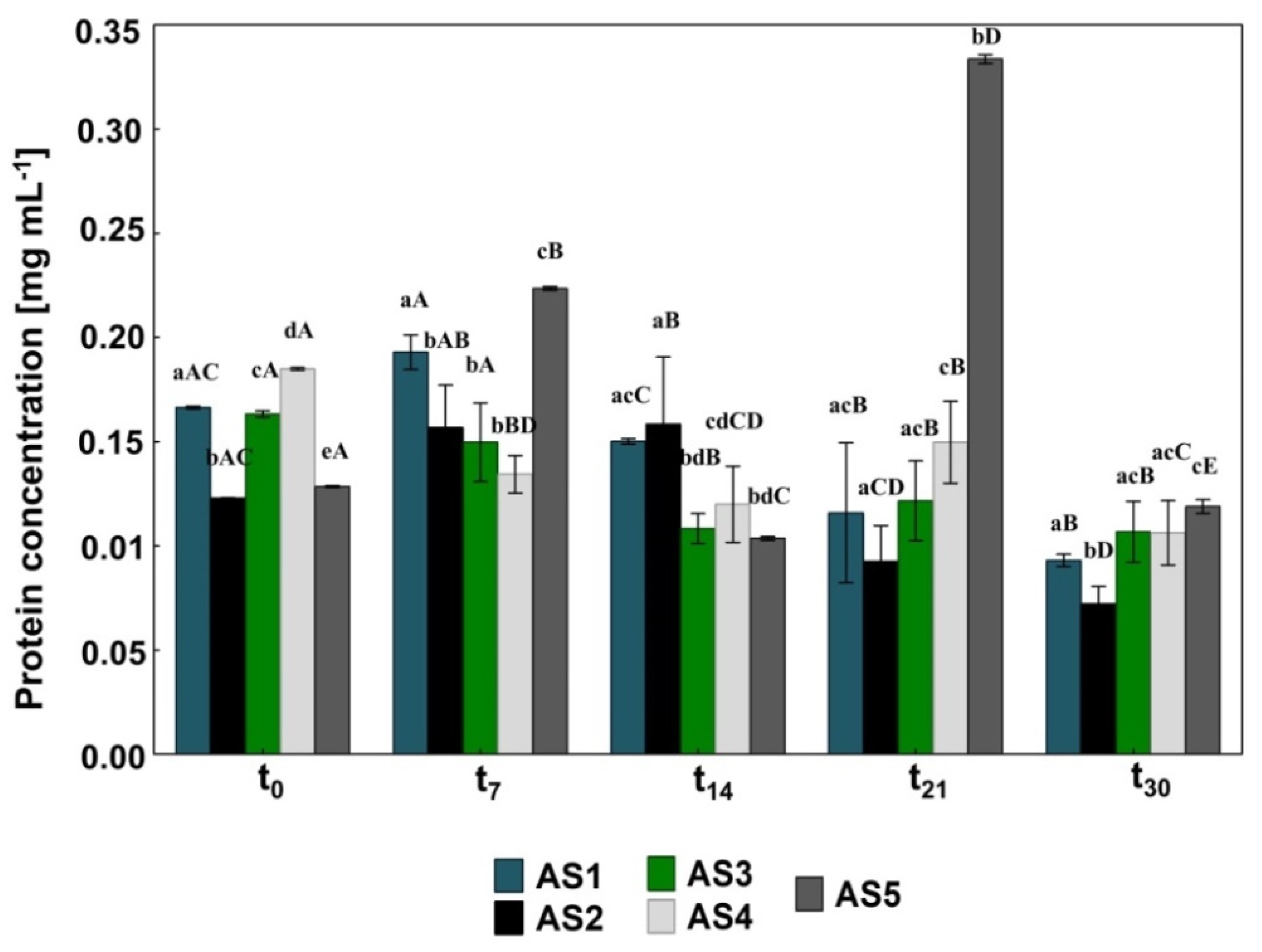
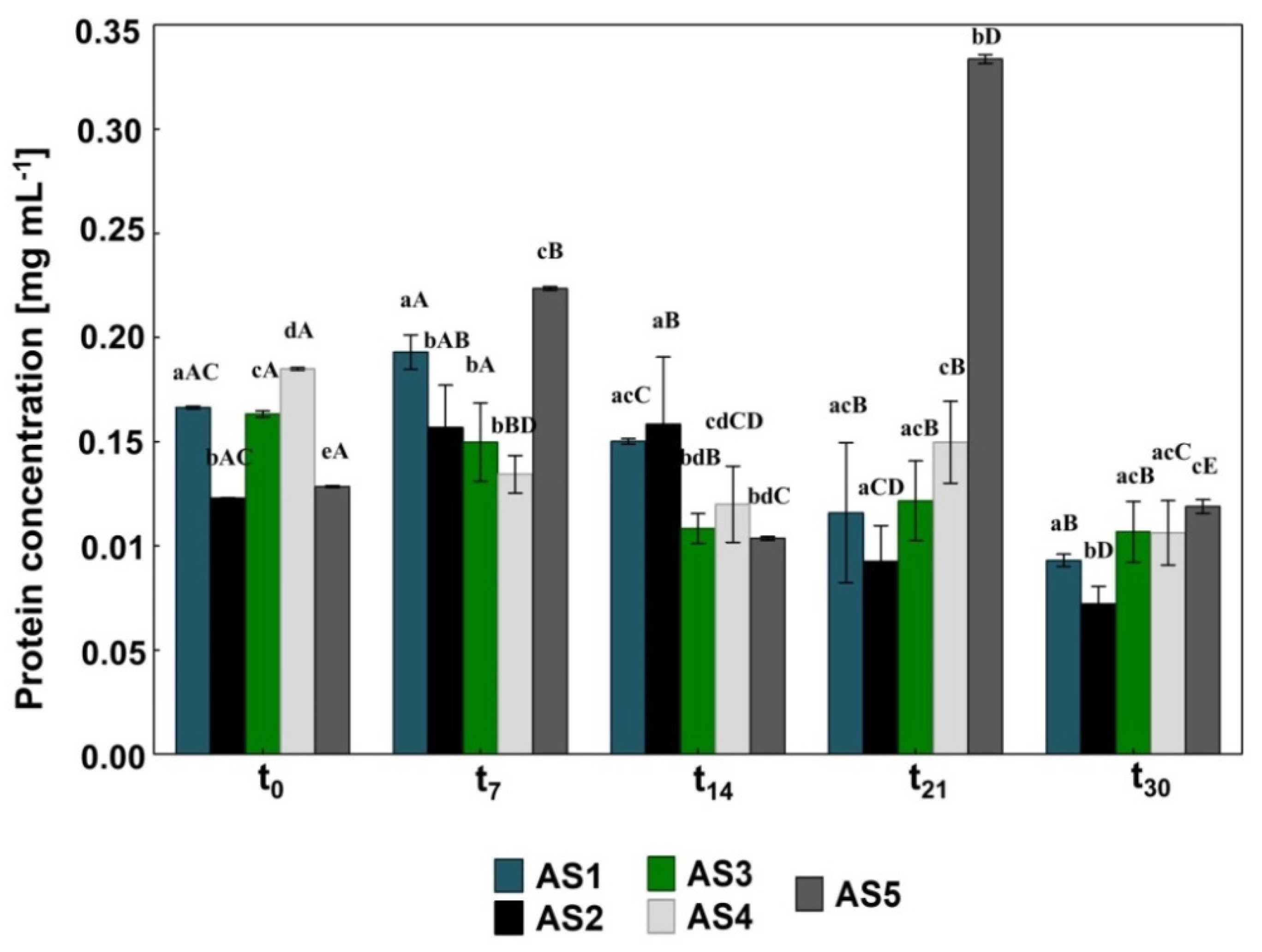
2.5. Measuring the Protein Concentration
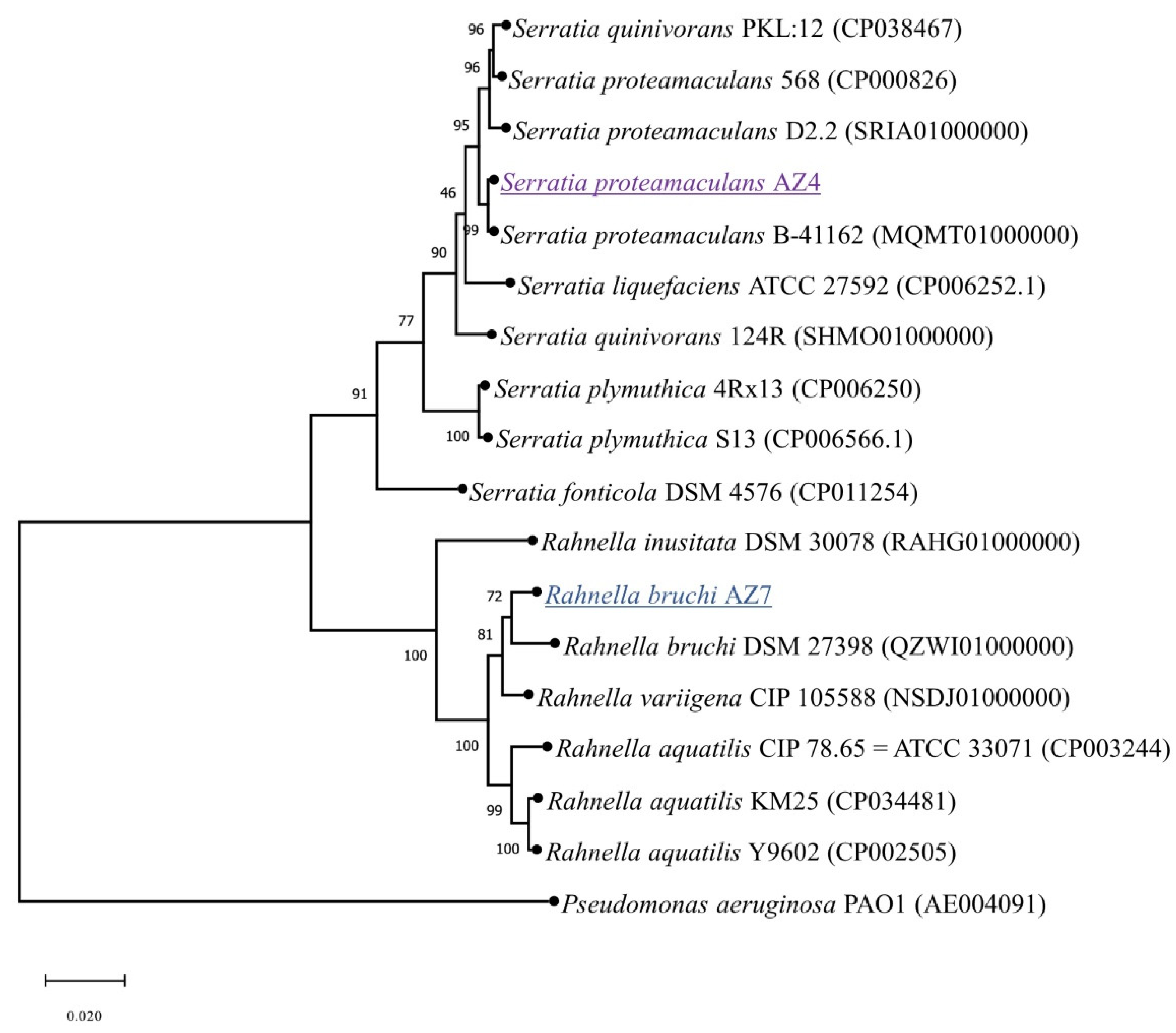
2.6. Multi-Locus Sequence Typing and Phylogenetic Analysis of the Newly Isolated Strains
2.7. Measuring the Analgesics Concentrations
2.8. Identifying the Intermediates Formed During Degradation
2.9. Statistical Analyses
3. Results
3.1. The Impact of Pharmaceuticals and Bioaugmentation on the AS Quality
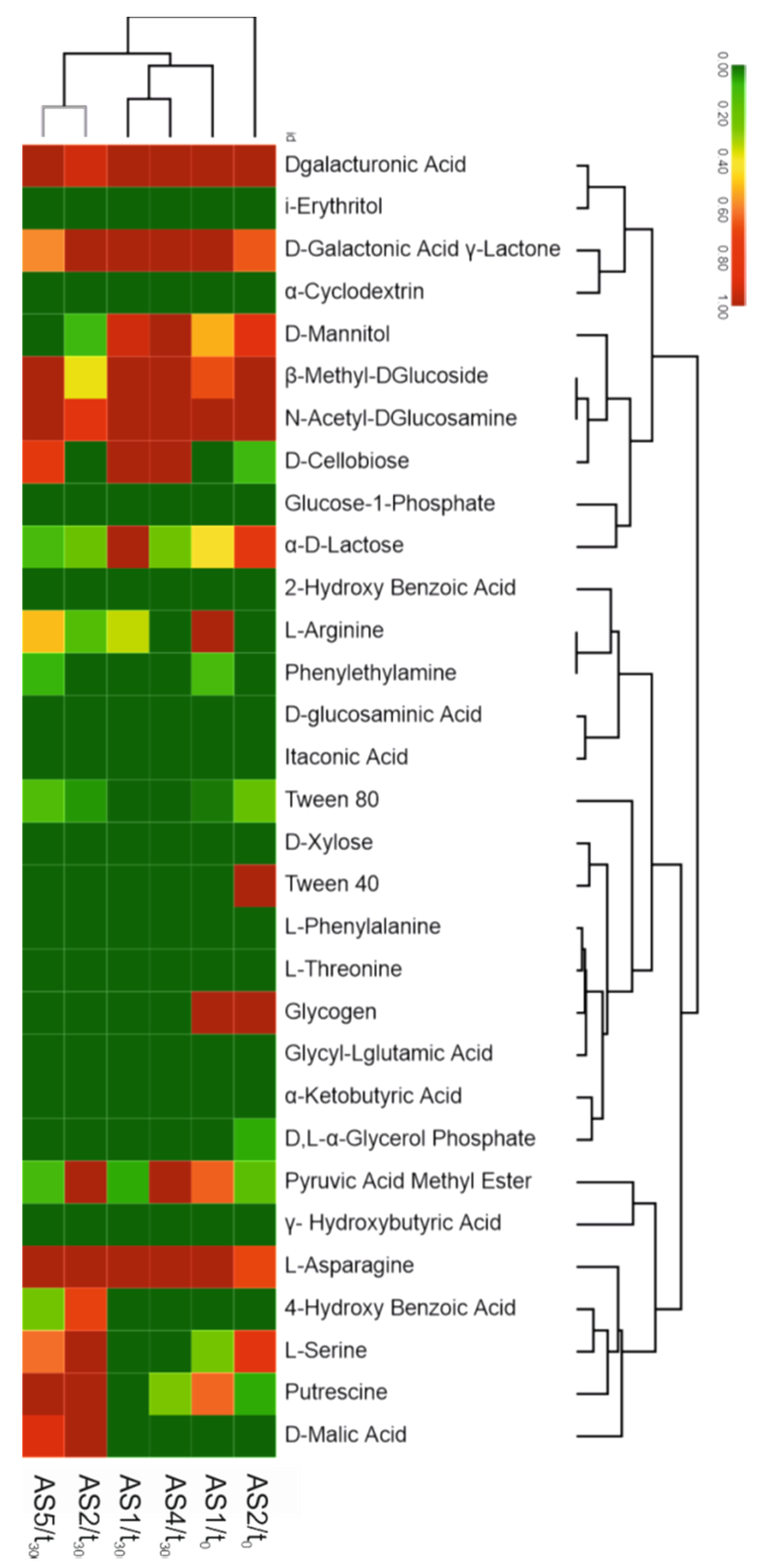
3.2. Microbial Activity in the Activated Sludge
3.3. Removal of Pharmaceuticals
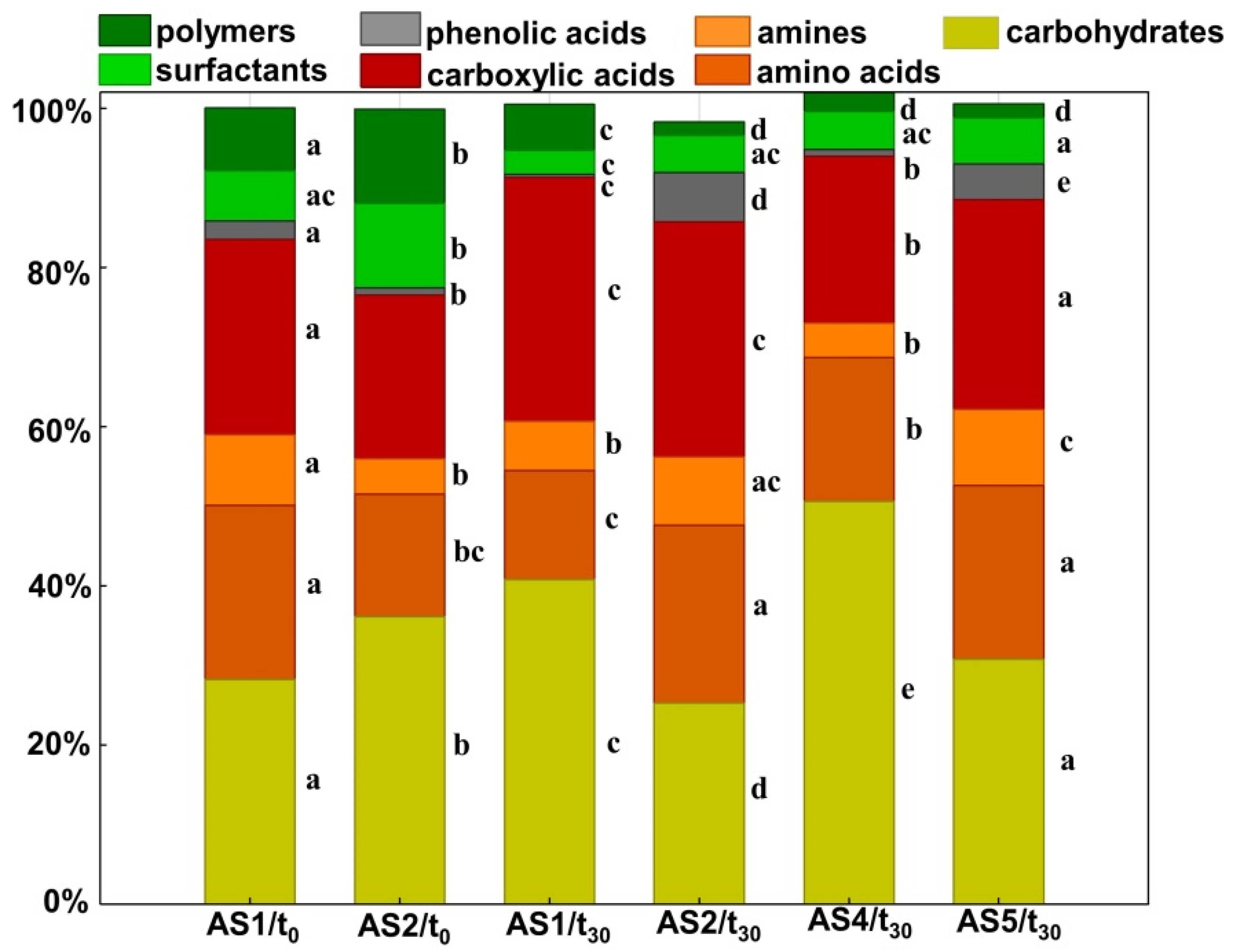
3.4. Secondary Metabolites Formation
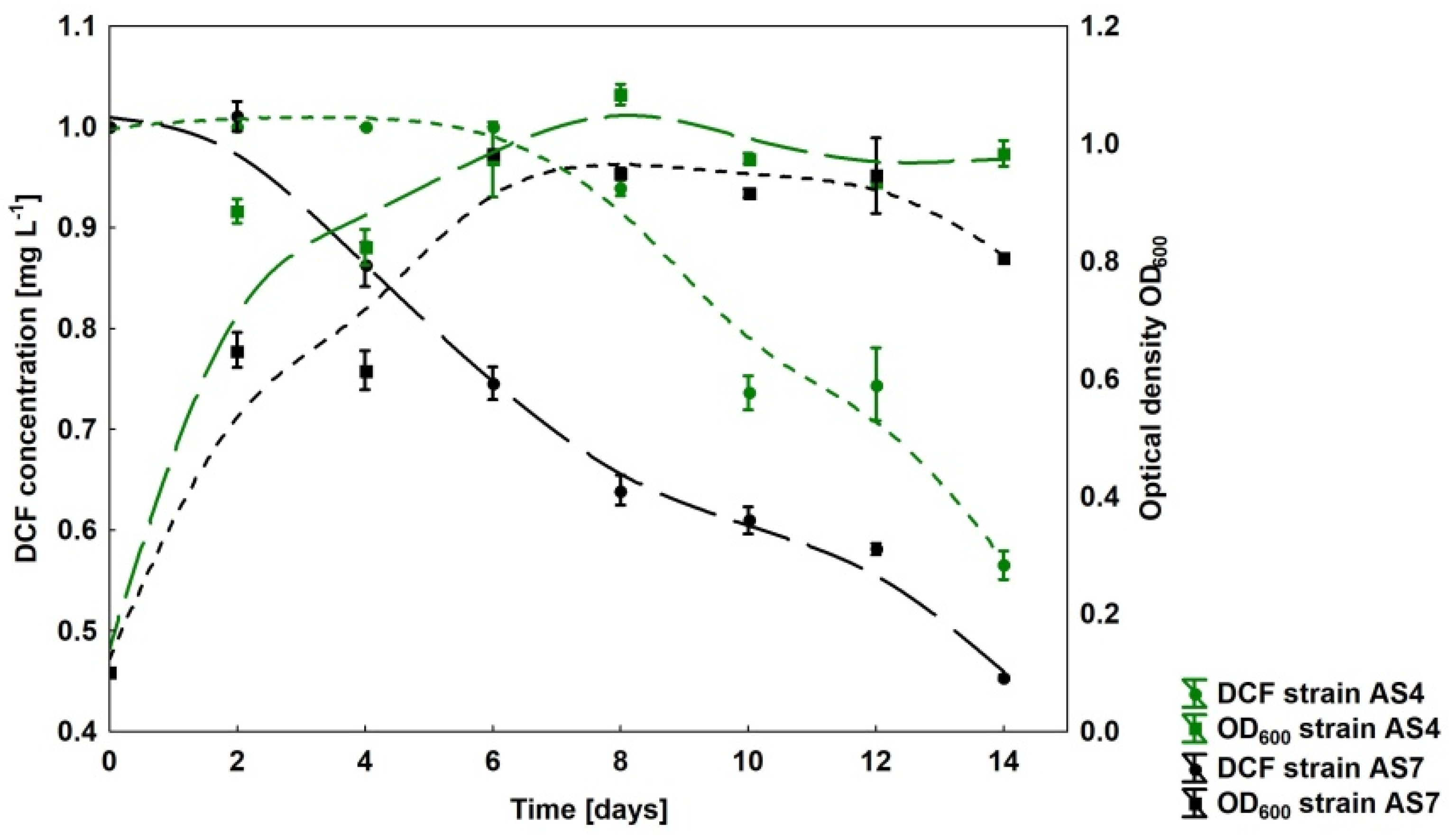
3.5. New Diclofenac-Degrading Strains
4. Discussion
4.1. The Impact of Bioaugmentation and Pharmaceuticals on the AS Quality and Metabolic Diversity
4.2. Pharmaceuticals Removal and Secondary Metabolites Formation
4.3. Isolation of New DCF-Degrading Strains
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Benotti, M.J.; Brownawell, B.J. Microbial degradation of pharmaceuticals in estuarine and coastal seawater. Environ. Pollut. 2009, 157, 994–1002. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Chu, K.H.; Al-Hamadani, Y.A.J.; Park, C.M.; Jang, M.; Kim, D.-H.; Yu, M.; Heo, J.; Yoon, Y. Removal of contaminants of emerging concern by membranes in water and wastewater: A review. Chem. Eng. J. 2018, 335, 896–914. [Google Scholar] [CrossRef]
- Roberts, P.H.; Thomas, K.V. The occurrence of selected pharmaceuticals in wastewater effluent and surface waters of the lower Tyne catchment. Sci. Pollut. Res. 2006, 356, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Esterhuizen-Londt, M.; Schwartz, K.; Pflugmacher, S. Using aquatic fungi for pharmaceutical bioremediation: Uptake of acetaminophen by Mucor hiemalis does not result in an enzymatic oxidative stress response. Fungal Biol. 2016, 120, 1249–1257. [Google Scholar] [CrossRef] [PubMed]
- Guerra, P.; Kim, M.; Shah, A.; Alaee, M.; Smyth, S.A. Occurrence and fate of antibiotic, analgesic/anti-inflammatory, and antifungal compounds in five wastewater treatment processes. Sci. Total Environ. 2014, 473, 235–243. [Google Scholar] [CrossRef]
- Luo, Y.; Guo, W.; Ngo, H.H.; Nghiem, L.D.; Hai, F.I.; Zhang, J.; Liang, J.; Wang, X. A review on the occurrence of micropollutants in the aquatic environment and their fate and removal during wastewater treatment. Sci. Total Environ. 2014, 473–474, 619–641. [Google Scholar] [CrossRef]
- Matongo, S.; Birungi, G.; Moodley, B.; Ndungu, P. Pharmaceutical residues in water and sediment of Msunduzi River, Kwa Zulu-Natal, South Africa. Chemosphere 2015, 134, 133–140. [Google Scholar] [CrossRef]
- Ashfaq, M.; Noor, N.; Saif-Ur-Rehman, M.; Sun, Q.; Mustafa, G.; Nazar, M.F.; Yu, C.-P. Determination of commonly used pharmaceuticals in hospital waste of Pakistan and evaluation of their ecological risk assessment. Clean Water Air Soil 2017, 45, 1500392. [Google Scholar] [CrossRef]
- Tiwari, B.; Sellamuthu, B.; Ouarda, Y.; Drogui, P.; Tyagi, R.D.; Buelna, G. Review on fate and mechanism of removal of pharmaceutical pollutants from wastewater using biological approach. Bioresour. Technol. 2017, 224, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Żur, J.; Piński, A.; Marchlewicz, M.; Hupert-Kocurek, K.; Wojcieszyńska, D.; Guzik, U. Organic micropollutants paracetamol and ibuprofen—Toxicity, biodegradation, and genetic background of their utilization by bacteria. Environ. Sci. Pollut. Res. 2018, 25, 21498–21524. [Google Scholar] [CrossRef] [Green Version]
- Falås, P.; Baillon-Dhumez, A.; Andersen, H.R.; Ledin, A.; la Cour Jansen, J. Suspended biofilm carrier and activated sludge removal of acidic pharmaceuticals. Water Res. 2012, 46, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- Festa, S.; Coppotelli, B.M.; Morelli, I.S. Comparative bioaugmentation with a consortium and a single strain in a phenanthrene-contaminated soil: Impact on the bacterial community and biodegradation. Appl. Soil Ecol. 2016, 98, 8–19. [Google Scholar] [CrossRef]
- Loperana, L.; Saravia, V.; Murro, D.; Ferrari, M.D.; Lareo, C. Kinetic properties of a commercial and a native inoculum for aerobic milk fat degradation. Bioresour. Technol. 2006, 97, 2160–2165. [Google Scholar] [CrossRef] [PubMed]
- Loperana, L.; Ferrari, M.D.; Saravia, V.; Murro, D.; Lima, C.; Ferrando, L.; Fernández, A.; Lareo, C. Performance of a commercial inoculum for the aerobic biodegradation of a high fat content dairy wastewater. Bioresour. Technol. 2007, 98, 1045–1051. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.C.; van der Gast, C.J.; Thompson, I.P. Perspectives and vision for strain selection in bioaugmentation. Trends Biotechnol. 2005, 23, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Michalska, J.; Greń, I.; Żur, J.; Wasilkowski, D.; Mrozik, A. Impact of the biological cotreatment of the Kalina Pond leachate on laboratory sequencing batch reactor operation and activated sludge quality. Water 2019, 11, 1539. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Raquet, G.; Durand, E.; Braun, F.; Cravo-Laureau, C.; Godon, J.-J. Impact of microbial diversity depletion on xenobiotic degradation by sewage-activated sludge. Environ. Microbiol. Rep. 2013, 4, 588–594. [Google Scholar] [CrossRef]
- Domaradzka, D.; Guzik, U.; Wojcieszyńska, D. Biodegradation and biotransformation of polycyclic non-steroidal anti-inflammatory drugs. Rev. Environ. Sci. Biotechnol. 2015, 14, 229–239. [Google Scholar] [CrossRef] [Green Version]
- Thompson, I.P.; Van Der Gast, C.J.; Ciric, L.; Singer, A. Bioaugmentation for bioremediation: The challenge of strain selection. Environ. Microbiol. 2005, 7, 909–915. [Google Scholar] [CrossRef]
- Nzila, A.; Razzak, S.A.; Zhu, J. Bioaugmentation: An emerging strategy of industrial wastewater treatment for reuse and discharge. Int. J. Environ. Res. Public Health 2016, 13, 846. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, J.; Zhu, R.; Zhou, Q.; Chen, J. Degradation of paracetamol by pure bacterial cultures and their microbial consortium. Appl. Microbiol. Biotechnol. 2013, 97, 3687–3698. [Google Scholar] [CrossRef] [PubMed]
- Kolvenbach, B.A.; Helbling, D.E.; Kohler, H.-P.E.; Corvini, P.F.-X. Emerging chemicals and the evolution of biodegradation capacities and pathways in bacteria. Curr. Opin. Biotechnol. 2014, 27, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; McLachlan, M.S. Biodegradation of chemicals in unspiked surface waters downstream of wastewater treatment plants. Environ. Sci. Technol. 2019, 53, 1884–1892. [Google Scholar] [CrossRef] [Green Version]
- APHA. Standard Methods for the Examination of Water and Wastewater; American Public Health Association, American Water Works Association, Water Environment Federation: Washington, DC, USA, 1999. [Google Scholar]
- Miksch, K. Application of dehydrogenase activity determinations in biodegradation of refinery sewage. Gas Water Civ. Technol. 1977, 51, 234–235. [Google Scholar]
- Schumacher, T.E.; Eynard, A.; Chintala, R. Rapid cost-effective analysis of microbial activity in soils using modified fluorescein diacetate method. Environ. Sci. Pollut. Res. 2015, 22, 4759–4762. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.W.; Yan, J.Z.; Seng, C.E. Performance of phenol-acclimated activated sludge in the presence of various phenolic compound. Appl. Water Sci. 2013, 3, 515–525. [Google Scholar] [CrossRef] [Green Version]
- EN ISO 8199:2010. Water Quality—General Requirements and Guidance for Microbiological Examinations by Culture; International Organization for Standardization: Geneva, Switzerland, 2010. [Google Scholar]
- Marchlewicz, A.; Domaradzka, D.; Guzik, U.; Wojcieszyńska, D. Bacillus thuringiensis B1(2015b) is a Gram-positive bacteria able to degrade naproxen and ibuprofen. Water Air Soil Pollut. 2016, 227, 197. [Google Scholar] [CrossRef] [Green Version]
- Guzik, U.; Greń, I.; Wojcieszyńska, D.; Łabużek, S. Isolation and characterization of a novel strain of Stenotrophomonas maltophilia possessing various dioxygenases for monocyclic hydrocarbon degradation. Braz. J. Microbiol. 2009, 40. [Google Scholar] [CrossRef] [Green Version]
- Żur, J.; Wojcieszyńska, D.; Hupert-Kocurek, K.; Marchlewicz, A.; Guzik, U. Paracetamol—Toxicity and microbial utilization. Pseudomonas moorei KB4 strain as a case study for exploring degradation pathway. Chemosphere 2018, 206, 192–202. [Google Scholar] [CrossRef]
- Wojcieszyńska, D.; Guzik, U.; Greń, I.; Perkosz, M.; Hupert-Kocurek, K. Induction of aromatic ring: Cleavage dioxygenases in Stenotrophomonas maltophilia strain KB2 in cometabolic systems. World J. Microbiol. Biotechnol. 2011, 27, 805–811. [Google Scholar] [CrossRef] [Green Version]
- Miksch, K. The influence of the concentration of dissolved oxygen on the determination of the dehydrogenase activity of activated sludge using method of TTC test. Clean Soil Air Water. 1983, 11, 17–21. [Google Scholar] [CrossRef]
- Nowak, A.; Mrozik, A. Degradation of 4-chlorophenol and microbial diversity in soil inoculated with single Pseudomonas sp. CF600 and Stenotrophomonas maltophilia KB2. J. Environ. Manag. 2018, 215, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Eisen, J. A simple, fast, and accurate method of phylogenomic inference. Genome Biol. 2008, 9, R151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial bioinformatics database and analysis resource center. Nucl. Acid Res. 2017, 45, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Rozas, O.; Vidal, C.; Baeza, C.; Jardim, W.F.; Rossner, A.; Mansilla, H.D. Organic micropollutants (OMPs) in natural waters: Oxidation by UV/H2O2 treatment and toxicity assessment. Water Res. 2016, 98, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Grujíć, S.; Vasiljević, T.; Laušević, M. Determination of multiple pharmaceutical classes in surface and ground waters by liquid chromatography-ion-trap-tandem-mass spectrometry. J. Chromatogr. A 2009, 1216, 4989–5000. [Google Scholar] [CrossRef]
- Paxéus, N. Removal of selected non-steroidal anti-inflammatory drugs (NSAIDs), gemfibrozil, carbamazepine, β-blockers, trimethoprim and triclosan in conventional wastewater treatment plants in five EU countries and their discharge to the aquatic environment. Water Sci. Technol. 2004, 50, 253–260. [Google Scholar] [CrossRef]
- Quintana, J.B.; Weiss, S.; Reemtsma, T. Pathways and metabolites of microbial degradation of selected acidic pharmaceutical and their occurrence in municipal wastewater treated by membrane bioreactor. Water Res. 2005, 39, 2654–2664. [Google Scholar] [CrossRef]
- Salgado, R.; Marques, R.; Noronha, J.P.; Carvalho, G.; Oehmen, A.; Reis, M.A. Assessing the removal of pharmaceuticals and personal care products in a full-scale activated sludge plant. Environ. Sci. Pollut. Res. Int. 2012, 19, 1818–1827. [Google Scholar] [CrossRef]
- Aissaoui, S.; Ouled-Haddar, H.; Sifour, M.; Harrouche, K.; Sghaier, H. Metabolic and co-metabolic transformation of diclofenac by Enterobacter hormachei D15 isolated from activated sludge. Curr. Microbiol. 2016, 74, 381–388. [Google Scholar] [CrossRef]
- Jiménez-Silva, V.A.; Santoyo-Tepole, F.; Ruiz-Ordaz, N.; Galíndez-Mayer, J. Study of the ibuprofen impact on wastewater treatment mini-plants with bioaugmented sludge. Process Saf. Environ. 2019, 123, 140–149. [Google Scholar] [CrossRef]
- Almeida, B.; Oehmen, A.; Marques, R.; Brito, D.; Carvalho, G.; Barreto Crespo, M.T. Modelling the biodegradation of non-steroidal anti-inflammatory drugs (NSAIDs) by activated sludge and a pure culture. Bioresour. Technol. 2013, 133, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Reardon, K.F.; Mosteller, D.C.; Rogers, J.B.; Du Teau, N.M.; Kim, K.H. Biodegradation kinetics of aromatic hydrocarbon mixtures by pure and mixed bacterial cultures. Environ. Health Perspect. 2002, 6, 1005–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Wen, X.; Zhang, B.; Yang, Z. Diversity and assembly patterns of activated sludge microbial communities: A review. Biotechnol. Adv. 2018, 36, 1038–1047. [Google Scholar] [CrossRef]
- Hailei, W.; Ping, L.; Ying, W.; Lei, L.; Jianming, Y. Metagenomic insight into the bioaugmentation mechanism of Phanerochaete chrysosporium in an activated sludge system treating coking wastewater. J. Hazard. Mater. 2017, 321, 820–829. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Yang, Q.; Wang, J.L. Biodegradation of di-n-butyl phthalate in sequencing batch reactor bioaugmented with Micrococcus sp. and the bacterial community analysis. Int. J. Environ. Sci. Technol. 2015, 12, 2819–2828. [Google Scholar] [CrossRef] [Green Version]
- De Gusseme, B.; Vanhaecke, L.; Verstraete, W.; Boona, N. Degradation of acetaminophen by Delftia tsuruhatensis and Pseudomonas aeruginosa in a membrane bioreactor. Water Res. 2011, 45, 1829–1837. [Google Scholar] [CrossRef]
- Musson, S.E.; Campo, P.; Tolaymat, T.; Suidan, M.; Townsend, T.G. Assessment of the anaerobic degradation of six active pharmaceutical ingredients. Sci. Total. Environ. 2010, 408, 2068–2074. [Google Scholar] [CrossRef]
- Marchlewicz, A.; Guzik, U.; Smułek, W.; Wojcieszyńska, D. Exploring the degradation of ibuprofen by Bacillus thuringiensis B1(2015b): The new pathway and factors affecting degradation. Molecules 2017, 22, 1676. [Google Scholar] [CrossRef]
- Parolini, M.; Binelli, A.; Provini, A. Chronic effects induced by ibuprofen on the freshwater bivalve Dreissena polymorpha. Ecotoxicol. Environ. Saf. 2011, 74, 1586–1594. [Google Scholar] [CrossRef]
- Kosjek, T.; Heath, E.; Perez, S.; Petrovic, M.; Barcelo, D. Metabolism studies of diclofenac and clofibric acid in activated sludge bioreactors using liquid chromatography with quadrupole and time-of-flight mass spectrometry. J. Hydrol. 2009, 372, 109–117. [Google Scholar] [CrossRef]
- Al-Rajab, A.J.; Sabourin, L.; Lapen, D.R.; Topp, E. The non-steroidal anti-inflammatory drug diclofenac is readily biodegradable in agricultural soils. Sci. Total Environ. 2010, 409, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Langenhoff, A.; Inderfurth, N.; Veuskens, T.; Schraa, G.; Blokland, M.; Kujawa-Roeleveld, K.; Rijnaarts, H. Microbial removal of the pharmaceuticals compounds ibuprofen and diclofenac from wastewater. BioMed Res. Int. 2013, 2013, 325806. [Google Scholar] [CrossRef]
- Jewell, K.S.; Falås, P.; Wick, A.; Joss, A.; Ternes, T.A. Transformation of diclofenac in hybrid biofilm activated sludge processes. Water Res. 2016, 105, 559–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matamoros, V.; Arias, C.; Brix, H.; Bayona, J.M. Preliminary screening of small-scale domestic wastewater treatment systems for removal of pharmaceutical and personal care products. Water Res. 2009, 43, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Górny, D.; Guzik, U.; Hupert-Kocurek, K.; Wojcieszyńska, D. A new pathway for naproxen utilisation by Bacillus thuringiensis B1(2015b) and its decomposition in the presence of organic and inorganic contaminants. J. Environ. Manag. 2019, 239, 1–7. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, L.; Chen, J. Paracetamol in the environment and its degradation by microorganisms. Appl. Microbiol. Biotechnol. 2012, 96, 875–884. [Google Scholar] [CrossRef]
- Żur, J.; Piński, A.; Michalska, J.; Hupert-Kocurek, K.; Nowak, A.; Wojcieszyńska, D.; Guzik, U. A whole-cell immobilization system on bacterial cellulose for the paracetamol-degrading Pseudomonas moorei KB4 strain. Int. Biodeterior. Biodegrad. 2020, 149, 104919. [Google Scholar] [CrossRef]
- Murdoch, R.W.; Hay, A.G. The biotransformation of ibuprofen to trihydroxyibuprofen in activated sludge by Variovorax Ibu-1. Biodegradation 2015, 26, 105–113. [Google Scholar] [CrossRef]
- Patiño, L.H.; Camargo, M.; Muñoz, M.; Ríos-Chaparro, D.I.; Patarroyo, M.A.; Ramírez, J.D. Unveiling the Multilocus Sequence Typing (MLST) schemes and core genome phylogenies for genotyping Chlamydia trachomatis. Front. Microbiol. 2018, 9, 1854. [Google Scholar] [CrossRef]
- Hu, Y.; Wei, L.; Feng, Y.; Xie, Y.; Zong, Z. Klebsiella huaxiensis sp. nov., recovered from human urine. Int. J. Syst. Evol. Microbiol. 2019, 69, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Domaradzka, D.; Guzik, U.; Hupert-Kocurek, K.; Wojcieszyńska, D. Toxicity of diclofenac and its biotransformation by Raoultella sp. DD4. Pol. J. Environ. Stud. 2016, 25, 2211–2216. [Google Scholar] [CrossRef]
- Bessa, V.S.; Moreira, I.S.; Tiritan, M.E.; Castro, P.M.L. Enrichment of bacterial strains for the biodegradation of diclofenac and carbamazepine from activated sludge. Int. Biodeterior. Biodegrad. 2017, 120, 135–142. [Google Scholar] [CrossRef]
- Palyzová, A.; Zahradník, J.; Marešová, H.; Sokolová, L.; Kyslíková, E.; Grulich, M.; Štěpánek, V.; Řezanka, T.; Kyslík, P. Potential of the strain Raoultella sp. KDF8 for removal of analgesics. Folia Microbiol. (Praha) 2018, 63, 273–282. [Google Scholar] [CrossRef]
- Moreira, I.S.; Bessa, V.S.; Murgolo, S.; Piccirillo, C.; Mascolo, G.; Castro, P.M.L. Biodegradation of diclofenac by the bacterial strain Labrys portucalensis F11. Ecotoxicol. Environ. Saf. 2018, 152, 104–113. [Google Scholar] [CrossRef]
- Stylianou, K.; Hapeshi, E.; Vasquez, M.; Fatta-Kassinos, D.; Vyrides, I. Diclofenac biodegradation by newly isolated Klebsiella sp. KSC: Microbial intermediates and ecotoxicological assessment. J. Environ. Chem. Eng. 2018, 6, 3242–3248. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| pH | 7.713 ± 0.071 |
| SVI, g cm−3 | 114.182 ± 1.665 |
| MLSS, g L−1 | 3.667 ± 0.180 |
| SOUR, mg O2 g−1 MLSS h−1 | 8.424 ± 0.621 |
| DHA, mg TPF g−1 MLSS h−1 | 16.096 ± 0.952 |
| NSEA, µg fluorescein g−1 MLSS h−1 | 23.012 ± 0.697 |
| Number of the THB, log CFU g−1 MLSS−1 | 8.226 ± 0.992 |
| Bacterial Strain | Site of Isolation | Degradation of Aromatics | Reference |
|---|---|---|---|
| Bacillus thuringiensis B1(2015b) | Contaminated soil Jaworzno, Poland | Ibuprofen, naproxen | [29] |
| Stenotrophomonas maltophilia KB2 | Activated sludge Miechowice, Poland | Naproxen, phenol, mononithrophenols, methylphenols | [30] |
| Pseudomonas moorei KB4 | Activated sludge Chorzów, Poland | Paracetamol, diclofenac, phenol, 4-hydroxybenzoic acid | [31] |
| Parameter | AS1/t0 | AS2/t0 | AS1/t30 | AS2/t30 | AS4/t30 | AS5/t30 |
|---|---|---|---|---|---|---|
| Functional Diversity Indices | ||||||
| H’Eco | 1.27 ± 0.02 a | 1.24 ± 0.02 b | 1.28 ± 0.02 a | 1.28 ± 0.01 a | 1.29 ± 0.01 c | 1.31 ± 0.02 d |
| AWCD | 1.26 ± 0.04 a | 1.12 ± 0.11 b | 0.99 ± 0.09 c | 0.94 ± 0.05 c | 0.92 ± 0.07 c | 0.78 ± 0.04 d |
| G | 0.55 ± 0.04 a | 0.68 ± 0.05 b | 0.62 ± 0.03 c | 0.59 ± 0.03 c | 0.54 ± 0.03 ad | 0.51 ± 0.04 d |
| S | 27.11 ± 3.10 a | 22.00 ± 1.23 b | 25.89 ± 1.90 a | 26.56 ± 0.88 a | 27.67 ± 1.22 a | 26.22 ± 3.07 a |
| E | 0.88 ± 0.02 a | 0.92 ± 0.02 bc | 0.90 ± 0.01 ac | 0.90 ± 0.01 a | 0.90 ± 0.01 a | 0.93 ± 0.03 b |
| Parameter | AS1/t0 | AS2/t0 | AS1/t30 | AS2/t30 | AS4/t30 | AS5/t30 |
|---|---|---|---|---|---|---|
| Kinetic Parameters | ||||||
| Amax | 1.29 ± 0.04 ac | 1.04 ± 0.13 b | 1.32 ± 0.09 a | 1.19 ± 0.05 ab | 1.25 ± 0.10 ac | 1.10 ± 0.06 bc |
| b | 50.07 ± 5.63 a | 129.88 ± 19.04 b | 221.59 ± 34.59 cd | 180.78 ± 19.50 bc | 227.67 ± 30.22 d | 674.21 ± 60.82 e |
| k | 0.069 ± 0.004 a | 0.082 ± 0.006 b | 0.098 ± 0.003 c | 0.100 ± 0.004 c | 0.090 ± 0.002 d | 0.148 ± 0.005 e |
| Avt50 | 57.15 ± 2.91 ac | 58.054 ± 1.70 ad | 54.78 ± 1.18 bc | 52.65 ± 1.17 b | 60.46 ± 1.46 d | 44.62 ± 1.19 e |
| Pharmaceutical | Time of Analysis | Assay | Degraded Dose (mg L−1) |
|---|---|---|---|
| APAP | t7 | AS1 | 1.0 |
| AS2 | 1.0 | ||
| AS3 | 1.0 | ||
| t14 | AS1 | 1.0 | |
| AS2 | 1.0 | ||
| AS3 | 1.0 | ||
| t21 | AS1 | 1.0 | |
| AS2 | 1.0 | ||
| AS3 | 1.0 | ||
| t30 | AS1 | 1.0 | |
| AS2 | 1.0 | ||
| AS3 | 1.0 | ||
| IBU | t7 | AS1 | 1.0 |
| AS2 | 1.0 | ||
| AS3 | 1.0 | ||
| t14 | AS1 | 1.0 | |
| AS2 | 1.0 | ||
| AS3 | 1.0 | ||
| t21 | AS1 | 1.0 | |
| AS2 | 1.0 | ||
| AS3 | 1.0 | ||
| t30 | AS1 | 1.0 | |
| AS2 | 1.0 | ||
| AS3 | 0.94 ± 0.02 | ||
| NPX | t7 | AS1 | 0.5 |
| AS2 | 0.5 | ||
| AS3 | 0.5 | ||
| t14 | AS1 | 0.5 | |
| AS2 | 0.5 | ||
| AS3 | 0.5 | ||
| t21 | AS1 | 0.5 | |
| AS2 | 0.5 | ||
| AS3 | 0.5 | ||
| t30 | AS1 | 0.34 ± 0.00 | |
| AS2 | 0.32 ± 0.05 | ||
| AS3 | 0.44 ± 0.01 | ||
| DCF | t7 | AS1 | 0.14 ± 0.08 |
| AS2 | 0.12 ± 0.04 | ||
| AS3 | 0.33 ± 0.04 | ||
| t14 | AS1 | 0.5 ± 0.00 | |
| AS2 | 0.34 ± 0.04 | ||
| AS3 | 0.39 ± 0.02 | ||
| t21 | AS1 | 0.46 ± 0.01 | |
| AS2 | 0.31 ± 0.02 | ||
| AS3 | 0.5 | ||
| t30 | AS1 | 0.49 ± 0.00 | |
| AS2 | 0.43 ± 0.05 | ||
| AS3 | 0.48 ± 0.01 |
| Pharmaceutical | Metabolite | Time of Analysis | RT | UHPLC MS/MS (ng mL−1) |
|---|---|---|---|---|
| APAP | 4-aminophenol | t7 | 1.93 | <40.0 |
| t14 | <40.0 | |||
| t21 | <40.0 | |||
| t30 | <40.0 | |||
| Hydroquinone | t7 | nd | nd | |
| t14 | nd | |||
| t21 | nd | |||
| t30 | nd | |||
| NPX | O-desmethyl-NPX | t7 | 4.89 | <20.0 |
| t14 | <20.0 | |||
| t21 | <20.0 | |||
| t30 | <20.0 | |||
| IBU | 1-OH-IBU | t7 | 5.25 | <5.0 |
| t14 | <5.0 | |||
| t21 | <5.0 | |||
| t30 | <5.0 | |||
| 2-OH-IBU | t7 | 4.81 | <5.0 | |
| t14 | <5.0 | |||
| t21 | 8.6 ± 0.1 | |||
| t30 | <5.0 | |||
| DCF | DCF lactam | t7 | 8.98 | <20.0 |
| t14 | <20.0 | |||
| t21 | <20.0 | |||
| t30 | <20.0 | |||
| 4-OH-DCF | t7 | 6.11 | 48.9 ± 0.7 | |
| t14 | 72.1 ± 2.2 | |||
| t21 | 45.9 ± 3.4 | |||
| t30 | <20.0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Żur, J.; Michalska, J.; Piński, A.; Mrozik, A.; Nowak, A. Effects of Low Concentration of Selected Analgesics and Successive Bioaugmentation of the Activated Sludge on Its Activity and Metabolic Diversity. Water 2020, 12, 1133. https://doi.org/10.3390/w12041133
Żur J, Michalska J, Piński A, Mrozik A, Nowak A. Effects of Low Concentration of Selected Analgesics and Successive Bioaugmentation of the Activated Sludge on Its Activity and Metabolic Diversity. Water. 2020; 12(4):1133. https://doi.org/10.3390/w12041133
Chicago/Turabian StyleŻur, Joanna, Justyna Michalska, Artur Piński, Agnieszka Mrozik, and Agnieszka Nowak. 2020. "Effects of Low Concentration of Selected Analgesics and Successive Bioaugmentation of the Activated Sludge on Its Activity and Metabolic Diversity" Water 12, no. 4: 1133. https://doi.org/10.3390/w12041133
APA StyleŻur, J., Michalska, J., Piński, A., Mrozik, A., & Nowak, A. (2020). Effects of Low Concentration of Selected Analgesics and Successive Bioaugmentation of the Activated Sludge on Its Activity and Metabolic Diversity. Water, 12(4), 1133. https://doi.org/10.3390/w12041133