1. Introduction
Grass carp,
Ctenopharyngodon idella (Valenciennes, 1844) were originally imported from Asia to the Mississippi River Basin for control of macrophytes. Nowadays, they are one of the most notorious invasive species of aquaculture in the central United States [
1]. Different types of barrier and deterrent have been proposed to control the spread of such invasive species. However, the efficacy of each strategy still remains inconclusive and requires further investigation [
2,
3,
4]. Among them, sound barriers are typical non-physical barriers which inject traveling pressure waves as a stimulus to block carp passage and confine their habitat [
4]. Sound barriers are relatively easy to implement at relatively low cost, without causing any physical change in water flow or damaging the surrounding environment, which makes them a promising strategy to control invasive carp species [
3].
Another advantage of acoustic stimuli is that carps have greater capacity to detect a wide frequency range of sound compared to most fish inhabiting in Midwest Rivers, which could allow for the design of selective acoustic barriers targeting only invasive species [
5,
6,
7]. Common Midwestern fishes such as Lake Sturgeon,
Acipenser fulvescens (Rafinesque, 1817) and Bluegill Sunfish,
Lepomis macrochirus (Rafinesque, 1819) detect sound up to 400 Hz in maximum, while carps can hear much broader frequency range depending on their taxonomy [
6,
8]. It has been reported that both Silver carp,
Hypophthalmichthys molitrix (Valenciennes, 1844) and bighead carp,
Hypophthalmichthys nobilis (Richardson, 1845) can detect up to 5000 Hz for pure tones and can even go further for mixed tones [
9]. Grass carp, in particular, have shown to alter their behavior in response to sounds of up to 1000 Hz [
10,
11].
Despite research efforts focused on the hearing ability of carp, the most effective frequency range of acoustic stimuli has not been yet defined. One major challenge is that results are highly dependent on experimental conditions, and the various techniques to define and analyze the response of individual specimens [
9,
10,
11,
12]. A consistent, robust experimental method is needed to apply to various circumstances with high reliability. In addition, compared to active research on hearing capabilities of mature carp, much less is known about the frequency range that carp can perceive, and respond to, during their early developmental stages [
13]. The reproduction strategy of grass carp is to release eggs directly into the ambient water and let them drift until hatched larvae find habitats suitable for their growth [
5]. If the navigation of carp larvae can be controlled with acoustic stimuli, it can limit the spread of carp from their early stages.
The main objective of this study is to analyze the behavior of grass carp larvae exposed to sound of different frequencies, using proven quantitative imaging techniques used in hydrodynamic studies, to obtain reliable conclusions about the relation between sound frequency and larvae response. We introduce a robust framework to quantify the response to acoustic signals by capturing their movements with a camera and identifying trajectories via particle-tracking velocimetry (PTV). The response of grass carp larvae to external (i.e., out-of-water) acoustic signals of 100 to 1000 Hz frequency range are analyzed based on two quantities: 1) bulk velocities at each frame computed by averaging the velocity of every larvae and 2) response ratio computed from the number of larvae experiencing large variation of velocities and acceleration at the tone frame.
2. Materials and Methods
2.1. Experimental Design
Fertilized grass carp eggs were collected and transported as described in Prada et al. [
14]. All procedures were authorized by the Illinois Department of Natural Resources. Larvae hatched in a recirculating flume at a controlled temperature of 23–24 °C and stayed there for 5 days (reaching developmental stages 37–39) before the present experiment to ensure that larvae developed an ability to swim and were able to displace to their preferred locations [
14,
15].
Experiments were conducted at the Ven Te Chow Hydrosystems Laboratory (VTCHL), at the University of Illinois at Urbana Champaign. Approximately 200 larvae were placed inside a fish tank with dimensions 0.5 m long and 0.25 m wide, with a water depth of 0.075 m. After an acclimation period, the response of larvae to acoustic stimuli was tested. Since we are interested in their response to external noise (i.e., environmental noise generated out of the water in the stream, rather than a signal generated underwater), we used computer speakers (Logitech LS21) installed outside of the tank as our source. Acoustic stimuli consisted of single tones ranging from 100–1000 Hz, varying in 100 Hz increments with a constant amplitude of 60 dB. The tones were generated using Matlab’s ‘sound’ function, creating a sinusoidal signal with the respective frequency between 100–1000 Hz. Each signal lasted for 1 s for every case. Reactions were recorded with two GoPro Hero 4 Black cameras (4K resolution, frame size of 3840 × 2160 pixels, 30 frames per second), installed above and to the side of the fish tank. Recording started 5 s prior to the sound and continued for another 5 s after the sound was ceased to capture baseline and recovery images.
2.2. Acquisition of Larval Trajectories
Recordings from a top-view camera were processed using a custom particle image processing Matlab routine to obtain larvae trajectories based on a two-stage process: (1) identification of larvae from individual frames and (2) matching larvae between two consecutive frames [
16]. Raw videos were converted into time series of individual grayscale frames. The background was removed through an image-preprocessing procedure to only leave motile larvae on the image. The larvae were initially specified as circular shapes without considering their tails, defining a radius of 2.5 mm (similar dimension as matured eggs, e.g., [
5,
17,
18]). While other geometries were considered, the spherical approach performed better at both recognizing a larger number of larvae (
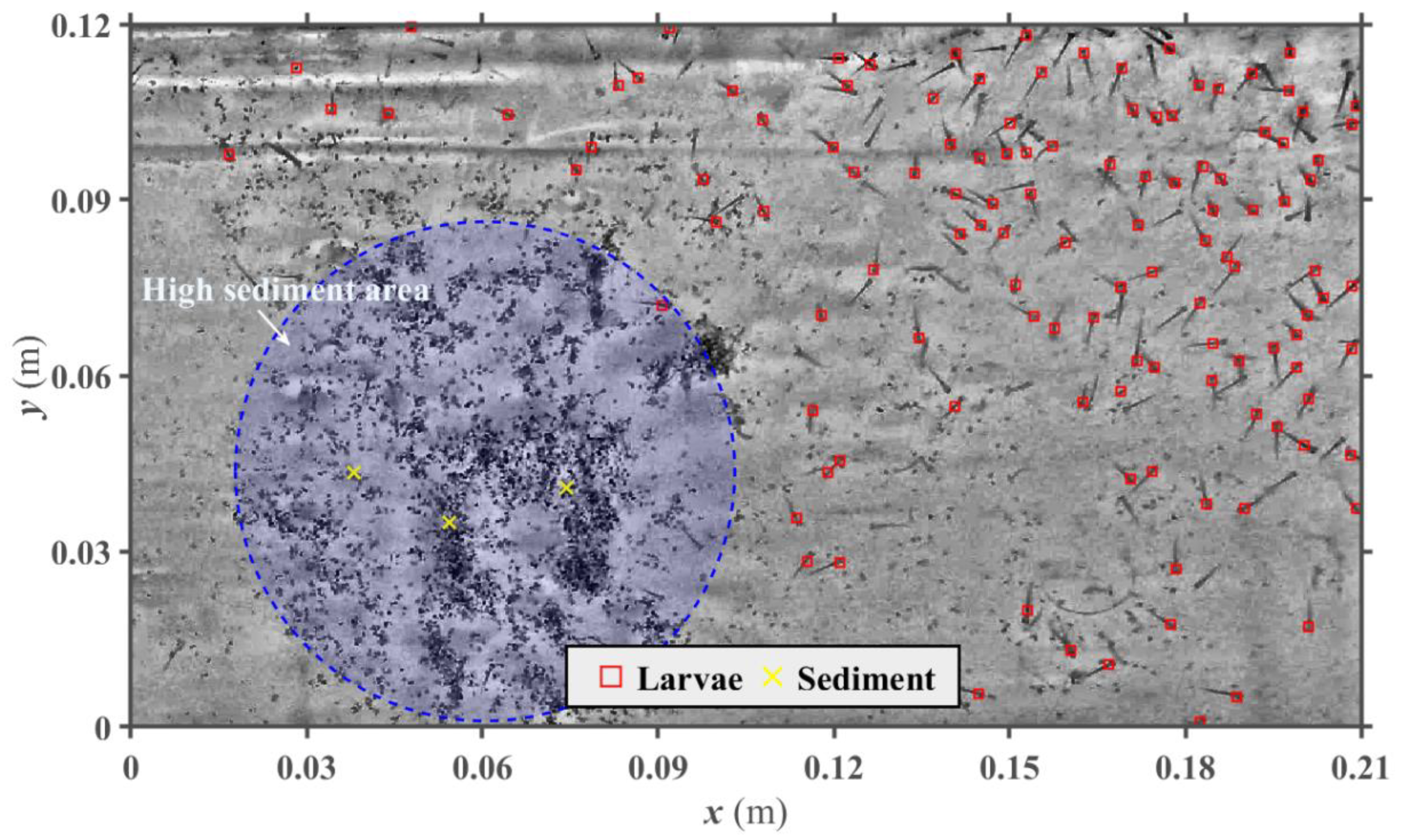
Figure 1) and experiencing a smaller “loss” of larvae between frames (i.e., missing a larva on consecutive frames if a larva moves or changes orientation). The spherical approach yielded the coordinates of the larvae centroids in each frame, which allowed us to proceed with particle matching for displacement calculations.
Figure 1 (top-view camera) shows the location where larvae were detected from an individual image, focusing on a highly populated region inside the fish tank. Although the algorithm missed some larvae, the majority was captured by their heads, and even closely located larvae were distinguished from each other as separate objects. The algorithm was also successful to capture larvae while neglecting other elements on the image, such as sediment and light reflections. While sediment grains were present in the fish tank (circle shown in
Figure 1), only a few were mis-identified as larvae (yellow cross-marks in
Figure 1).
In the second stage of the routine, the matching algorithm searched for larvae in consecutive frames to build trajectories over time, limited by a maximum distance that larvae can travel between two frames, and using the distribution of pixel intensities around larvae centroids for identification and matching. Since the experiment was conducted with still water conditions, the algorithm solely depended on larvae movements between two consecutive frames without considering any surrounding flow speeds. Displacements of larvae were quantified by velocity and acceleration based on the assumption that vertical movements are negligible, and larvae are moving in the horizontal plane. We use the magnitudes of velocity and acceleration, computed considering the components in the x and y directions. For consistency of the analysis, positions of larvae were tracked at identical numbers of images (30 frames) corresponding to prior to the sound stimuli, throughout the sound exposure and after the sound ceased. The instantaneous velocity and acceleration of larvae at each frame were quantified based on their displacement between frames.
2.3. Quantification of Larval Response to Sound Stimuli
The bulk behavior of larvae when exposed to abrupt sound stimuli are quantified by their velocity statistics at each frame. Mean velocity represents the overall movements of every identified larvae. To account for the largest displacements between frames, a 95th percentile was used to only account for extreme velocities, which reflects behaviors of larvae that are most active or sensitive to surrounding environments. Mean velocity and the 95 percentile are calculated based on Equations (1) and (2), respectively, where
is the mean velocity,
is the total number of larvae,
is the velocity of
larva and
is the rank of velocity to satisfy the 95th percentile (i.e., the 95th percentile,
would be located at the
position when velocities of each larva are aligned in descending order).
Mean and 95th percentile velocities are averaged over frames: (1) before the exposure to sound (1–29 frame), (2) during continuous sound (30–59 frame) and (3) after sound was ceased (60–89 frame) to quantify larval movements at each frame range and see if sound caused significant changes. Averages are computed according to Equations (3) and (4), where
is the averaged mean,
is the averaged 95 percentile,
is the number of frames,
is the initial frame and
is the final frame for each stage.
In order to represent velocities with dimensionless values and compare their variation at the tone frame between different larva, instantaneous velocities are divided by the maximum velocity of each larva, as shown in Equation (5), where
is the normalized velocity of
larva at
frame,
is the instantaneous velocity of
larva at
frame and
is the maximum velocity of
larva during 90 frames.
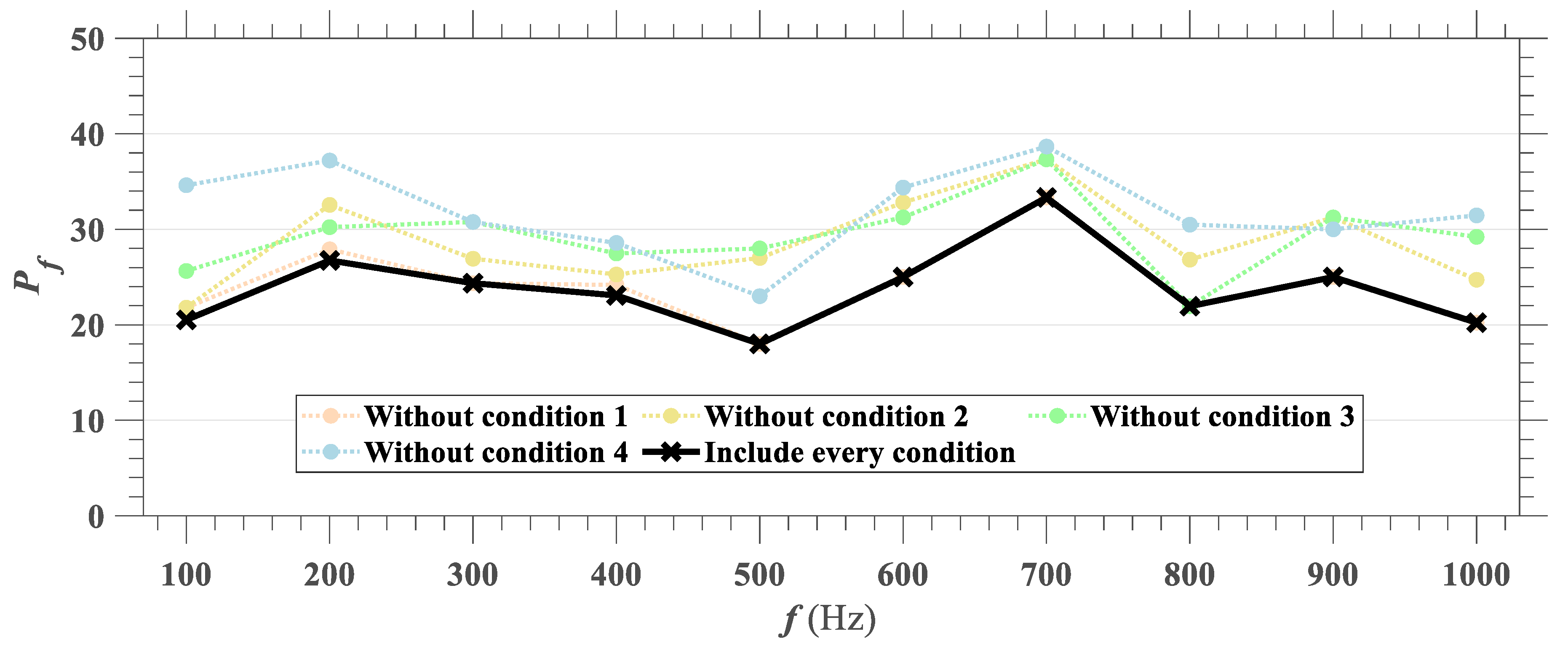
Another method of quantification of larval response is to compute the ratio of larvae that change movements at the exposure to sound. We defined four conditions to detect a change of behavior based on instantaneous velocity and acceleration: (1) peak acceleration should occur close to the tone frame, (2) 90th percentile of instantaneous velocities during the duration of the tone should increase compared to those prior to the sound, (3) maximum velocity should occur when larvae were exposed to sound, and (4) the minimum velocity should exceed
for larvae to be considered as moving. When all four conditions were satisfied, it is considered as larvae ‘responding’ to sound stimuli. The four conditions are set to avoid considering larvae that were already moving before the signal, or those that experienced a change in speed before or long after the stimulus, to ensure the response was due to larvae perceiving the tone. Based on the four conditions established to define larval response when exposed to sound, the response ratio
, was calculated using Equation (6), where
is the number of larvae satisfying all four conditions.
4. Discussion
4.1. Analysis Based on Bulk Behavior of Larvae in Response to Sound Stimuli: Possibilities of Misrepresenting Individual Larval Behaviors
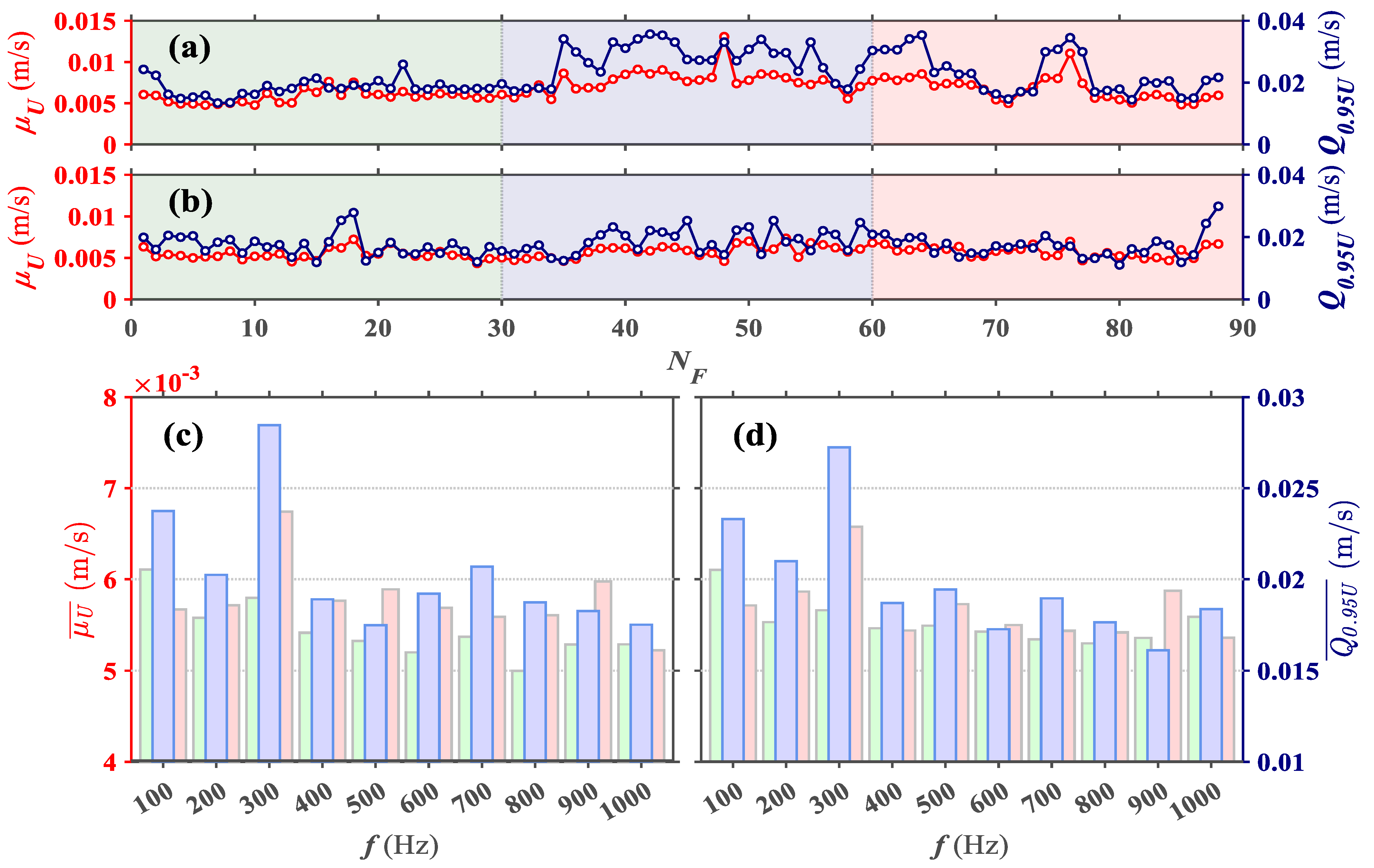
The analysis based on bulk quantifications of larval velocity could lead to conclusions that: (a) grass carp larvae can perceive tones in the range of 100–1000 Hz; (b) tones with frequency of 300 Hz could exert a more significant response in grass carp larvae; and (c) increasing frequencies past 300 Hz will not produce significant responses. However, large fluctuations in velocities among individual larva (
Figure 3a, with vertical error-bars representing variance of larval velocity), hints at misrepresentations of carp behavior using only a mean value. The large disparity of velocity trend between responding and non-responding larvae (
Figure 3b,c) demonstrates that bulk velocities are not sufficient to quantify the response of larvae to sound, since behaviors vary by individual. Although the larvae were exposed to identical acoustic stimuli at the same time, high velocities (marked by red dots in
Figure 3b) are concentrated right after the tone frame for larvae that are responding to sound, while high velocities (red dots in
Figure 3c) and low velocities (green dots in
Figure 3c) are randomly distributed for larvae that are not responding to sound. Thus, we suggest that number of individual larvae reacting to sound should be counted, and its ratio should be used to better quantify the extent of larvae response to sound stimuli.
4.2. Analysis Based on Individual Behavior of Larvae in Response to Sound Stimuli: Methodology Considerations to Improve Assessment of Larval Response
The most conspicuous difference between analysis based on bulk behaviors (i.e., quantification with mean and 95th percentile velocities) and individual behavior (i.e., quantification with response ratio) is whether carp larvae are reacting more actively at certain frequency ranges of sound. From the perspective of individual larval response, we didn’t observe any clear relation between frequency and response ratio, but rather a consistent proportion of larvae were reacting to sound stimuli, with response ratio ranging between 20–30% regardless of the frequency.
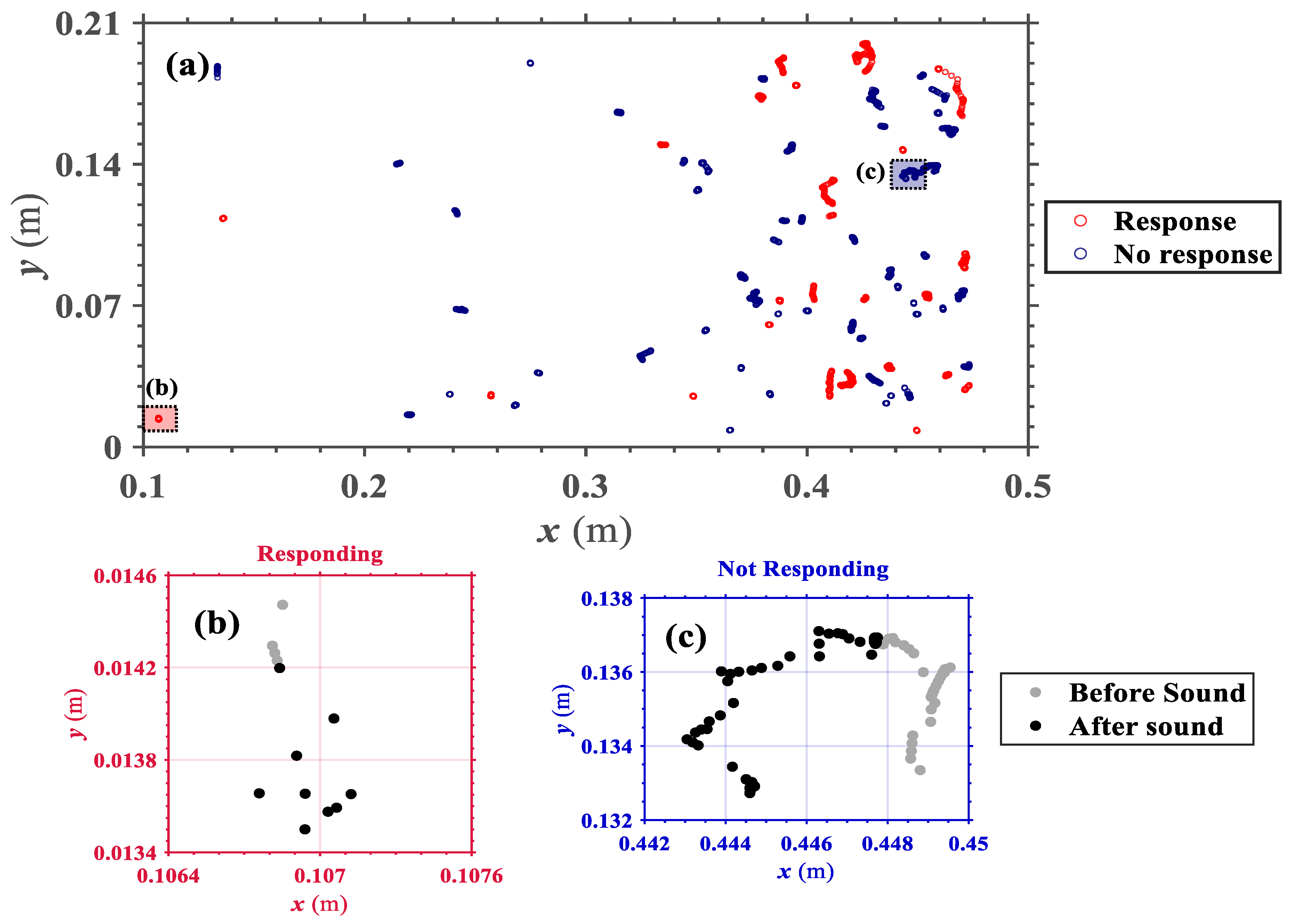
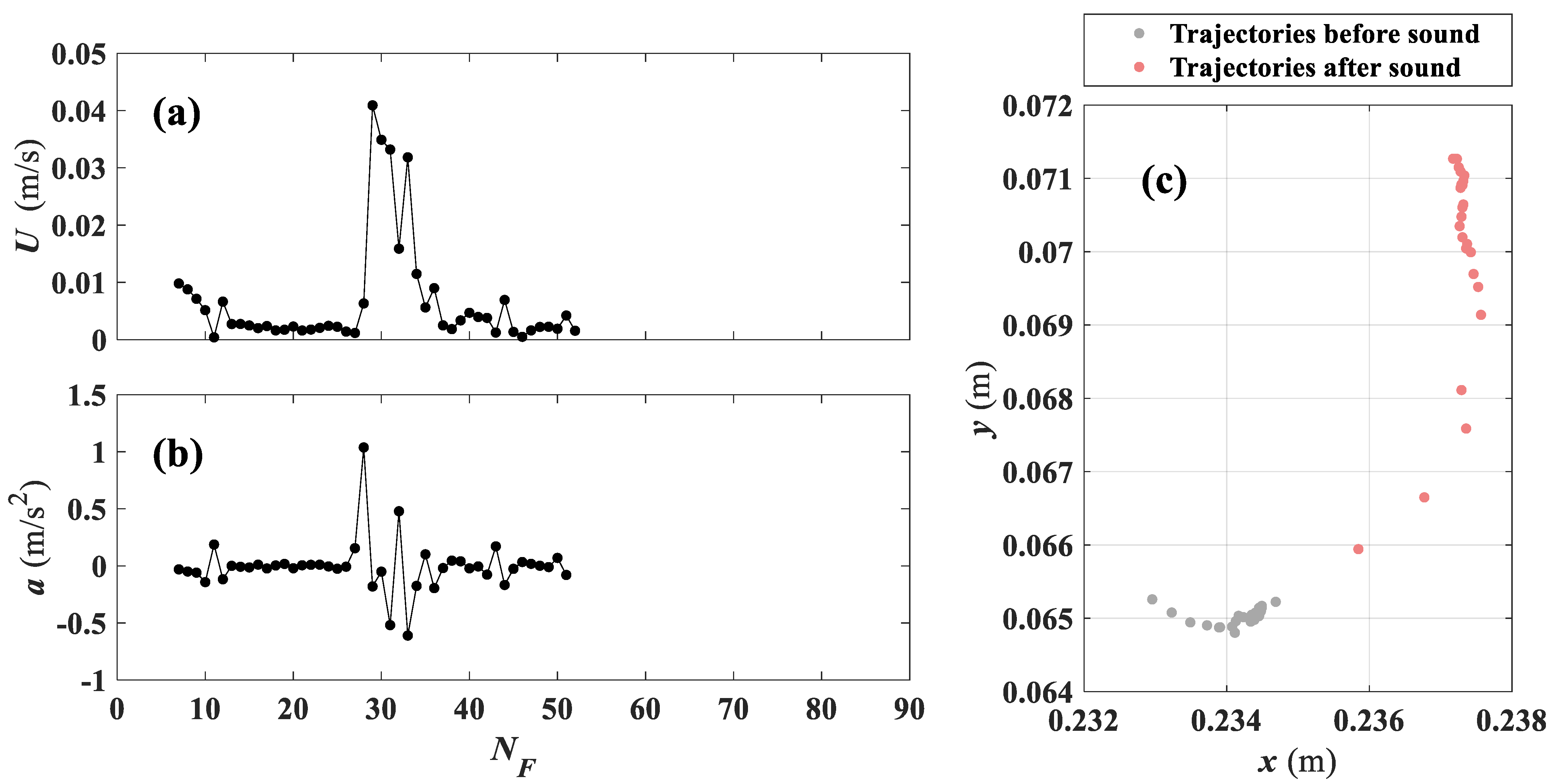
Looking at the individual response, time-series of velocity and acceleration for a single larva (
Figure 6), these show a clear change near the tone frame, for a larva responding to the tone. In
Figure 6a,b, velocities and accelerations are close to zero before the tone, which shows that movements of larvae were negligible. At the moment of exposure to sound, velocity jumps to a peak value and active larvae movements are maintained for a short period of time. A large fluctuation in acceleration is also observed at the tone. Trajectories of larvae in
Figure 6c also confirm the larval response to sound, as the distance between instantaneous position on each frame reaches a maximum right after larvae were exposed to sound (i.e., even if larvae were already moving prior to the tone, which would report a pre-existing velocity prior to the stimulus, they clearly accelerate when they perceive the sound). Thus, trajectories and time-series of velocity and acceleration are appropriate indices of larval response to sound which are considered to compute response ratio in this study.
The duration of the response varied within individuals. The larva shown in
Figure 6, for example, had a comparatively short duration of response, lasting only 4–5 frames. In general, the response lasted shorter than a second and died out before the sound ceased. Although we provide data from one typical larva in
Figure 6, peak velocities, acceleration observed near the tone frame, and larger displacements right after the sound were common behaviors among larvae that responded to sound.
As large number of larvae were exposed to sound at the same time, the interaction between neighboring larvae to generate additional motions were also observed. It is shown from
Figure 5 that the movement of each larva was independent from one another, and sudden initiation of motion among neighbors did not startle nearby larvae to start their motions as well. Thus, we concluded that sudden change in motion at the tone purely results from each larva acknowledging the sound, and their decision on how to react to that stimulus.
4.3. Limitations and Applications of Current Research
Several limitations remain in this type of studies, which requires future tasks to design effective sound barriers. Our experiment was designed to test larvae response to acoustic stimuli at still water condition while the actual habitat of carp larvae would be flowing rivers. Thus, we expect higher variability among larval responses to sound in the fields. Due to short duration of sound in our experiment, the consistency of larvae response was not considered, while carps’ fast adaptation to acoustic stimuli has been recently raised as a concern when implementing sound barriers [
19]. In addition, we only analyzed the response of larvae at pure tones whereas previous studies reported various effective designs of acoustic barriers such as exposing mixed tones and combining acoustic stimuli with other types of barrier [
2,
20].
No type of sensory deterrent systems can be 100% effective but different combinations of deterrents can provide an effective solution for expansion of invasive carp species in Midwest Rivers. Although the response ratio was comparatively moderate, our study showed that grass carp larvae can acknowledge acoustic signals, which suggests the applicability of sound to be used in a different combination of deterrents.
Although we only focused on the larval response to acoustic signal in this paper, efficacy of other types of deterrents can be also quantified based on similar framework of analysis based on PTV by tracking trajectories of larvae and analyzing the variation of velocities and acceleration when they are exposed to external stimuli. For actual application of deterrents in real rivers, field tests are required to consider the effect of diverse biotic and abiotic variables on the interaction between carps and deterrents. Our framework of analysis based on particle tracking velocimetry is advantageous in that it can be applied to both lab and field work since we only need the recordings of trajectories of carps to identify altered navigation due to external stimuli. We expect that the negative impact of invasive carps on freshwater ecosystem and surrounding morphodynamics can be alleviated by active future research on the efficacy of various deterrents, including acoustic stimuli, with careful consideration of both bulk and individual behaviors based on our proposed framework.
5. Conclusions
Grass carp larvae demonstrated a consistent response ratio ranging between 20–30% when exposed to acoustic signals of pure tones ranging from 100 to 1000 Hz. Maximum response ratio was calculated as 33% at 700 Hz, although we did not observe any clear relation between frequency and response ratio. This result differs from our observations based on mean bulk velocities, which showed a larger response at a 300 Hz tone. Sharply increased velocities at the tone frame did not last long, and larvae returned to the original inactive state even before the sound ceased. Larvae were also insensitive to movements of neighbors which indicates that sudden motions at the tone frame were caused purely as a reaction to the acoustic signal.
The most significant finding in this study is that bulk quantification of larval motion can misrepresent their actual sensitivity and response to acoustic signal. Analysis based on bulk velocities indicate that larvae are sensitive to sound of low frequencies while in fact, larval response was not prominent at any specific frequency ranges when analyzed based on individual trajectories. Consistent response ratio implies that several larvae were startled by sound even when bulk velocity increased negligibly at the tone frame. High standard deviation of velocities is another indicator to suggest that larval response to acoustic signals is highly dependent on the behavior of each individual larva. Our result points out that quantifications of an optimal or threshold frequency to alter grass carp behavior depend on how the ‘response’ is defined. High deviation among individual behaviors highlights that it is necessary to simultaneously test a sufficient number of carp rather than repeating the experiment with a limited number of individual specimens, which will help us determine the general sensitivity and overall response to acoustic signals.



 : mean and
: mean and  : 95th percentile at (a) f = 300 Hz and (b) f = 400 Hz where f is the frequency. Bar plot of (c) time averaged mean velocity and (d) time averaged 95 percentile velocity computed for
: 95th percentile at (a) f = 300 Hz and (b) f = 400 Hz where f is the frequency. Bar plot of (c) time averaged mean velocity and (d) time averaged 95 percentile velocity computed for  : frames 1–29 (before sound tone),
: frames 1–29 (before sound tone),  : frames 30–59 (during sound tone) and
: frames 30–59 (during sound tone) and  : frames 60–89 (after sound tone ceased).
: frames 60–89 (after sound tone ceased).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}