
Diversity and Structure of Pelagic Zooplankton (Crustacea, Rotifera) in NE Poland



Abstract
:1. Introduction
2. Study Area and Methods
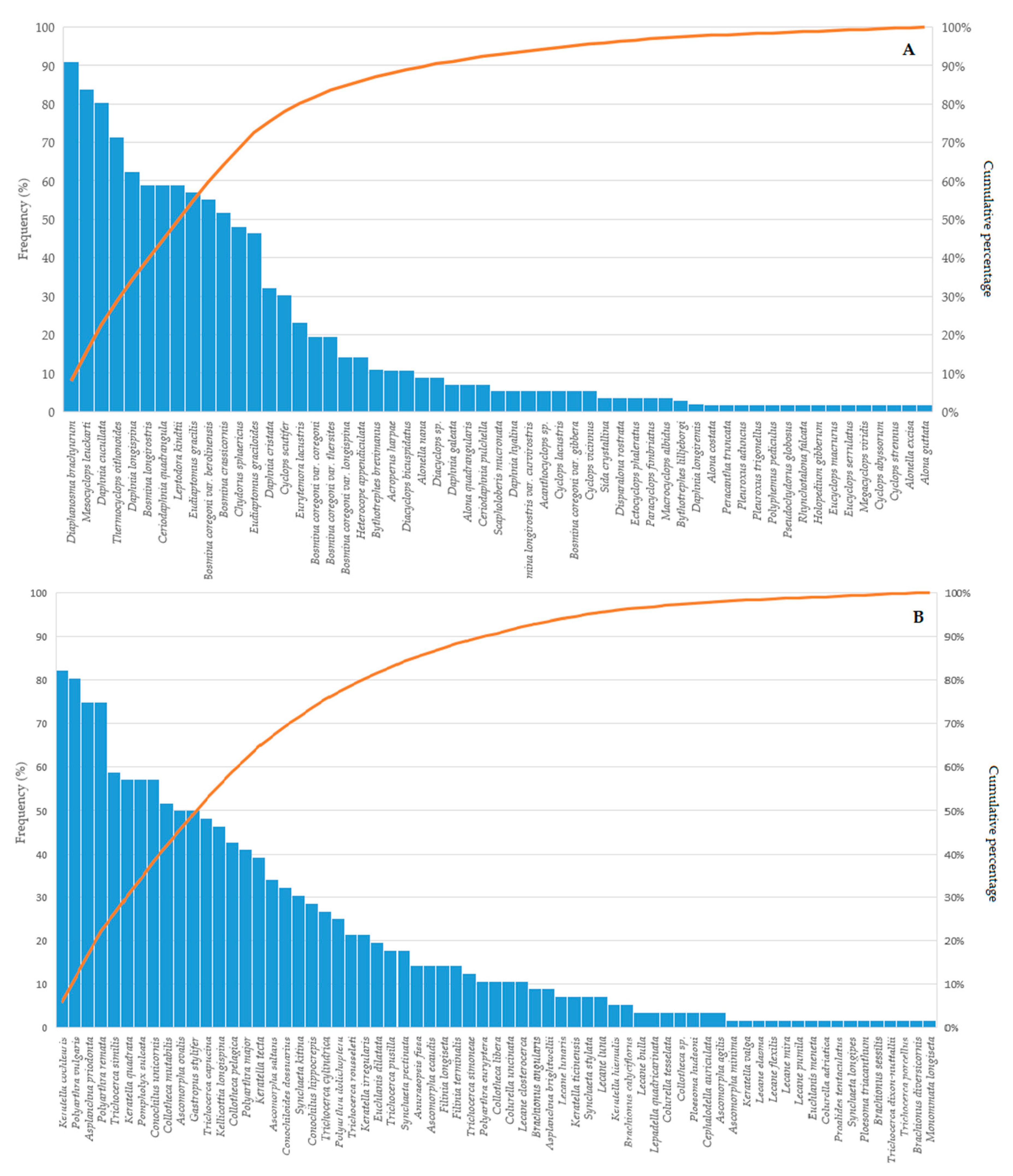
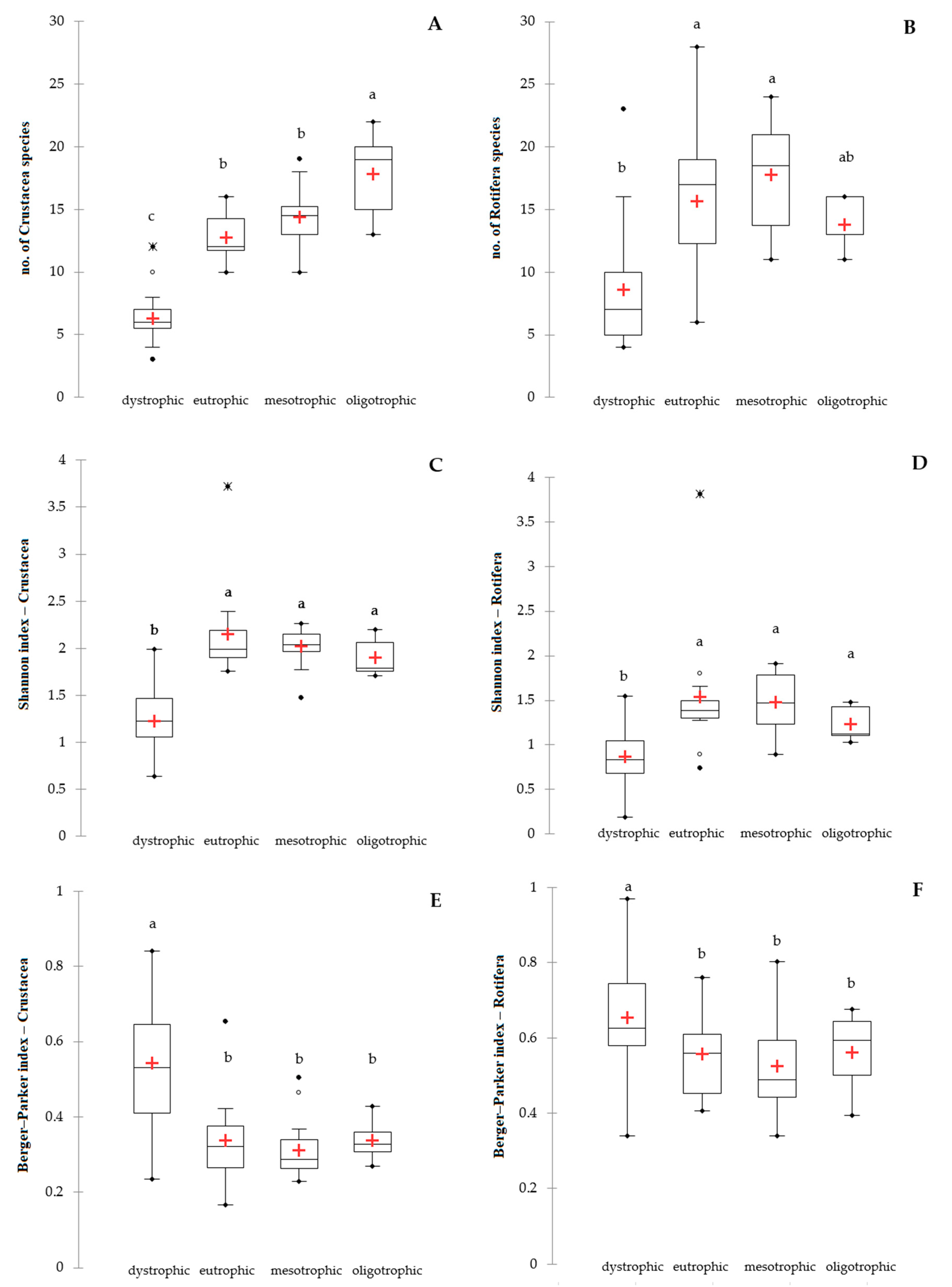
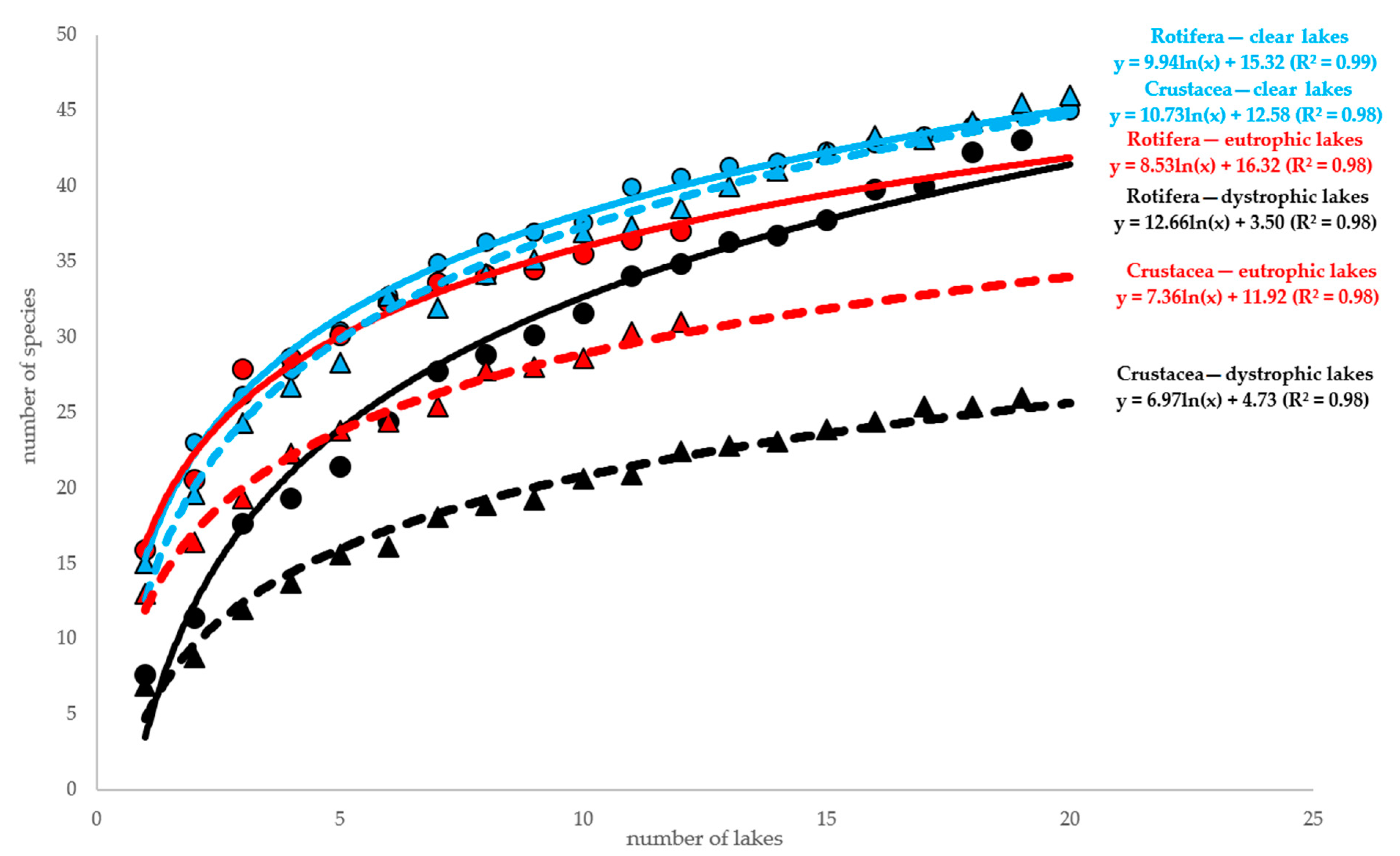
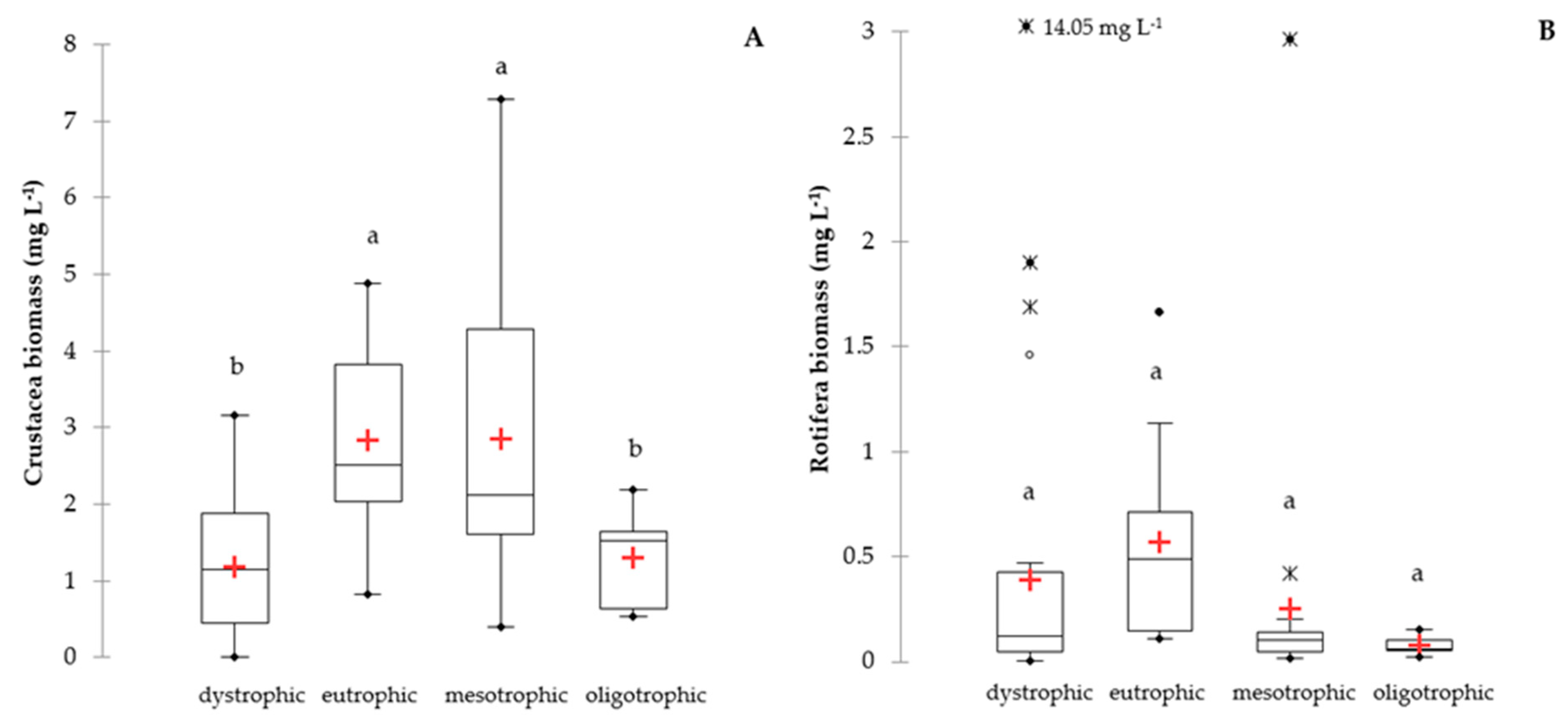
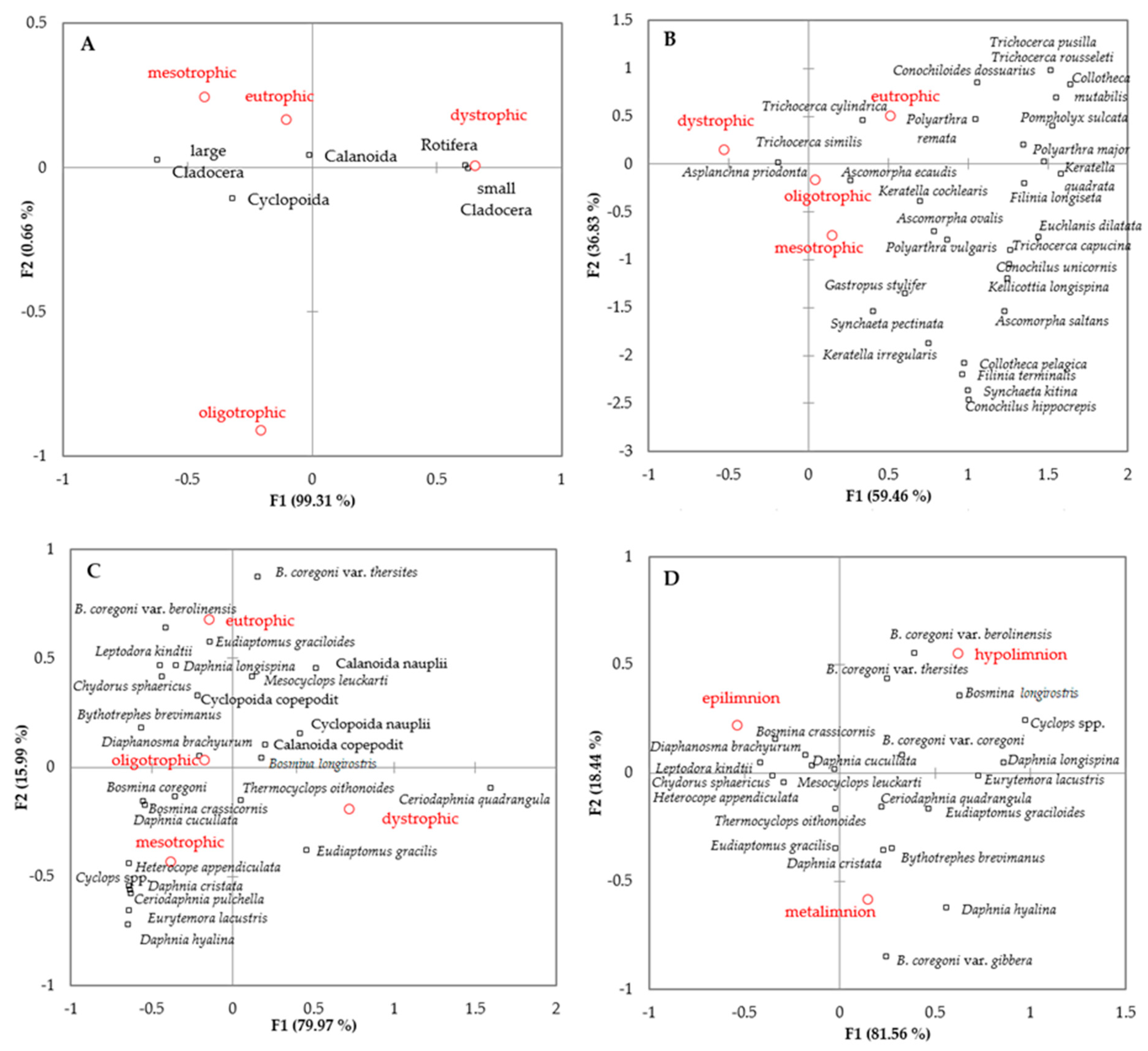
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karpowicz, M.; Zieliński, P.; Grabowska, M.; Ejsmont-Karabin, J.; Kozłowska, J.; Feniova, I. Effect of eutrophication and humification on nutrient cycles and transfer efficiency of matter in the freshwater food web. Hydrobiologia 2020, 847, 2521–2540. [Google Scholar] [CrossRef]
- Gliwicz, Z.M.; Pijanowska, J. The role of predation in zooplankton succession. In Plankton Ecology: Succession in Plankton Communities; Sommer, U., Ed.; Springer: Berlin/Heidelberg, Germany, 1989; pp. 253–296. [Google Scholar]
- Gliwicz, Z.M. Between Hazards of Starvation and Risk of Predation: The Ecology of Off-Shore Animals; International Ecology Institute: Oldendorf, Germany, 2003; ISBN 978-3-946729-12-9. [Google Scholar]
- Polishchuk, L.V.; Vijverberg, J.; Voronov, D.A.; Mooij, W.M. How to measure top–down vs bottom–up effects: A new population metric and its calibration on Daphnia. Oikos 2013, 122, 1177–1186. [Google Scholar] [CrossRef]
- Feniova, I.; Dawidowicz, P.; Ejsmont-Karabin, J.; Gladyshev, M.; Kalinowska, K.; Karpowicz, M.; Kostrzewska-Szlakowska, I.; Majsak, N.; Petrosyan, V.; Razlutskij, V.; et al. Effects of zebra mussels on cladoceran communities under eutrophic conditions. Hydrobiologia 2018, 822, 37–54. [Google Scholar] [CrossRef] [Green Version]
- Gliwicz, Z.M. Filtering rates, food size selection, and feeding rates in cladocerans-another aspect of interspecific competition in filter feeding zooplankton. In Evolution and Ecology of Zooplankton Communities; Kerfoot, W.C., Ed.; New England Press: Hanover, Germany, 1980; pp. 282–291. [Google Scholar]
- Sinistro, R. Top-down and bottom-up regulation of planktonic communities in a warm temperate wetland. J. Plankton Res. 2010, 2, 209–220. [Google Scholar] [CrossRef]
- Walseng, B.; Hessen, D.O.; Halvorsen, G.; Schartau, A.K. Major contribution from littoral crustaceans to zooplankton species richness in lakes. Limnol. Oceanogr. 2006, 51, 2600–2606. [Google Scholar] [CrossRef]
- Karpowicz, M.; Ejsmont-Karabin, J.; Więcko, A.; Górniak, A.; Cudowski, A. A place in space-the horizontal vs vertical factors that influence zooplankton (Rotifera, Crustacea) communities in a mesotrophic lake. J. Limnol. 2019, 78, 243–258. [Google Scholar] [CrossRef]
- Williamson, C.E.; Fischer, J.M.; Bollens, S.M.; Overholt, E.P.; Brenckenridge, J.K. Toward a more comprehensive theory of zooplankton diel vertical migration: Integrating ultraviolet radiation and water transparency into the biotic paradigm. Limnol. Oceanogr. 2011, 56, 1603–1623. [Google Scholar] [CrossRef] [Green Version]
- Stich, H.B.; Lampert, W. Predator evasion as an explanation of diurnal vertical migration by zooplankton. Nature 1981, 293, 396–398. [Google Scholar] [CrossRef]
- Lampert, W. Ultimate causes of diel vertical migration of zooplankton: New evidence for the predator avoidance hypothesis. Arch. Hydrobiol. Beih. Ergebn. Limnol. 1993, 39, 70–88. [Google Scholar]
- Loose, C.J.; Dawidowicz, P. Trade-offs in diel vertical migration by zooplankton: The costs of predator avoidance. Ecology 1994, 75, 2255–2263. [Google Scholar] [CrossRef]
- Karpowicz, M.; Ejsmont-Karabin, J.; Kozłowska, J.; Feniova, I.; Dzialowski, A.R. Zooplankton Community Responses to Oxygen Stress. Water 2020, 12, 706. [Google Scholar] [CrossRef] [Green Version]
- Camacho, A. On the occurrence and ecological features of deep chlorophyll maxima (DCM) in Spanish stratified lakes. Limnetica 2006, 25, 453–478. [Google Scholar] [CrossRef]
- Vincent, W.F.; Goldman, C.R. Evidence for algal heterotrophy in Lake Tahoe, California-Nevada. Limnol. Oceanogr. 1980, 25, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Ptacnik, R.; Diehl, S.; Berger, S. Performance of sinking and nonsinking phytoplankton taxa in a gradient of mixing depths. Limnol. Oceanogr. 2003, 48, 1903–1912. [Google Scholar] [CrossRef] [Green Version]
- Arvola, L.; Ojala, A.; Barbosa, F.; Heaney, S.I. Migration behaviour of three cryptophytes in relation to environmental gradients: An experimental approach. Brit. Phycol. J. 1991, 26, 361–373. [Google Scholar] [CrossRef]
- Reynolds, C.S. Dynamics, selection and composition of phytoplankton in relation to vertical structure in lakes. Arch. Hydrobiol. Beih. Ergebn. Limnol. 1992, 35, 13–31. [Google Scholar]
- Winder, M.; Buergi, H.R.; Spaak, P. Seasonal vertical distribution of phytoplankton and copepod species in a high-mountain lake. Arch. Hydrobiol. 2003, 158, 197–213. [Google Scholar] [CrossRef]
- Gliwicz, Z.M. Metalimnetic gradients and phosphorus loss from the epilimnion in stratified eutrophic lakes. Develop. Hydrobiol. 1980, 2, 23–27. [Google Scholar] [CrossRef]
- Fee, E.J.; Hecky, R.E.; Kasian, S.E.M.; Cruikshank, D.R. Effects of lake size, water clarity, and climatic variability on mixing depths in Canadian Shield lakes. Limnol. Oceanogr. 1996, 41, 912–920. [Google Scholar] [CrossRef]
- Karpowicz, M.; Ejsmont-Karabin, J. Effect of metalimnetic gradient on phytoplankton and zooplankton (Rotifera, Crustacea) communities in different trophic conditions. Environ. Monit. Assess. 2017, 189, 367. [Google Scholar] [CrossRef] [Green Version]
- Christensen, D.L.; Carpenter, S.R.; Cottingham, K.L. Predicting chlorophyll vertical distribution in response to epilimnetic nutrient enrichment in small stratified lakes. J. Plankton Res. 1995, 17, 1461–1477. [Google Scholar] [CrossRef]
- Lampert, W.; Grey, J. Exploitation of deep-water algal maximum by Daphnia: A stable-isotope tracer study. Hydrobiologia 2003, 500, 95–101. [Google Scholar] [CrossRef]
- Cantin, A.; Beisner, B.E.; Gunn, J.M.; Prairie, Y.T.; Winter, J.G. Effects of thermocline deepening on lake plankton communities. Can. J. Fish. Aquat. Sci. 2011, 68, 260–276. [Google Scholar] [CrossRef]
- Francis, T.B.; Schindler, D.E.; Holtgrieve, G.W.; Larson, E.R.; Scheuerell, M.D.; Semmens, B.X.; Ward, E.J. Habitat structure determines resource use by zooplankton in temperate lakes. Ecol. Lett. 2011, 14, 364–372. [Google Scholar] [CrossRef]
- Klumb, R.A.; Bunch, K.L.; Mills, E.L.; Rudstam, L.G.; Brown, G.; Knauf, C.; Burton, R.; Arrhenius, F. Establishment of a metalimnetic oxygen refuge for zooplankton in a productive Lake Ontario embayment. Ecol. Appl. 2004, 14, 113–131. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Ludsin, S.A.; Ruberg, S.A.; Hook, T.O.; Pothoven, S.A.; Brandt, S.B.; Lang, G.A.; Liebig, J.R. Hypoxia affects spatial distributions and overlap of pelagic fish, zooplankton, and phytoplankton in Lake Erie. J. Exp. Mar. Biol. Ecol. 2009, 381, 92–107. [Google Scholar] [CrossRef]
- Larsson, P.; Lampert, W. Experimental evidence of a low oxygen refuge for large zooplankton. Limnol. Oceanogr. 2011, 56, 1682–1688. [Google Scholar] [CrossRef]
- Hofmann, W. Population dynamics of hypolimnetic rotifers in the Pluss-see (North Germany). Hydrobiologia 1987, 147, 197–201. [Google Scholar] [CrossRef]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef]
- Lomolino, M.V.; Riddle, B.R.; Whittaker, R.J.; Brown, J.H. Biogeography, 4th ed.; Sinauer Associates: Sunderland, MA, USA, 2010. [Google Scholar]
- Pinel-Alloul, B.; André, A.; Legendre, P.; Cardille, A.J.; Patalas, K.; Salki, A. Large-scale geographic patterns of diversity and community structure of pelagic crustacean zooplankton in Canadian lakes. Global Ecol. Biogeogr. 2013, 22, 784–795. [Google Scholar] [CrossRef]
- Dumont, H.J.; Segers, H. Estimating lacustrine zooplankton species richness and complementarity. Hydrobiologia 1996, 341, 125–132. [Google Scholar] [CrossRef]
- Patalas, J.; Patalas, K. The crustacean plankton communities in Polish lakes. Verh. Int. Ver. Limnol. 1966, 16, 204–215. [Google Scholar] [CrossRef]
- Shurin, J.; Havel, J.; Leibold, M.; Pinel-Alloul, B. Local and regional zooplankton species richness: A scale-independent test for saturation. Ecology 2000, 81, 3062–3073. [Google Scholar] [CrossRef]
- Shurin, J.B.; Arnott, S.E.; Hillebrand, H.; Longmuir, A.; Pinel-Alloul, B.; Winder, M.; Yan, N.D. Diversity-stability relationship varies with latitude in zooplankton. Ecol. Lett. 2007, 10, 127–134. [Google Scholar] [CrossRef]
- Sługocki, Ł.; Czerniawski, R. Trophic state (TSISD) and mixing type significantly influence pelagic zooplankton biodiversity in temperate lakes (NW Poland). PeerJ 2018, 6, e5731. [Google Scholar] [CrossRef] [Green Version]
- Ejsmont-Karabin, J.; Kalinowska, K.; Karpowicz, M. Structure of ciliate, rotifer, and crustacean communities in lake systems of Northeastern Poland. In Polish River Basins and Lakes—Part II; Springer: Cham, Switzerland, 2020; Volume 87, pp. 77–101. [Google Scholar] [CrossRef]
- Hakkari, L. Zooplankton species as indicators of environment. Aqua Fennica 1972, 1, 46–54. [Google Scholar]
- Andronikova, I. Zooplankton characteristics in monitoring of Lake Ladoga. Hydrobiologia 1996, 322, 173–179. [Google Scholar] [CrossRef]
- Ejsmont-Karabin, J. The usefulness of zooplankton as lake ecosystem indicators: Rotifer trophic state index. Pol. J. Ecol. 2012, 60, 339–350. [Google Scholar]
- Ejsmont-Karabin, J.; Karabin, A. The suitability of zooplankton as lake ecosystem indicators: Crustacean trophic state index. Pol. J. Ecol. 2013, 61, 561–573. [Google Scholar]
- Ochocka, A.; Pasztaleniec, A. Sensitivity of plankton indices to lake trophic conditions. Environ. Monit. Assess. 2016, 188, 622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpowicz, M.; Sługocki, Ł.; Kozłowska, J.; Ochocka, A.; López, C. Body size of Daphnia cucullata as an indicator of the ecological status of temperate lakes. Ecol. Indic. 2020, 17, 106585. [Google Scholar] [CrossRef]
- Neal, C.; Neal, M.; Wickham, H. Phosphate measurement in natural waters: Two examples of analytical problems associated with silica interference using phosphomolybdic acid methodologies. Sci. Total Environ. 2000, 251, 513–542. [Google Scholar] [CrossRef]
- Carlson, R.E. A trophic state index for lakes. Limnol. Oceanogr. 1977, 22, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Górniak, A. A new version of the Hydrochemical Dystrophy Index to evaluate dystrophy in lakes. Ecol. Indic. 2017, 78, 566–573. [Google Scholar] [CrossRef]
- Błędzki, L.A.; Rybak, J.I. Freshwater Crustacean Zooplankton of Europe; Springer: Cham, Switzerland, 2016; pp. 1–918. [Google Scholar]
- Ejsmont-Karabin, J. Empirical equations for biomass calculation of planktonic rotifers. Pol. Arch. Hydrobiol. 1998, 45, 513–522. [Google Scholar]
- Chao, A. Non-parametric estimation of the number of classes in a population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar]
- Muirhead, J.R.; Ejsmont-Karabin, J.; MacIsaac, H.J. Quantifying rotifer species richness in temperate lakes. Freshw. Biol. 2006, 51, 1696–1709. [Google Scholar] [CrossRef]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples. Version 7.5; User’s Guide and Application. 2005. Available online: http://purl.oclc.org/estimates (accessed on 1 November 2020).
- Ejsmont-Karabin, J. 2014-Rotifer invasion? On appearance and abundance of tropical species in lakes of North-Eastern Poland. Pol. J. Ecol. 2014, 62, 727–733. [Google Scholar]
- Benzie, J.A.H. The genus Daphnia (including Daphniopsis) (Anomopoda: Daphniidae). In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World 21; Kenobi Productions, Ghent & Backhuys Publishers: Leiden, The Netherlands, 2005; p. 376. [Google Scholar]
- Petrusek, A.; Hobæk, A.; Nilssen, J.P.; Skage, M.; Černý, M.; Brede, N.; Schwenk, K. A taxonomic reappraisal of the European Daphnia longispina complex (Crustacea, Cladocera, Anomopoda). Zool. Scr. 2008, 37, 507–519. [Google Scholar] [CrossRef]
- Hudec, I. Anomopoda, Ctenopoda, Haplopoda, Onychopoda (Crustacea: Branchiopoda). Fauna Slovenska; VEDA: Bratislava, Slovakia, 2010; Volume 3, p. 496. [Google Scholar]
- Leibold, M.A.; Tessier, A.J. Contrasting patterns of body size for coexisting Daphnia species that segregate by habitat. Oecologia 1991, 86, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Brosseau, C.J.; Cline, T.J.; Cole, J.J.; Hodgson, J.R.; Pace, M.L.; Weidel, B.C. Do Daphnia use metalimnetic organic matter in a north temperate lake? An analysis of vertical migration. Inland Waters 2012, 2, 193–198. [Google Scholar] [CrossRef]
- Brzeziński, T.; Fronk, J.; Trzcińska-Danielewicz, J.; Dawidowicz, P. Interspecific hybridization in sympatric species of Daphnia inhabiting lakes in north-eastern Poland. Oceanol. Hydrobiol. Stud. 2012, 41, 1–6. [Google Scholar] [CrossRef]
- Lityński, A. Próba klasyfikacji biologicznej jezior Suwalszczyzny na zasadzie składu zooplanktonu [Biological classification of lakes of the Suwałki District based on zooplankton composition]. Spraw. Stacji Hydrob. Wigrach 1925, 1, 37–56. (In Polish) [Google Scholar]
- Kamiński, M. Lake Wigry, the lake “adopted” by international association of theoretical and applied limnology (SIL “Lake Adoption” Project). Pol. J. Ecol. 1999, 47, 215–224. [Google Scholar]
- Taylor, D.J.; Ishikane, C.R.; Haney, R.A. The systematics of Holarctic bosminids and a revision that reconciles molecular and morphological evolution. Limnol. Oceanogr. 2002, 47, 1486–1495. [Google Scholar] [CrossRef] [Green Version]
- Kotov, A.A.; Ishida, S.; Taylor, D.J. Revision of the genus Bosmina Baird, 1845 (Cladocera: Bosminidae), based on evidence from male morphological characters and molecular phylogenies. Zool. J. Linn. Soc. 2009, 156, 1–56. [Google Scholar] [CrossRef] [Green Version]
- Hołyńska, M.; Dimante-Deimantovica, I. Redescription and taxonomic notes on Cyclops bohater Koźmiński, 1933 and Cyclops lacustris G.O. Sars, 1863 (Arthropoda, Crustacea), with an identification key to the Cyclops species of Fenno-Scandinavia. Eur. J. Taxon. 2016, 212, 1–31. [Google Scholar] [CrossRef] [Green Version]
- Hołyńska, M. On the morphology and geographical distribution of some problematic South Palearctic Cyclops (Copepoda: Cyclopidae). J. Nat. Hist. 2008, 42, 2011–2039. [Google Scholar] [CrossRef]
- Hołyńska, M.; Wyngaard, G.A. Towards a phylogeny of Cyclops (Copepoda): (In) congruences among morphology, molecules and zoogeography. Zool. Scr. 2019, 48, 376–398. [Google Scholar] [CrossRef]
- Litvinchuk, L.F.; Litvinchuk, S.N. Morphological diversity and widespread hybridization in the genus Bythotrephes Leydig, 1860 (Branchiopoda, Onychopoda, Cercopagidae). Arch. Biol. Sci. 2016, 68, 67–79. [Google Scholar] [CrossRef]
- Korovchinsky, N.M. Further revision of the genus Bythotrephes Leydig (Crustacea: Cladocera: Onychopoda): Redescription of B. brevimanus Lilljeborg, reevaluation of B. cederströmii Schödler, and description of a new species of the genus. Zootaxa 2018, 4379, 347–387. [Google Scholar] [CrossRef]
- Korovchinsky, N.M. Morphological assessment of the North Eurasian interspecific hybrid forms of the genus Bythotrephes Leydig, 1860 (Crustacea: Cladocera: Cercopagididae). Zootaxa 2019, 4550, 340–356. [Google Scholar] [CrossRef] [PubMed]
- Fenchel, T.; Finlay, B.J. The ubiquity of small species: Patterns of local and global diversity. BioScience 2004, 54, 777–784. [Google Scholar] [CrossRef]
- Finlay, B.J.; Fenchel, T. Cosmopolitan metapopulations of free-living microbial eukaryotes. Protist 2004, 155, 237–244. [Google Scholar] [CrossRef]
- Hairston, N.G., Jr. Zooplankton egg banks as biotic reservoirs in changing environments. Limnol. Oceanogr. 1996, 41, 1087–1092. [Google Scholar] [CrossRef]
- Schröder, T. Colonizing Strategies and diapause of planktonic rotifers (Monogononta, Rotifera) during aquatic and terrestrial phases in a floodplain (Lower der Valley, Germany). Internat. Rev. Hydrobiol. 2001, 86, 635–660. [Google Scholar] [CrossRef]
- Dumont, H.J. Workshop on taxonomy and biogeography. Hydrobiologia 1980, 73, 205–206. [Google Scholar] [CrossRef]
- Dumont, H.J. Biogeography of rotifers. Hydrobiologia 1983, 104, 19–30. [Google Scholar] [CrossRef]
- Arora, J.; Mehra, N.K. Species diversity of planktonic and epiphytic rotifers in the backwaters of the Delhi segment of the Yamuna River, with remarks on new records from India. Zool. Stud. 2003, 42, 239–247. [Google Scholar]
- Kutikova, L.A. Kolovratki Fauny SSSR. [Rotifer Fauna of USSR]; Nauka: Leningrad, Russia, 1970; p. 742. [Google Scholar]
- Belmonte, G. Species richness in isolated environments: A consideration on the effect of time. Biodivers. J. 2012, 3, 273–280. [Google Scholar]
- Zhang, H.H.; Chen, S.N.; Huang, T.L.; Ma, W.X.; Xu, J.L.; Sun, X. Vertical distribution of bacterial community diversity and water quality during the reservoir thermal stratification. Int. J. Environ. Res. Public Health 2015, 12, 6933–6945. [Google Scholar] [CrossRef] [Green Version]
- Rahkola, M.; Avinsky, V.; Holopainen, A.-L.; Jurvelius, J.; Karjalainen, J.; Viljanen, M. Interacting in the dark: A study of the diel vertical migrations of plankton and fish in Lake Ladoga. Boreal Environ. Res. 1999, 4, 245–255. [Google Scholar]
- Kiørboe, T. How zooplankton feed: Mechanisms, traits and trade-offs. Biol. Rev. 2011, 86, 311–339. [Google Scholar] [CrossRef]
- Karabin, A. Pelagic zooplankton (Rotatoria + Crustacea) variation in the process of Lake Eutrophication. I. Structural and Quantitative Features. Ekol. Pol. 1985, 33, 567–616. [Google Scholar]
- Jeppesen, E.; Noges, P.; Davidson, T.A.; Haberman, J.; Noges, T.; Blank, K.; Lauridsen, T.; Sondergaard, M.; Sayer, C.; Laugaste, R.; et al. Zooplankton as indicators in lakes: A scientific based plea for including zooplankton in the ecological quality assessment of lakes according to the European water framework directive (WFD). Hydrobiologia 2011, 676, 279–297. [Google Scholar] [CrossRef]
- Gliwicz, Z. Food thresholds and body size in cladocerans. Nature 1990, 343, 638–640. [Google Scholar] [CrossRef]
- Zaret, T.M.; Suffern, J.S. Vertical migration in zooplankton as a predator avoidance mechanism. Limnol. Oceanogr. 1976, 21, 804–813. [Google Scholar] [CrossRef]
- Brooks, J.L.; Dodson, S.I. Predation, body size and composition of plankton. Science 1965, 150, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Lass, S.; Boersma, M.; Spaak, P. How do migrating daphnids cope with fish predation risk in the epilimnion under anoxic conditions in the hypolimnion. J. Plankton Res. 2000, 22, 1411–1418. [Google Scholar] [CrossRef]
- Robidoux, M.; del Giorgio, P.; Derry, A. Effects of humic stress on the zooplankton from clear and DOC-rich lakes. Freshw. Biol. 2015, 60, 1263–1278. [Google Scholar] [CrossRef]
- Yang, X.; Yuan, J.; Yue, F.-J.; Li, S.; Wang, B.; Mohinuzzaman, M.; Liu, Y.; Senesi, N.; Lao, X.; Li, L.; et al. New insights into mechanisms of sunlight- and dark-mediated high-temperature accelerated diurnal production-degradation of fluorescent DOM in lake waters. Sci. Total Environ. 2020. [Google Scholar] [CrossRef] [PubMed]
- Taipale, S.J.; Vuorioc, K.; Strandberg, U. Lake eutrophication and brownification downgrade availability and transfer of essential fatty acids for human consumption. Environ. Int. 2016, 96, 156–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rautio, M.; Korhola, A. Effects of ultraviolet radiation and dissolved organic carbon on the survival of subarctic zooplankton. Polar Biol. 2002, 25, 460–468. [Google Scholar] [CrossRef]
- Rautio, M.; Tartarotti, B. UV radiation and freshwater zooplankton: Damage, protection and recovery. Freshw. Rev. 2010, 3, 105–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpowicz, M.; Ejsmont-Karabin, J. Influence of environmental factors on vertical distribution of zooplankton communities in humic lakes. Ann. Limnol. Int. J. Lim. 2018, 54, 17. [Google Scholar] [CrossRef]
- Podshivalina, V.N.; Sheveleva, N.G.; Bayanov, N. Biology and Ecology of Holopedium gibberum (Branchiopoda: Cladocera: Ctenopoda) in the Palearctic. Hydrobiol. J. 2012, 48, 28–36. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| no. | Lake Name | Date | Latitude (N) | Longitude (E) | Surface (ha) | Max Depth (m) | Trophic Status | TSI | HDI | SDV (m) |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Białe Filipowskie | 29 July 2019 | 54°11′56″ | 22°38′59″ | 132.4 | 52.0 | oligo | 29.6 | 35.0 | 7.5 |
| 2 | Gaładuś | 29 July 2019 | 54°10′31″ | 23°25′22″ | 728.6 | 54.8 | oligo | 33.8 | 35.3 | 5.7 |
| 3 | Serwy | 29 July 2019 | 53°54′06″ | 23°12′15″ | 460.3 | 41.5 | oligo | 38.5 | 35.0 | 3.8 |
| 4 | Jegocin | 23 July 2019 | 53°39′52″ | 21°41′54″ | 127.4 | 36.1 | oligo | 39.7 | 37.7 | 7.0 |
| 5 | Leleskie | 24 July 2019 | 53°38′37″ | 20°50′38″ | 423.5 | 49.5 | oligo | 39.7 | 37.3 | 5.5 |
| 6 | Jaczno | 22 July 2015 | 54°16′48″ | 22°52′25″ | 41.0 | 19.0 | meso | 40.5 | 29.1 | 2.9 |
| 7 | Hańcza | 24 July 2015 | 54°15′48″ | 22°48′35″ | 311.4 | 108.5 | meso | 40.8 | 31.1 | 4.2 |
| 8 | Buwełno | 22 July 2019 | 53°52′48″ | 21°51′37″ | 360.3 | 49.1 | meso | 42.4 | 30.7 | 2.4 |
| 9 | Majcz Wielki | 31 July 2019 | 53°46′49″ | 21°27′14″ | 163.5 | 16.4 | meso | 42.8 | 34.1 | 2.7 |
| 10 | Kuc | 30 July 2019 | 53°49′12″ | 21°24′23″ | 98.8 | 28.0 | meso | 43.5 | 34.7 | 4.0 |
| 11 | Białe Wigierskie | 16 July 2018 | 54°01′53″ | 23°05′26″ | 100.2 | 34.0 | meso | 44.3 | 33.3 | 5.7 |
| 12 | Busznica | 19 July 2018 | 53°56′38″ | 23°05′00″ | 49.4 | 48.0 | meso | 44.3 | 34.8 | 6.6 |
| 13 | Szurpiły | 24 July 2015 | 54°13′43″ | 22°53′51″ | 89.0 | 46.8 | meso | 45.8 | 31.3 | 2.9 |
| 14.1 | Wigry, South Basin | 18 July 2018 | 54°00′54″ | 23°03′38″ | 2118.3 | 74.2 | meso | 45.4 | 30.9 | 5.2 |
| 14.2 | Wigry, Central Basin | 7August 2015 | 54°02′53″ | 23°05′40″ | 2118.3 | 74.2 | meso | 48.3 | 36.6 | 2.0 |
| 14.2 * | Wigry, Central Basin | 26 July 2016 | 54°02′53″ | 23°05′40″ | 2124.3 | 74.2 | meso | 45.9 | 32.3 | 3.2 |
| 14.3 | Wigry, North Basin | 7 August 2015 | 54°03′50″ | 23°04′50″ | 2118.3 | 74.2 | meso | 48.5 | 36.6 | 2.2 |
| 14.3 * | Wigry, North Basin | 26 July 2016 | 54°03′50″ | 23°04′50″ | 2118.3 | 74.2 | meso | 46.9 | 31.5 | 3.5 |
| 14.4 | Wigry Zadworze Bay | 7 August 2015 | 54°04′21″ | 23°05′09″ | 2118.3 | 74.2 | meso | 47.6 | 36.1 | 2.6 |
| 14.4 * | Wigry Zadworze Bay | 26 July 2016 | 54°04′21″ | 23°05′09″ | 2118.3 | 74.2 | meso | 44.8 | 33.2 | 4.9 |
| 15 | Probarskie | 30 July 2019 | 53°49′26″ | 21°22′40″ | 201.4 | 31.0 | meso | 46.3 | 33.6 | 3.8 |
| 16 | Brzozolasek | 1 August 2019 | 53°36′58″ | 21°44′14″ | 155.9 | 17.2 | meso | 49.4 | 36.0 | 1.2 |
| 17 | Kalwa | 24 July 2019 | 53°38′36″ | 20°45′27″ | 562.2 | 31.7 | meso | 49.6 | 34.8 | 2.0 |
| 18 | Okrągłe | 16 July 2018 | 54°01′14″ | 23°01′21″ | 10.7 | 4.6 | meso | 49.7 | 31.4 | 3.1 |
| 19 | Mikołajskie | 31 July 2019 | 53°47′22″ | 21°34′56″ | 497.9 | 25.9 | meso | 49.9 | 34.8 | 2.0 |
| 20 | Jagodne | 22 July 2019 | 53°55′19″ | 21°42′33″ | 942.7 | 37.4 | eu | 50.3 | 29.6 | 2.2 |
| 21 | Boczne | 31 July 2019 | 53°57′40″ | 21°44′46″ | 183.3 | 17.0 | eu | 51.1 | 32.2 | 1.4 |
| 22 | Wiartel | 1 August 2019 | 53°36′04″ | 21°41′49″ | 178.6 | 29.0 | eu | 51.7 | 37.7 | 1.6 |
| 23 | Ryńskie | 31 July 2019 | 53°54′43″ | 21°29′40″ | 670.8 | 50.8 | eu | 52.5 | 34.3 | 1.0 |
| 24 | Kierźlińskie | 24 July 2019 | 53°48′03″ | 20°44′32″ | 92.8 | 44.5 | eu | 53.1 | 32.7 | 1.5 |
| 25 | Długie Wigierskie | 18 July 2018 | 54°01′33″ | 23°01′23″ | 80.0 | 14.8 | eu | 53.5 | 30.3 | 1.6 |
| 26 | Leszczewek | 19 July 2018 | 54°04′21″ | 23°03′47″ | 21.0 | 6.5 | eu | 54.4 | 32.1 | 2.3 |
| 27 | Nidzkie | 23 July 2019 | 53°37′46″ | 21°32′38″ | 1818.0 | 23.7 | eu | 55.4 | 35.4 | 0.9 |
| 28 | Garbaś | 24 July 2018 | 54°08′05″ | 22°37′20″ | 140.6 | 48.0 | eu | 58.0 | 30.1 | 2.0 |
| 29 | Necko | 24 July 2018 | 53°51′47″ | 22°57′50″ | 400.0 | 25.0 | eu | 58.9 | 32.2 | 1.1 |
| 30 | Miłkowskie | 22 July 2019 | 53°56′31″ | 21°52′14″ | 23.7 | 15.0 | eu | 59.4 | 32.3 | 0.5 |
| 31 | Juno | 31 July 2019 | 53°53′38″ | 21°17′47″ | 380.7 | 33 | eu | 62.9 | 32.7 | 0.7 |
| 32 | Widne | 28 July 2016 | 54°00′44″ | 23°07′25″ | 1.9 | 4.0 | dy | 57.5 | 58.6 | 1.4 |
| 33 | Klimunt | 26 July 2019 | 53°42′23″ | 21°26′55″ | 12.8 | 4.0 | dy | 63.8 | 60.4 | 0.3 |
| 34 | Wesołek | 25 July 2019 | 53°35′33″ | 21°30′44″ | 7.0 | 3.0 | dy | 50.7 | 66.9 | 1.2 |
| 35 | Zdrużno | 26 July 2019 | 53°38′13″ | 21°20′59″ | 6.8 | 5.0 | dy | 51.2 | 69.8 | 1.8 |
| 36 | Suchar Wielki | 28 July 2016 | 54°01′40″ | 23°03′20″ | 11.0 | 9.6 | dy | 50.9 | 67.1 | 2.5 |
| 36 * | Suchar Wielki | 18 July 2018 | 54°01′40″ | 23°03′20″ | 11.0 | 9.6 | dy | 56.7 | 68.2 | 2.2 |
| 37 | Suchar I | 19 July 2018 | 54°05′07″ | 23°00′54″ | 1.3 | 2.0 | dy | 62.0 | 66.5 | 1.1 |
| 38 | Suchar II | 28 July 2016 | 54°05′14″ | 23°01′03″ | 2.6 | 9.5 | dy | 53.7 | 69.4 | 1.9 |
| 38 * | Suchar II | 19 July 2018 | 54°05′14″ | 23°01′03″ | 2.6 | 9.5 | dy | 58.1 | 71.1 | 1.5 |
| 39 | Wądołek | 28 July 2016 | 54°06′39″ | 23°02′38″ | 1.2 | 15.0 | dy | 70.0 | 68.6 | 1.2 |
| 39 * | Wądołek | 24 July 2018 | 54°06′39″ | 23°02′38″ | 1.2 | 15.0 | dy | 62.9 | 70.0 | 1.1 |
| 40 | Dembowskich | 28 July 2016 | 54°02′18″ | 23°03′33″ | 3.1 | 3.5 | dy | 55.2 | 74.5 | 2.5 |
| 41 | Sęczek | 23 July 2019 | 53°43′41″ | 21°32′47″ | 3.8 | 3.5 | dy | 56.0 | 72.2 | 0.9 |
| 42 | Gryżlewskie | 23 July 2019 | 53°43′26″ | 21°33′03″ | 4.3 | 5.0 | dy | 50.7 | 71.4 | 1.8 |
| 43 | Borkowskie | 23 July 2019 | 53°43′16″ | 21°32′57″ | 2.9 | 5.0 | dy | 53.7 | 73.2 | 1.2 |
| 44 | Kruczy Staw | 25 July 2019 | 53°39′41″ | 21°24′21″ | 2.1 | 8.0 | dy | 47.7 | 73.3 | 2.0 |
| 45 | Kruczek | 25 July 2019 | 53°39′36″ | 21°24′08″ | 4.2 | 4.0 | dy | 51.3 | 74.2 | 1.5 |
| 46 | Kruczek Mały | 25 July 2019 | 53°39′28″ | 21°25′01″ | 2.6 | 9.0 | dy | 46.7 | 74.6 | 1.7 |
| 47 | Konopniak | 29 July 2019 | 53°35′07″ | 21°33′09″ | 9.5 | 4.0 | dy | 50.4 | 79.9 | 1.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karpowicz, M.; Ejsmont-Karabin, J. Diversity and Structure of Pelagic Zooplankton (Crustacea, Rotifera) in NE Poland. Water 2021, 13, 456. https://doi.org/10.3390/w13040456
Karpowicz M, Ejsmont-Karabin J. Diversity and Structure of Pelagic Zooplankton (Crustacea, Rotifera) in NE Poland. Water. 2021; 13(4):456. https://doi.org/10.3390/w13040456
Chicago/Turabian StyleKarpowicz, Maciej, and Jolanta Ejsmont-Karabin. 2021. "Diversity and Structure of Pelagic Zooplankton (Crustacea, Rotifera) in NE Poland" Water 13, no. 4: 456. https://doi.org/10.3390/w13040456
APA StyleKarpowicz, M., & Ejsmont-Karabin, J. (2021). Diversity and Structure of Pelagic Zooplankton (Crustacea, Rotifera) in NE Poland. Water, 13(4), 456. https://doi.org/10.3390/w13040456