
Environmental Flows as a Proactive Tool to Mitigate the Impacts of Climate Warming on Freshwater Macroinvertebrates




Abstract
:1. Introduction
2. Materials and Methods
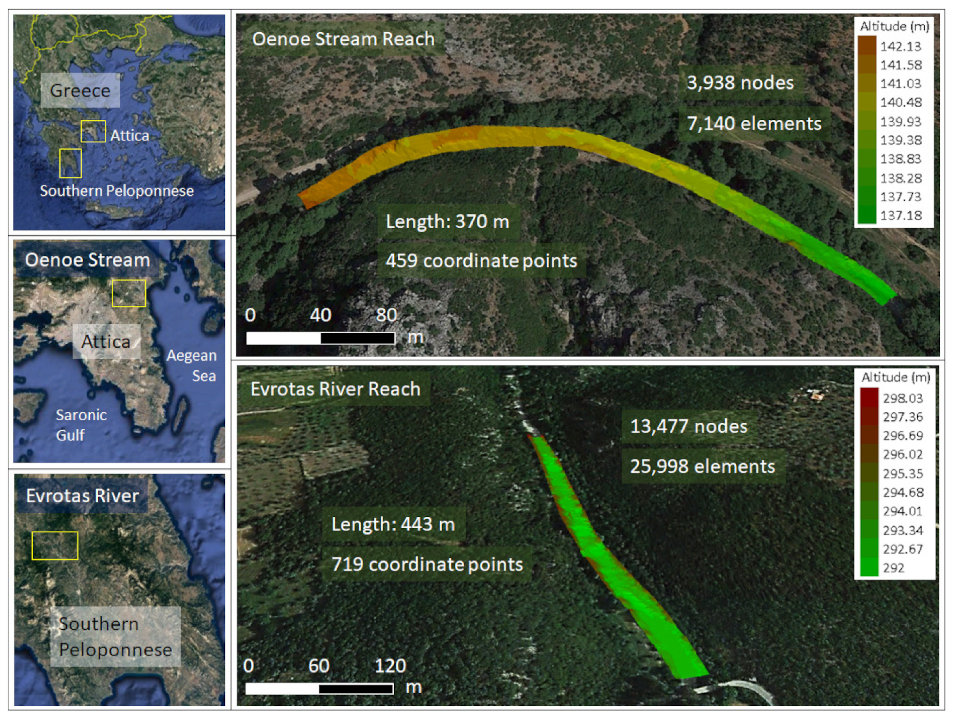
2.1. Case Studies
2.2. Climatic Data and Climate Change Scenarios
2.3. Hydraulic Models
2.4. Habitat Suitability Curves and Habitat Suitability Models
2.5. Reach-Scale Habitat Suitability and Comparisons between Scenarios
3. Results
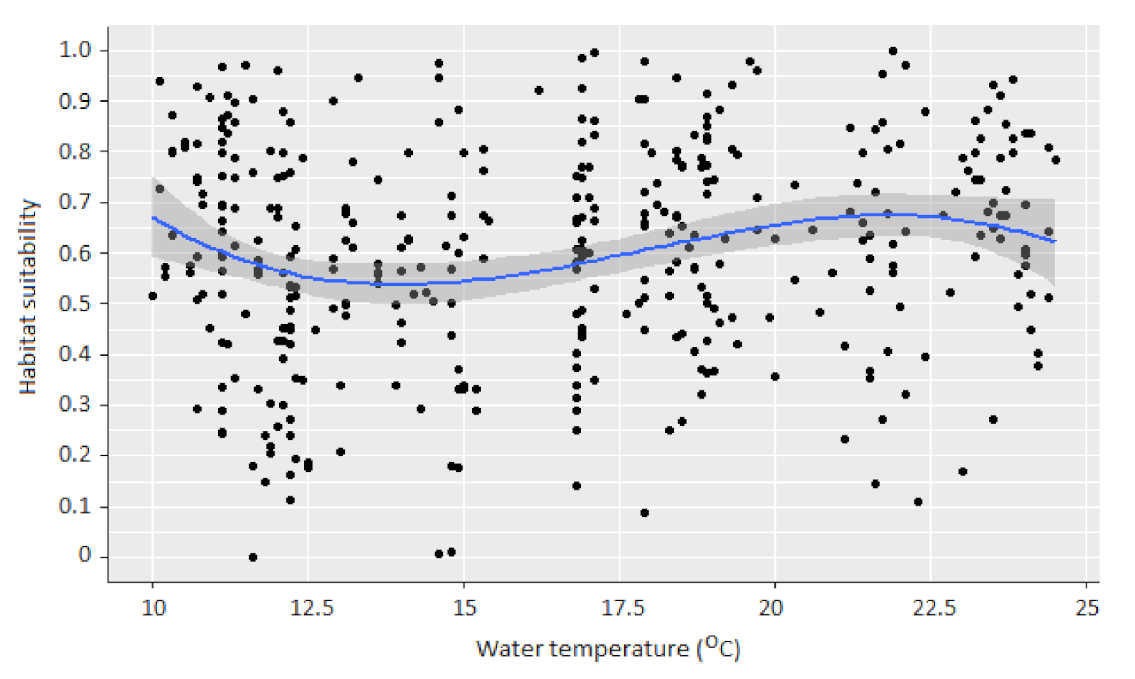
3.1. Water Temperature and Macroinvertebrate Habitat Suitability
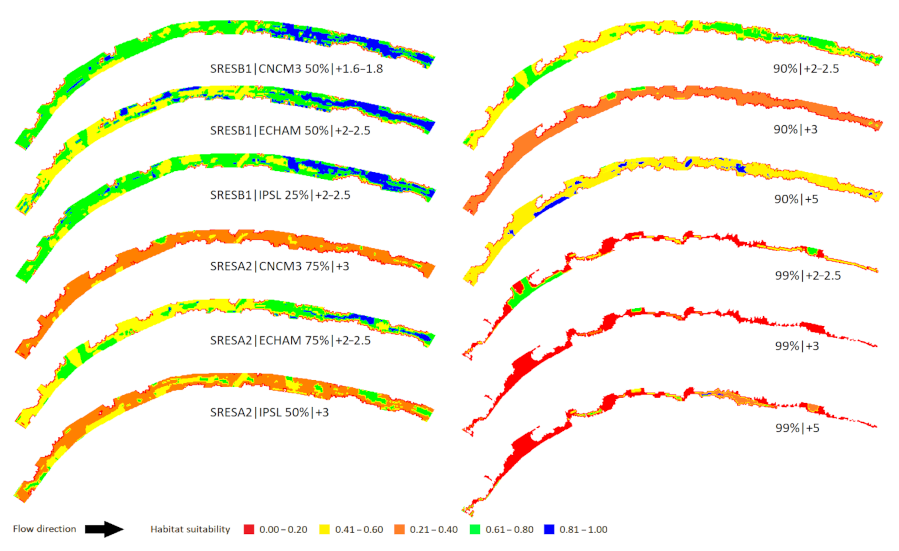
3.2. Case Study I: The Oenoe Stream (a Losing Watershed)
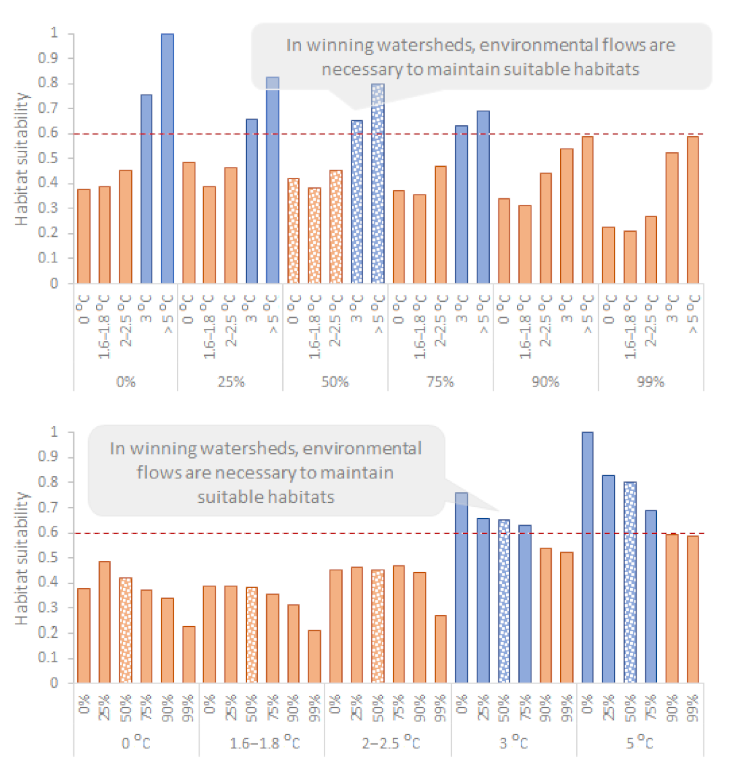
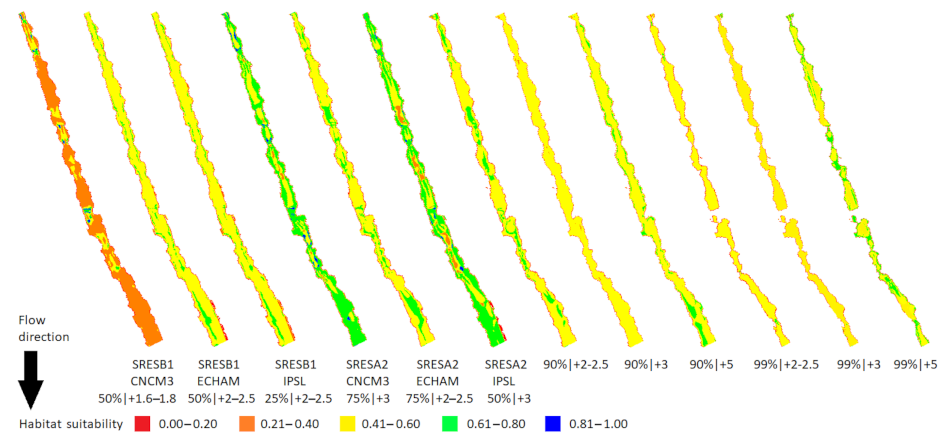
3.3. Case Study II: The Evrotas River (a Winning Watershed)
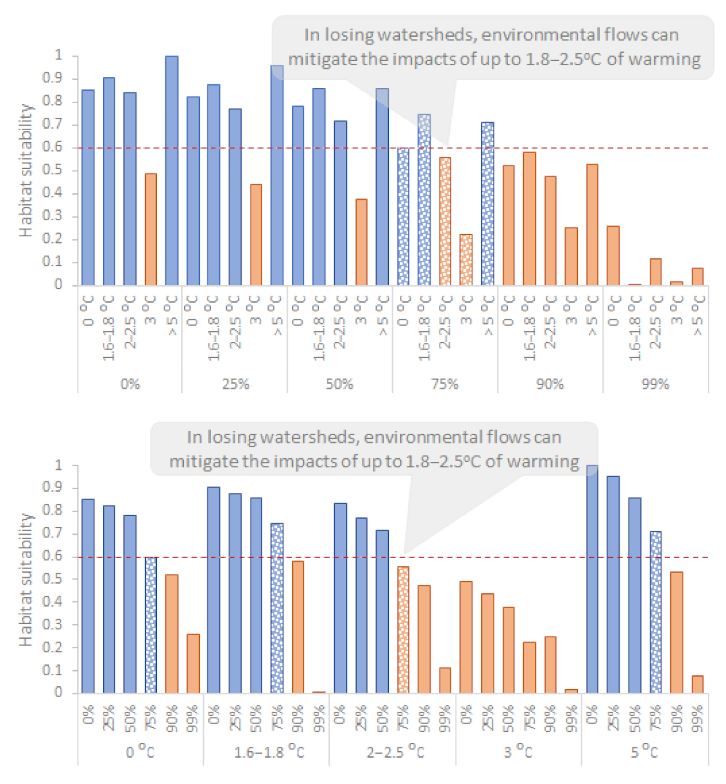
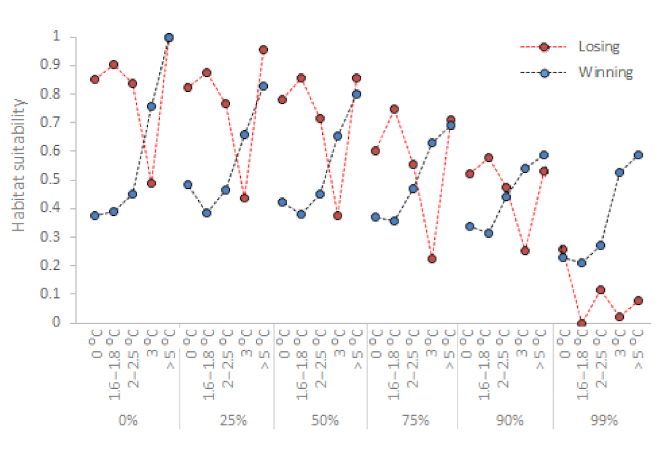
3.4. Losing vs. Winning Watersheds
4. Discussion
4.1. In Losing Watersheds, Environmental Flows Can Mitigate the Impacts of up to 1.8–2.5 °C of Warming
4.2. In Winning Watersheds, Environmental Flows Are Necessary to Maintain Suitable Habitats
4.3. Securing Environmental Flows Builds the Capacity of the Ecosystem to Better Adapt to Impacts from Climate Change
4.4. Limitations and Future Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1: T ≤ 18.96 °C | Group 2: T ≥ 19.96 °C | |||||
|---|---|---|---|---|---|---|
| Taxa | Average abundance | Average abundance | Average dissimilarity | Dissimilarity/SD | Contribution (%) | Cumulative contribution (%) |
| Baetidae | 24.84 | 7.6 | 16.9 | 1.01 | 20.83 | 20.83 |
| Leuctridae | 4.47 | 16.99 | 11.27 | 0.9 | 13.9 | 34.73 |
| Chironomidae | 14.22 | 7.1 | 10.81 | 0.82 | 13.33 | 48.06 |
| Caenidae | 0.83 | 11.58 | 6.63 | 0.79 | 8.18 | 56.23 |
| Gomphidae | 0.16 | 5.33 | 4.74 | 0.5 | 5.84 | 62.08 |
| Heptageniidae | 5.17 | 1.42 | 4.58 | 0.67 | 5.65 | 67.73 |
| Hydropsychidae | 2.87 | 4.88 | 4.24 | 0.78 | 5.23 | 72.95 |
| Elmidae | 1.84 | 2.42 | 2.65 | 0.69 | 3.27 | 76.22 |
| Hydraenidae | 1.98 | 2.13 | 2.33 | 0.54 | 2.87 | 79.09 |
| Ephemerellidae | 2.82 | 0.35 | 2.23 | 0.54 | 2.75 | 81.84 |
| Perlidae | 1.75 | 0.5 | 1.52 | 0.5 | 1.87 | 83.71 |
| Hydroptilidae | 0.18 | 1.4 | 1.29 | 0.53 | 1.59 | 85.3 |
| Oligochaeta | 0.11 | 1.08 | 1.23 | 0.4 | 1.51 | 86.81 |
| Simuliidae | 1.58 | 0.36 | 1.11 | 0.34 | 1.37 | 88.18 |
| Hydracarina | 0.27 | 1.35 | 1.06 | 0.48 | 1.31 | 89.49 |
| Athericidae | 0.9 | 0.44 | 1.05 | 0.51 | 1.29 | 90.78 |
References
- Houghton, J. Global Warming: The Complete Briefing, 4th ed.; Cambridge University Press: Cambridge, UK, 2009. [Google Scholar]
- Collins, M.; Knutti, R.; Arblaster, J.; Dufresne, J.L.; Fichefet, T.; Friedlingstein, P.; Gao, X.; Gutowski, W.J.; Johns, T.; Krinner, G.; et al. Long-term Climate Change: Projections, Commitments and Irreversibility. In Climate Change 2013: The Physical Science Basis; Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate, Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: New York, NY, USA, 2013; pp. 1029–1136. [Google Scholar]
- Trenberth, K.E. The impact of climate change and variability on heavy precipitation, floods, and droughts. In Encyclopedia of Hydrological Sciences; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2006. [Google Scholar] [CrossRef] [Green Version]
- Milly, P.C.D.; Dunne, K.A.; Vecchia, A.V. Global pattern of trends in streamflow and water availability in a changing climate. Nature 2005, 438, 347–350. [Google Scholar] [CrossRef] [PubMed]
- Erol, A.; Randhir, T.O. Climatic change impacts on the ecohydrology of Mediterranean watersheds. Clim. Chang. 2012, 114, 319–341. [Google Scholar] [CrossRef]
- Stahl, K.; Tallaksen, L.M.; Hannaford, J.; Van Lanen, H.A.J. Filling the white space on maps of European runoff trends: Estimates from a multi-model ensemble. Hydrol. Earth Syst. Sci. 2012, 16, 2035–2047. [Google Scholar] [CrossRef] [Green Version]
- Van Vliet, M.T.H.; Franssen, W.H.P.; Yearsley, J.R.; Ludwig, F.; Haddeland, I.; Lettenmaier, D.P.; Kabat, P. Global river discharge and water temperature under climate change. Glob. Environ. Chang. 2013, 23, 450–464. [Google Scholar] [CrossRef]
- Pascual, D.; Pla, E.; Lopez-Bustins, J.A.; Retana, J.; Terradas, J. Impacts of climate change on water resources in the Mediterranean Basin: A case study in Catalonia, Spain. Hydrol. Sci. J. 2015, 60, 2132–2147. [Google Scholar] [CrossRef] [Green Version]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldgerb, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef]
- Hogg, I.D.; Williams, D.D. Response of stream invertebrates to a global-warming thermal regime: An ecosystem-level manipulation. Ecology 1996, 77, 395–407. [Google Scholar] [CrossRef] [Green Version]
- USEPA (US Environmental Protection Agency). Climate Change Effects on Stream and River Biological Indicators: A Preliminary Analysis; EPA/600/R-07/085; Global Change Research Program, National Center for Environmental Assessment: Washington DC, USA, 2008. [Google Scholar]
- Jourdan, J.; O’Hara, R.B.; Bottarin, R.; Huttunen, K.L.; Kuemmerlen, M.; Monteith, D.; Muotka, T.; Ozoliņš, D.; Paavola, R.; Pilotto, F.; et al. Effects of changing climate on European stream invertebrate communities: A long-term data analysis. Sci. Total Environ. 2018, 621, 588–599. [Google Scholar] [CrossRef]
- Haase, P.; Pilotto, F.; Fengqing, L.; Sundermann, A.; Lorenz, A.W.; Tonkin, J.D.; Stoll, S. Moderate warming over the past 25 years has already reorganized stream invertebrate communities. Sci. Total Environ. 2019, 658, 1531–1538. [Google Scholar] [CrossRef] [PubMed]
- Baranov, V.; Jourdan, J.; Pilotto, F.; Wagner, R.; Haase, P. Complex and nonlinear climate-driven changes in freshwater insect communities over 42 years. Conserv. Biol. 2020, 34, 1241–1251. [Google Scholar] [CrossRef]
- Forslund, A. Securing Water for Ecosystems and Human Well-Being: The Importance of Environmental Flows; Swedish Water House Report 24; Swedish Water House: Stockholm, Sweden, 2009. [Google Scholar]
- Theodoropoulos, C.; Karaouzas, I. Climate change and the future of mediterranean freshwater macroinvertebrates. Hydrobiologia 2021, in press. [Google Scholar] [CrossRef]
- Nakicenovic, N.; Alcamo, J.; Davis, G.; de Vries, H.J.M.; Fenhann, J.; Gaffin, S.; Gregory, K.; Grubler, A.; Jung, T.Y.; Kram, T.; et al. Emissions Scenarios: A Special Report of Working Group III of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Theodoropoulos, C.; Georgalas, S.; Mamassis, N.; Stamou, A.; Rutschmann, P.; Skoulikidis, N. Comparing environmental flow scenarios from hydrological methods, legislation guidelines, and hydrodynamic habitat models downstream of the Marathon Dam (Attica, Greece). Ecohydrology 2018, 11, e2019. [Google Scholar] [CrossRef]
- Theodoropoulos, C.; Papadaki, C.; Vardakas, L.; Dimitriou, E.; Kalogianni, E.; Skoulikidis, N. Conceptualization and pilot application of a model-based environmental flow assessment adapted for intermittent rivers. Aquat. Sci. 2019, 81, 10. [Google Scholar] [CrossRef] [Green Version]
- Hervouet, J.M. Hydrodynamics of Free Surface Flows: Modelling with the Finite Element Method; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2007. [Google Scholar]
- Theodoropoulos, C.; Vourka, A.; Skoulikidis, N.; Rutschmann, P.; Stamou, A. Evaluating the performance of habitat models for predicting the environmental flow requirements of benthic macroinvertebrates. J. Ecohydraulics 2018, 3, 30–44. [Google Scholar] [CrossRef]
- Theodoropoulos, C.; Skoulikidis, N.; Rutschmann, P.; Stamou, A. Ecosystem-based environmental flow assessment in a Greek regulated river with the use of 2D hydrodynamic habitat modelling. River Res. Appl. 2018, 34, 538–547. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 8 September 2021).
- Stasinopoulos, D.M.; Rigby, R.A. Generalized additive models for location, scale, and shape (GAMLSS) in R. J. Stat. Softw. 2007, 23, 1–46. [Google Scholar] [CrossRef] [Green Version]
- Kooperberg, C. Package ‘logspline’ v2.1.9. Available online: https://cran.r-project.org/web/packages/logspline/index.html (accessed on 8 September 2021).
- Zhang, D. Package ‘rsq’. R-Squared and Related Measures. Available online: https://cran.r-project.org/web/packages/rsq/rsq.pdf (accessed on 8 September 2021).
- Theodoropoulos, C.; Skoulikidis, N.; Stamou, A. HABFUZZ: A tool to calculate the instream hydraulic habitat suitability using fuzzy logic and fuzzy Bayesian inference. J. Open Source Softw. 2016, 1, 82. [Google Scholar] [CrossRef]
- Van Broekhoven, E.; Adriaenssens, V.; De Baets, B.; Verdonschot, P.F.M. Fuzzy rule-based macroinvertebrate habitat suitability models for running waters. Ecol. Model. 2006, 198, 71–84. [Google Scholar] [CrossRef]
- Marziali, L.; Rossaro, B. Response of chironomid species (Diptera, Chironomidae) to water temperature: Effects on species distribution in specific habitats. J. Entomol. Acarol. Res. 2013, 45, 73–89. [Google Scholar] [CrossRef] [Green Version]
- Suter, P.J. Baetidae. In Mayfly Nymphs of Australia. A guide to Genera; Identification Guide No. 7; Dean, J.C., Suter, P.J., Eds.; Cooperative Research Centre for Freshwater Ecology: Albury, NSW, Australia; pp. 13–28.
- Zuellig, R.E.; Kondratieff, B.C.; Ruiter, D.E.; Thorp, R.A. An Annotated List of the Mayflies, Stoneflies, and Caddisflies of the Sand Creek Basin, Great Sand Dunes National Park and Preserve, Colorado, 2004 and 2005. USGS Data series 183. Available online: https://pubs.usgs.gov/ds/ds183/#heptageniidae (accessed on 23 June 2021).
- Elliott, J.M. Effect of temperature on the hatching time of eggs of Ephemerella ignita (Poda) (Ephemeroptera: Ephemerellidae). Freshw. Biol. 1978, 8, 51–58. [Google Scholar] [CrossRef]
- Puckett, R.T.; Cook, J.L. Physiological tolerance ranges of larval Caenis latipennis (Ephemeroptera: Caenidae) in response to fluctuations in dissolved oxygen concentration, pH and temperature. Tex. J. Sci. 2004, 56, 123–130. [Google Scholar]
- Braune, E.; Richter, O.; Söndgerath, D.; Suhling, F. Voltinism flexibility of a riverine dragonfly along thermal gradients. Glob. Chang. Biol. 2008, 14, 470–482. [Google Scholar] [CrossRef]
- Dallas, H. The Effect of Water Temperature on Aquatic Organisms: A Review of Knowledge and Methods for Assessing Biotic Responses to Temperature; WRC Report No. KV 213/09; The Freshwater Consulting Group Freshwater Research Unit, Department of Zoology, University of Cape Town: Cape Town, South Africa, 2009. [Google Scholar]
- Elliott, J.M. Temperature-induced changes in the life cycle of Leuctra nigra (Plecoptera: Leuctridae) from a Lake District stream. Freshw. Biol. 1987, 18, 177–184. [Google Scholar] [CrossRef]
- Puig, M.A. Distribution and ecology of the stoneflies (Plecoptera) in Catalonian rivers (NE-Spain). Int. J. Limnol. 1984, 20, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Karaouzas, I.; Andriopoulou, A.; Kouvarda, T.; Murányi, D. An annotated checklist of the Greek Stonefly Fauna (Insecta: Plecoptera). Zootaxa 2016, 4111, 301–333. [Google Scholar] [CrossRef] [PubMed]
- Murray-Darling Basin Authority—Delivering Water for the Environment. Available online: https://www.mdba.gov.au/issues-murray-darling-basin/water-for-environment/delivering-water-environment (accessed on 8 September 2021).
- Bonada, N.; Rieradevall, M.; Prat, N. Macroinvertebrate community structure and biological traits related to flow permanence in a Mediterranean river network. Hydrobiologia 2007, 589, 91–106. [Google Scholar] [CrossRef]
- Woodward, G.; Perkins, D.M.; Brown, L.E. Climate change and freshwater ecosystems: Impacts across multiple levels of organization. Philos. Trans. R. Soc. B 2010, 365, 2093–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoks, R.; Geerts, A.N.; De Meester, L. Evolutionary and plastic responses of freshwater invertebrates to climate change: Realized patterns and future potential. Evol. Appl. 2014, 7, 42–55. [Google Scholar] [CrossRef]
- Palmer, M.A.; Lettenmaier, D.P.; Poff, N.L.; Postel, S.L.; Richter, B.; Warner, R. Climate change and river ecosystems: Protection and adaptation options. Environ. Manag. 2009, 44, 1053–1068. [Google Scholar] [CrossRef]







| Climate Change Scenarios for 2071–2100 | Mean Discharge Change for 2071–2100 Compared to Current (%) | Mean Water Temperature Change for 2071–2100 Compared to Current (°C) | Evrotas-Specific Discharge (m3/s) | Evrotas-Specific Temperature (°C) | Oenoe-Specific Discharge (m3/s) | Oenoe-Specific Temperature (°C) |
|---|---|---|---|---|---|---|
| Standard (IPCC; [7,17]) | ||||||
| SRES B1/CNCM3 | 25–50 | 1.6–1.8 | 1.25 | 17.06 | 0.3 | 18.06 |
| SRES B1/ECHAM | 25–50 | 2–2.5 | 1.25 | 17.96 | 0.3 | 18.96 |
| SRES B1/IPSL | 0–25 | 2–2.5 | 1.875 | 17.96 | 0.45 | 18.96 |
| SRES A2/CNCM3 | >50 | 3–5 | 0.625 | 18.96 | 0.15 | 19.96 |
| SRES A2/ECHAM | >50 | 2–2.5 | 0.625 | 17.96 | 0.15 | 18.96 |
| SRES A2/IPSL | 25–50 | 3–5 | 1.25 | 18.96 | 0.3 | 19.96 |
| Extreme scenarios (study-specific) | ||||||
| EXT1 | 90 | 2–2.5 | 0.25 | 17.96 | 0.06 | 18.96 |
| EXT2 | 90 | 3–5 | 0.25 | 18.96 | 0.06 | 19.96 |
| EXT3 | 90 | >5 | 0.25 | 20.46 | 0.06 | 21.46 |
| EXT4 | 99 | 2–2.5 | 0.025 | 17.96 | 0.006 | 18.96 |
| EXT5 | 99 | 3–5 | 0.025 | 18.96 | 0.006 | 19.96 |
| EXT6 | 99 | >5 | 0.025 | 20.46 | 0.006 | 21.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Theodoropoulos, C.; Karaouzas, I.; Stamou, A. Environmental Flows as a Proactive Tool to Mitigate the Impacts of Climate Warming on Freshwater Macroinvertebrates. Water 2021, 13, 2586. https://doi.org/10.3390/w13182586
Theodoropoulos C, Karaouzas I, Stamou A. Environmental Flows as a Proactive Tool to Mitigate the Impacts of Climate Warming on Freshwater Macroinvertebrates. Water. 2021; 13(18):2586. https://doi.org/10.3390/w13182586
Chicago/Turabian StyleTheodoropoulos, Christos, Ioannis Karaouzas, and Anastasios Stamou. 2021. "Environmental Flows as a Proactive Tool to Mitigate the Impacts of Climate Warming on Freshwater Macroinvertebrates" Water 13, no. 18: 2586. https://doi.org/10.3390/w13182586
APA StyleTheodoropoulos, C., Karaouzas, I., & Stamou, A. (2021). Environmental Flows as a Proactive Tool to Mitigate the Impacts of Climate Warming on Freshwater Macroinvertebrates. Water, 13(18), 2586. https://doi.org/10.3390/w13182586