Abstract
The Gulf of La Spezia (GLS) in Northwest Italy is a rocky embayment with low fluvial influence facing the Mediterranean Sea. Past landscape dynamics were investigated through a multi-proxy, facies-based analysis down to a core depth of 30 m. The integration of quantitative ostracod, foraminifera, and pollen analyses, supported by radiocarbon ages, proved to be a powerful tool to unravel the late Quaternary palaeoenvironmental evolution and its forcing factors. The complex interplay between relative sea-level (RSL), climatic changes, and geomorphological features of the embayment drove four main evolution phases. A barrier–lagoon system developed in response to the rising RSL of the Late Pleistocene (likely the Last Interglacial). The establishment of glacial conditions then promoted the development of an alluvial environment, with generalised erosion of the underlying succession and subsequent accumulation of fluvial strata. The Holocene transgression (dated ca. 9000 cal year BP) caused GLS inundation and the formation of a low-confined lagoon basin, which rapidly turned into a coastal bay from ca. 8000 cal year BP onwards. This latter environmental change occurred in response to the last Holocene stage of global sea-level acceleration, which submerged a morphological relief currently forming a drowned barrier-island complex in the embayment.
1. Introduction
Mediterranean coastal areas include a variety of aquatic and wetland environments and habitats (swamps, marshes, lakes, lagoons, bays, etc.) that deserve to be protected against the threat of global change through the application of rigorous conservation or restoration plans, based on both monitoring techniques and geological data [1,2,3]. Being located at the edge between land and sea, coastal zones provide a wide range of resources (biodiversity hotspots, archaeological sites), commercial activities (fishing industries, port infrastructures), and urban areas. On the other hand, subsurface successions in coastal regions preserve precious information about past environmental responses to natural and anthropogenic impacts, including changes in climate conditions and RSL, as well as amounts, types, and paths of sediments delivered to the sea. Thus, in-depth knowledge of late Quaternary dynamics and related forcings represent a key step towards proper management and sustainable development of these highly valuable and vulnerable areas.
It is not by chance that studies looking at Mediterranean coastal successions have dramatically increased during the last decade, taking advantage of the availability and good preservation of palaeobiological proxies (molluscs, foraminifers, ostracods, pollen, plant macroremains, etc.). If framed into a stratigraphic perspective, these fossils are known to be extremely useful to achieve robust and detailed palaeoenvironmental (facies-based), vegetation-derived palaeoclimate, and chronological (mainly 14C age-based) reconstructions [4,5,6,7,8,9,10,11,12]. However, multidisciplinary studies are generally restricted to Holocene successions and the integrated analysis of superposed transgressive-regressive (T-R) Quaternary depositional cycles is still scarce [13,14,15,16]. Besides, palaeoenvironmental reconstructions of embayments/lagoons located along rocky shorelines or of drowned estuaries are relatively scarce [11,17,18,19], since wide, low-lying coastal plains have drawn the most attention. Geomorphological studies of rocky shores are rapidly increasing, especially in the Mediterranean, where rocky cliffs, platforms, and rock pools represent more than 50% of coastline habitat types [20]. Rocky shores include several landforms such as platforms/marine terraces, tidal notches, and cliffs, which can provide potential RSL markers or data in terms of erosion rates and decay/shaping processes [20,21,22,23,24].
In this study, we present a stratigraphic-based reconstruction of late Quaternary landscape dynamics from the Gulf of La Spezia (Figure 1) based on the integration of sedimentological, benthic foraminifers and ostracods, and pollen data. GLS is a relatively deep embayment located along the Ligurian rocky coastline (Figure 1). It is locally interrupted by coastal plains of limited extent whose subsurface stratigraphy is only partially known [25]. The gulf area, which is currently affected by intense anthropization due to the commercial port of La Spezia (Figure 2), represents a particular coastal, shallow marine setting protected by steep promontories and almost deprived of river inputs. In 2016, a drilling campaign, requested by the Italian Superintentency, was conducted as part of the rescue archaeology survey associated with an important infrastructural harbour project. This campaign gave us the opportunity to investigate the shallow (30 m) subsurface of the innermost portion of the GLS (Figure 2A,B). The present study provides new insights into the late Quaternary evolution trends of depositional environments and vegetation patterns of rocky coastal tracts.
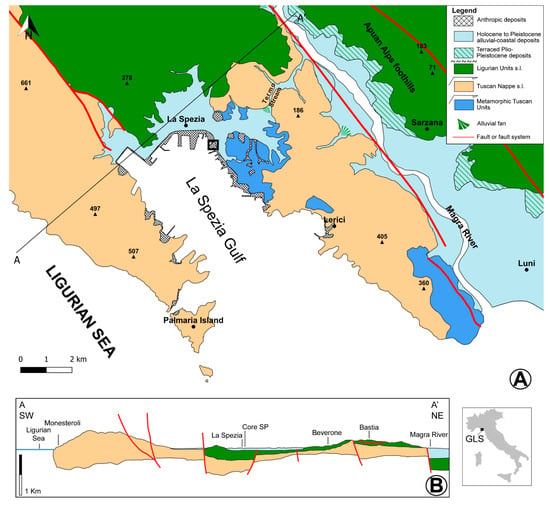
Figure 1.
(A) Geological and geomorphological sketch map of the Gulf of La Spezia (GLS) and the surrounding areas; modified from Sheet 248–CARG Project and [21]. The inset shows the site of the coring campaign (see Figure 2). (B) Simplified geological cross-section modified from Sheet 248–CARG Project. Transect A-A’ is shown in Figure 1A. The location of the studied core SP is projected on the section.

Figure 2.
Close-up view of the harbour area of GLS (A) and its eastern terminal, where the coring campaign (cores are reported as white dots) was undertaken (B). The studied core SP is highlighted in red; other cores discussed in the text are highlighted in white. Basemap (aerial photos) from the “Geoportale della Regione Liguria” [26].
The main purpose of our research is to document the depositional and environmental variability of rocky embayments, highlighting the impact of late Quaternary climate and sea-level changes on the variety of subaqueous and humid rocky shore settings. A particular aim of this study is to clarify the complex interrelationship between coastal palaeo-plant, faunal communities, and sedimentary dynamics within a sheltered setting influenced by low fluvial impact.
2. Study Area
2.1. Geology and Geomorphology
The Gulf of La Spezia represents a marked indentation in the easternmost portion of the Ligurian rocky shoreline in NW Italy (Figure 1), which is characterised by steep slopes, seacliffs, and platforms with local embayments. The latter include narrow coastal plains with sandy-to-gravelly pocket beaches formed under the predominant action of sea currents (prevailing winds blowing from south and south-west; [27]), and the contribution of alluvial fluxes from the hinterland. Especially in E Liguria, reliefs are so close to the plains that altitudes of 700–800 m a.s.l. are reached in less than 5 km inland from the shore [28]. The Ligurian coastline has a microtidal regime and the tidal range does not exceed 0.4 m [29].
The GLS is ~3–4.5 km wide and ~10–15 m deep [30]. It is bordered by two NW-SE oriented promontories (i.e., headlands of the gulf; Figure 1) belonging to the Northern Apennines, a fold-and-thrust belt developed since the Late Cretaceous in response to the Adria-Europe plate convergence [31,32,33]. The eastern of these promontories, predominantly composed of Mesozoic limestones and Late Oligocene-Early Miocene turbidite sandstones (Tuscan Nappe), separates the semi-graben of GLS from that of the Magra River (Figure 1B; [34]). The graben systems, which have developed since the Late Miocene-Early Pliocene, are characterised by a set of faults with predominant NW-SE orientation, some of which have been interpreted as being still active during the Quaternary period [35]. Ligurian and sub-Ligurian Units of the Late Cretaceous-Paleocene age crop out a few hundreds of meters inland of the present shoreline (Figure 1).
At present, GLS does not receive any significant fluvial inputs, as only channelized streams flow in the area. Accordingly, the mean salinity of the gulf is ~37‰ [27]. Possible Pleistocene alluvial terraces are widely recognised along the stream valleys, documenting a phase of predominant fluvial incision on pre-existing alluvial deposits (mainly pebbles, gravels, and sands) that may have resulted from climate oscillations (i.e., glacial periods). These geomorphological-stratigraphic features, along with the occurrence of an inactive alluvial fan in the eastern portion of GLS (Termo alluvial fan; Figure 1A), point to a palaeodrainage network feeding the area during the Pleistocene [35]. Unfortunately, no age determinations are available from these alluvial deposits, thus preventing any further interpretations.
Few data are available from the subsurface of the narrow coastal plain, while published stratigraphic data (e.g., cores, trenches) refer almost exclusively to the urban area of La Spezia (Figure 1 and Figure 2), where up to 15–20 m of fine-grained deposits locally rest on tens of meters of sand, gravel, and pebbles which, in turn, overlie the Mesozoic limestones [36]. In the submerged part of the gulf, seismic lines calibrated through cores highlight the occurrence of a complex palaeochannel network and a NE-SW elongated sandy bar located ~3 km from the present coastline, at a water depth of ~13 m. Chronology and stratigraphic relations between these two morpho-stratigraphic features are still largely unknown. However, the bar is likely to have developed after the palaeochannels, leading to the establishment of a protected setting in the gulf [30,35].
2.2. Climate and Vegetation
The climate of the GLS is Mediterranean/sub-Mediterranean, with hot summers and precipitations prevailing in the autumn-winter months. Mean temperature is 15 °C and mean precipitation is ~950 mm/year, as recorded at the weather station in La Spezia [37]. The gulf area is influenced by the Atlantic bioclimate, as the easternmost Ligurian coastline corresponds to the northern limit of the Mediterranean bioclimate [38,39]. Peculiar geomorphological features, such as steepness and orientation of the reliefs and proximity of the reliefs to the sea (Figure 1), result in a complex pattern of vegetation types in the Ligurian region. The littoral area is characterised by a meso-Mediterranean vegetation corresponding to a mosaic of Mediterranean maquis (e.g., Quercus ilex, Juniperus oxycedrus, Erica arborea) and open meso-hygrophilous woodlands (Alnus glutinosa, Fraxinus oxycarpa, Populus, Ulmus minor). Forests of Quercus deciduous, Carpinus betulus, and Ostrya carpinifolia occur on the nearby foothills [28]. In the study area (GLS), natural coastal wetlands have been progressively destroyed by intense urbanization that accelerated during the last century as a result of the expansion of the commercial port of La Spezia.
3. Materials and Methods
The stratigraphic record of the innermost portion of the GLS was studied through the recovery of several sedimentary cores, drilled down to a maximum depth of 40 m and at water depths ranging between about 13.5 m below mean sea-level and 1.6 m above mean sea-level (Figure 2). The cores were closely spaced in a regular grid distribution across the eastern platform area of La Spezia harbour (Figure 2B). The continuous coring system adopted has guaranteed an almost undisturbed stratigraphy and a high recovery percentage of about 80–90%. The description of the sediment cores stored at the Centro Agricoltura e Ambiente—CAA G. Nicoli laboratory included grain size, colour, primary sedimentary structures (if visible), type of contacts between lithofacies, and the occurrence of a variety of accessory materials such as plant remains, mollusc shells or fragments, layers rich in decomposed organic matter, and pedogenic features (Fe-Mn oxides, calcareous nodules).
The 30 m-long core SP (Figure 2B; 44°6′22″ N, 9°51′23″ E) was selected for detailed palaeoenvironmental and sedimentary facies analysis owing to its high lithofacies variability and abundance of clay and silt, where rich palaeobiological assemblages were expected. In order to reach a high degree of resolution in facies characterization, the multi-proxy study combined sedimentological data with the quantitative analysis of the microfossil content, including benthic foraminifers and ostracods as well as pollens and spores. Combining benthic foraminifer and ostracod data is known to provide robust information about even the most subtle changes in a variety of parameters (e.g., salinity, hydrodynamic conditions, vegetation cover, turbidity, and amount/type of organic matter), especially within transitional-coastal settings [40,41,42,43]. Pollens and spores are valuable indicators of the palaeovegetation communities growing in the low-lying areas close to the coastline [9,11,12,15,44]. Taking into account the proximity of the surrounding reliefs (Figure 1), the regional signal of the pollen rain can be particularly strong [28], allowing the reconstruction of climate-related vegetation dynamics.
Twenty-six and 28 samples were collected for meiofaunal and palynological analyses, respectively. All the lithofacies identified from the visual inspection of reference core SP were sampled, focusing on fine-grained, organic-rich intervals, and maximizing the integration of palaeobiological data from the same stratigraphic interval. Meiofauna samples (~50 g of dry weight) were treated by following standard procedures adopted for subsurface Quaternary successions of the Mediterranean area [45,46]. This included: (i) soaking dry samples in water or water + H2O2 (35% vol.) for 24 h; (ii) wet-sieving at 63 µm; and (iii) oven-drying of the residue. Samples containing autochthonous meiofauna (i.e., well-preserved specimens or valves) were quantitatively analysed in the size fraction >125 µm. Where possible, at least 300 foraminifers were counted. Both adult and juvenile ostracod valves with distinct morphological features that allowed identification at species level were counted in the same foraminiferal sub-sample (carapaces were considered as two valves). The meiofauna counts are reported in Table S1 of the Supplementary Material. A list of references used for taxonomic identification and species autoecological information is provided in Table S2. To fully capture the variations in meiofauna composition and structure, two diversity indices were calculated using the PAST (PAlaeontological Statistics ver. 4.04) data analysis package [47]. The α-index [48], which guarantees a reliable account of rare species and the Shannon–Weaver index (H), which corresponds to a measure of entropy (i.e., distribution of taxa among the total individuals [49]), were applied.
Palynological samples were prepared following a standard extraction technique [50], with only minor modifications. About 8–10 g of sample was treated in 10% Na pyrophosphate to deflocculate the sediment matrix; a Lycopodium tablet was added to calculate pollen concentration; the sediment residue was subsequently washed through a 7 μm sieve. Samples were resuspended in 10% HCl to remove calcareous material and subsequently acetolysed. Heavy-liquid separation was performed using Na metatungstate hydrate with a specific gravity of 2.0, followed by centrifugation at 2000 rpm for 20 min. The retained fractions were treated with 40% HF for 24 h, and the sediment residue was washed in distilled water and then in ethanol with glycerol. The final residue was desiccated and mounted on slides using glycerol jelly and sealed with paraffin.
Identification and counting of the pollen grains was performed at x400 magnification. For each sample, where possible, 300 pollen grains were counted. Pollen percentages were calculated on the basis of the total pollen sum (Table S3). Seven main pollen groups were identified according to their vegetational and ecological characteristics, taking into account the regional versus local components of the pollen rain. The Montane taxa group mainly comprises taxa living at medium and/or high altitudes in the Apennines [51], such as Alnus cf. viridis, Betula, Fagus sylvatica, Abies alba, Pinus sylvestris, and Pinus mugo. Quercus and the group of other deciduous trees correspond to the mixed oak forest that can be found in association with Mediterranean or sub-Mediterranean woods [51]. It is composed of Quercetum taxa such as Quercus deciduous, Carpinus betulus, Corylus avellana, Fraxinus, Ostrya carpinifolia, Tilia, Ulmus, and other deciduous trees. The Mediterranean group includes taxa belonging to the Mediterranean evergreen and sclerophyllous forest such as Quercus ilex, Pinus cf. pinea, Erica, and Myrtus. Pasture/meadow herbs mainly include Poaceae, Fabaceae, and Cichorioideae. The group of herbaceous hygrophytes and aquatics (hygro+aquatic) was also distinguished; it includes hygrophilous herbs (e.g., Cyperaceae), helophytes, and hydrophytes (i.e., Butomus, Typha, Lemna, Callitriche, Myriophyllum, Nymphaea, Potamogeton, and Sparganium), which are considered a good proxy for wetlands conditions. Alnus cf. glutinosa, Populus, and Salix compose the group of hygrophilous trees that can tolerate only short periods of inundation. Halophytes represented by Artemisia and Amaranthaceae are indicators of brackish conditions.
Pasture/meadow, hygro + aquatic and halophyte herbs, as well as hygrophilous trees, represent the main components of the local pollen rain, mainly reflecting environmental conditions. Montane taxa, Mediterranean taxa, and Quercus + other deciduous trees typify the regional signal of the pollen rain. References for taxonomic identification and ecological characterization of pollen taxa are shown in Table S2. Monilophyta spores were also identified and counted as number of spores/g. Undeterminable, poorly preserved (crumpled, fragmented) grains and secondary grains (i.e., redeposited from older deposits), characterised by enhanced reddening due to acetolysis or belonging to extinct taxa, were excluded from the total pollen sum. Both types of grains proved to be scarce within the studied succession, according to a generally low fluvial influence on the embayment.
The chronological framework of the studied succession was obtained through seven AMS radiocarbon ages derived from key stratigraphic layers of the reference core SP (Table 1). Conventional ages were calibrated using the CALIB 8.2 program [52] along with the IntCal20 or Mixed Marine NoHem (50% of marine component) calibration curves of [53] for organic matter derived from terrestrial or transitional settings, respectively.

Table 1.
Radiocarbon ages from core SP (see Figure 2 for location).
4. Results
4.1. Facies Associations
The eight facies associations identified in this study are based on the integration of sedimentological and palaeobiological (i.e., meiofauna and pollen taxa/groups strictly related to the local pollen rain) data from the reference core SP, which is considered representative of the 30 m-thick subsurface succession of the study area (Figure 2). Facies description and interpretation of depositional environments, ranging from alluvial to shallow-marine systems, are reported below. Depth intervals refer to core SP stratigraphy (Figure 3, Figure 4 and Figure 5; Tables S1 and S3).
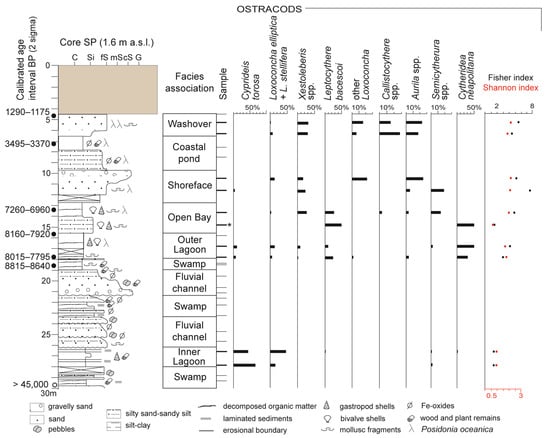
Figure 3.
Stratigraphy, facies characterization, and vertical distribution of main ostracod taxa and diversity indices of core SP. Samples with an autochthonous ostracod fauna are reported as bold lines. One sample containing less than 20 valves is highlighted by an asterisk (*). C: Clay; Si: Silt; fS: Fine sand; mS: Medium sand; cS: Coarse sand; G: Gravel. The uppermost 4 m are composed of anthropogenic materials probably due to harbour construction.
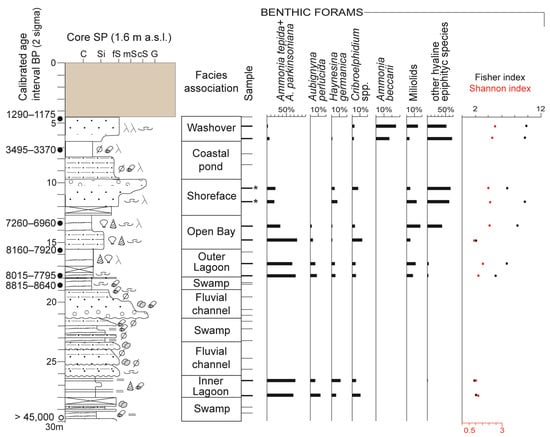
Figure 4.
Stratigraphy, facies characterization, and vertical distribution of main benthic foraminiferal taxa and diversity indices of core SP. Samples with autochthonous foraminiferal fauna are reported as bold lines. Two samples containing less than 300 specimens are highlighted by an asterisk (*). C: Clay; Si: Silt; fS: Fine sand; mS: Medium sand; cS: Coarse sand; G: Gravel. The uppermost 4 m are composed of anthropogenic materials probably due to harbour construction. See legend in Figure 3 for lithological information.
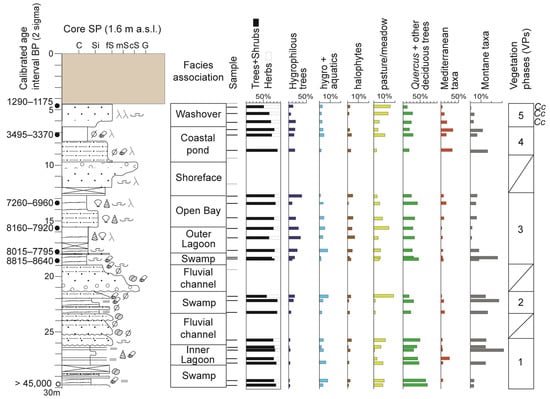
Figure 5.
Stratigraphy, facies characterization, and vertical distribution of main pollen groups of core SP. Samples with countable pollen grains are reported as bold lines. The vegetation phases are also shown. Cc indicates the presence of cultivated plants in the pollen record. C: Clay; Si: Silt; fS: Fine sand; mS: Medium sand; cS: Coarse sand; G: Gravel. The uppermost 4 m are composed of anthropogenic materials probably due to harbour construction. See legend in Figure 3 for lithological information.
4.1.1. Fluvial-Channel Facies Association (~26–23.50 m; ~21.50–19 m)
Description
This facies association, which has a maximum thickness of 2.5 m, consists of light- grey to yellow-brown, fine-sand bodies or gravelly sand to medium-fine sand bodies both grading upwards into silty sands and sandy silts (i.e., fining-upward trend—FU trend). The lower boundary is erosional and may exhibit pebble layers. Wood and other plant fragments, unidentifiable fragments of mollusc shells, and mottles produced by Fe oxides are found locally, especially within silty intercalations. Only a few samples were collected from this facies association. Owing to their coarse grain size, the samples are barren of both pollen and autochthonous meiofauna, with the exception of the sample collected at ~25.75 m core depth. This sample shows a rich pollen assemblage, characterised by considerable percentages of pasture/meadow pollen taxa (~10.5%), with the second-level occurrence of hygrophilous tree pollen (4.5%).
Interpretation
Based on its diagnostic sedimentological features (granulometry; vertical grain size variations-FU trend; lower erosional surface) and absence of meiofauna, this facies association is interpreted as sandy fluvial-channel bodies occasionally fed by coarser (i.e., gravel) inputs. Taking into account the geological-geomorphological setting, these deposits could be related to a fluvial network developed at the fringe of alluvial fans.
4.1.2. Swamp Facies Association (~30–28 m; ~23.50–21.50 m; 19–18 m)
Description
This facies association, ~1–2 m-thick, is composed of grey-to-dark grey, organic-rich silts and sandy silts locally interrupted by mm- to cm-thick sandy layers, at places showing an internal FU trend. Small-sized, well-preserved plant remains occur within mm-thick layers giving rise to a laminated deposit. Few scattered Fe-oxides and unidentifiable fragments of mollusc shells are also encountered. Meiofauna samples collected from this facies association are barren or contain few, poorly preserved benthic foraminifers and ostracods belonging to Ammonia beccarii, Haynesina germanica, Cribroelphidium ex gr. poeyanum, and Cytheridea neapolitana.
The total pollen and spore concentration is highly variable within this interval, ranging between 1.3 k and 69 k grains/g. The local pollen rain is mainly characterised by considerable percentages of hygro + aquatic herbs (<7%) and hygrophilous trees (~1–14.5%) with the co-occurrence of pasture-meadow taxa (<7.5%), almost exclusively represented by Poaceae, that can become co-dominant at places. Within the hygro + aquatic group, Cyperaceae (hygrophytes) and Myriophyllum spicatum (hydrophytes) are the most common pollen taxa; Alnus and subordinately Populus mainly represent the hygrophilous trees. Halophytes pollen taxa show percentages < 3%.
Interpretation
This facies association records the deposition of fine-grained sediments and organic detritus in a stagnant, paludal environment, such as a swampy area. The considerably abundant occurrence of hygrophyte and aquatic plants along with Poaceae (wild grass), pasture/meadow taxa tolerant of wet conditions, and the variable amount of Polypodiophyta spores strongly support this interpretation, suggesting a low, oscillating water table. The absence of an autochthonous meiofauna reflects standing and hypohaline waters, as also documented by the very low abundance of halophytes. On the other hand, the occurrence of transported brackish-marine meiofauna species suggests relative proximity to the sea. Individual sand layers are interpreted to reflect episodic terrigenous inputs from inland locations.
4.1.3. Coastal pond Facies Association (~9.80–6.50 m)
Description
This facies association includes ~1.5 m of mottled, grey silty fine sands abruptly passing upwards to a layer of ~1.5 m-thick silty clays containing sparse unidentified plant remains as well as Posidonia oceanica (Neptune grass) fibers. Samples are barren of meiofauna. The average pollen and spore concentration is generally below 3k grains/g (with a peak at 14k grains/g) and 1.5k grains/g, respectively. Hygrophilous trees represent the most abundant local pollen group (~14–17%), showing a dominance of Alnus cf. glutinosa and subordinately Populus. Pollens of hygrophyte herbs (Cyperaceae and Ranunculaceae) occur at low percentages (<3%), while aquatic herbs are almost absent. Pasture/meadow herbs show highly variable pollen abundances, between ~1.5 and 10.5%, and are mainly represented by Poaceae (wild grass). Halophytes show percentages < 4%.
Interpretation
The abundance of plant remains, including Posidonia, and the dominance of hygrophilous pollen taxa point to a slightly depressed, generally wet area close to the coeval coastline, but shielded from the direct influence of saline waters and subject to periodic subaerial exposure (i.e., water table oscillations), as testified by the diffuse presence of Fe-oxides. All these features suggest a coastal pond located behind a beach sand bar. The observed FU trend may document a progressively increasing distance from the shoreline, with the establishment of more protected conditions.
4.1.4. Inner Lagoon Facies Association (~28–26 m)
Description
This facies association is formed by a dark grey, laminated clay-silt succession containing mm-thick silty-sandy layers. Plant remains and shell fragments of brackish-marine gastropods (e.g., Cerithium sp.) are abundant. Few gypsum crystals and rosettes are also encountered. Samples contain an abundant, but low diverse meiofauna (low values of α-index; Figure 3 and Figure 4), dominated by few species (low values of H index; Figure 3 and Figure 4). The assemblage is mainly composed of the euryhaline, opportunistic taxa Ammonia tepida-Ammonia parkinsoniana (~70%), Cyprideis torosa (~45–67%), and Loxoconcha elliptica + L. stellifera (15–48%), with the second-level occurrence of Haynesina germanica, Aubignyna perlucida, Cribroelphidium species (mainly C. ex gr. poeyanum), Leptocythere and Semicytherura species (mainly L. psammophila and S. sulcata). Total pollen and spore concentration ranges between ~5k and 7.5k grains/g and 1k–1.8k grains/g, respectively. The pollens of hygrophilous trees and hygro + aquatic herbs show variable percentages, between ~1 and 4%, and 0.7 and 5%, respectively. Pasture/meadow herbs pollen locally reach considerable abundance of ~7% (mainly Poaceae). Halophytes taxa show percentages <2%.
Interpretation
The distinctive sedimentological features (i.e., fine-grained deposits, dark colour, and abundance of plant remains) and the meiofauna dominated by euryhaline species indicate that this facies association was formed in a low-energy, semi-protected brackish environment, namely a lagoonal basin. The C. torosa+Ammonia tepida-A. parkinsoniana assemblage is considered a sensitive and reliable indicator of back-barrier settings subject to salinity oscillations in the Mediterranean area ([54] and references therein). More specifically, the low species diversity accompanied by the paucity of brackish-marine taxa and halophytes point to restricted, highly confined conditions typical of an inner lagoon surrounded by wet grasslands, as suggested by the remarkable abundance of Poaceae pollen. However, the absence of freshwater-hypohaline ostracods and the co-occurrence of gypsum crystals document a negligible degree of river influence on the basin.
4.1.5. Outer Lagoon Facies Association (~18–15.50 m)
Description
This 2.5 m-thick-facies association is formed by a homogeneous succession of dark-to-light grey soft silty clays, containing brackish-marine mollusc shells (e.g., Cerithium sp.), echinoderm spines, and thin layers of decomposed organic matter. Crystals and rosettes of gypsum and few Posidonia oceanica fibers are documented. The meiofauna is abundant, rich, and quite diverse, as documented by the remarkably high values of both α-index and H index (Figure 3 and Figure 4). The foraminiferal assemblage is dominated by Ammonia tepida-Ammonia parkinsoniana (~67–74%), with the second-level occurrence of miliolids (~7–12%) and A. perlucida (~7–8%). Among the miliolids, Adelosina and Lachnella species are the most common. Cytheridea neapolitana, a euhaline mud-loving species commonly thriving in the shallow marine environment enriched with organic matter, is the dominant ostracod (~30–50%). This is accompanied by the euryhaline C. torosa and L. elliptica (~12–25% as a total) along with a set of brackish-marine taxa including Leptocythere bacescoi (~9–23%) and Xestoleberis spp. (<6%). A local peak in the euhaline Carinocythereis whitei, reaching about 24%, is also found. Pollens and spores display a high total concentration, between 28k and 109k grains/g and 10k–35k grains/g, respectively. Pollens of hygrophilous trees, in particular Alnus cf. glutinosa, are very abundant (~23–33%). The accompanying humid herbaceous community includes hygro + aquatics (~3–7.5%) and Poaceae (~3–13%). Halophytes reach up to ~7% of relative abundance.
Interpretation
This facies association is inferred to have been deposited in a semi-enclosed, calm basin characterised by brackish to marine waters and low confined conditions typical of an outer lagoon. A good connection with the open sea is testified by the richness of the meiofauna and by the remarkable abundance of euhaline ostracods, miliolids, and halophytes. The development of wide paludal areas on land is evidenced by the high percentages of hygrophilous + aquatic plants.
4.1.6. Open bay Facies Association (~15.50–12.60 m)
Description
This 3 m-thick-facies association consists of light grey sandy silts passing upwards to silty clays containing several fragments and shells of brackish-marine molluscs (e.g., Cerithium sp.) and echinoderm spines. Thin organic-rich layers and sparse Posidonia fibers are recorded locally. Few, small-sized gypsum rosettes are encountered. The foraminiferal assemblage is abundant throughout the facies, but a different composition distinguishes the lower silty portion from the overlying clays (Figure 4). Silts contain a poorly diversified fauna (α-index is ~2), strongly dominated by Ammonia tepida-Ammonia parkinsoniana (~78%; H index is ~0.9). Only a few other taxa occur, with Cribroelphidium spp. (mainly C. ex gr. poeyanum) showing ~13% of relative abundance. The overlying clayey portion includes a more diversified assemblage (α-index is ~8) containing high percentages of hyaline epiphytic taxa (~40%), mainly represented by Planorbulina mediterranensis and, to a lesser extent, by Rosalina species. Ammonia tepida-Ammonia parkinsoniana (~34%) and Miliolids (~18.5%) dominated by the epiphytic Siphonaperta aspera are also well represented.
Similarly, the richness of the ostracod fauna shows an upward increase (Figure 3). Within the lower portion, ostracods are almost absent and only two opportunistic species are encountered (L. bacescoi and C. neapolitana). In contrast, the clayey portion contains several species (α-index of ~5) with the dominance (~55%) of brackish-marine (Xestoleberis spp. and L. bacescoi) taxa, accompanied by euhaline ostracods mainly belonging to the genera Semicytherura (~12%), Callistocythere, and Aurila (~3% each). Pollen shows relatively high total concentrations, ranging between ca. 16k and 30k grains/g. Alnus cf. glutinosa and Populus pollen reach remarkable abundances (~9–38%) and suggest a dominance of both taxa in the vegetation. Pollen of hygro + aquatic herbs and halophytes show relative percentages <4%. Poaceae, which represents the main taxon of the pasture/meadow group, accounts for ~3.5–8%. Spores display a total concentration of ~15k and 25k grains/g, similar to pollen.
Interpretation
The absence of C. torosa and the co-dominance of brackish-marine and euhaline ostracods, the latter thriving in shallow-marine environments, point to a transitional setting characterised by very high rates of water exchange with the sea and salinity close to marine values. Benthic foraminifers highlight an upward decrease in the degree of confinement, which is reflected by the substitution of Ammonia tepida-Ammonia parkinsoniana with epiphytic taxa. This suggests the establishment of an open-bay environment with a vegetated bottom of Posidonia oceanica meadows. This interpretation is consistent with the sedimentological features, since fine-grained sedimentation is enhanced by the morphological configuration of GLS (i.e., promontories provided protection against sea currents; Figure 1). In the pollen record, the abundance of hygrophilous woody taxa may document the presence of wooded wetlands surrounding the bay.
4.1.7. Shoreface Facies Association (~12.60–9.80 m)
Description
A coarsening-upward (CU) succession formed by light grey sandy silts, fine sands, and gravelly sands characterises this 3 m-thick-facies association. Scattered fragments of mollusc shells, gypsum rosettes, and abundant fibers of Posidonia are encountered. The meiofauna is rather scarce, but with a large number of species (α-index > 5; Figure 3 and Figure 4). Among the foraminifers, hyaline epiphytic taxa are dominant (~60%; Rosalina and Elphidium species), with the second-level occurrence of Ammonia tepida-Ammonia parkinsoniana and miliolids. The ostracod fauna is made up of phytal transitional-coastal taxa (Xestoleberis, Loxoconcha and Aurila species) and euhaline taxa of shallow-marine environments (Palmoconcha and Semicytherura species). The former taxa become dominant within the uppermost, coarser layers. The pollen in this facies association is extremely scarce.
Interpretation
Sedimentological features and meiofauna content point to a high-energy, shallow-marine environment with a vegetated bottom, such as, for example, a shoreface. The upward increase in grain size documents a shallowing trend, consistent with the concomitant increase in transitional-coastal ostracods.
4.1.8. Washover Facies Association (~6.50–4.50 m)
Description
This 2-m-thick-facies association consists of dark-to-light grey sandy silts grading upwards into fine sands (CU trend). Posidonia fibers are very abundant throughout the succession. Scattered mollusc fragments are recorded. Meiofauna is abundant (especially benthic foraminifers) and highly diversified (Figure 3 and Figure 4), as indicated by the high values of the α-index for both benthic foraminifers (α-index ~5) and ostracods (α-index ~9). The H index, which reaches a value of ~2, suggests the co-dominance of various taxa including several hyaline epiphytic species (mainly Ammonia beccarii, Rosalina and Elphidium species; for a total of ~50–60%) and miliolids (mainly Adelosina and Lachnella; for a total of ~3–15%) among benthic foraminifers, and phytal ostracods of the genera Xestoleberis, Aurila, and Loxoconcha.
Both pollens and spores show low concentrations between 2k and 3k grains/g. A sample from the uppermost sandy layer was barren of pollen and spores. The pollen of hygrophylous trees (Alnus cf. glutinosa and Populus) and of pasture/meadow herbs (Poaceae and to a lesser extent Cichorioideae and Fabaceae) co-dominate the local pollen rain, with relative abundances: 7.5–20% and 4.5–16.5%, respectively. The pollen of hygro + aquatic herbs and halophytes is scarce (<2%).
Interpretation
Sedimentological characteristics (thickness, CU trend), the pervasive occurrence of Posidonia fibers, and the presence of a highly diversified meiofauna, dominated by taxa preferring shallow-marine environments with highly vegetated sandy bottoms, indicate a coastal setting subject to high-energy events. The stratigraphic position of this facies association, enclosed between anthropogenic materials and coastal pond deposits, allows to refine the interpretation as that of washover deposits formed behind the shoreline under the action of storms. The meiofauna may have been transported inland by storms along with the Posidonia fibers. Accordingly, the pollen data indicate a paucity of hygro + aquatic herbs and a co-dominance of hygrophilous trees and Poaceae, suggesting wet, but not permanently submerged, conditions.
4.2. Vegetation Phases
Five vegetation phases (VPs 1–5) were identified based on the three pollen groups that best depict the regional component of the vegetation: (i) Quercus + other deciduous trees; (ii) Mediterranean taxa; and (iii) Montane taxa. These five vegetation phases are strictly connected to changing palaeoclimate conditions in the GLS area, at least at the scale of glacial-interglacial variability (Figure 5). Throughout the cored succession, arboreal pollen (AP) is invariably dominant with respect to the non-arboreal pollen (NAP); the only exception is the sample at 4.70 m core depth, where substantial equality is recorded. The VPs described below follow the stratigraphic order. The radiocarbon ages available suggest a pre-Holocene age (Late Pleistocene?) for VPs 1 and 2.
4.2.1. VP 1
This is the oldest vegetation phase, recorded at a core depth between ca. 30 and 26 m. It is characterised invariably by high percentages of trees (ca. 75–87%) and by the dominance of Quercus + other deciduous trees (>40%) relative to Mediterranean (commonly < 6%) and Montane taxa (commonly < 17%). This pollen composition points to a mixed oak forest consistent with an interglacial period affected by mild conditions and relatively wet summers (i.e., low degree of mediterraneity). The sample at 27.40 m core depth shows a peak in Mediterranean taxa (e.g., Quercus ilex) that reach 25% of relative abundance; however, this value is considered to be overestimated since it is affected by a very low pollen counting (40 counted pollen grains). High percentages of pioneer deciduous trees (Ostrya carpinifolia+Corylus avellana: ~25–50%) at the base of VP1 and their progressive replacement by Quercus species, such as Quercus cf. robur, Q. cf. petraea and Q. caducifolia, document the natural achievement of Potential Natural Vegetation (PNV; sensu [55]).
Throughout VP 1, Quercus + other deciduous trees show an upward decreasing trend (from ~70% to 40%), which is paralleled by a sudden increase in Montane taxa (from ~3% up to 20–30%), mainly Abies alba. A peak in Pinus cf. sylvestris (~14%) is also recorded at ~26.6 m core depth. This indicates a vegetation turnover resulting in an expansion of montane trees toward the plain, reasonably linked to an overall change towards cooler (and wetter) conditions. Morphological features of GLS (i.e., proximity of the reliefs to the sea) might have enhanced the montane pollen signal.
4.2.2. VP 2
This vegetation phase, recorded at a core depth of ~23–21 m, shows very high percentages of trees pollen (~90%), although a relative drop (~60%) is recorded close to the upper zone boundary, where the expansion of Poaceae and hygro + aquatic herbs suggests an increase in local humidity. The decrease in Quercus + other deciduous trees (~18–22%) leads to a substantial co-dominance with the Montane taxa (~14–28%), mainly represented by A. alba; Mediterranean taxa (i.e., Q. ilex) occur with highly variable abundances (~2–10%). This composition documents the establishment of a mixed forest mainly composed of both cool-temperate and meso-thermophilous trees. The relative abundance of montane trees points to cool (interstadial) climate conditions that likely stabilized during the last glacial period. Because of its proximity to the sea and morphological configuration (embayment protected by high promontories; Figure 1), GLS may have acted as an important tree refuge during this period.
4.2.3. VP 3
This vegetation phase, recorded between ~19 and 12 m core depth, is characterised by a high percentage of trees (~64–86%; frequently >80%), with the dominance of Quercus + other deciduous trees (~30–45%), mainly represented by Q. cf. robur, Q. caducifolia, C. avellana and O. carpinofoglia. Mediterranean taxa, essentially corresponding to Q. ilex, display variable abundances (~2–14% range). Montane taxa show a generally decreasing trend from ~17 m core depth upwards (<6%); interestingly, a peak of 26% (almost entirely accounted for by A. alba) is recorded at ca. 18.5 m core depth. This pollen assemblage reflects the progressive spread of a mixed oak forest in the plain. Consistent with radiocarbon dates, which constrain this interval between about 8700 and 7000 cal year BP, VP 3 is interpreted to reflect the Holocene climatic optimum. The variable amounts of Q. ilex pollen suggest a low degree of mediterraneity, whereas the montane component of the vegetation reflects, in part at least, proximity to the promontories.
4.2.4. VP 4
This vegetation phase is encountered at a core depth between ~9 and 6.5 m, where the arboreal pollen is very high (~75–92%). The main feature is the marked expansion of Q. ilex pollen (~15–30%), which partially substitutes the group of Quercus + other deciduous trees (~18–30%), documenting the establishment of a holm oak-mixed oak forest, typical of a Mediterranean/sub-Mediterranean climate. Unfortunately, the presence of a thick shoreface sand body devoid of pollen and the lack of a precise chronologic control do not allow to firmly constrain this phase, which ended around 3000 cal year BP. The montane component of the vegetation shows considerable percentages, with two peaks at ~11–16%, mainly accounted for by A. alba, F. sylvatica, and A. cf. viridis and likely to have recorded episodic expansions of cool-temperate forests on the nearby promontories.
4.2.5. VP 5
The youngest vegetation phase is recorded at ~6.5–4.7 m core depth. A slight decrease in arboreal pollen (from ~70% to 50%) characterises the uppermost sample, which also displays a marked peak of Castanea sativa (~16%), a typical tree cultivated since pre-Roman times in Italy [56], but which started to spread in the Middle Ages [57]. The whole VP 5 is characterised by a considerable abundance of cultivated plants, mainly C. sativa and, to a very low extent, Cannabis sativa and Vitis vinifer, which show upward increasing values from ~3% to 17.5%. This trend marks the beginning and progressive increase of the human impact on the gulf and on the surrounding areas, consistent with radiocarbon dating that constrains VP 5 between ~3000 and 1170 cal year BP. Moreover, there is a gradual decrease of Mediterranean taxa (i.e., Q. ilex percentages from 17% to 3%) accompanied by a drop in montane trees from base to top of VP 5 (from 3% to 0.5%). These data could reflect, at least in part, the anthropogenic clearance of woodland.
5. Discussion
The vertical stacking pattern of facies associations recorded in core SP, along with the observed turnovers of both meiofauna and pollen, provide robust information about late Quaternary landscape dynamics at GLS. Radiocarbon data constrain the uppermost 19 m to the Holocene period, while the underlying sedimentary succession is ascribed to the (Late) Pleistocene, based on the age (>45,000 years BP) obtained from swamp deposits at the core bottom (Figure 3, Figure 4 and Figure 5; Table 1). These deposits overlie dozens of m-thick alluvial successions mainly formed by gravel and sand, as revealed by the stratigraphy of cores SC, SD, and SI, located less than 500 m seaward of core SP (Figure 2B). A radiocarbon date from a fine-grained interval of core SI at 42.5 m below sea level (s.l.) yielded an age of >45,000 years BP.
Above this thick alluvial succession, likely reflecting an advancing alluvial fan system into the GLS during an intense glacial period (MIS 6), our multi-proxy data highlight the four-fold, Late Pleistocene-Holocene environmental evolution of the rocky embayment under the predominant forcing of Milankovitch-scale palaeoclimate oscillations (100,000-year cycles).
5.1. Late Pleistocene Barrier-Lagoon System
The vertical succession of organic-rich swamps and fossiliferous lagoon silty clays of the pre-Holocene age (~28.5–24.5 m below s.l.) testifies to progressive flooding of the GLS via the backstepping of a barrier-lagoon system, as a result of rising sea-level conditions. This interpretation is strengthened by the pollen record of an interglacial vegetation phase (VP 1; Figure 5) characterised by mixed oak forests progressively dominated by the Quercus species, suggesting the establishment of PNV concomitant with lagoon formation. The generally low amount of halophytes (<2%) and the overwhelming dominance of euryhaline, opportunistic meiofauna species (C. torosa, L. elliptica and A. tepida + A. parkinsoniana) point to a high-confined lagoonal environment surrounded by freshwater-fed wet grasslands deprived of direct fluvial inputs, as suggested by the absence of freshwater-hypohaline ostracods and by the presence of gypsum crystals. The stratigraphic context supported by regional VP 1 pollen data suggests the attribution of this transitional setting to the Last Interglacial, when RSL was sufficiently high [58,59] to inundate the inland portion of GLS. At comparable stratigraphic depths (~22–20 m below sea level) a brackish lagoon succession ascribed to the Last Interglacial is recorded ~60 km SE, in the shallow subsurface of the Arno River plain supporting this interpretation [15]. The unconformable upper boundary with the overlying fluvial-channel sand body (Figure 3, Figure 4 and Figure 5) reasonably implies that widespread fluvial erosion partly eroded the lagoonal deposits in GLS, preventing the full record of the Last Interglacial succession. This could explain the absence of a shallow-marine environment and the low degree of VP 1 mediterraneity, as both might not have been preserved within the subsurface record of GLS.
5.2. Late Pleistocene Alluvial System
Above the lagoon deposits, the presence of two coarse-grained fluvial intervals, separated by thin freshwater swamp silts, documents the establishment of a fully continental setting within the GLS (~24.5–17.5 m below s. l.). The regional vegetation pattern (i.e., the co-dominance of cool-temperate and meso-thermophilous trees; VP 2 in Figure 5) and the overall stratigraphic framework suggest a last glacial age for this alluvial system. As the uppermost fluvial interval is barren of pollen, no further pollen stratigraphic inference is possible. The palynological data from the fine-grained swamp succession record the development of a forest landscape with abundant A. alba (~12–27%), a conifer with a preference for high moisture availability [60], during a phase of fluvial-channel inactivity and wetland formation (hygrophyte plants + aquatics reach ~10–20%). Interestingly, the sample from the base of the lower fluvial interval, close to the erosive boundary with the underlying lagoon deposits, still belongs to VP 1. This suggests a delayed response of vegetation to the establishment of glacial conditions with respect to the rapid change in sedimentary dynamics that promoted fluvial incision and channel fill. It is difficult to relate past fluvial activity in the gulf to a specific drainage network owing to the paucity of river courses flowing into the GLS, especially along its eastern margin where our data are located (Figure 2). A potential candidate is the inactive alluvial fan of the Pleistocene age [35] outcropping less than 2 km inland of the studied core SP (Termo alluvial fan; Figure 1A). Accordingly, a pre-Holocene phase of alluvial fan progradation is reported from the nearby Magra River plain (Figure 1A) and the Apuo-Versilia plain, located ~25 km SE [61].
5.3. Early Holocene Barrier-Lagoon System
At around 17.5 m below s.l., the studied succession records the Holocene flooding of the GLS through the abrupt superposition of transitional deposits onto the Pleistocene alluvial system. This vertical stacking pattern of facies associations is also documented in the shallow subsurface of the nearby Magra plain (at Luni archaeological site; Figure 1A), where lagoon sediments onlap on top of a piedmont alluvial fan, deepening towards SW, in response to the Holocene transgression [45].
A swamp environment with clear evidence of proximity to the coeval coastline (i.e., transported brackish-shallow marine meiofauna and gypsum crystals) first occurred around 9000 cal year BP. This was rapidly replaced by the establishment of a lagoon basin highly connected to the open sea (Figure 3 and Figure 4). The abrupt appearance of mud-loving marine ostracods (i.e., C. neapolitana) and miliolids, accompanied by a remarkable amount of halophytes (~4–7%), marks a rapid marine ingression and the formation of a barrier-lagoon system into the gulf under the predominant forcing of high RSL rates during the early Holocene [29]. Interestingly, seismic profiles from the gulf area [30] document the occurrence of a sand bar at ~13–12 m water depths (about 3 km away from the present-day coastline) and the bar’s crest shoals to less than 3 m below the sea floor. This raised cross-gulf barrier-bar is consistent with the stratigraphic depths of the lagoon interval (Figure 3, Figure 4 and Figure 5) and with the estimated RSL values for the W Mediterranean (~15 m lower than present s.l.; [29]). The palynological data indicate a simultaneous development of wide wetlands along the lagoon margins (hygrophyte plants + aquatics reach ~30%) and, on a regional scale, the progressive achievement of an optimum-like vegetation (VP 3 in Figure 5) pointing to a landscape deeply shaped by early Holocene transgressive conditions. The peak (~25%) in A. alba within the swampy interval possibly reflects a minor cool and wet oscillation around 8500–8000 cal year BP (8200 event; [62]), which is well-recorded in the Renella and Corchia caves of the Apuane Alps [63,64,65], ~20–40 km SE of the study area.
Coeval barrier-lagoon systems are documented from several coastal-delta plains facing the Ligurian-Tyrrhenian seas (Apuo-Versilia plain; Arno River plain; Posada River plain; Ombrone Delta; Tiber Delta; [9,15,66,67,68]) as well as the Adriatic sea (Po Delta; Biferno Delta; [69,70]), irrespective of their physiographic context and dimension, and the presence (or not) of an important river drainage network. This stratigraphic evidence clearly testifies a predominant climate-eustatic control on the Western Mediterranean coastal landscape during the Early Holocene, which was characterised by very high RSL rising rates [29].
5.4. Mid-Late Holocene Coastal Bay System
At around 8000 cal year BP, the stratigraphic record and the faunal assemblage (disappearance of C. torosa and Loxoconcha elliptica) document a further change in the environmental conditions within the gulf, reasonably caused by the flooding of the barrier that protected the early Holocene lagoon system. This flooding event, chronologically consistent with the climax of RSL rising rates in the W Mediterranean [29], led to the submersion of the barrier and to the establishment of a shallow-marine/open bay environment. The decreasing-increasing trend of hygrophyte plants + aquatics (in particular the hygrophilous trees; Figure 5) mimics the achievement of the maximum water-depth around 7000 cal year BP and the following coastline progradation, which culminates with the establishment of a shoreface environment. This progressive shallowing trend traces the effects of the RSL highstand in the gulf from about 7000 cal year BP until the construction of La Spezia harbour (i.e., anthropogenic material in core SP; Figure 3, Figure 4 and Figure 5). This phenomenon is particularly well-recorded by the meiofauna through the quite abrupt replacement of A. tepida-A. parkinsoniana with epiphytic, hyaline foraminifers (e.g., Planorbulina, Elphidium and Rosalina species) and the concomitant occurrence of a more diverse ostracod fauna containing species that prefer sandy, vegetated bottoms (e.g., Xestoleberis spp.; Aurila spp.; L. gr rhomboidea). This evolution phase corresponds to the development of the present-day GLS: a shallow marine area physically sheltered by rocky promontories, covered with Posidonia meadows and marked by sandy beaches passing seaward to finer-grained bottoms. The regional vegetation landscape is very similar to the one that characterised the early Holocene period (optimum-like vegetation; VP 3), except for a slight increase in Montane taxa (mainly A. alba, Alnus cf. viridis and Betula) (Figure 5). However, the palynological record is incomplete owing to the absence of palynomorphs within shoreface deposits.
Finally, shoreline progradation led to the establishment of a coastal plain featured by a set of emerged sand bars and humid areas (as coastal ponds), the latter formed within topographic depressions where storm deposits had accumulated since historical times (after about 3000 cal year BP—Bronze-Iron Age). The presence of coastal wetlands is testified by remarkable amounts of hygrophyte plants + aquatics (commonly 15–18% in total) up to the early Middle Ages (1290–1175 cal year BP), when the city of La Spezia was founded [71]. However, the pollen records document a vegetation community increasingly impacted by humans since the Bronze-Iron Age through agricultural activities, mainly related to Castanea sativa plantations in the surrounding hills (Figure 5, Table S3).
The replacement of a back-barrier environment by an open bay was likely favoured by the GLS morphology (Section 2.1), which prevented the formation of a recent and more proximal cross-gulf bar. This phenomenon contributed to differentiate the Middle-Late Holocene evolutionary trend of the rocky embayment from several Mediterranean coastal-delta plains, where a progressive closing and emersion of lagoons occurred since ~5000 cal year BP ([7,12,15,68]).
6. Conclusions
A multi-proxy approach including sedimentological analysis integrated with meiofauna, pollen content and radiocarbon dating was applied to the 30 m-thick succession of core SP, in the eastern part of the Gulf of La Spezia (GLS), a relatively deep embayment protected by steep promontories. This study provides new insights into the late Quaternary environmental evolution of a rocky embayment over the last two interglacial-glacial cycles.
Our data and interpretations support the following conclusions:
- The interplay between Milankovitch-scale eustatic-climate oscillations and inherited basin morphologies shaped the GLS landscape through time, leading to the alternation of transitional-shallow marine and alluvial environments during interglacial and glacial periods, respectively;
- The GLS rocky embayment proved to be a very dynamic setting able to host a variety of transitional-shallow marine environments during interglacial periods. All sedimentary facies developed under highstand or transgressive RSL conditions showed a low degree of fluvial influence, consistent with the present-day hydrographic context. Even under high confined conditions (those of inner lagoons, coastal ponds, etc.) river fluxes played a negligible role in the environmental dynamics, as testified by the absence of a freshwater-hypohaline ostracod fauna and by the formation of gypsum crystals/rosettes. Only during the last glacial period was the gulf subject to strong sedimentary inputs from the hinterland, and an alluvial fan progradation occurred favoured by the proximity to steep promontories;
- Pollen taxa/groups mainly reflecting the local pollen rain proved to be excellent palaeoenvironmental indicators, which strengthened and complemented the information derived from the meiofaunal and sedimentological records. More specifically, pollen assemblages reflect the degree of hygrophily/humidity in the coastal plain (hygrophilous trees and hygro + aquatic herbs), offering a complete picture of gulf palaeolandscapes. Moreover, the group of halophytes made it possible to update and refine the environmental information derived from the meiofauna in terms of salinity and degree of confinement;
- The relative abundances of mesophilous taxa (Quercetum taxa + other deciduous trees), Mediterranean taxa (mainly Q. ilex) and Montane taxa (mainly A. alba) helped to track the main changes in regional vegetation cover during the late Quaternary, and to provide vegetation-derived palaeoclimate inferences at a Milankovitch timescale. Apart from present interglacial (Holocene) vegetation phases (VPs 5–3), two older VPs were recognised and assigned to the last glacial (VP 2) and interglacial (VP 1) periods. The high tree cover (>50–60%) that characterised all the VPs and the particular composition of VP 2 (co-dominance of cool-temperate and meso-thermophilous trees) suggest that GLS acted as an important tree refuge, especially during glacial times, when favourable physiographic features (high promontories surrounding the gulf and proximity to the sea) are likely to have fostered mild temperatures and high moisture conditions;
- The Holocene RSL rise caused GLS inundation and the establishment at around 9000–8500 cal year BP of a low-confined lagoon environment, protected seaward by an inherited relief (sandy barrier) located at ~13–12 m water depths on the gulf bottom. Around 8000 cal year BP, the barrier was submerged under accelerating RSL rise, which led to the development of the modern coastal bay, only partially sheltered by the surrounding promontories. This phase, which also includes shoreline progradation under RSL highstand (last 7000 years), occurred during optimum-like vegetation conditions that showed an increase in mediterraneity during the last few millennia (VPs 4–5);
- The Middle-Late Holocene depositional and palaeoenvironmental dynamics, occurred within the GLS, reflect evolutionary patterns that move away from those already documented for several Western Mediterranean plains. The superposition of shoreface and coastal wetlands deposits onto open bay sediments tracks the progradation of the coastline into a rocky embayment subject to predominant marine processes;
- Until the foundation of the old town of La Spezia in the early Middle Ages, coastal wetlands represented natural topographic lows in the coastal plain, where the hygrophyte plants flourished. The earliest evidence of human disturbance in the vegetation landscape dates back to pre-Roman times, about 3000 cal year BP, as highlighted by the stratigraphic records of cultivated plants. A dramatic increase of Castanea sativa is recorded along the hill slopes during the early medieval period.
Supplementary Materials
The following are available online at https://www.mdpi.com/2073-4441/13/4/427/s1, Table S1: Benthic foraminifers and ostracod counts; values of the diversity indices presented in the text are also reported. Samples barren of meiofauna are not shown, Table S2: List of reference works used for the identification and autoecological interpretation of benthic foraminiferal, ostracod and pollen taxa, Table S3: Relative abundances of pollen taxa and groups. Pollen concentration data are also reported.
Author Contributions
Conceptualization, V.R., M.B., M.M., S.M., and A.A.; sampling, sample treatment, methodology, laboratory analyses, M.M., S.M., A.C., and V.R.; pollen data interpretation, M.M. and S.M.; All the authors contributed to the discussion (V.R., A.A., M.M., S.M., A.C., L.L., S.L.T., G.V., M.B.); writing V.R., A.A., M.B., M.M., and S.M.; figure preparation, L.L., M.B., and V.R.; All authors have read and agreed to the submitted version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available at supplementary material.
Acknowledgments
We are very thankful to the reviewers and to the Editor, who made a careful review of the manuscript and greatly improved it through their comments and remarks. We are indebted to Marco Cacciari and Marco Luppichini for the technical support during the sampling procedure and reference formatting activity. We are very grateful to L. Cignoni for the English language editing and revision.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Carbognin, L.; Teatini, P.; Tomasin, A.; Tosi, L. Global Change and Relative Sea Level Rise at Venice: What Impact in Term of Flooding. Clim. Dyn. 2010, 35, 1055–1063. [Google Scholar] [CrossRef]
- Marsico, A.; Lisco, S.; Lo Presti, V.; Antonioli, F.; Amorosi, A.; Anzidei, M.; Deiana, G.; De Falco, G.; Fontana, A.; Fontolan, G.; et al. Flooding Scenario for Four Italian Coastal Plains Using Three Relative Sea Level Rise Models. J. Maps 2017, 13, 961–967. [Google Scholar] [CrossRef]
- Antonioli, F.; De Falco, G.; Lo Presti, V.; Moretti, L.; Scardino, G.; Anzidei, M.; Bonaldo, D.; Carniel, S.; Leoni, G.; Furlani, S.; et al. Relative Sea-Level Rise and Potential Submersion Risk for 2100 on 16 Coastal Plains of the Mediterranean Sea. Water 2020, 12, 2173. [Google Scholar] [CrossRef]
- Di Rita, F.; Simone, O.; Caldara, M.; Gehrels, W.R.; Magri, D. Holocene Environmental Changes in the Coastal Tavoliere Plain (Apulia, Southern Italy): A Multiproxy Approach. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 310, 139–151. [Google Scholar] [CrossRef]
- Di Rita, F.; Molisso, F.; Sacchi, M. Late Holocene Environmental Dynamics, Vegetation History, Human Impact, and Climate Change in the Ancient Literna Palus (Lago Patria; Campania, Italy). Rev. Palaeobot. Palynol. 2018, 258, 48–61. [Google Scholar] [CrossRef]
- Longhitano, S.G.; Della Luna, R.; Milone, A.L.; Cilumbriello, A.; Spilotro, G. The 20,000-years-long sedimentary record of the Lesina coastal system (southern Italy): From alluvial, to tidal, to wave process regime change. Holocene 2015, 5, 18–21. [Google Scholar] [CrossRef]
- Bellotti, P.; Calderoni, G.; Dall’Aglio, P.L.; D’Amico, C.; Davoli, L.; Di Bella, L.; D’Orefice, M.; Esu, D.; Ferrari, K.; Bandini Mazzanti, M.; et al. Middle-to Late-Holocene Environmental Changes in the Garigliano Delta Plain (Central Italy): Which Landscape Witnessed the Development of the Minturnae Roman Colony? Holocene 2016, 26, 1457–1471. [Google Scholar] [CrossRef]
- Ejarque, A.; Julià, R.; Reed, J.M.; Mesquita-Joanes, F.; Barba, J.M.; Riera, S. Coastal Evolution in a Mediterranean Microtidal Zone: Mid to Late Holocene Natural Dynamics and Human Management of the Castelló Lagoon, NE Spain. PLoS ONE 2016, 11, e0155446. [Google Scholar] [CrossRef] [PubMed]
- Melis, R.T.; Di Rita, F.; French, C.; Marriner, N.; Montis, F.; Serreli, G.; Sulas, F.; Vacchi, M. 8000 years of Coastal Changes on a Western Mediterranean Island: A Multiproxy Approach from the Posada Plain of Sardinia. Mar. Geol. 2018, 403, 93–108. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, A.; Blázquez, A.M.; Guillem, J.; Usera, J. Maximum Flood Area during MIS 1 in the Almenara Marshland (Western Mediterranean): Benthic Foraminifera and Sedimentary Record. Holocene 2018, 28, 1452–1466. [Google Scholar] [CrossRef]
- Revelles, J.; Ghilardi, M.; Rossi, V.; Currás, A.; López-Bultó, O.; Brkojewitsch, G.; Vacchi, M. Coastal Landscape Evolution of Corsica Island (W. Mediterranean): Palaeoenvironments, Vegetation History and Human Impacts since the Early Neolithic Period. Quat. Sci. Rev. 2019, 225. [Google Scholar] [CrossRef]
- D’Orefice, M.; Bellotti, P.; Bertini, A.; Calderoni, G.; Neri, P.C.; Di Bella, L.; Fiorenza, D.; Foresi, L.M.; Louvari, M.A.; Rainone, L.; et al. Holocene Evolution of the Burano Paleo-Lagoon (Southern Tuscany, Italy). Water 2020, 12, 1007. [Google Scholar] [CrossRef]
- Amorosi, A.; Colalongo, M.L.; Fusco, F.; Pasini, G.; Fiorini, F. Glacio-Eustatic Control of Continental-Shallow Marine Cyclicity from Late Quaternary Deposits of the Southeastern Po Plain, Northern Italy. Quat. Res. 1999, 52, 1–13. [Google Scholar] [CrossRef]
- Aguzzi, M.; Amorosi, A.; Colalongo, M.L.; Lucchi, M.R.; Rossi, V.; Sarti, G.; Vaiani, S.C. Late Quaternary Climatic Evolution of the Arno Coastal Plain (Western Tuscany, Italy) from Subsurface Data. Sediment. Geol. 2007, 202, 211–229. [Google Scholar] [CrossRef]
- Cacciari, M.; Amorosi, A.; Campo, B.; Marchesini, M.; Rossi, V. Palynology of the Late Quaternary Succession of the Arno Plain (Northern Italy): New Insights on Palaeoenvironmental Trends and Climate Dynamics. Boll. Soc. Paleontol. Ital. 2019, 58, 201–221. [Google Scholar] [CrossRef]
- Campo, B.; Bruno, L.; Amorosi, A. Basin-Scale Stratigraphic Correlation of Late Pleistocene-Holocene (MIS 5e–MIS 1) Strata across the Rapidly Subsiding Po Basin (Northern Italy). Quat. Sci. Rev. 2020, 237. [Google Scholar] [CrossRef]
- Dolez, L.; Salel, T.; Bruneton, H.; Colpo, G.; Devillers, B.; Lefèvre, D.; Muller, S.D.; Sanchez, C. Holocene Palaeoenvironments of the Bages-Sigean Lagoon (France). Geobios 2015, 48, 297–308. [Google Scholar] [CrossRef]
- Milli, S.; Girasoli, D.E.; Tentori, D.; Tortora, P. Sedimentology and Coastal Dynamics of Carbonate Pocket Beaches: The Ionian-Sea Apulia Coast between Torre Colimena and Porto Cesareo (Southern Italy). J. Mediterr. Earth Sci. 2017, 9, 29–66. [Google Scholar] [CrossRef]
- Aiello, G.; Amato, V.; Barra, D.; Caporaso, L.; Caruso, T.; Giaccio, B.; Parisi, R.; Rossi, A. Late Quaternary Benthic Foraminiferal and Ostracod Response to Palaeoenvironmental Changes in a Mediterranean Coastal Area, Port of Salerno, Tyrrhenian Sea. Reg. Stud. Mar. Sci. 2020, 40, 101498. [Google Scholar] [CrossRef]
- Furlani, S.; Pappalardo, M.; Gómez-Pujol, L.; Chelli, A. The rock coast of the Mediterranean and Black seas. In Rock Coast Geomophology; Kennedy, D.M., Stephenson, W.J., Naylor, L.A., Eds.; Geological Society, Memoirs: London, UK, 2014; pp. 89–123. [Google Scholar]
- Chelli, A.; Pappalardo, M.; Llopis, I.A.; Federici, P.R. The Relative Influence of Lithology and Weathering in Shaping Shore Platforms along the Coastline of the Gulf of La Spezia (NW Italy) as Revealed by Rock Strength. Geomorphology 2010, 118, 93–104. [Google Scholar] [CrossRef]
- Rovere, A.; Vacchi, M.; Firpo, M.; Carobene, L. Underwater Geomorphology of the Rocky Coastal Tracts between Finale Ligure and Vado Ligure (Western Liguria, NW Mediterranean Sea). Quat. Int. 2011, 232, 187–200. [Google Scholar] [CrossRef]
- Antonioli, F.; Lo Presti, V.; Rovere, A.; Ferranti, L.; Anzidei, M.; Furlani, S.; Mastronuzzi, G.; Orru, P.E.; Scicchitano, G.; Sannino, G.; et al. Tidal Notches in Mediterranean Sea: A Comprehensive Analysis. Quat. Sci. Rev. 2015, 119, 66–84. [Google Scholar] [CrossRef]
- Furlani, S. The Rocky Coasts of the Mediterranean Sea: Field Studies. In Proceedings of the Conférence Méditerranéenne Côtière et Maritime, Split, Croatia, 29 November–1 December 2017; pp. 1–6. [Google Scholar]
- Arobba, D.; Caramiello, R.; Firpo, M.; Mercalli, L.; Morandi, L.F.; Rossi, S. New Evidence on the Earliest Human Presence in the Urban Area of Genoa (Liguria, Italy): A Multi-Proxy Study of a Mid-Holocene Deposit at the Mouth of the Bisagno River. Holocene 2018, 28, 1918–1935. [Google Scholar] [CrossRef]
- Geoportale. Available online: http://srvcarto.regione.liguria.it/geoviewer2/pages/apps/geoportale/index.html (accessed on 27 December 2020).
- Gasparini, G.P.; Abbate, M.; Bordone, A.; Cerrati, G.; Galli, C.; Lazzoni, E.; Negri, A. Circulation and Biomass Distribution during Warm Season in the Gulf of La Spezia (North-Western Mediterranean). J. Mar. Syst. 2009, 78, 548–562. [Google Scholar] [CrossRef]
- Bellini, C.; Mariotti-Lippi, M.; Montanari, C. The Holocene Landscape History of the NW Italian Coasts. Holocene 2009, 19, 1161–1172. [Google Scholar] [CrossRef]
- Vacchi, M.; Marriner, N.; Morhange, C.; Spada, G.; Fontana, A.; Rovere, A. Multiproxy Assessment of Holocene Relative Sea-Level Changes in the Western Mediterranean: Sea-Level Variability and Improvements in the Definition of the Isostatic Signal. Earth Sci. Rev. 2016, 155, 172–197. [Google Scholar] [CrossRef]
- Breslau, L.R.; Edgerton, H.E. The Gulf of La Spezia, Italy: A case history of seismic-sedimentologic correlation. In The Mediterranean Sea: A Natural Sedimentation Laboratory; Stanlet, D.J., Ed.; Dowden, Hutchinson and Ross, Inc.: Stroudsburg, PA, USA, 1972; pp. 177–188. [Google Scholar]
- Mariani, M.; Prato, R. I Bacini Neogenici Costieri Del Margine Tirrenico: Approccio Sismico-Stratigrafico. Mem. Soc. Geol. Ital. 1988, 41, 519–531. [Google Scholar]
- Boccaletti, M.; Corti, G.; Martelli, L. Recent and Active Tectonics of the External Zone of the Northern Apennines (Italy). Int. J. Earth Sci. 2011, 100, 1331–1348. [Google Scholar] [CrossRef]
- Conti, P.; Cornamusini, G.; Carmignani, L. An Outline of the Geology of the Northern Apennines (Italy), with Geological Map at 1:250,000 Scale. Ital. J. Geosci. 2020, 139, 149–194. [Google Scholar] [CrossRef]
- Gandolfi, G.; Paganelli, L. Il Macigno Costiero fra La Spezia e Grosseto. G. Geol. 1992, 54, 163–179. [Google Scholar]
- Abbate, E.; Fanucci, F.; Benvenuti, M.; Bruni, P.; Cipriani, N.; Falorni, P.; Fazzuoli, M.; Morelli, D.; Pandeli, E.; Papini, M.; et al. Note Illustrative della Carta Geologica d’Italia alla Scala 1:50.000. Foglio 248 La Spezia; S.EL.CA. s.r.l.: Florence, Italy, 2005; 288p. [Google Scholar]
- Nisio, S.I. Sinkholes in Liguria. Mem. Descr. Cart. Geol. Ital. 2008, 85, 399–408. [Google Scholar]
- Climate Data. Dati Climatici Sulle Città del Mondo. Available online: https://it.climate-data.org/ (accessed on 27 December 2020).
- Barberis, G.; Peccenini, S.; Vagge, I. Formazioni Arbustive Mediterranee in Liguria. Fitosociologia 1997, 32, 61–71. [Google Scholar]
- Vagge, I. La Diffusione Del Bioclima Mediterraneo in Liguria (Italia Nord Occidentale). Fitosociologia 2014, 36, 95–109. [Google Scholar]
- Debenay, J.P.; Guillou, J.J. Ecological Transitions Indicated by Foraminiferal Assemblages in Paralic Environments. Estuaries 2002, 25, 1107–1120. [Google Scholar] [CrossRef]
- Frenzel, P.; Boomer, I. The Use of Ostracods from Marginal Marine, Brackish Waters as Bioindicators of Modern and Quaternary Environmental Change. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 225, 68–92. [Google Scholar] [CrossRef]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Horne, D.J.; Curry, B.B.; Mesquita-Joanes, F. Mutual climatic range methods for Quaternary ostracods. In Ostracoda as Proxies for Quaternary Climate Change. Developments in Quaternary Science; Horne, D.J., Holmes, J.A., Rodriguez-Lazaro, J., Viehberg, F.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 65–84. [Google Scholar]
- Milli, S.; D’Ambrogi, C.; Bellotti, P.; Calderoni, G.; Carboni, M.G.; Celant, A.; Di Bella, L.; Di Rita, F.; Frezza, V.; Magri, D.; et al. The Transition from Wave-Dominated Estuary to Wave-Dominated Delta: The Late Quaternary Stratigraphic Architecture of Tiber River Deltaic Succession (Italy). Sediment. Geol. 2013, 284–285, 159–180. [Google Scholar] [CrossRef]
- Bini, M.; Brückner, H.; Chelli, A.; Pappalardo, M.; Da Prato, S.; Gervasini, L. Palaeogeographies of the Magra Valley Coastal Plain to Constrain the Location of the Roman Harbour of Luna (NW Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 337–338, 37–51. [Google Scholar] [CrossRef]
- Barbieri, G.; Vaiani, S.C. Benthic Foraminifera or Ostracoda? Comparing the Accuracy of Palaeoenvironmental Indicators from a Pleistocene Lagoon of the Romagna Coastal Plain (Italy). J. Micropalaeontol. 2018, 37, 203–230. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T. Paleontological Data Analysis; Wiley-Blackwell: New York, NY, USA, 2005. [Google Scholar]
- Fisher, R.A.; Corbet, A.S.; Williams, C.B. The Relation between the Number of Species and the Number of Individuals in a Random Sample of an Animal Population. J. Anim. Ecol. 1943, 12, 42–58. [Google Scholar] [CrossRef]
- Shannon, C.A. Mathematical Theory of Communication. Bell Syst. Technol. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Lowe, J.J.; Accorsi, C.A.; Bandini Mazzanti, M.; Bishop, V.; der Kaars, S.; Forlani, L.; Mercuri, A.M.; Rivalenti, C.; Torri, P.; Watson, C. Pollen Stratigraphy of Sediment Sequences from Carter Lakes Albano and Nemi (near Rome) and from the Central Adriatic, Spanning the Interval from Oxygen Isotope Stage 2 to the Present Day. Mem. Ist. Ital. Idrobiol. 1996, 55, 71–98. [Google Scholar]
- Pignatti, S. I Piani Di Vegetazione in Italia. G. Bot. Ital. 1979, 113, 411–428. [Google Scholar] [CrossRef]
- Stuiver, M.; Reimer, P.J.; Reimer, R.W. CALIB 8.2. Available online: http://calib.org (accessed on 25 December 2020).
- Reimer, P.J.; Austin, W.E.N.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Bronk Ramsey, C.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0–55 Cal KBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Amorosi, A.; Rossi, V.; Scarponi, D.; Vaiani, S.C.; Ghosh, A. Biosedimentary Record of Postglacial Coastal Dynamics: High-Resolution Sequence Stratigraphy from the Northern Tuscan Coast (Italy). Boreas 2014, 43, 939–954. [Google Scholar] [CrossRef]
- Cacciari, M.; Amorosi, A.; Marchesini, M.; Kaniewski, D.; Bruno, L.; Campo, B.; Rossi, V. Linking Holocene Vegetation Dynamics, Palaeoclimate Variability and Depositional Patterns in Coastal Successions: Insights from the Po Delta Plain of Northern Italy. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2020, 538, 109468. [Google Scholar] [CrossRef]
- Mercuri, A.M.; Mazzanti, M.B.; Torri, P.; Vigliotti, L.; Bosi, G.; Florenzano, A.; Olmi, L.; N’siala, I.M. A Marine/Terrestrial Integration for Mid-Late Holocene Vegetation History and the Development of the Cultural Landscape in the Po Valley as a Result of Human Impact and Climate Change. Veg. Hist. Archaeobot. 2012, 21, 353–372. [Google Scholar] [CrossRef]
- Accorsi, C.A.; Bandini Mazzanti, M.; Forlani, L.; Mercuri, A.M.; Trevisan Grandi, G. An Overview of Holocene Forest Pollen Flora/ Vegetation of the Emilia Romagna Region—Northern Italy. Arch. Geobot. 1999, 5, 3–37. [Google Scholar]
- Federici, P.R.; Pappalardo, M. Evidence of Marine Isotope Stage 5.5 Highstand in Liguria (Italy) and Its Tectonic Significance. Quat. Int. 2006, 145–146, 68–77. [Google Scholar] [CrossRef]
- Ferranti, L.; Antonioli, F.; Mauz, B.; Amorosi, A.; Dai Pra, G.; Mastronuzzi, G.; Monaco, C.; Orrù, P.; Pappalardo, M.; Radtke, U.; et al. Markers of the Last Interglacial Sea-Level High Stand along the Coast of Italy: Tectonic Implications. Quat. Int. 2006, 145–146, 30–54. [Google Scholar] [CrossRef]
- Tinner, W.; Colombaroli, D.; Heiri, O.; Henne, P.D.; Steinacher, M.; Untenecker, J.; Vescovi, E.; Allen, J.R.M.; Carraro, G.; Conedera, M.; et al. The Past Ecology of Abies Alba Provides New Perspectives on Future Responses of Silver Fir Forests to Global Warming. Ecol. Monogr. 2013, 83, 419–439. [Google Scholar] [CrossRef]
- Federici, P.R. La tettonica recente dell’Appennino: I—Il bacino villafranchiano di Sarzana e il suo significato nel quadro dei movimenti distensivi a nord-ovest delle Alpi Apuane. Boll. Soc. Geol. Ital. Ital. J. Geosci. 1973, 92, 287–301. [Google Scholar]
- Bond, G.; Showers, W.; Cheseby, M.; Lotti, R.; Almasi, P.; DeMenocal, P.; Priore, P.; Cullen, H.; Hajdas, I.; Bonani, G. A Pervasive Millennial-Scale Cycle in North Atlantic Holocene and Glacial Climates. Science 1997, 278, 1257–1266. [Google Scholar] [CrossRef]
- Zanchetta, G.; Isola, I.; Piccini, L.; Dini, A. The Corchia Cave (Alpi Apuane): A 2 Ma Long Temporal Window on the Earth Climate. Geol. Field Trips 2011, 3, 1–55. [Google Scholar] [CrossRef]
- Zhornyak, L.V.; Zanchetta, G.; Drysdale, R.N.; Hellstrom, J.C.; Isola, I.; Regattieri, E.; Piccini, L.; Baneschi, I.; Couchoud, I. Stratigraphic Evidence for a “Pluvial Phase” between ca 8200–7100 Ka from Renella Cave (Central Italy). Quat. Sci. Rev. 2011, 30, 409–417. [Google Scholar] [CrossRef]
- Regattieri, E.; Zanchetta, G.; Drysdale, R.N.; Isola, I.; Hellstrom, L.D. Lateglacial to Holocene Trace Element Record (Ba, Mg, Sr) from Corchia Cave (Apuan Alps, Central Italy): Paleoenvironmental Implications. J. Quat. Sci. 2014, 29, 381–392. [Google Scholar] [CrossRef]
- Carboni, M.G.; Bergamin, L.; Di Bella, L.; Esu, D.; Cerone, E.P.; Antonioli, F.; Verrubbi, V. Palaeoenvironmental Reconstruction of Late Quaternary Foraminifera and Molluscs from the ENEA Borehole (Versilian Plain, Tuscany, Italy). Quat. Res. 2010, 74, 265–276. [Google Scholar] [CrossRef]
- Bellotti, P.; Caputo, C.; Davoli, L.; Evangelista, S.; Garzanti, E.; Pugliese, F.; Valeri, P. Morpho-Sedimentary Characteristics and Holocene Evolution of the Emergent Part of the Ombrone River Delta (Southern Tuscany). Geomorphology 2004, 61, 71–90. [Google Scholar] [CrossRef]
- Di Rita, F.; Celant, A.; Milli, S.; Magri, D. Lateglacial-Early Holocene Vegetation History of the Tiber Delta (Rome, Italy) under the Influence of Climate Change and Sea Level Rise. Rev. Palaeobot. Palynol. 2015, 218, 204–216. [Google Scholar] [CrossRef]
- Amorosi, A.; Bruno, L.; Campo, B.; Morelli, A.; Rossi, V.; Scarponi, D.; Hong, W.; Bohacs, K.M.; Drexler, T.M. Global Sea-Level Control on Local Parasequence Architecture from the Holocene Record of the Po Plain, Italy. Mar. Pet. Geol. 2017, 87, 99–111. [Google Scholar] [CrossRef]
- Amorosi, A.; Bracone, V.; Campo, B.; D’Amico, C.; Rossi, V.; Rosskopf, C.M. A Late Quaternary Multiple Paleovalley System from the Adriatic Coastal Plain (Biferno River, Southern Italy). Geomorphology 2016, 254, 146–159. [Google Scholar] [CrossRef]
- Treccani. La Spezia. Available online: https://www.treccani.it/enciclopedia/la-spezia (accessed on 28 December 2020).





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).