
Stem Photosynthesis Affects Hydraulic Resilience in the Deciduous Populusalba but Not in the Evergreen Laurus nobilis


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Condition
2.2. Morphological Measurements
2.3. Leaf Water Potential Isotherms
2.4. Gas Exchange and Water Potential Measurements
2.5. Hydraulic Conductivity Measurements
2.6. NSC Content
2.7. Statistical Analysis
3. Results
3.1. Stem Shading Effects on Plant Growth
3.2. Plant Water Relations
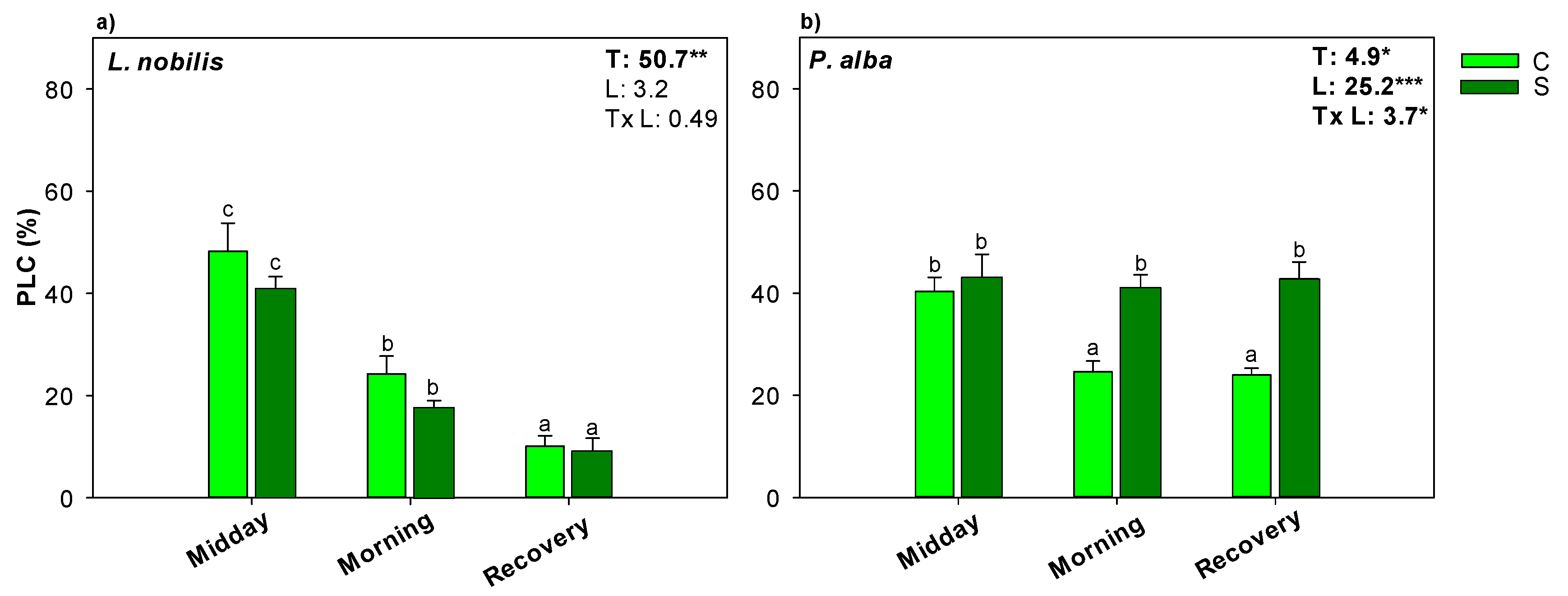
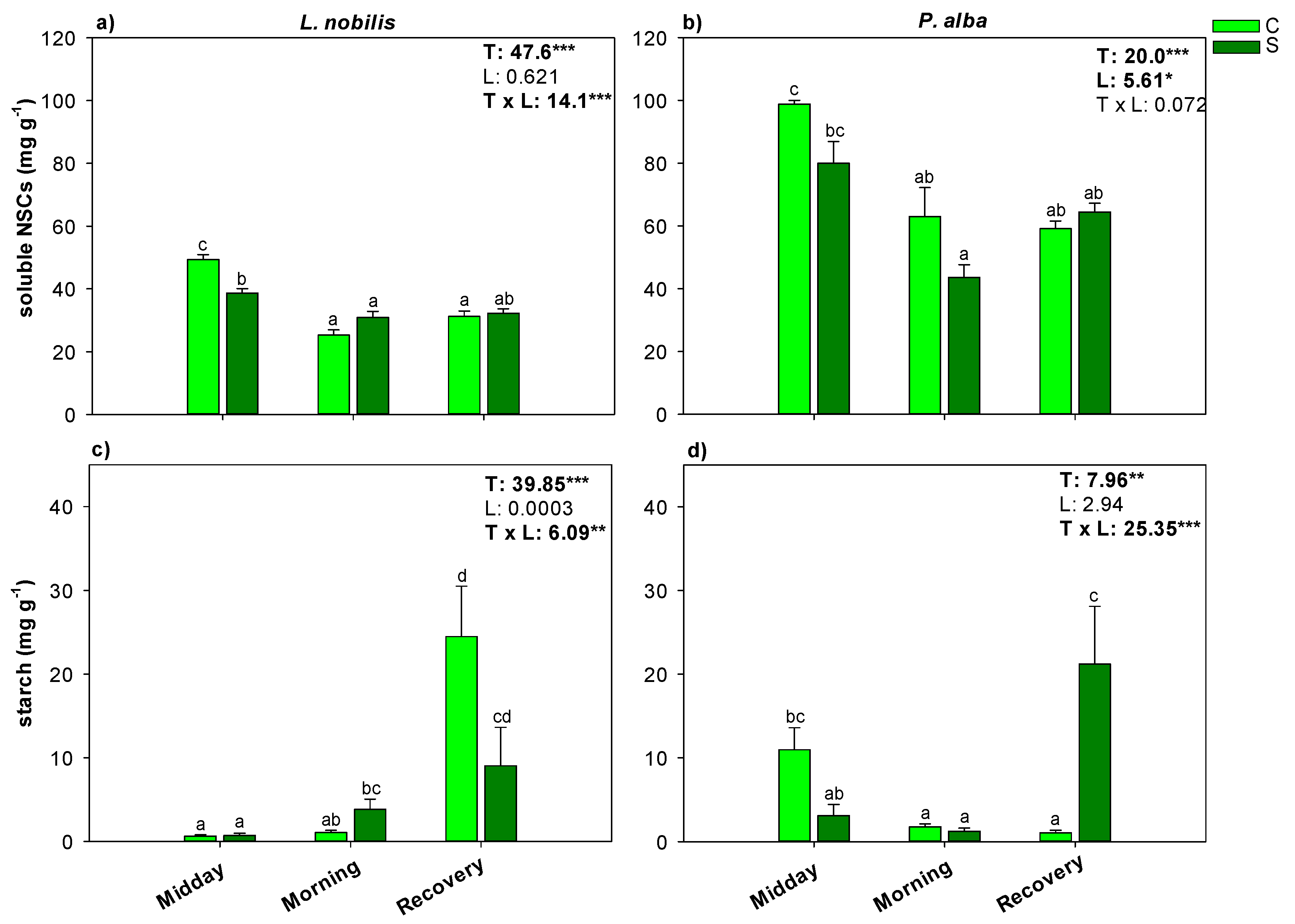
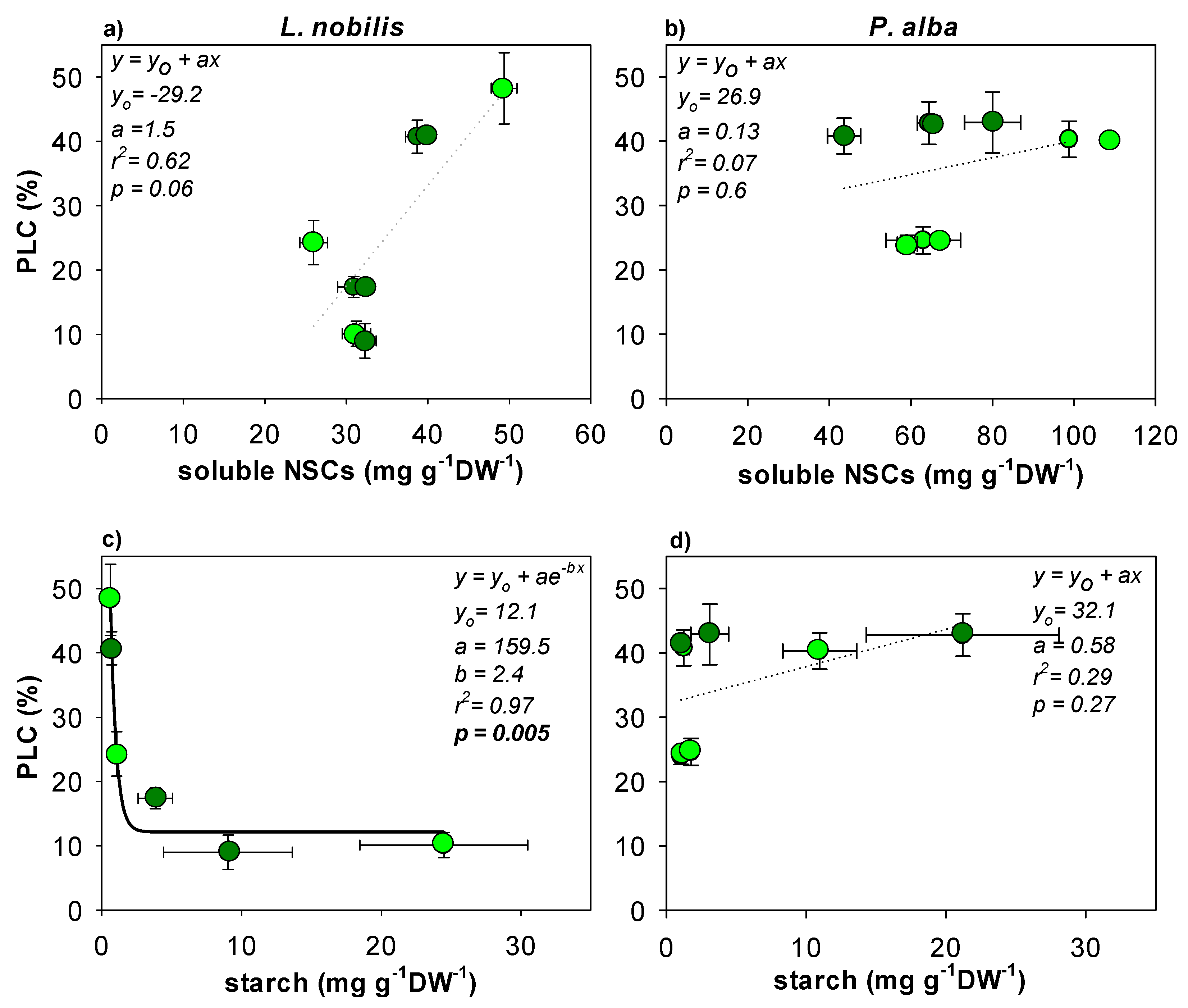
3.3. Hydraulic Recovery and NSCs Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wiebe, H.H. Photosynthesis in wood. Physiol. Plant. 1975, 33, 245–246. [Google Scholar] [CrossRef]
- Van Cleve, B.; Apel, K. Induction by nitrogen and low temperature of storage-protein synthesis in poplar trees exposed to long days. Planta 1993, 189, 157–160. [Google Scholar] [CrossRef]
- Berveiller, D.; Kierzkowski, D.; Damesin, C. Interspecific variability of stem photosynthesis among tree species. Tree Physiol. 2007, 27, 53–61. [Google Scholar] [CrossRef]
- Rentzou, G.K.; Psaras, A. Green plastids, maximal PSII photochemical efficiency and starch content of inner stem tissues of three Mediterranean woody species during the year. Flora 2008, 203, 350–357. [Google Scholar] [CrossRef]
- Nilsen, E.T. Stem photosynthesis: Extent, patterns, and role in plant carbon economy. In Plant. Stems; Gartner, B.L., Ed.; Elsevier: Amsterdam, The Netherlands, 1995; pp. 223–240. [Google Scholar]
- Teskey, R.O.; Saveyn, A.; Steppe, K.; McGuire, M.A. Origin, fate and significance of CO2 in tree stems. New Phytol. 2008, 177, 17–32. [Google Scholar] [CrossRef] [PubMed]
- Avila-Lovera, E.; Herrera, A.; Tezara, W. Contribution of stem CO2 fixation to whole-plant carbon balance in nonsucculent species. Photosyntica 2014, 52, 3–15. [Google Scholar] [CrossRef]
- Aschan, G.; Pfanz, H. Non foliar photosynthesis—A strategy of additional carbon acquisition. Flora Morphol. Distrib. Funct. Ecol. Plants 2003, 198, 81–97. [Google Scholar] [CrossRef]
- Ehleringer, J.R.; Phillips, S.L.; Comstock, J.P. Seasonal variation in the carbon isotopic composition of desert plants. Funct. Ecol. 1992, 6, 396–404. [Google Scholar] [CrossRef]
- Nilsen, E.T.; Sharifi, M.R. Carbon isotopic composition of legumes with photosynthetic stems from mediterranean and desert habitats. Am. J. Bot. 1997, 84, 1707–1713. [Google Scholar] [CrossRef]
- Avila-Lovera, E.; Tezara, W. Water use efficiency is higher in green stems than in leaves of a tropical tree species. Trees 2018, 32, 1547–1558. [Google Scholar] [CrossRef]
- Avila-Lovera, E.; Urich, R.; Coronel, I.; Tezara, W. Seasonal gas exchange and resource-use efficiency in evergreen versus deciduous species from a tropical dry forest. Tree Physiol. 2019, 39, 1561–1571. [Google Scholar] [CrossRef] [PubMed]
- Santiago, L.S.; Goldstein, G.; Meinzer, F.C.; Fisher, J.B.; Machado, K.; Woodruff, D.; Jones, T. Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees. Oecol 2004, 140, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Osmond, C.B.; Smith, S.D.; Gui-Ying, B.; Sharkey, T.D. Stem photosynthesis in a desert ephemeral, Eriogonum inflatum. Characterization of leaf and stem CO2 fixation and H2O vapor exchange under controlled conditions. Oecol 1987, 72, 542–549. [Google Scholar] [CrossRef]
- Nilsen, E.T.; Meinzer, F.C.; Rundel, P.W. Stem photosynthesis in Psorothamnus spinosus (smoke tree) in the Sonorandesert of California. Oecol 1989, 79, 193–197. [Google Scholar] [CrossRef]
- Nilsen, E.T.; Bao, Y. The influence of water stress on stem and leaf photosynthesis in Glycine max and Spartium junceum (leguminosae). Am. J. Bot. 1990, 77, 1007–1015. [Google Scholar] [CrossRef]
- Tinoco-Ojanguren, C. Diurnal and seasonal patterns of gas exchange and carbon gain contribution of leaves and stems of Justicia californica in the Sonoran Desert. J. Arid Environ. 2008, 72, 127–140. [Google Scholar] [CrossRef]
- Damesin, C. Respiration and photosynthesis characteristics of current-year stems of Fagus sylvatica: From the seasonal pattern to an annual balance. New Phytol. 2003, 158, 465–475. [Google Scholar] [CrossRef]
- De Roo, L.; Salomon, R.L.; Steppe, K. Woody tissue photosynthesis reduces stem CO2 efflux by half and remains unaffected by drought stress in young Populus tremula trees. Plant. Cell Enviorn. 2020, 43, 981–991. [Google Scholar] [CrossRef] [PubMed]
- Saveyn, K.A.; Steppe, N.; Ubierna, T.E. Woody tissue photosynthesis and its contribution to trunk growth and bud development in young plants. Plant. Cell Environ. 2010, 33, 1949–1958. [Google Scholar] [CrossRef]
- Bloemen, J.; Overlaet-Michiels, L.; Steppe, K. Understanding plant responses to drought: How important is woody tissue photosynthesis? Acta Hortic. 2013, 991, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Cernusak, L.A.; Hutley, L.B. Stable isotopes reveal the contribution of corticular photosynthesis to growth in branches of Eucalyptus miniata. Plant. Physiol. 2011, 155, 515–523. [Google Scholar] [CrossRef] [Green Version]
- Simbo, D.J.; Van den Bilcke, N.; Samson, R. Contribution of corticular photosynthesis to bud development in african baobab (Adansonia digitata L.) and castor bean (Ricinus communis L.) seedlings. Environ. Exp. Bot. 2013, 95, 1–5. [Google Scholar] [CrossRef]
- Liu, J.; Gu, L.; Yu, J.; Ju, G.; Sun, Z. Stem photosynthesis of twig and its contribution to new organ development in cutting seedlings of Salix Matsudana Koidx. Forests 2018, 9, 207. [Google Scholar] [CrossRef] [Green Version]
- Bossard, C.C.; Rejmanek, M. Why have green stems? Funct. Ecol. 1992, 6, 197–205. [Google Scholar] [CrossRef]
- Pfanz, H.; Aschan, G. The existence of bark and stem photosynthesis in woody plants and its significance for the overall carbon gain. An eco-physiological and ecological approach. In Progress in Botany; Essr, K., Luttge, U., Kadereit, J.W., Beyshlag, W., Eds.; Springer: Berlin, Germany, 2001; Volume 62, pp. 477–510. [Google Scholar]
- Eyles, A.; Pinkard, E.A.; Mohammed, C. Shifts in biomass and resource allocation patterns following defoliation in Eucalyptus globulus growing with varying water and nutrient supplies. Tree Physiol. 2009, 29, 753–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cernusak, L.A.; Marshall, J.D. Photosynthetic refixation in branches of western white pine. Funct. Ecol. 2000, 14, 300–311. [Google Scholar]
- Pfanz, H.; Aschan, G.; Langenfeld-Heyser, R.; Wittmann, C.; Loose, M. Ecology and ecophysiology of tree stems: Corticular and wood photosynthesis. Naturwissenschaften 2002, 89, 147–162. [Google Scholar]
- Wittmann, C.; Pfanz, H. Temperature dependency of bark photosynthesis in beech (Fagus sylvatica L.) and birch (Betula pendula Roth.) trees. J. Exp. Bot. 2007, 58, 4293–4306. [Google Scholar] [CrossRef]
- Wittmann, C.; Pfanz, H. Bark and woody tissue photosynthesis: A means to avoid hypoxia or anoxia in developing stem tissue. Funct. Plant. Biol. 2014, 41, 940–953. [Google Scholar] [CrossRef]
- Wittmann, C.; Pfanz, H. More than just CO2-recycling: Corticular photosynthesis as a mechanism to reduce the risk of an energy crisis induced by low oxygen. New Phythol. 2018, 219, 551–564. [Google Scholar] [CrossRef] [Green Version]
- Comstock, J.P.; Ehleringer, J.R. Contrasting photosynthetic behaviour in leaves and twigs of Hymenoclea salsola, a green-twigged warm desert shrub. Am. J. Bot. 1988, 75, 1360–1370. [Google Scholar] [CrossRef]
- Dima, E.; Manetas, Y.; Psaras, G.H. Chlorophyll distribution pattern in inner stem tissues: Evidence from epifluorescence microscopy and reflectance measurements in 20 woody species. Trees 2006, 20, 515–521. [Google Scholar] [CrossRef]
- Vandegehuchte, M.W.; Bloemen, J.; Vergeynst, L.L.; Steppe, K. Woody tissue photosynthesis in trees: Salve on the wounds of drought? New Phytol. 2015, 208, 998–1002. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, H.; Ziegler, W.; Kolle, O.; Trumbore, S. Thirst beats hunger declining hydration during drought prevents carbon starvation in Norway spruce saplings. New Phytol. 2013, 200, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Adams, H.D.; Zeppel, M.J.; Anderegg, W.R.; Hartmann, H.; Landhäusser, S.M.; Tissue, D.T.; Huxman, T.E.; Hudson, P.J.; Franz, T.E.; Allen, C.D.; et al. A multispecies synthesis of physiological mechanisms in drought-induced tree mortality. Nat. Ecol. Evol. 2017, 1, 1285–1291. [Google Scholar] [CrossRef]
- Hammond, W.M.; Yu, K.; Wilson, L.A.; Will, R.E.; Anderegg, W.R.L.; Adams, H.D. Dead or dying? Quantifying the point of no return from hydraulic failure in drought- induced tree mortality. New Phytol. 2019, 223, 1834–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDowell, N.G.; Beerling, D.J.; Breshears, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef]
- Hartmann, H.; Trumbore, S. Understanding the roles of non-structural carbohydrates in forest trees from what we can measure to what we want to know. New Phytol. 2016, 211, 386–403. [Google Scholar] [CrossRef] [Green Version]
- Nardini, A.; Savi, T.; Trifilò, P.; Lo Gullo, M.A. Drought stress and the recovery from xylem embolism in woody plants. In Progress in Botany; Canovas, F., Luttgge, U., Matyssek, R., Eds.; Springer: Berlin, Germany, 2018; Volume 79, pp. 197–231. [Google Scholar]
- Tomasella, M.; Petrussa, E.; Petruzzellis, F.; Nardini, A.; Casolo, V. The possible role of non-structural carbohydrates in the regulation of tree hydraulics. Int. J. Mol. Sci. 2020, 21, 144. [Google Scholar] [CrossRef] [Green Version]
- Sapes, G.; Sala, A. Relative water content consistently predicts drought mortality risk in seedling populations with different morphology, physiology, and times to death. Plant. Cell Environ. 2021, 44, 3322–3335. [Google Scholar] [CrossRef]
- Deslauriers, A.; Beaulieu, M.; Balducci, L.; Giovannelli, A.; Gagnon, M.J.; Rossi, S. Impact of warming and drought on varbon balance related to wood formation in black spruce. Ann. Bot. 2014, 33, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Falchi, R.; Petrussa, E.; Braidot, E.; Sivilotti, P.; Boscutti, F.; Vuerich, M.; Calligaro, C.; Filippi, A.; Herrera, J.C.; Sabbatini, P.; et al. Analysis of non-structural carbohydrates and xylem anatomy of leaf petioles offers new insights in the drought response of two grapevine cultivars. Int. J. Mol. Sci. 2020, 21, 1457. [Google Scholar] [CrossRef] [Green Version]
- Sala, A.; Woodruff, D.R.; Meinzer, F.C. Carbon dynamics in trees: Feast or famine? Tree Physiol. 2012, 32, 764–775. [Google Scholar] [CrossRef] [Green Version]
- Plavcova, L.; Jansen, S. The role of xylem parenchyma in the storage and utilization of non-structural carbohydrates. In Functional and Ecological Xylem Anatomy; Hacke, U., Ed.; Springer: Berin, Germany, 2015; pp. 209–234. [Google Scholar]
- Traversari, L.; Neri, M.; Traversi, L. Daily osmotic adjustments in stem may be good predictors of water stress intensity in poplar. Plant. Physiol. Biochem. 2020, 146, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Neuner, G. Frost resistance in alpine woody plants. Front. Plant. Sci. 2014, 5, 654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tixier, A.; Gambetta, G.A.; Godfrey, J.; Zwieniecki, M.A. Non-structural carbohydrates in dormant woody perennials: The tale of winter survival and spring arrival. Front. For. Glob. Chang. 2019, 2, 18. [Google Scholar] [CrossRef] [Green Version]
- Améglio, T.; Decourteix, M.; Alves, G.; Valentin, V.; Sakr, S.; Julien, J.; Guillot, A.; Lacointe, A. Temperature effects on xylem sap osmolarity in walnut trees: Evidence for a vitalistic model of winter embolism repair. Tree Physiol. 2004, 24, 785–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayr, S.; Kartusch, B.; Kikuta, S. Evidence for air-seeding: Watching the formation of embolism in conifer xylem. J. Plant. Hydraul. 2014, 1, e0004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayr, S.; Ameglio, T. Freezing stress in tree xylem. In Progress Bot; Luttge, U., Canovas, F., Matyssek, R., Eds.; Springer: Berlin, Germany, 2016; Volume 77, pp. 381–414. [Google Scholar]
- Secchi, F.; Zwieniecki, M.A. Accumulation of sugars in the xylem apoplast observed under water stress conditions is controlled by xylem pH. Plant. Cell Environ. 2016, 39, 2350–2360. [Google Scholar] [CrossRef]
- Pagliarani, C.; Casolo, V.; Beiragi, M.A.; Cavalletto, S.; Siciliano, I.; Schubert, A.; Gullino, M.L.; Zwieniechi, M.A.; Secchi, F. Priming xylem for stress recovery depends on coordinated activity of sugar metabolic pathways and changes in xylem sap pH. Plant. Cell Environ. 2019, 42, 1775–1787. [Google Scholar] [CrossRef] [Green Version]
- Oroian, M.; Ropciuc, S.; Amariei, S.; Gutt, G. Correlations between density viscosity, surface tension and ultrasonic velocity of different mono- and di-saccharides. J. Mol. Liq. 2015, 207, 145–151. [Google Scholar] [CrossRef]
- Losso, A.; Beikircher, B.; Damon, B.; Kikuta, S.; Schimd, P.; Mayr, S. Xylem sap surface tension may be crucial for hydraulic safety. Plant. Physiol. 2017, 175, 1135–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schenk, H.J.; Espino, S.; Romo, D.M.; Nima, N.; Do, A.Y.; Michaud, J.M.; Papahadjopoulos-Sternberg, B.; Yang, J.; Zuo, Y.Y.; Steppe, K.; et al. Xylem surfactants introduce a new element to the Cohesion-Tension theory. Plant. Physiol. 2017, 173, 1177–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloemen, J.; Vergeynst, L.; Overlaet-Michielis, L.; Stepps, K. How important is woody tissue photosynthesis in poplar during drought stress? Trees 2016, 30, 63–72. [Google Scholar] [CrossRef]
- De Baerdemaeker, N.J.F.; Salomon, R.L.; De Roo, L.; Steppe, K. Sugars from woody tissue photosynthesis reduce xylem vulnerability to cavitation. New Phytol. 2017, 216, 720–727. [Google Scholar] [CrossRef] [Green Version]
- Tomasella, M.; Casolo, V.; Natale, S.; Petruzzellis, F.; Kofler, W.; Beikircher, B.; Mayr, S.; Nardini, A. shade-induced reduction of stem nonstructural carbohydrates increases xylem vulnerability to embolism and impedes hydraulic recovery in Populus nigra. New Phytol. 2021, 231, 108–121. [Google Scholar] [CrossRef]
- Schmitz, N.; Egerton, J.J.G.; Lovelock, C.E.; Ball, M.C. Light-dependent maintenance of hydraulic function in mangrove branches: Do xylary chloroplasts play a role in embolism repair? New Phytol. 2012, 195, 40–46. [Google Scholar] [CrossRef]
- Liu, J.; Gun, L.; Yu, Y.; Huang, P.; Wu, Z.; Zhang, Q.; Qian, Y.; Wan, X.; Sun, Z. Corticular photosynthesis drives bark water uptake to refill embolized vessels in dehydrated branches of Salix matsudana. Plant. Cell Environ. 2019, 42, 2584–2596. [Google Scholar] [CrossRef]
- Trifilò, P.; Nardini, A.; Lo Gullo, M.A.; Barbera, P.M.; Raimondo, F. Diurnal changes in embolism rate in nine dry forest trees: Relationship with specie-specific xylem vulnerability, hydraulic strategy and wood traits. Tree Physiol. 2015, 35, 694–705. [Google Scholar] [CrossRef] [Green Version]
- Tyree, M.T.; Hammel, H.T. The measurement of the turgor pressure and the water relations of plants by the pressure-bomb technique. J. Exp. Bot. 1972, 23, 267–282. [Google Scholar] [CrossRef]
- Huckin, K.; Cochard, H.; Dreyer, E.; Le Thiec, D.; Bogeat-Triboulot, M.B. Cavitation vulnerability in roots and shoots: Does Populus euphratica Oliv., a poplar from arid areas of Central Asia, differ from other poplar species? J. Exp. Bot. 2005, 418, 2003–2010. [Google Scholar] [CrossRef]
- Nardini, A.; Savi, T.; Losso, A.A.; Petit, G.; Pacilè, S.; Tromba, G.; Mayr, S.; Trifilò, P.; Lo Gullo, M.A.; Salleo, S. X-ray micro-tomography observations of xylem embolism in stems of Laurus nobilis L. are consistent with hydraulic measurements of percent loss of conductance. New Phytol. 2017, 213, 1068–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperry, J.S.; Donnelly, J.R.; Tyree, M.T. A method for measuring hydraulic conductivity and embolism in xylem. Plant. Cell Environ. 1988, 11, 35–40. [Google Scholar] [CrossRef]
- Lo Gullo, M.A.; Salleo, S. Three different methods for measuring xylem cavitation and embolism: A comparison. Ann. Bot. 1991, 67, 417–424. [Google Scholar] [CrossRef]
- Nardini, A.; Gascó, A.; Trifilò, P.; Lo Gullo, M.A.; Salleo, S. Ion-mediated enhancement of xylem hydraulic conductivity is not always suppressed by the presence of Ca2+ in the sap. J. Exp. Bot. 2007, 58, 2609–2615. [Google Scholar] [CrossRef] [Green Version]
- Wheeler, J.K.; Huggett, B.A.; Tofte, A.N.; Rockwell, F.E.; Holbrook, N.M. Cutting xylem under tension or supersaturated with gas can generate PLC and the appearance of rapid recovery from embolism. Plant. Cell Environ. 2013, 36, 1938–1949. [Google Scholar] [CrossRef]
- Trifilò, P.; Raimondo, F.; Lo Gullo, M.A.; Barbera, P.M.; Salleo, S. Nardini, A. Relax and refill: Xylem rehydration prior to hydraulic measurements favours embolism repair in stems and generates artificially low PLC values. Plant. Cell Environ. 2014, 37, 2491–2499. [Google Scholar] [CrossRef]
- Quentin, A.G.; Pinkard, E.A.; Ryan, M.G.; Tissue, D.T.; Baggett, L.S.; Adams, H.D.; Maillard, P.; Marchand, J.; Landhäusser, S.M.; Lacointe, A.; et al. Non-structural carbohydrates in woody plants compared among laboratories. Tree Physiol. 2015, 35, 1146–1165. [Google Scholar] [CrossRef] [Green Version]
- Landhausser, S.M.; Chow, P.S.; Dickman, L.T.; Furze, M.E.; Kuhlman, I.; Schmid, S.; Wiesenbauer, J.; Wild, B.; Gleixner, G.; Hartmann, H.; et al. Standardized protocols and procedures can precisely and accurately quantify non-structural carbohydrates. Tree Physiol. 2018, 38, 1764–1778. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant exctract by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; Lopez, R.; Medlyn, B.E. Triggers of tree mortality under drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef]
- Hartmann, H.; Moura, C.F.; Anderegg, W.R.L.; Ruehr, N.K.; Salmon, Y.; Allen, C.D.; Arndt, S.K.; Breshears, D.D.; Davi, H.; Galbraith, D.; et al. Research frontiers for improving our understanding of drought- induced tree and forest mortality. New Phytol. 2018, 218, 15–28. [Google Scholar] [CrossRef] [Green Version]
- Kanneberg, A.S.; Philips, R.P. Non-structural carbohydrate pools not linked to hydraulic strategies or carbon supply in tree saplings during severe drought and subsequent recovery. Tree Physiol. 2020, 40, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Cernusak, L.A.; Cheeseman, A.W. The benefits of recycling: How photosynthetic bark can increase drought tolerance. New Phytol. 2015, 208, 995–997. [Google Scholar] [CrossRef]
- Karlsson, P.S. Effects of water and mineral nutrient supply on a deciduous and an evergreen dwarf shrub: Vaccinium uliginosum L. and V. vitisidaea L. Ecography 1985, 8, 1–8. [Google Scholar] [CrossRef]
- Hansen, J.; Beck, E. Seasonal changes in the utilization and turnover of assimilation products in 8-year-old Scots pine (Pinus sylvestris L.). Trees 1994, 8, 172–182. [Google Scholar] [CrossRef]
- Piispanen, R.; Saranpaa, P. Variation of non-structural carbohydrates in silver birch (Betula pendula Roth) wood. Trees 2001, 15, 445–451. [Google Scholar] [CrossRef]
- Schadel, C.; Blochl, A.; Richter, A.; Hoch, G. Short-term dynamics of non-structural carbohydrates and hemicelluloses in young branches of temperate forest trees during bud break. Tree Physiol. 2009, 29, 901–911. [Google Scholar] [CrossRef]
- Klein, T.; Vitasse, Y.; Hoch, G. coordination between growth, phenology and carbon storage in three coexisting deciduous tree species in a temperate forest. Tree Physiol. 2016, 7, 847–855. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Vilalta, J.; Sala, A.; Asensio, D.; Galiano, L.; Hoch, G.; Palacio, S.; Piper, F.I.; Lloret, F. Dynamics on non-structural carbohydrates in terrestrial plants: A global synthesis. Ecol. Monogr. 2016, 86, 495–516. [Google Scholar] [CrossRef]
- Furze, M.E.; Trumbore, S.; Hartmann, H. Detours on the phloem sugar highway: Stem carbon storage and remobilization. Curr. Opin. Plant Biol. 2018, 43, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Carbone, M.S.; Czimczik, C.I.; Keenan, T.F.; Murakami, P.F.; Pederson, N.; Schaberg, P.G.; Xu, X.; Richardson, A.D. Age, allocation and availability of nonstructural carbon in mature red maple trees. New Phytol. 2013, 200, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Trumbore, S.; Czimczik, C.I.; Sierra, C.A.; Muhr, J.; Xu, X. Non-structural carbon dynamics and allocation relate to growth rate and leaf habit in California oaks. Tree Physiol. 2015, 35, 1206–1222. [Google Scholar] [CrossRef] [Green Version]
- Piper, F.; Paula, S. The role of nonstructural carbohydrates storage in forest resilience under climate change. Curr. For. Rep. 2020, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Khaleghi, A.; Naderi, R.; Brunetti, C.; Maserti, B.E.; Salami, S.A.; Babalar, M. Morphological, physiochemical and antioxidant responses of Maclura pomifera to drought stress. Sci. Rep. 2019, 9, 19250. [Google Scholar] [CrossRef]
- Salleo, S.; Lo Gullo, M.A.; Trifilò, P.; Nardini, A. New evidence for a role of vessel-associated cells and phloem in the rapid xylem refilling of cavitated stems of Laurus nobilis L. Plant. Cell Environ. 2004, 27, 1065–1076. [Google Scholar] [CrossRef]
- Secchi, F.; Zwieniecki, M.A. Down-regulation of plasma intrinsic protein1 aquaporin in poplar trees. Is detrimental to recovery from embolism? Plant. Physiol. 2014, 164, 1789–1799. [Google Scholar] [CrossRef] [Green Version]
- Trifilò, P.; Casolo, V.; Raimondo, F.; Petrussa, E.; Boscutti, F.; Lo Gullo, M.A.; Nardini, A. Effects of prolonged drought on stem non-structural carbohydrates content and post-drought hydraulic recovery in Laurus nobilis L.: The possible link between carbon starvation and hydraulic failure. Plant. Physiol. Biochem. 2017, 120, 232–241. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | L. nobilis | P. alba | ||
|---|---|---|---|---|
| C | S | C | S | |
| Ncy | 2.7 ± 0.3 | 2.7 ± 0.3 | 7.7 ± 1.1 | 9.3 ± 1.9 |
| (p-value = 1.0) | (p-value = 0.292) | |||
| Lcy (cm) | 13.3 ± 0.9 | 13.4 ± 2.3 | 17.6 ± 3.1 | 13.7 ± 1.7 |
| (p-value = 1.0) | (p-value = 0.201) | |||
| AL (cm2) | 12.2 ± 0.5 | 10.6 ± 0.6 | 17.4 ± 0.9 | 17.1 ± 1.1 |
| (p-value = 0.022) | (p-value = 0.414) | |||
| LMA (g m−2) | 104.0 ± 2.6 | 111.5 ± 2.9 | 41.1 ± 1.5 | 41.6 ± 1.3 |
| (p-value = 0.06) | (p-value = 0.797) | |||
| Species | Time | NL | p-Value | |
|---|---|---|---|---|
| C | S | |||
| L. nobilis | June | 32.3 ± 3.5 | 33.7 ± 5.5 | T: 0.819 |
| L: 1 | ||||
| July | 32.3 ± 3.5 | 33.7 ± 5.5 | T × L: 1 | |
| P. alba | June | 177.3 ± 15.5a | 168.7 ± 9.6a | T: <0.001 |
| L: 0.1 | ||||
| July | 38.7 ± 10.11b | 95.7 ± 3.6c | T × L: <0.05 | |
| Parameter | Time | L. nobilis | P. alba | ||
|---|---|---|---|---|---|
| C | S | C | S | ||
| −Ψtlp (MPa) | June | 1.99 ± 0.05a | 1.97 ± 0.06a | 2.27 ± 0.03 | 2.21 ± 0.10 |
| July | 2.64 ± 0.11b | 2.62 ± 0.10b | 2.19 ± 0.01 | 2.19 ± 0.09 | |
| −πo (MPa) | June | 1.71 ± 0.04a | 1.72 ± 0.04a | 1.73 ± 0.03 | 1.72 ± 0.05 |
| July | 2.07 ± 0.0.3b | 2.08 ± 0.09b | 1.53 ± 0.11 | 1.72 ± 0.09 | |
| εmax (MPa) | June | 27.4 ± 3.6 | 31.2 ± 4.4 | 18.2 ± 4.5 | 25.7 ± 3.5 |
| July | 35.7 ± 2.5 | 37.9 ± 3.1 | 30.1 ± 3.9 | 27.6 ± 4.1 | |
| Parameter | L. nobilis | P. alba | ||||
|---|---|---|---|---|---|---|
| T | L | T × L | T | L | T × L | |
| Ψtlp (MPa) | 43.58 *** | 0.025 | 0.0006 | 0.18 | 0.08 | 0.04 |
| πo (MPa) | 33.91 *** | 0.05 | 0.01 | 1.23 | 0.91 | 1.11 |
| εmax (MPa) | 3.76 | 0.58 | 0.05 | 2.36 | 1.30 | 1.24 |
| Species | Soluble Sugars (mg g−1 DW−1) | Starch (mg g−1 DW−1) | ||
|---|---|---|---|---|
| Midday | Morning | Midday | Morning | |
| L. nobilis | 14.8 ± 7.3 | 13.2 ± 9.7 | 19.2 ± 1.8b | 17.9 ± 2.5b |
| P. alba | 13.1 ± 1.3 | 13.2 ± 1.2 | 6.9 ± 2.3a | 610.5 ± 2.8ab |
| Parameters | Sp | T | Sp × T |
|---|---|---|---|
| soluble NSCs | 0.53 | 0.41 | 0.62 |
| starch | 16.71 *** | 0.215 | 1.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trifilò, P.; Natale, S.; Gargiulo, S.; Abate, E.; Casolo, V.; Nardini, A. Stem Photosynthesis Affects Hydraulic Resilience in the Deciduous Populusalba but Not in the Evergreen Laurus nobilis. Water 2021, 13, 2911. https://doi.org/10.3390/w13202911
Trifilò P, Natale S, Gargiulo S, Abate E, Casolo V, Nardini A. Stem Photosynthesis Affects Hydraulic Resilience in the Deciduous Populusalba but Not in the Evergreen Laurus nobilis. Water. 2021; 13(20):2911. https://doi.org/10.3390/w13202911
Chicago/Turabian StyleTrifilò, Patrizia, Sara Natale, Sara Gargiulo, Elisa Abate, Valentino Casolo, and Andrea Nardini. 2021. "Stem Photosynthesis Affects Hydraulic Resilience in the Deciduous Populusalba but Not in the Evergreen Laurus nobilis" Water 13, no. 20: 2911. https://doi.org/10.3390/w13202911
APA StyleTrifilò, P., Natale, S., Gargiulo, S., Abate, E., Casolo, V., & Nardini, A. (2021). Stem Photosynthesis Affects Hydraulic Resilience in the Deciduous Populusalba but Not in the Evergreen Laurus nobilis. Water, 13(20), 2911. https://doi.org/10.3390/w13202911