
Regulation of Vegetation and Evapotranspiration by Water Level Fluctuation in Shallow Lakes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
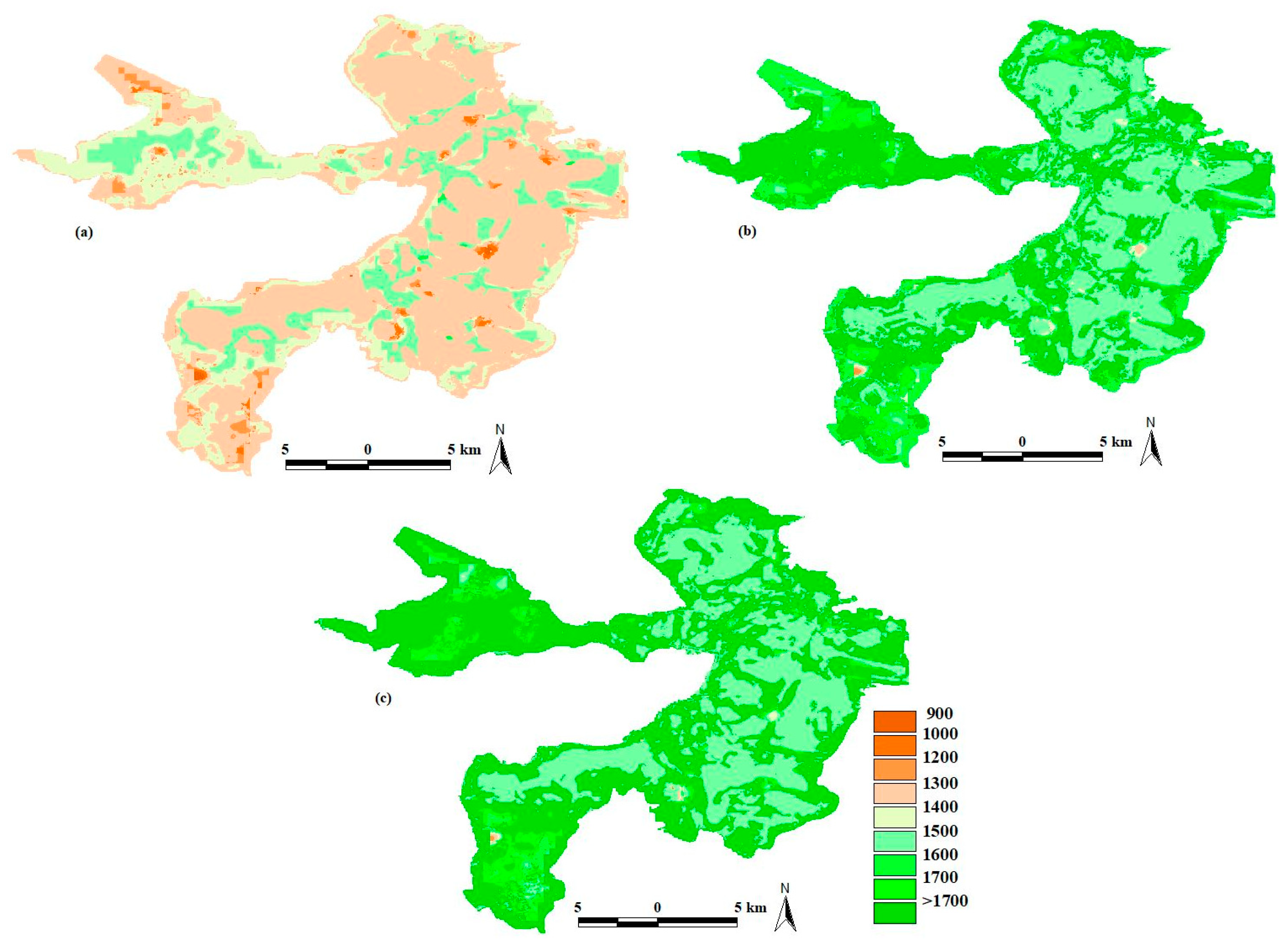
2.1. Study Site
2.2. Data and Field Experiments
2.3. Methods
3. Results
3.1. Alterations in Water-Fluctuations
3.2. Variation in Vegetation Patterns under Water Level Fluctuations
3.3. Evapotranspiration Variation under Water level Fluctuations
4. Discussion
4.1. Response of Ecological Effects to Water Level Fluctuations
4.2. Implications for Wetland Restoration and Management
5. Conclusions
- (i)
- The average water levels at annual scale or during germination stage (e.g., March and April) play a key role in regulating the vegetation community. However, the abrupt rise in water levels (due to ecological water transfer) inevitably caused disturbances to aquatic vegetation communities.
- (ii)
- The ET in different vegetation patterns influenced by water level fluctuations were estimated using the dynamic vegetation model combined with TVET model. Physical and biological processes influenced by varying macrophyte (e.g., P. australis) cover and open water area altered the partition of ET. Interestingly, high ET values appeared at low water levels instead of at high water levels.
- (iii)
- Suitable water levels, estimated for maintaining suitable habitat for P. australis (avoiding abrupt changes in ratio of vegetation cover to open water area, and maintaining higher Fv), ranged from 6.59 to 6.97 m. Furthermore, ecological water transfer projects and operations should be regulated under a specific mode of operation to reduce their impact on macrophytes while maintaining the integrity of Baiyangdian Lake ecosystems, e.g., avoiding abrupt water level rise during the generation stage.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ponce-Campos, G.E.; Moran, M.S.; Huete, A.; Zhang, Y.; Bresloff, C.; Huxman, T.E.; Eamus, D.; Bosch, D.D.; Buda, A.R.; Gunter, S.A.; et al. Ecosystem resilience despite large-scale altered hydroclimatic conditions. Nature 2013, 494, 349–352. [Google Scholar] [CrossRef]
- Knapp, A.K.; Hoover, D.L.; Wilcox, K.R.; Avolio, M.L.; Koerner, S.E.; La Pierre, K.J.; Loik, M.E.; Luo, Y.Q.; Sala, O.; Smith, M.D. Characterizing differences in precipitation regimes of extreme wet and dry years: Implications for climate change experiments. Glob. Chang. Biol. 2015, 21, 2624–2633. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; He, Q.; Yang, B.; He, W.; Xu, F.; Janssen, A.B.; Kuiper, J.J.; van Gerven, L.A.; Qin, N.; Jiang, Y.J.; et al. Hydrological regulation drives regime shifts: Evidence from paleolimnology and ecosystem modeling of a large shallow Chinese lake. Glob. Chang. Biol. 2017, 23, 737–754. [Google Scholar] [CrossRef] [PubMed]
- Reid, W.V.; Mooney, H.A.; Cropper, A.; Capistrano, D.; Zurek, M.B. Millennium Ecosystem Assessment; Synthesis Report; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Poff, N.L.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The Natural Flow Regime. Bioscience 1997, 47, 769–784. [Google Scholar] [CrossRef]
- Poff, N.L. Beyond the natural flow regime? Broadening the hydro-ecological foundation to meet environmental flows challenges in a non-stationary world. Freshw. Biol. 2018, 63, 1011–1021. [Google Scholar] [CrossRef]
- Palmer, M.; Ruhi, A. Linkages between flow regime, biota, and ecosystem processes: Implications for river restoration. Science 2019, 365, eaaw2087. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Liu, J.; Liu, H.; Liang, L.; Cai, Y.; Wang, X.; Li, C. Vegetation dynamics under water-level fluctuations: Implications for wetland restoration. J. Hydrol. 2020, 581, 124418. [Google Scholar] [CrossRef]
- Polvi, L.E.; Lind, L.; Persson, H.; Miranda-Melo, A.; Pilotto, F.; Su, X.; Nilsson, C. Facets and scales in river restoration: Nestedness and interdependence of hydrological, geomorphic, ecological, and biogeochemical processes. J. Environ. Manag. 2020, 265, 110288. [Google Scholar] [CrossRef] [PubMed]
- Barbarossa, V.; Schmitt, R.J.P.; Huijbregts, M.A.J.; Zarfl, C.; King, H.; Schipper, A.M. Impacts of current and future large dams on the geographic range connectivity of freshwater fish worldwide. Proc. Natl. Acad. Sci. USA 2020, 117, 3648–3655. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Wang, X.T.; Yang, Z.; Mao, C.; West, A.J.; Ji, J. Dam-triggered organic carbon sequestration makes the Changjiang (Yangtze) river basin (China) a significant carbon sink. J. Geophys. Res. Biogeosci. 2015, 120, 39–53. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.B.; Wen, Y.; Zou, L.F.; Wang, Z.Q.; He, C.G.; Zou, C.L. The effects of a wetland restoration project on the Siberian crane (Grus leucogeranus) population and stopover habitat in Momoge National Nature Reserve, China. Ecol. Eng. 2016, 96, 170–177. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, Z.F.; Liu, Q.; Zhao, Y. Water preservation and the ecological effects of removing leaves from stalks for a reed dominant wetland. Ecol. Eng. 2012, 49, 118–122. [Google Scholar] [CrossRef]
- Cui, B.; Li, X.; Zhang, K. Classification of hydrological conditions to assess water allocation schemes for Lake Baiyangdian in North China. J. Hydrol. 2010, 385, 247–256. [Google Scholar] [CrossRef]
- Liu, Q. Effects of groundwater level fluctuation on Phragmites australis evapotranspiration in the Baiyangdian Lake. Wetl. Sci. 2014, 12, 552–558. [Google Scholar]
- Zhao, Y.; Yang, Z.; Xia, X.; Wang, F. A shallow lake remediation regime with Phragmites australis: Incorporating nutrient removal and water evapotranspiration. Water Res. 2012, 46, 5635–5644. [Google Scholar] [CrossRef]
- Li, X.; Cui, B.; Yang, Q.; Lan, Y. Impacts of water level fluctuations on detritus accumulation in Lake Baiyangdian, China. Ecohydrology 2016, 9, 52–67. [Google Scholar] [CrossRef]
- Wang, Q.; Yan, D.; Yuan, Y.; Wang, D. Study on the Quantification of Drought in Freshwater Wetlands—A Case Study in Baiyangdian Wetland. Wetlands 2014, 34, 1013–1025. [Google Scholar] [CrossRef]
- Shuttleworth, W.J. Evaporation. In Handbook of Hydrology; Maidment, D.R., Ed.; McGraw-Hill: New York, NY, USA, 1993; Chapter 4. [Google Scholar]
- Guan, H.D.; Wilson, J.L. A hybrid dual-source model for potential evaporation and transpiration partitioning. J. Hydrol. 2009, 377, 405–416. [Google Scholar] [CrossRef]
- Serrano, A.; Mateos, V.L.; García, J.A. Trend analysis of monthly precipitation over the Iberian Peninsula for the period 1921–1995. Phys. Chem. Earth B 1999, 24, 85–90. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, Z. Quantitative estimation of the impact of climate change on actual evapotranspiration in the Yellow River Basin, China. J. Hydrol. 2010, 395, 226–234. [Google Scholar] [CrossRef]
- Mann, H.B. Non-parametric test against trend. Econometrika 1945, 13, 245–259. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods; Charles Griffin: London, UK, 1975. [Google Scholar]
- Birsan, M.-V.; Molnar, P.; Burlando, P.; Pfaundler, M. Streamflow trends in Switzerland. J. Hydrol. 2005, 314, 312–329. [Google Scholar] [CrossRef]
- Carlson, T.N.; Ripley, D.A. On the relation between NDVI, fractional vegetation cover, and leaf area index. Remote Sens. Environ. 1997, 62, 241–252. [Google Scholar] [CrossRef]
- Huang, H.M.; Zhang, L.Q.; Guan, Y.J.; Wang, D.H. A cellular automata model for population expansion of Spartina alterniflora at Jiuduansha Shoals, Shanghai, China. Estuar. Coast. Shelf Sci. 2008, 77, 47–55. [Google Scholar] [CrossRef]
- Qi, M.; Sun, T.; Xue, S.; Yang, W.; Shao, D.; Martinez-Lopez, J. Competitive ability, stress tolerance and plant interactions along stress gradients. Ecology 2018, 99, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Wolfram, S. Universality and complexity in cellular automata. Phys. D Nonlinear Phenom. 1984, 10, 1–35. [Google Scholar] [CrossRef]
- Foti, R.; del Jesus, M.; Rinaldo, A.; Rodriguez-Iturbe, I. Signs of critical transition in the Everglades wetlands in response to climate and anthropogenic changes. Proc. Natl. Acad. Sci. USA 2013, 110, 6296–6300. [Google Scholar] [CrossRef] [Green Version]
- Budelsky, R.A.; Galatowitsch, S.M. Effects of water regime and competition on the establishment of a native sedge in restored wetlands. J. Appl. Ecol. 2000, 37, 971–985. [Google Scholar] [CrossRef]
- Wilcox, D.A.; Nichols, S.J. The effects of water-level fluctuations on vegetation in a Lake Huron wetland. Wetlands 2008, 28, 487–501. [Google Scholar] [CrossRef]
- Schooler, S.S.; Salau, B.; Julien, M.H.; Ives, A.R. Alternative stable states explain unpredictable biological control of Salvinia molesta in Kakadu. Nature 2011, 470, 86–89. [Google Scholar] [CrossRef]
- van der Valk, A.G. Effects of prolonged flooding on the distribution and biomass of emergent species along a freshwater wetland coenocline. Vegetatio 1994, 110, 185–196. [Google Scholar] [CrossRef]
- Havens, K.E.; Fox, D.; Gornak, S.; Hanlon, C. Aquatic vegetation and largemouth bass population responses to water-level variations in Lake Okeechobee, Florida (USA). Hydrobiologia 2005, 539, 225–237. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Jeppesen, E.; Liu, X.H.; Qin, B.Q.; Shi, K.; Zhou, Y.Q.; Thomaz, S.M.; Deng, J.M. Global loss of aquatic vegetation in lakes. Earth-Sci. Rev. 2017, 173, 259–265. [Google Scholar] [CrossRef]
- An, Y.; Gao, Y.; Tong, S.Z. Emergence and growth performance of Bolboschoenus planiculmis varied in response to water level and soil planting depth: Implications for wetland restoration using tuber transplantation. Aquat. Bot. 2018, 148, 10–14. [Google Scholar] [CrossRef]
- Jacinthe, P.A. Carbon dioxide and methane fluxes in variably-flooded riparian forests. Geoderma 2015, 241, 41–50. [Google Scholar] [CrossRef]
- Ye, X.C.; Meng, Y.K.; Xu, L.G.; Xu, C.Y. Net primary productivity dynamics and associated hydrological driving factors in the floodplain wetland of China’s largest freshwater lake. Sci. Total Environ. 2019, 659, 302–313. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guan, H.; Batelaan, O.; McVicar, T.R.; Long, D.; Piao, S.; Liang, W.; Liu, B.; Jin, Z.; Simmons, C.T. Contrasting responses of water use efficiency to drought across global terrestrial ecosystems. Sci. Rep. 2016, 6, 23284. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Jaen, M.C.; Aide, T.M. Restoration Success: How Is It Being Measured? Restor. Ecol. 2005, 13, 569–577. [Google Scholar] [CrossRef]
- Bossuyt, B.; Honnay, O. Can the seed bank be used for ecological restoration? An overview of seed bank characteristics in European communities. J. Veg. Sci. 2008, 19, 875–884. [Google Scholar] [CrossRef]
- Frank, D.; Reichstein, M.; Bahn, M.; Thonicke, K.; Frank, D.; Mahecha, M.D.; Smith, P.; van der Velde, M.; Vicca, S.; Babst, F.; et al. Effects of climate extremes on the terrestrial carbon cycle: Concepts, processes and potential future impacts. Glob. Chang. Biol. 2015, 21, 2861–2880. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Anderson, N.J.; Yang, X.; Chen, X.; Shen, J. Carbon burial by shallow lakes on the Yangtze floodplain and its relevance to regional carbon sequestration. Glob. Chang. Biol. 2012, 18, 2205–2217. [Google Scholar] [CrossRef]
- Sánchez-Carrillo, S.; Angeler, D.G.; Sanchez-Andres, R.; Alvarez-Cobelas, M.; Garatuza-Payan, J. Evapotranspiration in semi-arid wetlands: Relationships between inundation and the macrophyte-cover: Open-water ratio. Adv. Water Resour. 2004, 27, 643–655. [Google Scholar] [CrossRef]
- Midwood, J.D.; Chow-Fraser, P. Changes in aquatic vegetation and fish communities following 5 years of sustained low water levels in coastal marshes of eastern Georgian Bay, Lake Huron. Glob. Chang. Biol. 2012, 18, 93–105. [Google Scholar] [CrossRef]
- Blodau, C.; Moore, T.R. Experimental response of peatland carbon dynamics to a water table fluctuation. Aquat. Sci. Res. Across Boundaries 2003, 65, 47–62. [Google Scholar] [CrossRef]
- Chimner, R.A.; Pypker, T.G.; Hribljan, J.A.; Moore, P.A.; Waddington, J.M. Multi-decadal Changes in Water Table Levels Alter Peatland Carbon Cycling. Ecosystems 2016, 20, 1042–1057. [Google Scholar] [CrossRef]
- Reich, P.; Lake, P.S. Extreme hydrological events and the ecological restoration of flowing waters. Freshw. Biol. 2015, 60, 2639–2652. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | P. australis | Open-Water Area | Other Land-Use Types | Total | Accuracy (%) |
|---|---|---|---|---|---|
| P. australis | 129,275 | 11,607 | 0 | 140,882 | 91.76 |
| Open-water area | 38,819 | 73,894 | 0 | 112,713 | 65.56 |
| Other land-use types | 3018 | 1141 | 82,502 | 86,661 | 95.20 |
| Total | 171,112 | 86,642 | 82,502 | ||
| Accuracy (%) | 75.55 | 85.29 | 100 | ||
| Overall accuracy | 84% | Kappa | 0.75 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.; Liang, L.; Yuan, X.; Yan, S.; Li, M.; Li, S.; Wang, X.; Li, C. Regulation of Vegetation and Evapotranspiration by Water Level Fluctuation in Shallow Lakes. Water 2021, 13, 2651. https://doi.org/10.3390/w13192651
Liu Q, Liang L, Yuan X, Yan S, Li M, Li S, Wang X, Li C. Regulation of Vegetation and Evapotranspiration by Water Level Fluctuation in Shallow Lakes. Water. 2021; 13(19):2651. https://doi.org/10.3390/w13192651
Chicago/Turabian StyleLiu, Qiang, Liqiao Liang, Xiaomin Yuan, Sirui Yan, Miao Li, Shuzhen Li, Xuan Wang, and Chunhui Li. 2021. "Regulation of Vegetation and Evapotranspiration by Water Level Fluctuation in Shallow Lakes" Water 13, no. 19: 2651. https://doi.org/10.3390/w13192651