
Evidence of Quorum Sensing in Cyanobacteria by Homoserine Lactones: The Origin of Blooms


Abstract
:1. Introduction
2. Experimental
2.1. Microcystis aeruginosa and Cylindrospermopsis raciborskii Cultures
2.2. Cell Counts
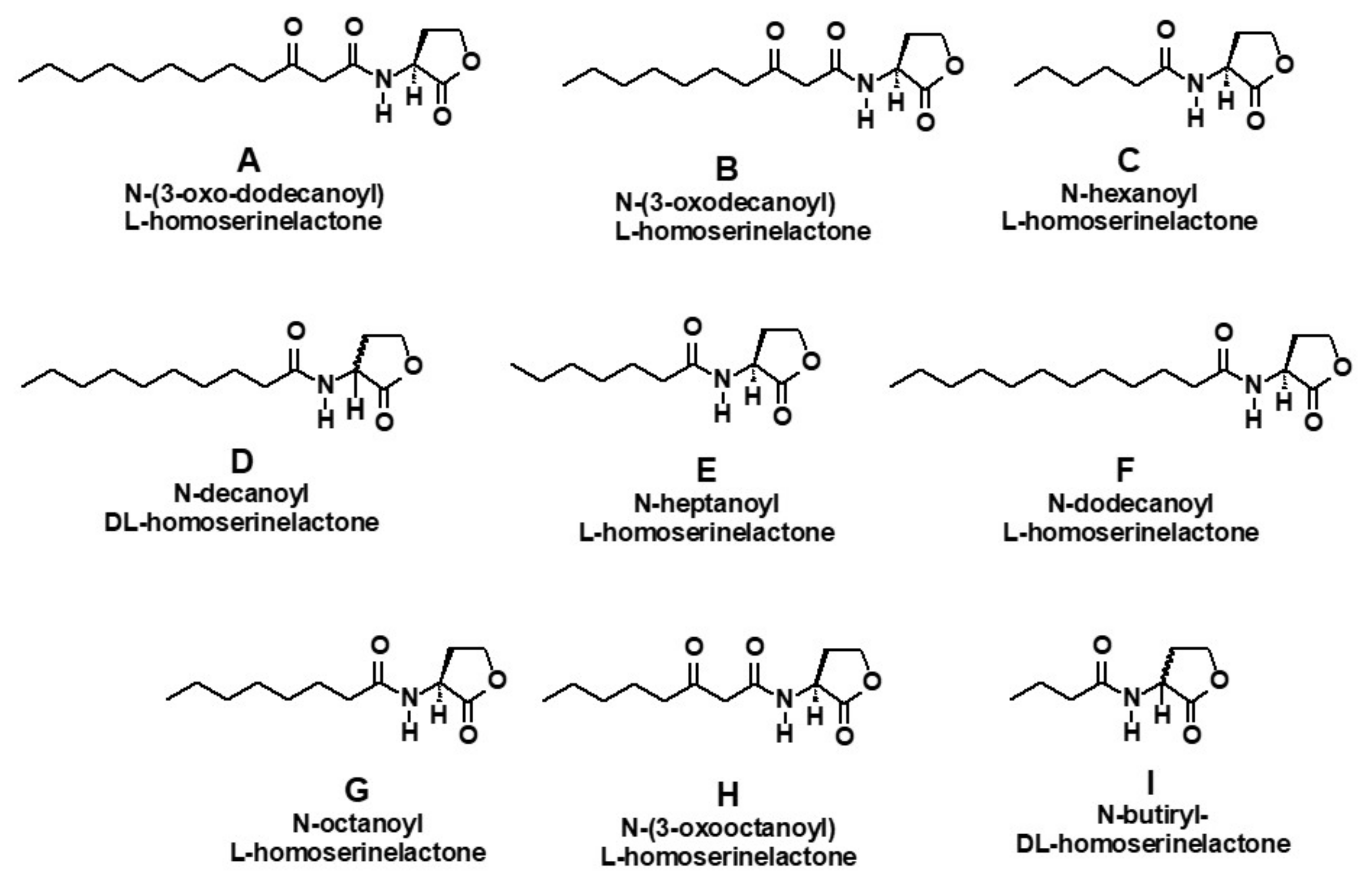
2.3. Inducer Lactones
2.4. Effect of Substances
2.4.1. Effect of Lactones on Cell Proliferation and Colony Formation of M. aeruginosa
2.4.2. Effect of Vanillin on Induction of M. aeruginosa Growth with Four Lactones
2.4.3. Effect of Lactones on Cell Proliferation of Cylindrospermopsis raciborskii
2.5. Spectrometric Readings
2.6. M. aeruginosa Cell and Colony Morphology
2.7. Analysis of Microcystins by ELISA
2.8. Analysis of Saxitoxin by ELISA
2.9. Data Analysis
3. Results
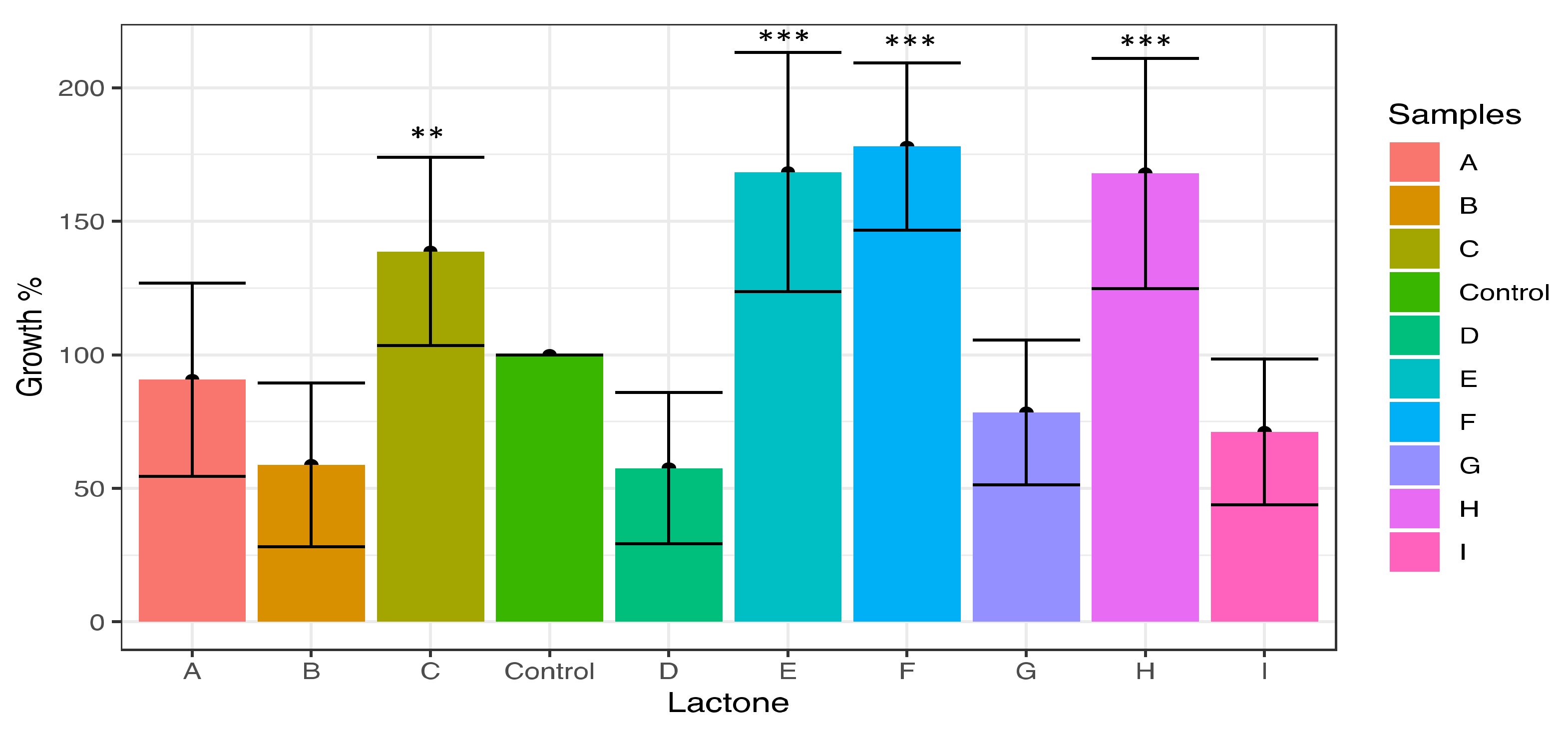
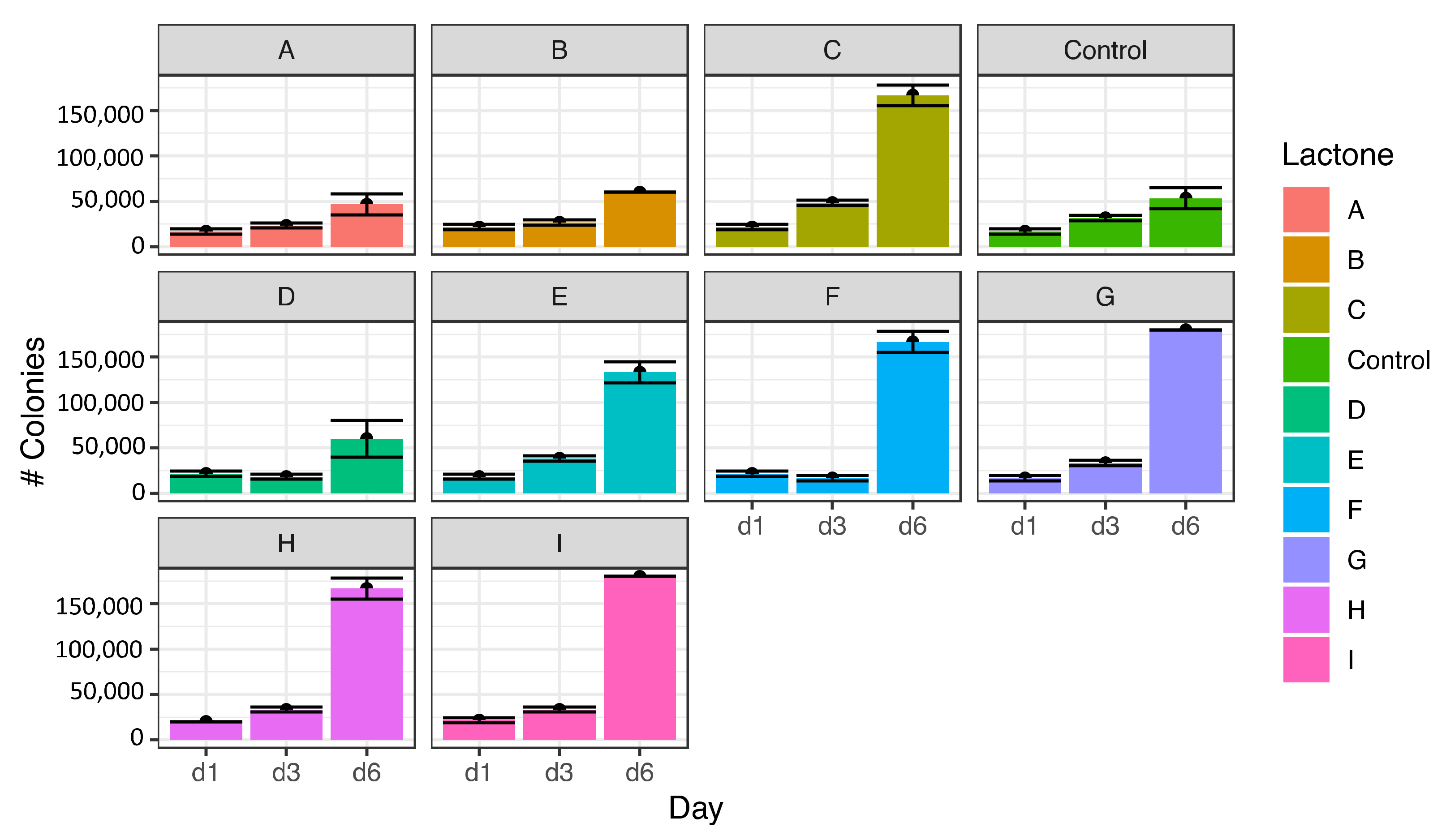
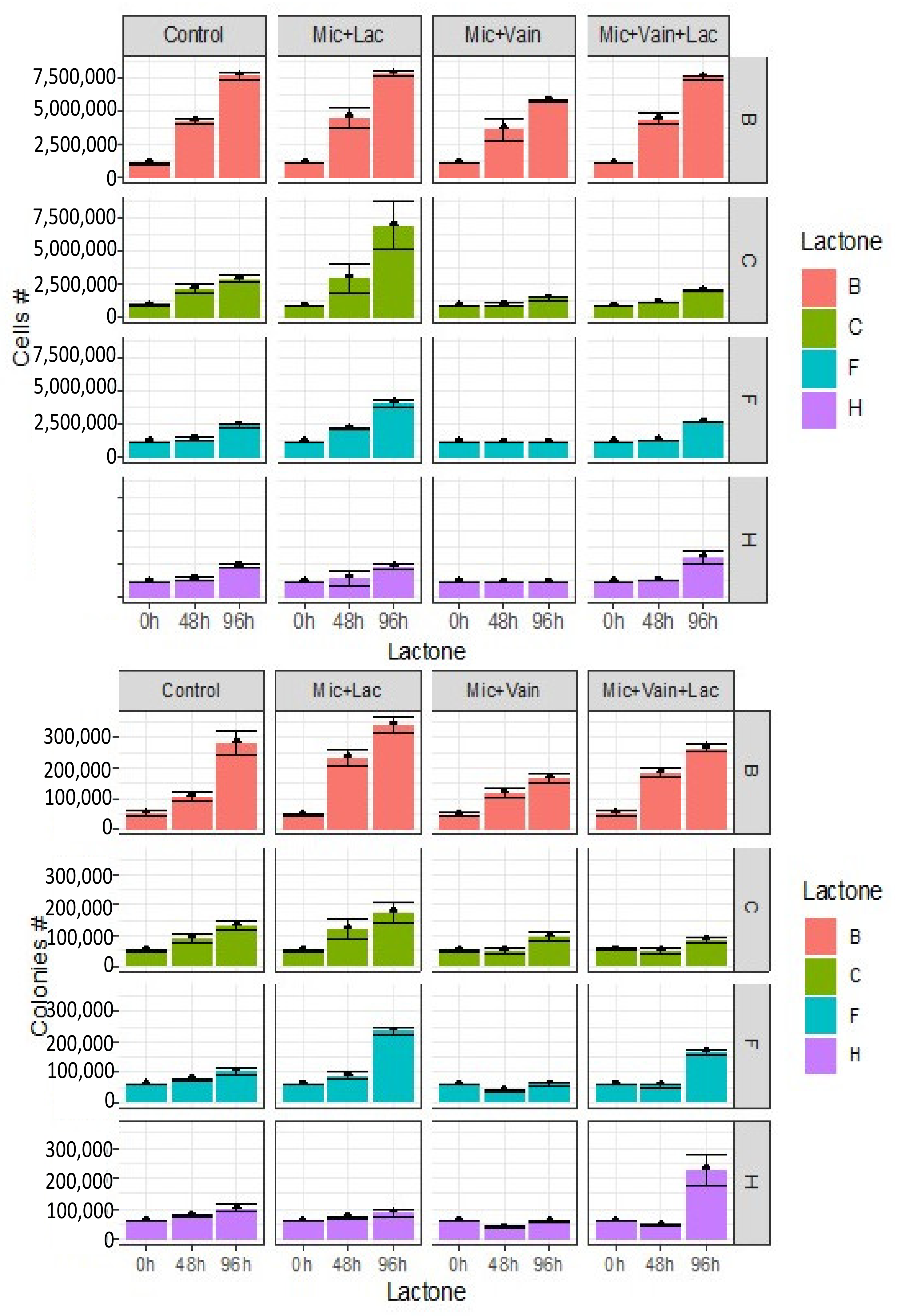
3.1. Effect of Lactones on Cell Proliferation of Microcystis aeruginosa
3.2. Effect of Lactones on the Induction of Colony Formation of Microcystis aeruginosa
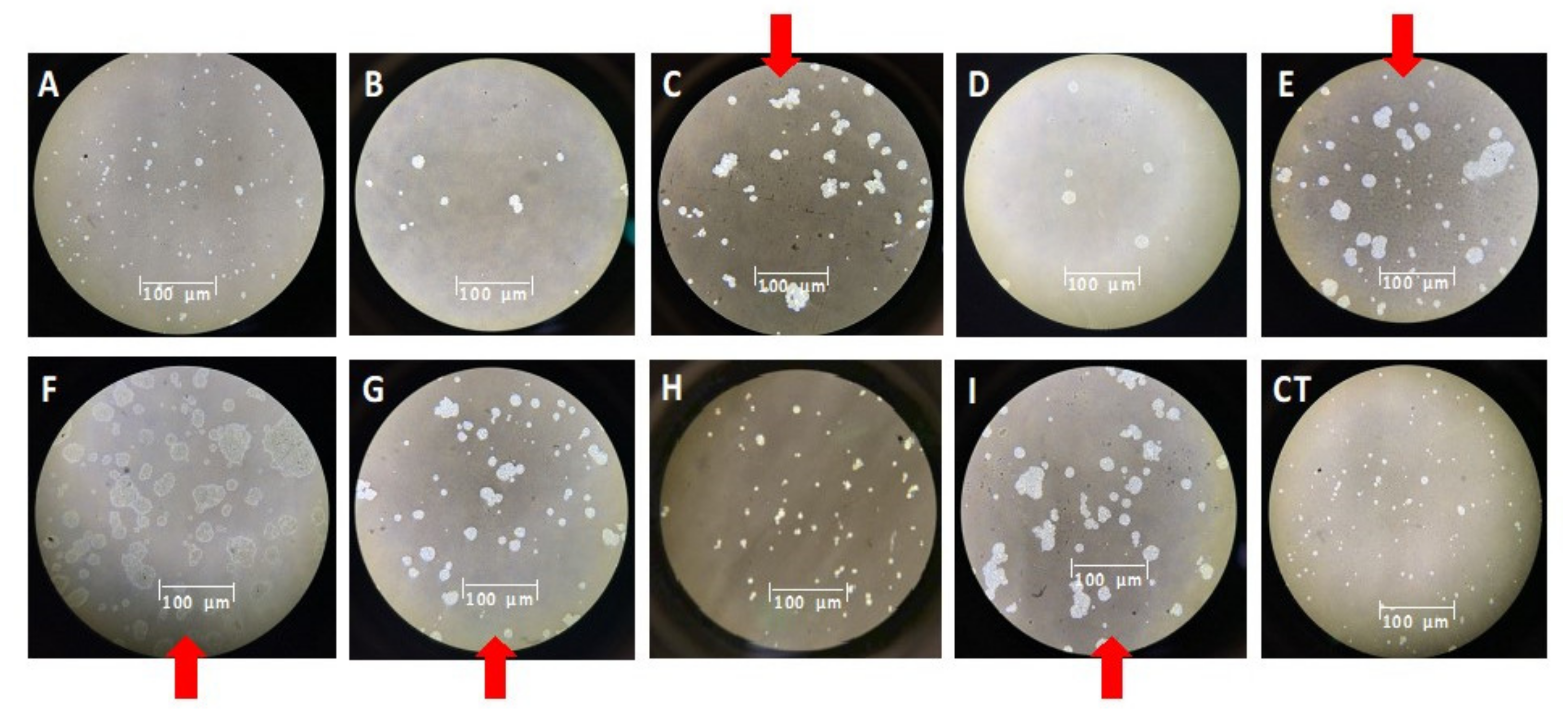
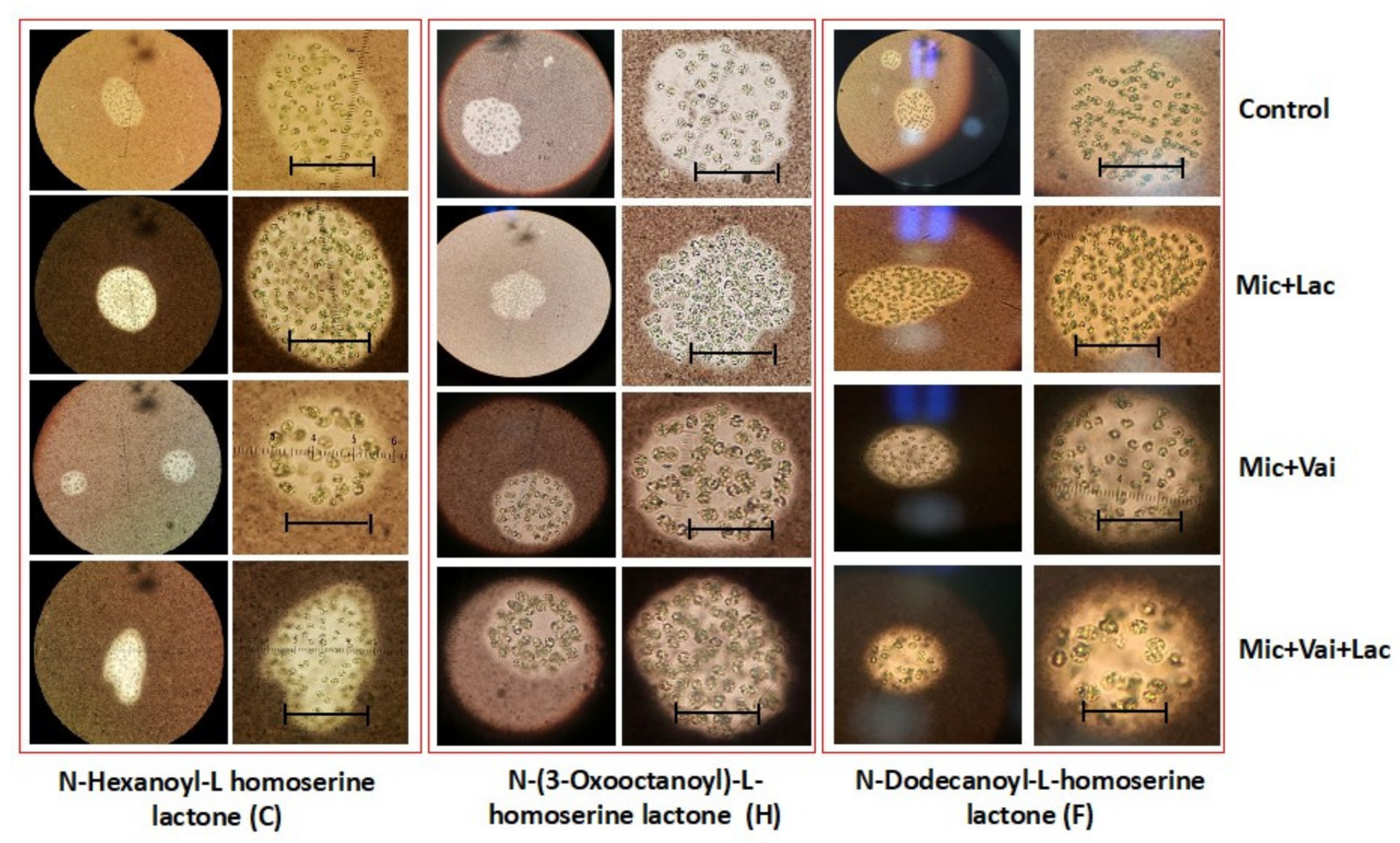
3.3. Effect of Lactones on Colony Morphology of M. aeruginosa
3.4. Effect of Vanillin in the Induction of Cell Proliferation and Colony Formation of M. aeruginosa by Lactones
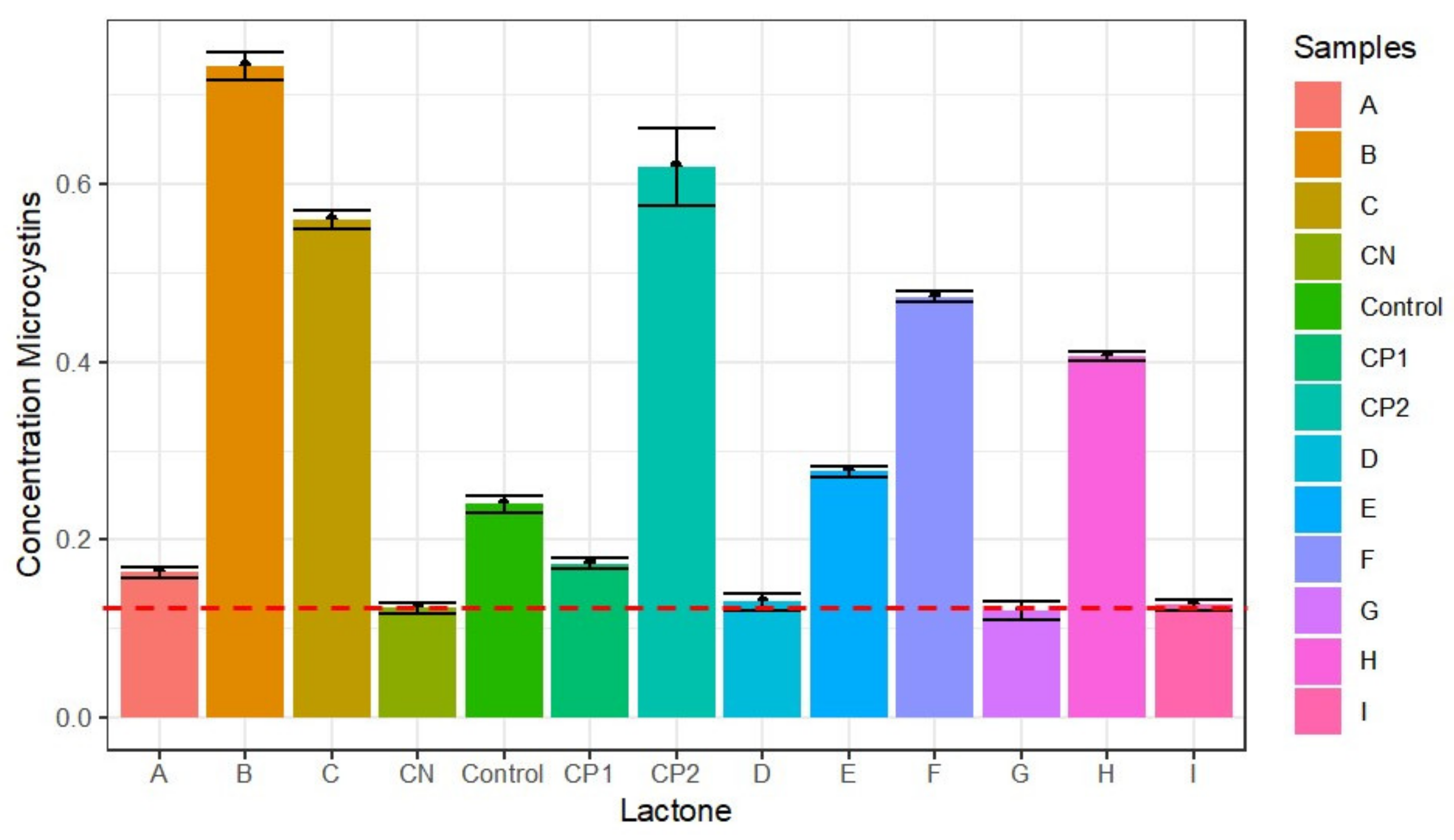
3.5. Effect of Lactones on Microcystin Toxins Production in M. aeruginosa
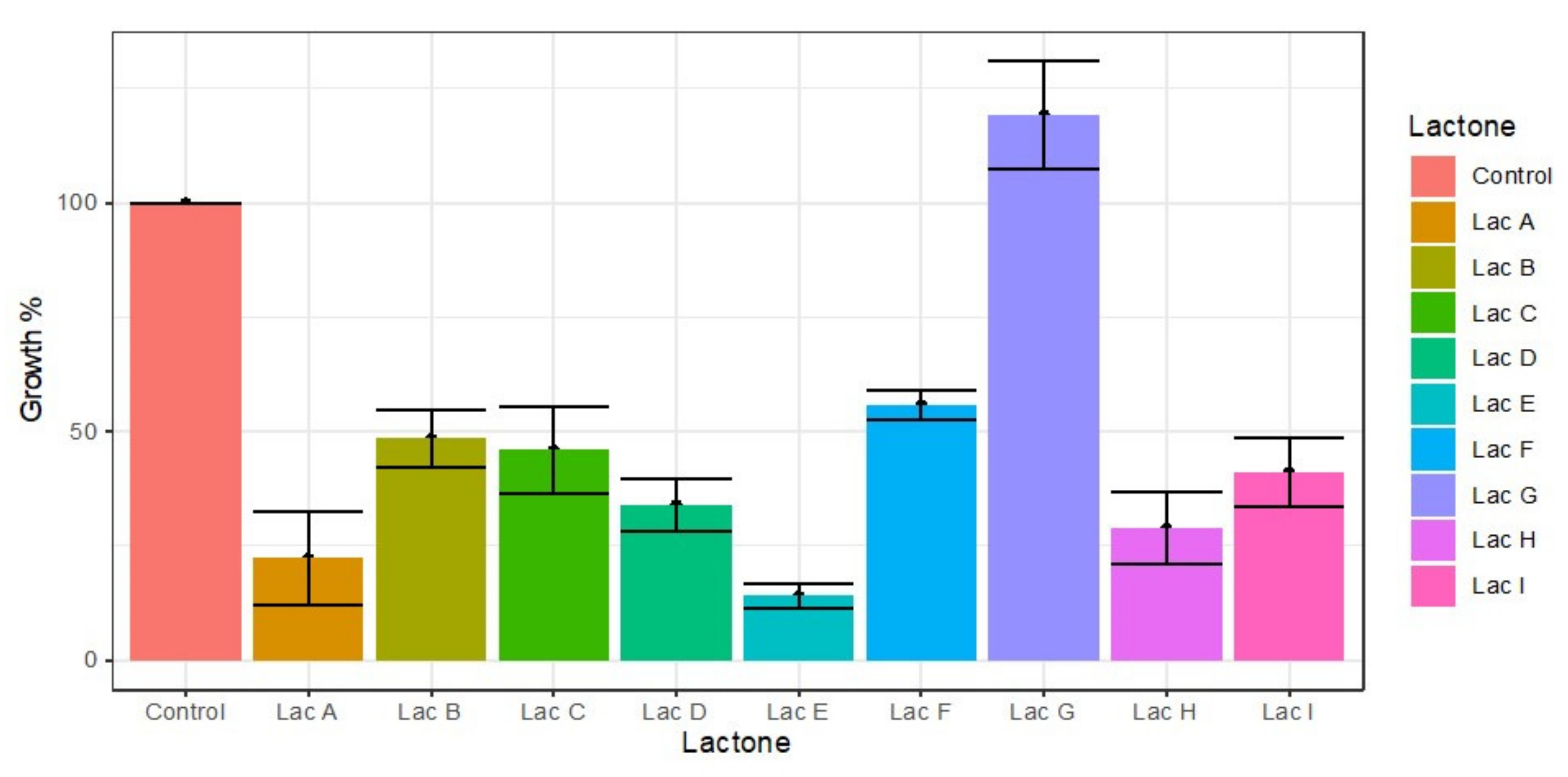
3.6. Induction of Cell Proliferation of Cylindrospermopsis raciborskii
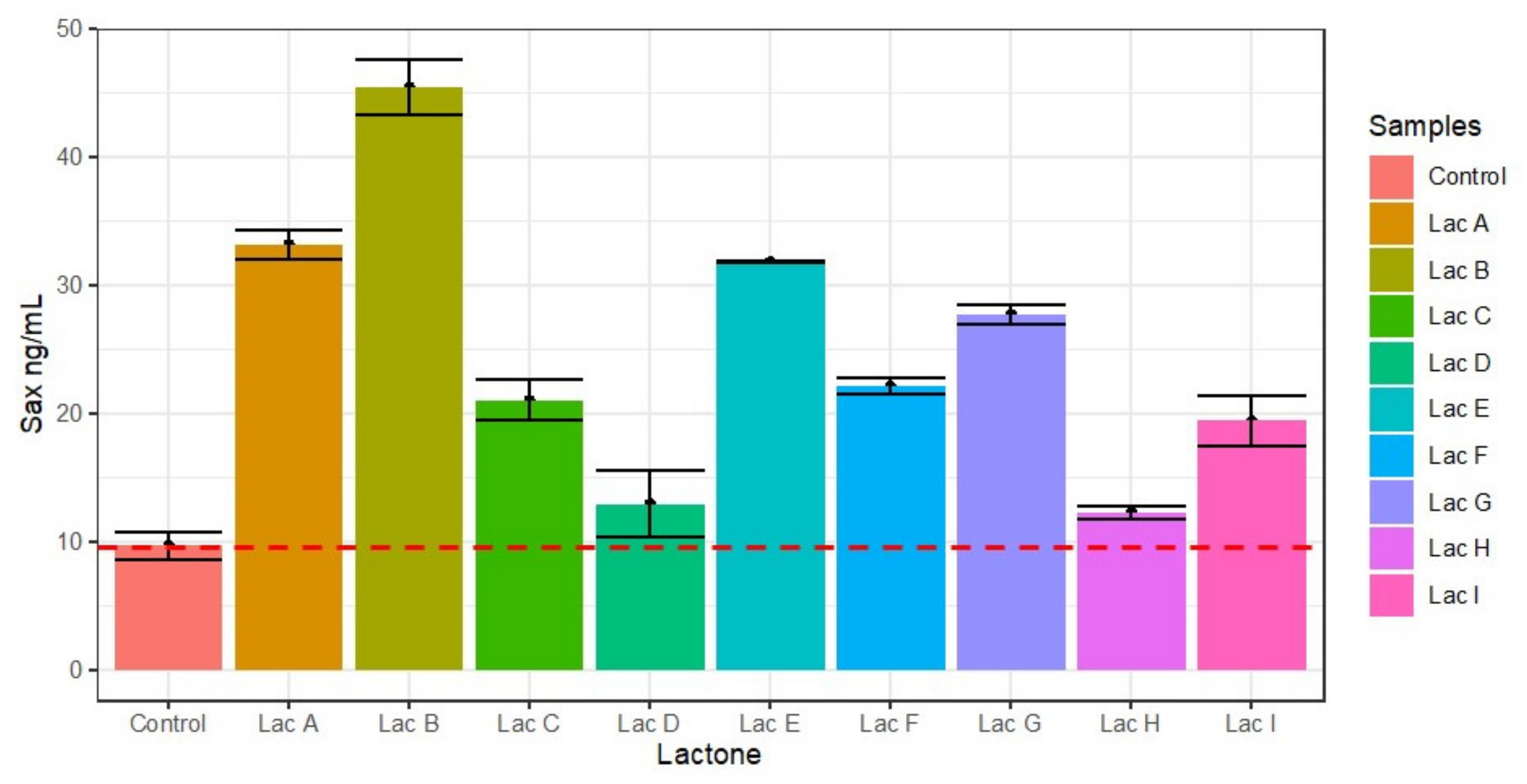
3.7. Effect of Lactones on Saxitoxin Production of Cylindrospermopsis raciborskii
4. Discussion
- In this regard, it has been said that C. raciborskii induces the formation of colonies in M. aeruginosa and inhibits their growth [25], confirming cyanobacterial communication. Experiments are currently underway to determine the production of lactones M. aeruginosa and C. raciborskii.
- There was a specific and selective response. Thus, each lactone appears to be involved in a quorum sensing process. This fact could explain why a mixture of lactones is produced. Similarly, there could be selectivity within the same cyanobacteria, as the effects found were different in M. aeruginosa and C. raciborskii.
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ding, L.; Li, K.; Huang, H.; Hu, H.; Geng, J.; Xu, K.; Ren, H. Estimation of spatial distribution of quorum sensing signaling in sequencing batch biofilm reactor (SBBR) biofilms. Sci. Total Environ. 2018, 612, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2021. [Google Scholar]
- Buratti, F.M.; Manganelli, M.; Vichi, S.; Stefanelli, V.; Scardala, S.; Testai, E.; Funari, E. Cyanotoxins: Producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation. Arch. Toxicol. 2017, 91, 1049–1130. [Google Scholar] [CrossRef] [PubMed]
- Loise, S.; Calado, D.M.; Souza, G.; Pires, T.; Leite, B.; Cristina, H.; Assis, S. Depuration time and sublethal effects of microcystins in a freshwater fi sh from water supply reservoir. Chemosphere 2018, 210, 805–815. [Google Scholar] [CrossRef]
- Gurbuz, F.; Uzunmehmetoğlu, O.Y.; Diler, Ö.; Metcalf, J.S.; Codd, G.A. Occurrence of microcystins in water, bloom, sediment and fish from a public water supply. Sci. Total Environ. 2016, 562, 860–868. [Google Scholar] [CrossRef] [PubMed]
- Lejars, M.; Hajnsdorf, E. Antisense RNA world in bacteria. Biochim. Biophys. Acta 2020, 1863, 194489. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.A. Algal Culturing Techniques; Elsevier/Academic Press: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Hoffmann, L.; Komárek, J.; Kaštovský, J. System of cyanoprokaryote (cyanobacteria)—State in 2004. Algol. Stud. 2005, 117, 95–115. [Google Scholar] [CrossRef]
- Papenfort, K.; Bassler, B. Quorum-Sensing Signal-Response Systems in Gram-Negative Bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef]
- Herrera, N.; Florez, M.T.; Velasquez, J.P.; Echeverri, F. Effect of Phenyl-Acyl Compounds on the Growth, Morphology, and Toxin Production of Microcystis aeruginosa Kützing. Water 2019, 11, 236. [Google Scholar] [CrossRef] [Green Version]
- Dos S Vieira, J.M.; de P Azevedo, M.T.; de Oliveira Azevedo, S.M.F.; Honda, R.Y.; Corrêa, B. Microcystin production by Radiocystis fernandoi (Chroococcales, Cyanobacteria) isolated from a drinking water reservoir in the city of Belém, PA, Brazilian Amazonia region. Toxicon 2003, 42, 709–713. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. 2020. Available online: http://www.r-project.org/index.html (accessed on 31 January 2021).
- Osti, J.A.S.; Podduturi, R.; Schlüter, L.; Liu, T.; Moraes, M.D.E.B.; Jorgensen, N.O.G. Monitoring of saxitoxin production in lakes in Denmark by molecular, chromatographic and microscopic approaches. Harmful Algae 2021, 101, 101966. [Google Scholar] [CrossRef]
- Hofer, U. Climate change boosts cyanobacteria. Nat. Rev. Microbiol. 2018, 16, 122–123. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- Moe, S.J.; Haande, S.; Couture, R.-M. Climate change, cyanobacteria blooms and ecological status of lakes: A Bayesian network approach. Ecol. Model. 2016, 337, 330–347. [Google Scholar] [CrossRef] [Green Version]
- Zhai, C.; Zhang, P.; Shen, F.; Zhou, C.; Liu, C. Does Microcystis aeruginosa have quorum sensing? FEMS Microbiol. Lett. 2012, 336, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif, D.I.; Gallon, J.; Smith, C.J.; Dudley, E. Quorum sensing in Cyanobacteria: N -octanoyl-homoserine lactone release and response, by the epilithic colonial cyanobacterium Gloeothece PCC6909. ISME J. 2008, 2, 1171–1182. [Google Scholar] [CrossRef] [Green Version]
- Van Mooy, B.A.S.; Hmelo, L.R.; Sofen, L.E.; Campagna, S.R.; May, A.L.; Dyhrman, S.T.; Heithoff, A.; Webb, E.A.; Momper, L.; Mincer, T.J. Quorum sensing control of phosphorus acquisition in Trichodesmium consortia. ISME J. 2011, 6, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Romero, M.; Muro-pastor, A.M.; Otero, A. Quorum sensing N-acylhomoserine lactone signals affect nitrogen fixation in the cyanobacterium Anabaena sp. PCC7120. FEMS Microbiol. Lett. 2011, 315, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Braun, E.; Bachofen, R. Homoserine-lactones and microcystin in cyanobacterial assemblages in Swiss lakes. Hydrobiologia 2004, 522, 280. [Google Scholar] [CrossRef]
- Gautam, K.; Tripathi, J.K.; Pareek, A.; Sharma, D.K. Growth and secretome analysis of possible synergistic interaction between green algae and cyanobacteria. J. Biosci. Bioeng. 2019, 127, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhu, J.; Cai, Z.; Lao, Y.; Jin, H.; Yu, K. Profiles of quorum sensing (QS)-related sequences in phycospheric microorganisms during a marine dinoflagellate bloom, as determined by a metagenomic approach. Microbiol. Res. 2018, 217, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mello, M.M.E.; Soares, M.C.S.; Roland, F.; Lürling, M. Growth inhibition and colony formation in the cyanobacterium Microcystis aeruginosa induced by the cyanobacterium Cylindrospermopsis raciborskii. J. Plankton Res. 2012, 34, 987–994. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lactone/# Carbon Atoms | M. aeruginosa | C. raciborskii Metabolic Effect | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Metabolic Effect | Effect of Vanillin | ||||||||||
| Cell Proliferation | Colonies Formation | ||||||||||
| Cell Prol | Col Form | Microc | Lact a | Van b | Lact + Van c | Lac a | Van b | Lac+ Van c | Cell Prol | Saxit | |
| Butiryl (I), C4 | - | ++++ | - | NA | NA | NA | NA | NA | NA | - - | ++ |
| Hexanoyl (C), C6 | ++ | ++++ | +++ | ++++ | WE | WE | + | - | - | - - | ++ |
| Heptanoyl (E), C7 | ++++ | ++++ | + | NA | NA | NA | NA | NA | NA | - - - - | +++ |
| Octanoyl (G), C8 | - | ++++ | - | NA | NA | NA | NA | NA | NA | ++ | +++ |
| N-(3-Oxooctanoyl) (H), C10 | +++ | ++++ | ++ | WE | - | WE | WE | WE | +++ | - - - | + |
| Decanoyl (D), C10 | - - | WE | - | NA | NA | NA | NA | NA | NA | - - | + |
| N-(3-Oxodecanoyl (A), C10 | - | WE | - - | NA | NA | NA | NA | NA | NA | - - - - | +++ |
| N-Dodecanoyl (F), C12 | ++++ | ++++ | +++ | ++ | WE | + | +++ | - - | - | - - | ++ |
| N-(3-Oxododecanoyl) (B), C12 | - - | WE | ++++ | WE | - | WE | + | - - - | - | - - | ++++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrera, N.; Echeverri, F. Evidence of Quorum Sensing in Cyanobacteria by Homoserine Lactones: The Origin of Blooms. Water 2021, 13, 1831. https://doi.org/10.3390/w13131831
Herrera N, Echeverri F. Evidence of Quorum Sensing in Cyanobacteria by Homoserine Lactones: The Origin of Blooms. Water. 2021; 13(13):1831. https://doi.org/10.3390/w13131831
Chicago/Turabian StyleHerrera, Natalia, and Fernando Echeverri. 2021. "Evidence of Quorum Sensing in Cyanobacteria by Homoserine Lactones: The Origin of Blooms" Water 13, no. 13: 1831. https://doi.org/10.3390/w13131831
APA StyleHerrera, N., & Echeverri, F. (2021). Evidence of Quorum Sensing in Cyanobacteria by Homoserine Lactones: The Origin of Blooms. Water, 13(13), 1831. https://doi.org/10.3390/w13131831