Abstract
The use of non-physical barriers, particularly based on acoustic and luminous stimuli has been historically used to influence the behavior of fish, mainly for fishing purposes. Nowadays, behavioral barriers and guidance systems have been developed, not only to deter movements of fish, but also to promote behavioral responses with the objective of native fish protection, in particular the potamodromous species, reducing their mortality in the hydraulic structures of dams and guiding them towards transposition systems or to replacement habitats in regularized water bodies. This review details the use of acoustic and luminous systems and their evolution in recent years (Scopus 2012–2019) for the development of selective behavioral barriers for fish. We found that recent technologies try to identify new acoustic and luminous sensory ranges. Ambient sound, sound of predators or luminous spectral bands with different wavelengths have been used to selectively stimulate target and non-target species, in order to improve the effectiveness of repulsive/attractive systems for fish. Guidelines for future research in the area are also present.
1. Introduction
Freshwater ecosystems of conservationist interest, known as hotspots of biodiversity, are particularly vulnerable to negative human activities (e.g., water pollution and eutrophication, water extraction, aquatic and riparian habitat degradation, river regulation and flow alterations) and, more recently, to climate change, which is responsible for an increase in the frequency and magnitude of drought and flood events [1,2,3]. Particularly, river fragmentation caused by the presence of instream obstacles is one of the major causes affecting the sustainability of native fish populations and contributing to the decline of more than 50% of threatened fish species in Europe [4]. The loss of river connectivity promoted by the construction of small and big dams is also accompanied by changes in flow and decrease in habitat heterogeneity and quality [3,5,6,7,8,9]. For example, in Iberia, during the past decades, not only increased the number of big dams (>15 m height), but also the number small hydropower plants and small weirs (in Portugal, it exists more than 8000 transversal obstacles, with less than 5 m height [10], contributing to the disruption of the longitudinal connectivity of lotic ecosystems [11,12]. Several researchers studied solutions to overcome or minimize this negative impact, such as through the implementation of fish passages [13,14,15,16]. However, often these structures represent a handicap for native potamodromous fish, namely to find the access and subsequently to transpose the obstacles during up and downstream reproductive migration. For these reasons, guidance systems can be crucial in order to increase the permeability of these infrastructures to species migration [17].
Various behavioral systems based on the use of sensory stimuli has been studied by several researchers with the objective to deter movements and protect fish from hydraulic traps as well as to increase the efficiency of transposition systems, through acoustic stimuli [18,19,20], luminous [21,22,23,24], electrical [25,26] chemical [27,28] among other systems. The effectiveness of these different stimuli is varied and depend essentially on the sensitive capacities of the species for a given stimulus, which may be related to their internal/external morphology, trophic chain, reproduction, or also related to environmental conditions [29,30].
The knowledge of the ecophysiology of target species, namely how they react to a given stimulus (acoustic, luminous or other) can be used to develop selective behavioral systems, either repulsive or attractive. High potential can be assigned to behavioral stimuli if they can be applied specifically to target species. In fact, the development of behavioral selectivity systems may help, in the future, for the protection and conservation of native threatened species. For instance, the longitudinal river connectivity and permeability in fragmented rivers is crucial for them to access particular areas, such as spawning zones, allowing the genetic flow in native populations. On the other hand, behavioral selectivity may prevent biological invasions and the introduction of undesirable species (e.g., invasive alien species) in priority habitats of rivers with high conservation value, avoiding population imbalance due, for example, to intense predation, hybridism or habitat competition.
The aspects related to the effectiveness of artisanal and industrial fishing methods dominated the first behavioral studies related to the sensory response of fish [31,32,33,34]. Fishing activity used this knowledge namely about hearing characteristics of fish species in a way to increase captures of commercial fish, but also to reduce catches of non-target fish [35]. Popper [31] recognizes that there is a growing demanding in fish attraction/repulsion systems, especially if they become cheap and reliable to the fishing industry. However, he adds that data in scientific literature presents ambiguous results. He also mentions that one of the most important and least studied issues is the nature and mechanism of fish habituation in relation to the stimuli used.
Noatch and Suski [36] makes a historical and extensive review of the available bibliography and they recognize the need to summarize and to compare existing and developing technologies, detailing the use and application of deterrence techniques: electrical, visual, acoustic, chemical and hydrological, which can be used to prevent fish movements. They consider that several options show some potential for the selective sorting of fish, by species or size, through behavioral and physiological mechanisms. They also indicate selectivity as a potential advantage in using systems based on species-specific alarm pheromones, or water velocity in its ability to select species/sizes according to their swimming abilities facing a deterrent effect. In the remaining stimuli analyzed in this study, namely acoustic and visual stimuli, the existence of references to selective filters, capable of developing selective behavioral systems by species or size, is not apparent. Noatch and Suski (2012) [36] is able to summarize and compare developments in methodologies and technologies associated with various behavioral systems for fish. The present work aims to continue an evolutionary analysis of new trends for those systems in recent years, although, specifically directed to acoustic and luminous stimuli. To this end, a survey of scientific publications of the last years (2012–2019) was carried out.
Thus, the main objective of this review is to highlight the recent use and development of selective behavior systems for fish based on acoustic, luminous and mixed stimuli (includes simultaneous stimuli), in a perspective of identifying selective filters, to understand the potential differentiating elements of the various types of stimuli for target or non-target species, to increase the effectiveness of repulsive/attractive behavioral systems. We also intend to identify the existing gaps and to register possible recommendations for future investigations, in order to provide a quicker development of this interesting and promising research line, towards the protection and safeguarding of threatened fish communities in freshwater regularized rivers.
2. Methods
A search was carried out in the Scopus database, which is the largest abstract and citation database of peer-reviewed research literature using specific keywords related to the present study, with the aim of identifying publications in the area-since 2012, with special emphasis on the most relevant ones (greater number of citations), considering the tested stimuli, target species, experimental methods and results. The research was carried out based on keywords insert in the “Article title” and “Abstract”, and further it was made a selection of the results obtained was made based on the relevance for the present review (objective of the study, freshwater species and diadromous migrators, analysis of fish behavior: repulsion, attraction, guide, stimuli used if only light, sound or mixed). The keywords considered are indicated in Table 1.

Table 1.
Keywords used in the search.
3. Results
Acoustic: The survey considering FIELD: Article title, Abstract, Keywords, returned 45 peer-reviewed articles. From these 45 articles, a selection was made based on the relevance for the present review (objective of the study, freshwater species and diadromous migrants, fish behavior analysis: repulsion, attraction, guide, stimuli used_ only sound or mixed effects) resulting in 22 articles considered relevant (Table 2).
Table 2.
Publications selected in this review, describing the stimulus tested, the intended effect, species tested, method, efficiency and number of citations.
Light: The search considering FIELD: Article title, Abstract, Keywords, returned 46 articles. A selection was made based on the relevance for the present review (objective of the study, freshwater species and diadromous migrators, analysis of fish behavior: repulsion, attraction, guide, stimuli used: only light and mixed) resulting in 16 articles considered relevant (Table 2).
Mixed: From the research carried out, in the 32 selected publications which included sound or light it was possible to identify nine articles with two or more combined stimuli: sound + light + bubble curtain; sound + light; sound + bubble curtain; light + bubble curtain, which deserved a particular analysis in this study (Table 2).
Another interesting aspect regarding the results obtained is the intended effect, which, in the case of acoustic stimulus, all studies (100%) corresponds to repulsive or orientation effects, whereas in the case of light-based systems, three studies were identified (19%) in which the intended effect is the attraction of fish.
In Figure 1, it is possible to observe graphically and summarized some relevant aspects listed in Table 1, as well as their geographic distribution.

Figure 1.
(A) Type and geographical distribution of publications, as well as the list of exotic/native species in each country. (B) Distribution of the total number of publications by type and by families. (C) Distribution of the main families for each of the studied stimuli. (D) Distribution of publications by year.
In Figure 1A it is possible to observe the geographical distribution of the selected publications, with special reference to the USA with 19 publications (six acoustic, one light, seven mixed and five studies) and 24 species tested, of which 83% are exotic, with emphasis on the cyprinids H. nobilis and H molitrix. Figure 1B indicates the distribution of families used in the experiments, by type of publication, with the acoustic stimulus having a greater number of publications (9) and number of trials with species/families: 20, with emphasis on cyprinids, tested in 19 experiences. Figure 1C shows the occurrence of the main families: cyprinids and salmonids in the various stimuli analyzed: Acoustic, Light and Mixed. In this graph we see that cyprinids occupy a prominent position in all studied stimuli. In Figure 1D we can see the distribution of publications over the years, with special emphasis on the 12 publications in 2019.
Following we present a more detailed analysis for each of the stimuli analyzed: Acoustic, light and mixed:
3.1. Acoustic
The analysis of the publications, based on the acoustic stimulus, allowed to identify 22 publications, namely: two reviews, tw directed studies (selectivity and sound frequencies) and 16 experimental studies. We observed that all had the objective of testing, in the fish species used, the repulsive or repulsive/orientation effect. No experiences were made with the objective of attraction. On the other hand, only three studies were carried out under field conditions, and the remaining ones were conducted under controlled laboratory conditions. Regarding the stimuli used in the experiments analyzed in this review, we can observe that the type of sound used was variable: modeled frequency—complex (sweep (2), outboard motor noise (8), animal (2), environment noise (1) and bubble curtains—while acoustic effect (2) and pure tone. It should be remarked that the modeled frequency (variation of sound frequencies (Hz) over time) was used in all experiments, and the pure tone was only used in a complementary way in four of the experiments performed. Considering the analyzed experiences, nine used isolated stimuli and nine used mixed stimuli, in this case with two or more stimuli simultaneously.
Two review articles, Noatch and Suski [36] and Putland and Mensinger [46], were published having different approaches:
(i) In Noatch and Suski [36] an in-depth review of the state of the art was made, regarding to the various types of behavioral stimuli for fish used in behavioral barriers. The authors referred that the use of acoustic stimuli, despite having shown little dependence on the effectiveness of the environmental conditions (e.g., turbidity or water flow), must be applied taking into account the local species in order define the respective frequencies to be used. Exposure levels with frequency ranges between 20 Hz and 20 KHz have been successful and, in contrast, the effectiveness of ultrasound (>20 KHz), was considered low or ineffective for most of the tested species. An approach to infrasound (<20 Hz) was also made, citing experiences with repulsive efficacy. This review also mentions the importance of obtaining a proper knowledge of the target species and their ecology, what is sometimes lacking in experiments. Two conclusions were presented: (a) future research must take into consideration the development of mixed systems with the simultaneous use of various stimuli (b) much of the research work in the area of behavioral barriers for fish was based on work carried out in field environment, which is generally difficult to validate, therefore, future studies should work intensively to validate and optimize behavioral systems under laboratory control conditions, prior to carrying out field tests.
(ii) Putland and Mensinger [46]: These authors made a very broad (1960–2019) and comprehensive review, oriented to the efficiency of the experiences according to the various taxonomic orders, making reference to some morphological aspects related to the auditory system that can affect those efficiencies. However, the authors mentioned that most studies do not allow conclusions to be drawn, as they did not indicate in a quantitative way the effectiveness of the studied acoustic stimulus, or whether there is a habituation to the stimulus.
Two publications assess cyprinid hearing range and the behavioral selectivity of fish species:
In the first study, Vetter et al. [47] characterize the hearing capacity of the cyprinid species: H. molitrix and H. nobilis, using auditory evoked potentials (AEPs). It was concluded that the effectiveness of the deterrent effect of acoustic stimuli was closely related to the auditory capacity of the tested species. These authors remark also that a frequency range between 100 Hz and 5 KHz comprised the sensitivity of these carps, identifying the frequency value 500 Hz as the lower threshold and 5 KHz (<150 dB re 1 μPa SPL rms) as the upper one for their hearing abilities. However, they admit that it is possible to find sensitivities greater than 5 KHz if values >150 dB re 1 μPa SPL rms are used.
In the second study, Rahel et al. [17] approached the selective fragmentation of fish and observed that the ideal acoustic filter would allow the passage of desirable species while preventing or greatly reducing the rate of passage of undesirable species ones. It also states that the development of selective fish passage requires that the desirable and undesirable species differ in one or more biological characteristics. These characteristics may influence fish’s ability to pass through a filter and can be related to fish movements (phenologies), namely physical capacity (temperature resistance, jumping, swimming), body morphology, sensory capacity, behavior and periods and periods of reproductive migration). These authors showed that acoustic barriers can be used as a potential filter, considering the morphological characteristics of the fish, namely in relation to species with or without a Weberian apparatus, which manifest different sensitivities in the presence of an acoustic stimulus. Thus, audible frequencies can be used for one species (target species) and not be audible for the remaining species present (non-target species). Finally, this work concludes that, by stating that the study of the species-specific sensitivity to sound, would make the development of selective passages for fish particularly attractive.
The efficiencies observed in the bibliography consulted are very varied, with the highest values corresponding to the cyprinid species: H. molitrix (93.7%) [37], L. bocagei (95.9%) [18] and H. nobili (>90%) [52]. The tested salmonids have low or very low efficiencies: O. tshawytscha (40%) [38] and S. trutta (8.7%) [18]. The effective stimuli for three cyprinid species showed to be complex acoustic stimuli (frequencies vary over time) and in one case [52] corresponded to a mixed stimulus (sound + light + bubble curtain). Murchy et al. (2017) [37] used in their experiments a complex sound from the audio recording of an outboard motor noise, producing broadband frequencies between 60 Hz and 10 KHz. The tests were carried out in a controlled environment and each test had 5 consecutive periods: (1) pre-sound (120 min); (2) sound playback 1 (30 min); (3) inter-sound (60–270 min); (4) sound playback 2 (30 min) and (5) post-sound (120 min). The results showed a high repulsive behavior for the two species tested: 82.5% (silver carp) and 93.7% (bighead carp). In the second referred case, Jesus et al. [18] used two acoustic stimuli in the experiments: sweep-up stimulus (up 2000 Hz) and tone pulses (140 Hz). The tests were performed in a controlled environment and each test, for each stimulus, had 4 periods: 15 min of a pre-playback control period (control), 15 min of sound playback (PBK1), 15 min of postplayback period (post-PBK), followed by 15 min of a second sound playback period (PBK2). The observed efficiencies were very different, with high efficiency for two cyprinids, L. bocagei and P. duriense, when the sweep up (2000 Hz) was used, with efficiencies of 95.9% and 87.7% respectively but lower efficiencies were obtained when the tone pulse was used (140 Hz) i.e., 27.4% and 30.71% respectively, for the mentioned species. In the case of the salmonid species, S. trutta, there were no records of repulsive efficiency for any of the stimuli used. In the third case mentioned, Ruebush et al. [52], tested the species H. nobilis and H. molitrix, with high efficiencies (especially for the bighead species) using a mixed system, which includes, in addition to sound, strobe light and bubble curtain and contrary to previous experiences, this study was carried out in-situ. Other species were also tested, and the authors observed that species with the Weberian apparatus had greater repulsive sensitivity than other species that do not have such apparatus.
The reproduction of animal sounds or of complex environmental sounds, appears in recent bibliography and tries to reproduce the deterrent effect that a known sound impact may have on the species, such as a sound related to a predatory animal [41,43], or a sound related to an impact recognized by the fish, such as a vessel’s engine [37,39,41,42,44,45,49,50]. In general, these complex sounds, and in particular the sounds that reproduce predators, have moderate to high efficacy.
3.2. Light
The analysis of the publications, based on the light stimulus allowed to identify 16 publications, namely: one review, two related to directed studies (vision study, selective filters) and 12 dealing with lab and field experiments. We observed that nine experiments were aimed towards the repulsive or repulsive/orientation effect and three implicated attraction behavior. On the other hand, only three experiments were carried out under field conditions, in spite that the majority of the studies was conducted in controlled laboratory conditions. Regarding the typology of the stimuli used in the experiments analyzed in this review, we can observe that 78% corresponded to the use of strobe light and only 12% use to continuous light, directed to induce attractive behavior, showing low or moderate efficacy. Although most of the experiments analyzed used white light, three experiments were observed under colored light beams [58,59,60].
Most studies with the light stimulus were carried out only during the day period, but four comprised both day and night periods. It should be also be emphasized that of the experiences analyzed, six used isolated stimuli and seven mixed stimuli, whereas two or more stimuli were applied simultaneously.
In Noatch and Suski [36] a careful review of the state of the art was made, considering publications with the various types of behavioral stimuli for fish until 2012. The authors referred that strobe lights, not being natural, promote repulsive behavior in fish. They also state that the reported efficacy, in relation to deterrence systems, with strobe light is variable and appears to be dependent on the life history of the target species, the brightness of the lights, water turbidity and environment light. However, the reported results showed to be very diverse when light is used in isolation, suggesting that strobe light may be more effective if combined with other behavioral stimuli, such as the bubble curtain.
The two directed studies analyzed the assessment of the visual capabilities of fish [56] and the behavioral selectivity of fish species present in the study sites [17]. Vetter et al. [56] proposed to optimize the effectiveness of non-physical barriers based on the use of strobe lights, either alone or simultaneously, with sound or with bubble curtains, with the study of visual sensitivity directed to the invasive species H. molitrix and H. nobilis. Their work indicates that, probably, the visual and spectral sensitivity and the environmental conditions (environment light and water turbidity), affect the behavior of fish and affect the efficiency of the strobe lights. It is suggested in this study that in dark (benthic) environments, the use of bright strobe flashes quickly affects the carp’s vision (the species studied) and that periods of visual recovery are longer with the duration of the flash. Rahel et al. [17] developed to the use of selectivity behavioral barriers, identifying some morphological characteristics or others that can act as filters in the selective behavior of fish. They indicate, in relation to light stimuli, that the morphological condition of most teleost fishes without mobile iris display a low ability to respond to rapid changes in light levels. In conclusion, strobe lights, which produce levels of unnatural light, induce repulsive reactions in fish. Despite mentioning in their study that fish species differ in repulsive behavior when faced with visual stimuli, they recognize that the potential for using these differences in the development of selective passageways remains largely untapped. Another interesting aspect to mention in this study is the approach to interspecific variation, this is, individuals of the same species that exhibit different or antagonistic behaviors, “outliers”, which can limit the effectiveness of the behavioral systems. Thus, they mention that in studies of high ecological sensitivity, in which efficiency is close to 100%, the limit values considered in the behavioral systems must take into account the interspecific variations of the target species. They propose the application of a safety factor in relation to the average values of the target species. They conclude by stating that, even when fish passage systems are only partially selective, they can play a very important role in the management of populations of undesirable species with a significant reduction in permeability rates.
The efficiencies observed in the bibliography consulted are very diverse: eight studies were identified in which the repulsive/attractive efficacy was low or moderate whereas the highest efficiencies corresponded to experiments involving S. trutta (>85%) [54], H. nobilis (>90%) [52], C. carpio (strong) [22]. Jesus et al. (2019) [54] used in the experiments two types of light frequency in isolation: 350 flashes/minute and 600 flashes/minute, in the day and night periods under laboratory conditions and tested separately three species: the salmonid S. trutta and the cyprinids L. bocagei and P. duriense. The tests were carried out in three phases: pre-test, test, and post-test. S. trutta was the species that showed the greatest repulsive behavior in the two stimuli tested, always showing greater efficiency during the day, but L. bocagei did not exhibit a repulsive behavior for any of the light stimuli used.
Ruebush et al. [52] used a mixed SBSLB stimulus through in-situ experiments, which included strobe light, sound and bubble curtain and compared this mixed technology with SBB, including sound and bubble curtain. They state that there is, apparently, no appreciable difference in effectiveness with the addition of strobe light in the SBSLB technology (in the range 500 Hz to 2000 Hz), both of which are very effective with Asian carp. Efficiency was assessed through recaptures upstream of the system, with previously marked individuals.
Kim et al. (2017) [22] used stroboscopic light (0.05–1.0 s flash rate) in their experiments in isolation, with the objective of promoting in the tested species (C. Carpio, A. nebulosus and M. salmoides) a repulsive effect. The tests were carried out in a controlled environment and each test consisted of three phases: control, treatment and post-treatment, with the use in each test of groups of three fish of each species, separately. The species C. Carpio and A. nebulosus, showed repulsive behavior, although C. Carpio, while M. salmoides did not exhibit a repulsive reaction under the strobe light in operation.
We report now the use of light stimuli to induce the effect of attraction in fish, in three studies analyzed [58,59,61]: Vowles et al. [61], used a continuous light at night to attract S. trutta into a downstream passage under controlled conditions and with various types of flow. They concluded that the attractive effect was not verified and, on contrary, light delayed fish orientation along the downstream passage. Lin et al. [58] evaluated the efficiency of several light colors (blue light, yellow light, white light and low light (control) to attract and capture fish of the species G. affinis. It was complemented with different flows and bubble curtain to assess the conditions of multimodal attractiveness. These authors showed that warm white light had greater attractiveness for the species tested, as well as the bubble curtain. Thus, they considered that multimodal systems, including flow, white light and bubble curtain can increase the efficiency of collecting fish after a previous attraction to the barge. Elvidge et al. [59], tested in laboratory conditions, a led strobe light device with various colors (green, blue, orange and white), evaluating the repulsive/attractive efficacy over A. fulvescens aged classes 1+ and 4+. They concluded that, for individuals aged 1+, blue strobe light may have an attractive effect and white strobe light a repulsive effect. For individuals aged 4+, the blue and white strobe light induced a repulsive effect, consequently we may consider that these ages negatively phototactic. These authors recognize, however, that the observations were based on a relatively small sample size and that further testing would is required for a proper validation.
The use of light stimuli through different colors (red, blue, etc.) appears in recent bibliography already mentioned and these works look to find behavioral differences in the fish tested when they face a varied light spectrum. The development of LED technology allowed to provide spectral stimulation at low cost, and it was possible to identify for each species their relative spectral sensitivity and behavior (attractive or repulsive). Hansen et al. (2018) [60], in their study, used combinations of spectra: red, green, blue and white light, in the night and RC and strobe frequencies in the day. Results were not conclusive, and they referred the need to carry out more studies related to the spectral sensitivity of the species tested.
3.3. Mixed
We analyzed publications where it was tested more than one stimulus simultaneously, with the aim of increasing the repulsive/attractive efficiency in the behavior of the tested fishes. The bubble curtain is the most applied complementary stimulus in this Mix typology (87.5%), being tested in seven of the eight experiments analyzed. The remaining complementary stimuli included strobe light (62.5%), sound (50%) and continuous light (12.5%). The simultaneous use of stimuli, sound, light and bubble curtain, was verified in 50% of the experiments. Only one of the experiments was designed to attract fish species, and the remaining ones were focused at the repulsive/guidance effect. Five experiments were carried out in a controlled laboratory environment and three of them under field conditions.
The efficiencies observed are very wide, with the highest values corresponding to the species C. carpio, [40,48], H. nobilis [52] (already described in “Acoustic”). In the study conducted by Zielinski et al. [40], it was analyzed the performance of three types of bubble curtains: fine, graded, and coarse-bubbles. This work is an innovative approach since it addresses other parallel stimuli to the visual ones characteristic of the bubble curtain, such as the sound emitted by the air of the tube diffusers. The authors demonstrated that, from the three bubble curtains tested, the system that emitted the thick bubble had the greatest repulsive efficacy, whereas the thin bubble exhibited the least repulsive efficacy. It stresses also the possibility that the parasitic sound emission generated in air diffusers (<300 Hz and <130 dB (ref 1 μPa)) may influence the repulsive behavior of C. carpio in shallow water, as the thin bubble curtain, coupled with an emitter it equivalent to the parasitic sound produced in the thick bubble curtain. This work included also the light stimulus together with the large bubble, although it did not result in an increase in efficiency; on the contrary, the stimulus was lower. The study concludes by suggesting that the role of visual stimuli is limited in altering the repulsive behavior in C. carpio.
The experience of Zielinski et al. [48] proposes a model to describe the movement of fish in the vicinity of a behavioral deterrence system and to examine what conditions would be necessary to alter the natural movement of C. carpio, using the bubble curtain (considering the parasitic acoustic stimulus already characterized). They concluded that the mathematical model may find general application in the evaluation of behavioral deterrence systems, particularly in the where long-term physical tests can be difficult to perform.
4. Discussion
4.1. Selectivity and Selective Filters
The need to guide and protect native fish species, in modified water courses (dams, weirs, water diversion, among others) threatened by habitat loss, prevented from accessing spawning, shelter and food areas, and by the introduction of invasive alien species, has intensified the search for solutions that aim to guide/attract/repel fish, minimizing those adverse effects. Physical barriers (e.g., grid) do not fully respond to this challenge, because in addition to interfering with the flow, are only selective regarding the size of the individuals, regardless of the fish species. Thus, management organizations and the scientific community have given greater relevance to behavioral (non-physical) barriers in order to respond as efficiently as possible to the challenge of safeguarding fish communities, especially in interventional water bodies and in face of the proliferation of invasive alien species [36].
Noatch and Suski [36] proposed a definition for non-physical deterrence systems: any non-solid stimulus or obstruction that discourages or prevents a selected species from passing through a target region. Most studied behavioral systems give special emphasis to acoustic and luminous stimuli taking advantage of the sensory and visual sensitivities of the fish in order to establish limits that allow defining behavioral selectivity. Over the years, it is possible to verify the development strategies for fisheries management based on behavioral systems due to the technological advance: for example, strobe light, despite being tested to change the behavior of fish for more than sixty years, stable technology systems able to operate in an underwater environment were only developed in the 90 s [62]. Noatch and Suski [36] mentioned in their study the future need to carry out more tests in the laboratory and under control conditions, given the low efficiencies recorded in most field studies conducted before 2012. In the bibliographic review of this study, most of the identified experiments were carried out under laboratory conditions (84%). The mentioned recommendation reflects the need to explore conveniently the integration of different types of sensory mechanisms of the target species to find specific behavioral differences in the individual and/or group of individuals, which may promote the behavioral selectivity required. As an example, is the analysis of more complex acoustic stimuli, integrating in the acoustic effect a predatory effect (vocalization of predators) [41,43]. On the other hand, it should be noted that, in recent years, light stimuli of different color ranges have been introduced in the experimental studies [58,59,60], increasing the sensitive specificity of each tested species in order to identify suitable selective filters. Anyway, the stroboscopic stimulus (white light) continues to deserve the preference of most researchers in experiments with visual/light stimuli.
4.2. Acoustic
The development of the use of acoustic stimuli to alter the behavior of fish lies essentially in their little dependence on environmental conditions [36,46], a reduced maintenance and low costs [63]. The effectiveness of the acoustic stimulus in a given fish species depends on the technical specifications characteristics of the stimulus and its suitability for the physiological and behavioral performance of the target and non-target species [46], besides, it must have strict conditions of repeatability. The presence of specialized auditory structures, such as the Weberian apparatus, enables fish species with superior auditory sensitivity in relation to species that do not have such apparatus [64], which constitutes a differentiating element between species, which should be explored in the search for selective filters in order to increase the effectiveness of behavioral barriers. Jesus et al. [18] explored exactly this possibility when comparing the results between the two families of Cyprinidae and Salmonidae, respectively with and without Weberian apparatus. It was observed that cyprinids, L. bocagei and P. duriense, presented a repulsive behavior far superior to that of the salmonid species, S. trutta, both facing the same acoustic stimuli, and concluded that it could be possible to develop behavioral systems with different responses to the same stimulus, considering the morphological and behavioral differences of the species present.
Complex sounds, with varying frequencies over time, such as sweeps [18], or the reproduction of surrounding sounds [37,39,41,42,44,45,49,50] or from animals [41,43], with a wider range of frequencies, have been suggested as the efficient solutions for most of the species, through the exploration of their wider auditory capacities and by avoiding potential habituation to a single frequency. To allow repeatability in the complex sounds, they must be properly characterized, not only in the ranges of frequencies they use (Hz), but also by the sound pressure level (SPL; re. 1 μPa). These sound characteristics can be used as selective filters between the target and non-target species, if their hearing abilities are known. Audiograms may allow to reveal some dimensions of the diversity of fish auditory capacities [65]. Their graphic representation establishes the relationship between frequencies (Hz) and sound pressure levels (SPL; re. 1 μPa). For example, in Figure 2 it is possible to verify the mentioned audiograms which show the differences in acoustic sensitivity between salmonids and cyprinids.
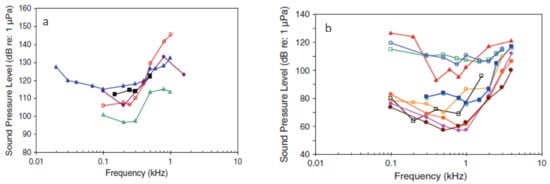
Figure 2.
(a) Audiogram of salmonid species: ▪ O. Mykissb, ჿ O. tshawytscha, ∆ C. laveratus, ∆ S. trutta, ◊ C. nasus. (b) Audiogram of cyprinid species: ▪ P. promelas, □ C. plumbeus, ∆ D. rerio, ◊ C. carpio, ● R. rutilus, ● P. parva, ჿ H. molitrix, ჿ C. carpio, □ A. Nobilis. Adapted from Ladich et al., 2013 [65].
In this figure it is possible to observe that cyprinids have a greater acoustic sensitivity, and that most species detect sound pressure levels below 90 dB (SPL; re. 1 μPa). On the contrary, salmonids have lower acoustic sensitivity and all species only detect sound pressure levels above 90 dB (SPL; re. 1 μPa). It should also be stressed that, despite these general trends, differences in sensitivity are visible within each family. Therefore, the development of audiograms for each species may be an essential tool in the development of selective barriers based on the acoustic sensitivities of species. For example, based on the audiograms in Figure 2, it will be possible to establish an acoustic behavioral barrier that only potentially interferes with the behavior of cyprinids, without exerting any acoustic stimulus in salmonids, provided that a range of sound pressure levels between 55 and 90 is used. dB (SPL; re. 1 μPa). On the other hand, if we consider the application of a frequency between 0.01 and 0.08 KHz and according to the audiograms in Figure 2, we may create a selective stimulus, which would only affect S. trutta, as long as the sound pressure level stays above 120 dB (SPL; re. 1 μPa). These differences in sensitivity can thus contribute to produce selective filters in the development of selective behavioral barriers.
Despite the physiological differences, determined by the elaboration of rigorous and credible audiograms, they are also of special importance for the determination of the acoustic sensitivity of the target species when we carry out tests in natural field conditions, but aquatic environmental variables must be considered too as they may interfere with the behavior of the fish [66,67,68]. For instance, the distribution of sound in limited tanks, built with artificial materials will be different from the distribution and decay of sound in natural field conditions, promoting different behavioral reactions [69]. Water temperature may influence also the acoustic sensitivity of fish [65,70], as well as the stress response, genetic history or surrounding noise [71,72].
These marked differences can really make the difference between the effectiveness of laboratory and field experiments. Murchy et al. [37], Vetter et al. [45], Vetter et al. [44] and Zielinski and Sorensen [50] observed high repulsive efficiencies with the use of a stimulus based on the “outboard motor noise” in the species H. nobilis and H. molitrix. However the field tests by Wamboldt et al. [39] observed low efficiencies with the same stimulus and for the same species and indicate the effect of the attenuation of the stimulus with depth as the main reason for these differences between laboratory and field tests. On the other hand, Zielinski and Sorensen [50], unlike Vetter et al. and Vetter et al. [44] identified habituation behaviors in their tests and indicated that, although using the same species and stimulus, differences in experimental protocol may be the cause of these apparent inconsistencies.
These examples testify to the limitations that still exist in the comparability of the tests performed, even with the use of the same species and stimulus. The absence of technical proportionality in the transition from laboratory to field tests, coupled with the high number of new variables to consider in field tests make the results unpredictable and apparently contradictory. This may mean, according to these examples, that despite the consistent evolution of laboratory tests with consistent results, the technical and scientific maturity to change this scenario has not yet been reached. In this regard, it should be noted that the effort of Zielinski and Sorensen [50], who in his experience seeks to evaluate, in the laboratory, the orientation/avoidance abilities of the species H. nobilis and H. molitrix, considering both the sound pressure (which they consider to play an important role in the early stages of repulsive behavior) or the movement of particles in water that may be responsible for guiding the movement of carp. These lines of investigation, along with others such as effective knowledge of the functions of the “lateral line”, improve the morphological and behavioral knowledge of the species present and will certainly be fundamental to increase the effectiveness of behavioral systems.
Another aspect to consider in future projects includes the elaboration of audiograms, not only of the target species, but of all the species present, thus promoting the proper knowledge of the acoustic sensitivities to find differentiating elements that allow to establish selective filters, in which they will only stimulate the target species, without affecting the other species and the integrity of the ecosystem.
4.3. Light
Light, widely used in experiments to induce an alteration of fish behavior is related to the visual and spectral sensitivity of the species and to the environmental conditions, such as surrounding light (daytime at night), turbidity [56] or the water temperature [73]. Fish, unlike most mammals, do not have a mobile iris, which reduces their ability to react to changes in light, seeking refuge in dark areas [36,74]. On the other hand, fish vision comprises photoreceptors specialized in the absorption and conversion of light into nerve impulses: rods and retinal cones. The rods are used for scotopic vision, in low light conditions, while the cones are used for photopic vision in bright light conditions [75]. The visual acuity of fish is closely related to the density of retinal cones, which varies greatly from species to species [73], whereas teleost fish increase their rod-to-cone ratio through life stage development [74] increasing their visual acuity. This aspect is very interesting because it allows the establishment of selective filters in visual stimuli based on the differences in the visual acuity of the fish according to their age. In this regard, Stenkamp et al. [75] suggested that adult fish are more sensitive to changes in light conditions than younger individuals, i.e., the age of the fish may have an differential influence in distinct age classes when exposed to light stimuli [74,76]. Therefore, this difference also constitutes another differentiating element to consider in the experiences for the development of behavioral systems based on light stimuli.
On the other hand, visual orientation behaviors, such as finding food and detecting partners and/or potential predators have been considered as the main drivers of the evolution of the eye, and have produced a multitude of ocular morphologies and visual strategies between taxa and environmental gradients [77,78,79,80,81]. These behavioral/morphological differences between species and/or habitats can also constitute additional elements for the identification of new selective filters.
As observed in this study the use of light, either in isolation or in a complementary way with another stimulus, is widely used or referenced in the bibliography, although the observed results are very varied in relation to its effectiveness. We analyzed two studies [22,49] with the particularity of testing in both studies the same specie: C. carpio, with very similar methodologies but different results; Bzonek et al. [49] observed in their experiments using strobe light (1–20 Hz) in an isolated way and mixed with the sound, that the repulsive efficacy tested only produced moderate behavior changes with strobe light. However, the authors mentioned some limitations of the tests, related to the lack of refuge and the large size of the wild fish used, in relation to the experimental tank. On the other hand, Kim et al. [22] using strobe light (0.05–1.0 s flash rate) found high repulsive efficacy, with significant differences between the control and emission of stroboscopic stimulus. However, the authors pointed out that the findings should not be extrapolated to other species or on a field scale, without further research. Based on the results obtained from these two experiments, carried out only during the day period, we can admit that the difference in repulsive effectiveness of the results between these studies may be related to the difference in the frequencies of the tested light stimuli, thus allowing to analyze and identify an efficient selective filter based on these differences between strobe light frequencies.
On the other hand, we analyzed a study using the same methodology and stimuli, but for different species: Jesus et al. [54] proposes in their experience (briefly described in the results chapter), the comparison of repulsive behavior between species from different families, but subject to exactly the same methodology, stimuli and test characteristics. Thus, it intended to identify the specific differences in behavior of each of the tested species (salmonid: S. trutta; cyprinids: P. duriense and L. bocagei), at the two stroboscopic stimuli of 350 and 600 flashes/minute. They observed significant differences in the efficacy (%) observed between families, specifically between the species S. trutta (87.87% 600 flashes/minute/day) and L. bocagei (27.06% flashes/minute/day). They conclude by admitting that the specific differences found are likely to be related to visual capacity and trophic factors. The high visual sensitivity of S. trutta may be related to the fact that this is a predominantly a visual predator [82] and a diurnal and twilight opportunistic [83] (period when the greatest repulsive efficacy was reported) [84] periods. Vinterstare et al. [81] mention in their study the importance of trophic and predatory factors in the evolutionary development of vision, as predators rely heavily on vision for the detection and capture of prey [85,86]. However, more trophic factors, other than predation, may be related to the low visual sensitivity to the light stimulus of the cyprinid L. bocagei, which is a species with a greater dependence on benthic and less illuminated habitats and, consequently, it presents other sensory systems better developed such as smell or hearing [87]. It is possible to observe in these experiences sensorial differences that must be explored in order to identify exactly the selective filters that allow to develop more effective behavioral systems.
The behavioral systems based on light stimulus, as they exploit the visual capacities of the fish, present as limiting factors for behavioral responses the conditions of the environment, mainly, the level of water turbidity. This aspect may influence the effectiveness of these systems in a natural setting in relation to laboratory results. Moreover, laboratory experiments often have poorly adjusted experimental protocols, either to the stimulus to be used, or to the species tested, which promotes behavioral deviations that are difficult to understand.
As it is possible to see in this review, the new lines of investigation apparently move towards the exploration of increasingly specific stimuli for each species, taking into account their visual acuity, as well as the environmental and habitat constraints that can influence. This specificity seeks the selectivity of behavioral systems and, consequently, their best effectiveness in safeguarding and defending these species.
4.4. Mixed
Mixed systems comprise the behavioral effects of the stimuli used, plus the potential synergy effect, such as, simultaneously, the effect of light or sound with the bubble curtain, which can be, according to some authors, more effective in controlling fish behavior [22,36,40,45,52,88].
The bubble curtain integrates 89% of the mixed experiences studied in this work. The curtain of small and air bubbles rising in the water column generates turbulence with recirculation currents [89,90], establishing an air/water interface that interferes with the propagation of sound and light, experiences refraction phenomena in propagation medium, with changes in speed and direction. This physical phenomenon may be the cause of the increased efficiency of the bubble curtain, when combined with other stimuli such as sound or light. Another aspect of interest in the analysis of bubble curtains is the emission of “parasitic” sound emitted in the passage of air in the holes of the diffuser sleeve [91,92,93], which can generate values up to 130 dB (ref 1 μPa) and frequencies up to 300 Hz [40]. Thus, we can consider that the bubble curtain includes in its structure a mixed system of visual and sound effects, with partial effects depending on the pressure exerted on the diffuser tube and on the diameter of the air outlet holes.
Mixed systems generally aim to disperse the repulsive/attractive effects with the use of several stimuli simultaneously and can, at certain frequencies or intensities, be redundant or, contrarily, reach sensitivities not achieved by the effects isolated the stimuli. Within this scope, if in one hand mixed systems may reach a greater range of sensitivities in the species present, but on the other one they reduce the possibility of controlling the specific selective filters for each species.
This apparent lack of selectivity of mixed systems does not reduce the high utility that these systems present in projects in which it is intended to remove as many species as possible, such as in the cooling water intake of a nuclear power plant. On the other hand, given their multimodal characteristics, mixed systems will have a greater capacity to limit the phenomenon of habituation characteristic of isolated systems.
In future terms, these systems are following the trend of the remaining isolated stimuli, with the use of more complex stimuli. Although the bubbles curtains remain the main option in the researched experiences, new visual and acoustic capacities are being successfully explored for this stimulus [40].
5. Conclusions
This work allowed to analyze, within each stimulus studied, a set of studies with more or less success in relation to the proposed objectives. Of this universe, 70% were performed under controlled laboratory conditions, with about 90% having the purpose of the repulsive effect of the stimulus and only about 10% having the objective of the attractive effect of the stimulus. Another interesting aspect to highlight is the fact that in the acoustic or mixed stimulus we observed the predominance of the use of complex and/or modeled sounds to the detriment of pure sounds. In the case of the light or mixed stimulus, this predominance was observed with the use of the strobe effect to the detriment of continuous light, the latter being used almost exclusively to obtain the attractive effect of the fish. The cyprinid family leads the use in tests with carp H. molitrix, H. nobilis and C. carpio, and in particular with acoustic stimuli, demonstrating the concern of the scientific community to find specific control mechanisms and filters for these species, considered in many areas of the globe as voracious invaders, with great negative impacts on the native fish communities of the areas colonizing.
It is notorious that there is almost no analysis of the fish’s habituation to the stimuli tested in the analyzed experiments until 2012 [36], a situation that remains after that date, although its importance in the results’ effectiveness is recognized, in spite that, different authors insistently mention the need to avoid this phenomenon. Future work should close this important gap, allowing to strengthen current knowledge by introducing specific analysis periods in the test protocols, both during stimulus in action and in the post-stimulus period.
Technological advances in areas such as acoustics or light have contributed and will in the near future be decisive to increase the knowledge of behavioral differentiation, thus allowing the development of selective filters that are increasingly specialized and directed to a specific species or set of species. It is evident in the reported studies, focused in the scope of this review, the importance given to the introduction of technology, in the future, either in the discovery of new visual sensitivities through the analysis of wavelength gradients, or in the search for new, more elaborate and complex acoustic stimuli.
Although these promising lines of research are yielding some interesting results, it still needs further studies in the laboratory phase. The variability of the observed results, managing to be contradictory in the levels of effectiveness, sometimes for the same species, may indicate the need to define well-characterized and truly reproducible methodologies, so that repeatability and validation reinforce the value of the results. It would be interesting for the scientific community to establish a set of theoretical sampling protocols, especially for acoustic and light stimuli, taking into account: preparation and adequate logistics of the experiments (material means and species tested), methodologies (phases, test times, periods of the day, stimuli and respective characterization with the definition of the parameters/units to be evaluated), to increase the potential for repeatability of tests with different age groups and/or with other species, whether native or non-native.
Author Contributions
Conceptualization, J.J., R.C. and A.T.; methodology, J.J., R.C. and A.T.; formal analysis, J.J., R.C. and A.T.; investigation, J.J.; writing—original draft preparation, J.J., R.C. and A.T.; writing—review and editing, J.J., R.C. and A.T.; project administration, J.J.; All authors have read and agreed to the published version of the manuscript.
Funding
This work is supported by National Funds by FCT—Portuguese Foundation for Science and Technology, under the project UIDB/04033/2020.
Institutional Review Board Statement
Ethical review and approval were waived for this study, due to the care and use of experimental animals complied with University of Trás-os-Montes e Alto Douro and Portuguese law: Decreto-lei nº 113/2013 de 7 de Agosto, animal welfare laws.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article or supplementary material.
Conflicts of Interest
The authors declare no conflict of interests.
References
- Aarts, B.G.; Van Den Brink, F.W.; Nienhuis, P.H. Habitat Loss as the Main Cause of the Slow Recovery of Fish Faunas of Regulated Large Rivers in Europe: The Transversal Floodplain Gradient. River Res. Appl. 2004, 20, 3–23. [Google Scholar] [CrossRef]
- Brink, K.; Gough, P.; Royte, J.; Schollema, P.P.; Wanningen, H. From Sea to Source 2.0. Available online: http://kalastajateselts.ee/fail/files_from_sea_to_source_2_0.pdf (accessed on 21 January 2021).
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J. Emerging Threats and Persistent Conservation Challenges for Freshwater Biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef]
- Northcote, T.G. Migratory Behaviour of Fish and Its Significance to Movement through Riverine Fish Passage Facilities. Fish Migr. Fish Bypasses 1998, 1, 3–18. [Google Scholar]
- Gozlan, R.E.; Karimov, B.K.; Zadereev, E.; Kuznetsova, D.; Brucet, S. Status, Trends, and Future Dynamics of Freshwater Ecosystems in Europe and Central Asia. Inland Waters 2019, 9, 78–94. [Google Scholar] [CrossRef]
- Boavida, I.; Díaz-Redondo, M.; Fuentes-Pérez, J.F.; Hayes, D.S.; Jesus, J.; Moreira, M.; Belmar, O.; Vila-Martínez, N.; Palau-Nadal, A.; Costa, M.J. Ecohydraulics of River Flow Alterations and Impacts on Freshwater Fish. Limnetica 2020, 39, 213–232. [Google Scholar]
- Boavida, I.; Santos, J.M.; Ferreira, T.; Pinheiro, A. Barbel Habitat Alterations Due to Hydropeaking. J. Hydro-Environ. Res. 2015, 9, 237–247. [Google Scholar] [CrossRef]
- Cortes, R.; Peredo, A.; Terêncio, D.P.; Sanches Fernandes, L.F.; Moura, J.P.; Jesus, J.J.; Magalhães, M.P.; Ferreira, P.J.; Pacheco, F.A. Undamming the Douro River Catchment: A Stepwise Approach for Prioritizing Dam Removal. Water 2019, 11, 693. [Google Scholar] [CrossRef]
- Terêncio, D.P.S.; Pacheco, F.A.L.; Fernandes, L.S.; Cortes, R.M.V. Is It Safe to Remove a Dam at the Risk of a Sprawl by Exotic Fish Species? Sci. Total Environ. 2021, 144768. [Google Scholar] [CrossRef] [PubMed]
- Ordeix, M.; González, G.; Sanz-Ronda, F.J.; Santos, J.M. Restoring Fish Migration in the Rivers of the Iberian Peninsula. Sea Source 2018, 2, 174–179. [Google Scholar]
- Kelly-Richards, S.; Silber-Coats, N.; Crootof, A.; Tecklin, D.; Bauer, C. Governing the Transition to Renewable Energy: A Review of Impacts and Policy Issues in the Small Hydropower Boom. Energy Policy 2017, 101, 251–264. [Google Scholar] [CrossRef]
- Couto, T.B.; Olden, J.D. Global Proliferation of Small Hydropower Plants–Science and Policy. Front. Ecol. Environ. 2018, 16, 91–100. [Google Scholar] [CrossRef]
- Larinier, M.; Marmulla, G. Fish Passes: Types, Principles and Geographical Distribution-an Overview. In Proceedings of the Second International Symposium on the Management of Large Rivers for Fisheries; RAP Publication: Bangkok, Thailand, 2004; Volume 2, pp. 183–206. [Google Scholar]
- Katopodis, C.; Williams, J.G. The Development of Fish Passage Research in a Historical Context. Ecol. Eng. 2012, 48, 8–18. [Google Scholar] [CrossRef]
- Amaral, S.D.; Branco, P.; Katopodis, C.; Ferreira, M.T.; Pinheiro, A.N.; Santos, J.M. Passage Performance of Potamodromous Cyprinids over an Experimental Low-Head Ramped Weir: The Effect of Ramp Length and Slope. Sustainability 2019, 11, 1456. [Google Scholar] [CrossRef]
- Santos, J.M.; Branco, P.J.; Silva, A.T.; Katopodis, C.; Pinheiro, A.N.; Viseu, T.; Ferreira, M.T. Effect of Two Flow Regimes on the Upstream Movements of the Iberian Barbel (Luciobarbus Bocagei) in an Experimental Pool-Type Fishway. J. Appl. Ichthyol. 2013, 29, 425–430. [Google Scholar] [CrossRef]
- Rahel, F.J.; McLaughlin, R.L. Selective Fragmentation and the Management of Fish Movement across Anthropogenic Barriers. Ecol. Appl. 2018, 28, 2066–2081. [Google Scholar] [CrossRef] [PubMed]
- Jesus, J.; Amorim, M.C.P.; Fonseca, P.J.; Teixeira, A.; Natário, S.; Carrola, J.; Varandas, S.; Pereira, L.T.; Cortes, R.M.V. Acoustic Barriers as an Acoustic Deterrent for Native Potamodromous Migratory Fish Species. J. Fish Biol. 2019, 95, 247–255. [Google Scholar] [CrossRef]
- Sand, O.; ENGER, P.; Karlsen, H.E.; Knudsen, F.R. To Intense Infrasound in Juvenile Salmonids. In Proceedings of the American Fisheries Society Symposium, Baltimore, MD, USA, 20–23 May 2001. [Google Scholar]
- Maes, J.; Turnpenny, A.W.H.; Lambert, D.R.; Nedwell, J.R.; Parmentier, A.; Ollevier, F. Field Evaluation of a Sound System to Reduce Estuarine Fish Intake Rates at a Power Plant Cooling Water Inlet. J. Fish Biol. 2004, 64, 938–946. [Google Scholar] [CrossRef]
- Hamel, M.J.; Brown, M.L.; Chipps, S.R. Behavioral Responses of Rainbow Smelt to in Situ Strobe Lights. N. Am. J. Fish. Manag. 2008, 28, 394–401. [Google Scholar] [CrossRef]
- Kim, J.; Mandrak, N.E. Effects of Strobe Lights on the Behaviour of Freshwater Fishes. Environ. Biol. Fishes 2017, 100, 1427–1434. [Google Scholar] [CrossRef]
- Maiolie, M.A.; Harryman, B.; Ament, B. Response of Free-Ranging Kokanee to Strobe Lights. In Proceedings of the Behavioral Technologies for Fish Guidance: American Fisheries Society Symposium, Baltimore, MD, USA, 20–23 May 2001. [Google Scholar]
- Königson, S.; Fjälling, A.; Lunneryd, S.-G. Reactions in Individual Fish to Strobe Light. Field and Aquarium Experiments Performed on Whitefish (Coregonus Lavaretus). Hydrobiologia 2002, 483, 39–44. [Google Scholar] [CrossRef]
- SpaRkS, R.E.; Barkley, T.L.; Creque, S.M.; Dettmers, J.M.; Stainbrook, K.M. Evaluation of an Electric Fish Dispersal Barrier in the Chicago Sanitary and Ship Canal. In Proceedings of the American Fisheries Society Symposium, Pittsburgh, PA, USA, 9–16 September 2010. [Google Scholar]
- Clarkson, R.W. Effectiveness of Electrical Fish Barriers Associated with the Central Arizona Project. N. Am. J. Fish. Manag. 2004, 24, 94–105. [Google Scholar] [CrossRef]
- Kates, D.; Dennis, C.; Noatch, M.R.; Suski, C.D. Responses of Native and Invasive Fishes to Carbon Dioxide: Potential for a Nonphysical Barrier to Fish Dispersal. Can. J. Fish. Aquat. Sci. 2012, 69, 1748–1759. [Google Scholar] [CrossRef]
- da Silva, L.G.M. Estudo de Sistemas Para Repulsão de Peixes Como Alternativas de Mitigação de Impacto Ambiental Em Usinas Hidrelétricas e Canais Para Abastecimento de Água Tese de Doutoramento; UFMG: Belo Horizonte, Brasil, 2010. [Google Scholar]
- Sager, D.R.; Hocutt, C.H.; Stauffer Jr, J.R. Estuarine Fish Responses to Strobe Light, Bubble Curtains and Strobe Light/Bubble-Curtain Combinations as Influenced by Water Flow Rate and Flash Frequencies. Fish. Res. 1987, 5, 383–399. [Google Scholar] [CrossRef]
- Johnson, R.L.; Simmons, M.A.; McKinstry, C.A.; Simmons, C.S.; Cook, C.B.; Brown, R.S.; Tano, D.K.; Thorsten, S.L.; Faber, D.M.; Lecaire, R. Strobe Light Deterrent Efficacy Test and Fish Behavior Determination at Grand Coulee Dam Third Powerplant Forebay; Pacific Northwest National Lab. (PNNL): Richland, WA, USA, 2005. [Google Scholar]
- Popper, A.N.; Schilt, C.R. Hearing and acoustic behavior: Basic and applied considerations. In Fish Bioacoustics; Springer: Berlin, Germany, 2008; pp. 17–48. [Google Scholar]
- Orbach, M.K. Hunters, Seamen, and Entrepreneurs; University of California Press: San Diego, CA, USA, 1977. [Google Scholar]
- Finneran, J.J.; Oliver, C.W.; Schaefer, K.M.; Ridgway, S.H. Source Levels and Estimated Yellowfin Tuna (Thunnus Albacares) Detection Ranges for Dolphin Jaw Pops, Breaches, and Tail Slaps. J. Acoust. Soc. Am. 2000, 107, 649–656. [Google Scholar] [CrossRef]
- Mitson, R.B. Underwater Noise of Research Vessels: Review and Recommendations; ICES Cooperative Research Report No. 209; ICES: Columbia, British, 1995; p. 61. ISSN 27077144. [Google Scholar]
- Broadhurst, M.K.; Kennelly, S.J.; Eayrs, S. Flow-Related Effects in Prawn-Trawl Codends: Potential for Increasing the Escape of Unwanted Fish through Square-Mesh Panels. Fish. Bull. 1999, 97, 1–8. [Google Scholar]
- Noatch, M.R.; Suski, C.D. Non-Physical Barriers to Deter Fish Movements. Environ. Rev. 2012, 20, 71–82. [Google Scholar] [CrossRef]
- Murchy, K.A.; Cupp, A.R.; Amberg, J.J.; Vetter, B.J.; Fredricks, K.T.; Gaikowski, M.P.; Mensinger, A.F. Potential Implications of Acoustic Stimuli as a Non-Physical Barrier to Silver Carp and Bighead Carp. Fish. Manag. Ecol. 2017, 24, 208–216. [Google Scholar] [CrossRef]
- Perry, R.W.; Romine, J.G.; Adams, N.S.; Blake, A.R.; Burau, J.R.; Johnston, S.V.; Liedtke, T.L. Using a Non-Physical Behavioural Barrier to Alter Migration Routing of Juvenile Chinook Salmon in the Sacramento-San Joaquin River Delta: Non-Physical Barrier for Routing of Juvenile Salmon. River Res. Appl. 2014, 30, 192–203. [Google Scholar] [CrossRef]
- Wamboldt, J.J.; Murchy, K.A.; Stanton, J.C.; Blodgett, K.D.; Brey, M.K. Evaluation of an Acoustic Fish Deterrent System in Shallow Water Application at the Emiquon Preserve, Lewistown, IL. Manag. Biol. Invasions 2019, 10, 536. [Google Scholar] [CrossRef]
- Zielinski, D.P.; Voller, V.R.; Svendsen, J.C.; Hondzo, M.; Mensinger, A.F.; Sorensen, P. Laboratory Experiments Demonstrate That Bubble Curtains Can Effectively Inhibit Movement of Common Carp. Ecol. Eng. 2014, 67, 95–103. [Google Scholar] [CrossRef]
- Qin, X.; Liu, Y.; Shen, X.; Wu, Y.; Tian, W.; Liu, Y.; Wang, X.; Shi, X.; Liu, G. Spatial avoidance of tu-fish Schizopygopsis younghusbandi for different sounds may inform behavioural deterrence strategies. Fish Manag. Ecol. 2019, 27, 10–19. [Google Scholar] [CrossRef]
- Nissen, A.C.; Vetter, B.J.; Rogers, L.S.; Mensinger, A.F. Impacts of Broadband Sound on Silver (Hypophthalmichthys Molitrix) and Bighead (H. Nobilis) Carp Hearing Thresholds Determined Using Auditory Evoked Potential Audiometry. Fish Physiol. Biochem. 2019, 45, 1683–1695. [Google Scholar] [CrossRef]
- Liu, G.; Wu, Y.; Shen, X.; Hu, Y.; Qin, X.; Tian, W.; Liu, L.; Wang, X.; Shi, X. Laboratory Experiments Demonstrate That the Hissing of the Chinese Alligator Can Effectively Inhibit Movement of Flower Fish Ptychobarbus Kaznakovi. Hydrobiologia 2019, 836, 97–108. [Google Scholar] [CrossRef]
- Vetter, B.J.; Murchy, K.A.; Cupp, A.R.; Amberg, J.J.; Gaikowski, M.P.; Mensinger, A.F. Acoustic Deterrence of Bighead Carp (Hypophthalmichthys Nobilis) to a Broadband Sound Stimulus. J. Gt. Lakes Res. 2017, 43, 163–171. [Google Scholar] [CrossRef]
- Vetter, B.J.; Cupp, A.R.; Fredricks, K.T.; Gaikowski, M.P.; Mensinger, A.F. Acoustical Deterrence of Silver Carp (Hypophthalmichthys Molitrix). Biol. Invasions 2015, 17, 3383–3392. [Google Scholar] [CrossRef]
- Putland, R.L.; Mensinger, A.F. Acoustic Deterrents to Manage Fish Populations. Rev. Fish Biol. Fish. 2019, 29, 789–807. [Google Scholar] [CrossRef]
- Vetter, B.J.; Brey, M.K.; Mensinger, A.F. Reexamining the Frequency Range of Hearing in Silver (Hypophthalmichthys Molitrix) and Bighead (H. Nobilis) Carp. PLoS ONE 2018, 13, e0192561. [Google Scholar] [CrossRef]
- Zielinski, D.P.; Hondzo, M.; Voller, V.R. Mathematical Evaluation of Behavioral Deterrent Systems to Disrupt Fish Movement. Ecol. Model. 2014, 272, 150–159. [Google Scholar] [CrossRef]
- Bzonek, P.A.; Kim, J.; Mandrak, N.E. Short-Term Behavioural Response of Common Carp, Cyprinus Carpio, to Acoustic and Stroboscopic Stimuli. Manag. Biol. Invasions 2020, 11, 279–292. [Google Scholar] [CrossRef]
- Zielinski, D.P.; Sorensen, P.W. Silver, Bighead, and Common Carp Orient to Acoustic Particle Motion When Avoiding a Complex Sound. PLoS ONE 2017, 12, e0180110. [Google Scholar] [CrossRef] [PubMed]
- Flammang, M.K.; Weber, M.J.; Thul, M.D. Laboratory Evaluation of a Bioacoustic Bubble Strobe Light Barrier for Reducing Walleye Escapement. North Am. J. Fish. Manag. 2014, 34, 1047–1054. [Google Scholar] [CrossRef]
- Ruebush, B.C.; Sass, G.G.; Chick, J.H.; Stafford, J.D. In-Situ Tests of Sound-Bubble-Strobe Light Barrier Technologies to Prevent Range Expansions of Asian Carp. Aquat. Invasions 2012, 7, 37–48. [Google Scholar] [CrossRef]
- Miehls, S.M.; Johnson, N.S.; Hrodey, P.J. Test of a Nonphysical Barrier Consisting of Light, Sound, and Bubble Screen to Block Upstream Movement of Sea Lampreys in an Experimental Raceway. N. Am. J. Fish. Manag. 2017, 37, 660–666. [Google Scholar] [CrossRef]
- Jesus, J.; Teixeira, A.; Natário, S.; Cortes, R. Repulsive Effect of Stroboscopic Light Barriers on Native Salmonid (Salmo Trutta) and Cyprinid (Pseudochondrostoma Duriense and Luciobarbus Bocagei) Species of Iberia. Sustainability 2019, 11, 1332. [Google Scholar] [CrossRef]
- Stewart, H.A.; Wolter, M.H.; Wahl, D.H. Laboratory Investigations on the Use of Strobe Lights and Bubble Curtains to Deter Dam Escapes of Age-0 Muskellunge. N. Am. J. Fish. Manag. 2014, 34, 571–575. [Google Scholar] [CrossRef]
- Vetter, B.J.; Rogers, L.S.; Mensinger, A.F. The Effect of Light Stimuli on Dark-Adapted Visual Sensitivity in Invasive Silver Carp Hypophthalmichthys Molitrix and Bighead Carp H. Nobilis. J. Fish Biol. 2019, 95, 256–262. [Google Scholar] [CrossRef]
- Kim, J.; Bondy, C.; Chandler, C.M.; Mandrak, N.E. Behavioural Response of Juvenile Common Carp (Cyprinus Carpio) and Juvenile Channel Catfish (Ictalurus Punctatus) to Strobe Light. Fishes 2019, 4, 29. [Google Scholar] [CrossRef]
- Lin, C.; Dai, H.; Shi, X.; Deng, Z.D.; Mao, J.; Zhao, S.; Luo, J.; Tan, J. An Experimental Study on Fish Attraction Using a Fish Barge Model. Fish. Res. 2019, 210, 181–188. [Google Scholar] [CrossRef]
- Elvidge, C.K.; Reid, C.H.; Ford, M.I.; Sills, M.; Patrick, P.H.; Gibson, D.; Backhouse, S.; Cooke, S.J. Ontogeny of Light Avoidance in Juvenile Lake Sturgeon. J. Appl. Ichthyol. 2019, 35, 202–209. [Google Scholar] [CrossRef]
- Hansen, M.J.; Cocherell, D.E.; Cooke, S.J.; Patrick, P.H.; Sills, M.; Fangue, N.A. Behavioural Guidance of Chinook Salmon Smolts: The Variable Effects of LED Spectral Wavelength and Strobing Frequency. Conserv. Physiol. 2018, 6, coy032. [Google Scholar] [CrossRef]
- Vowles, A.S.; Kemp, P.S. Effects of Light on the Behaviour of Brown Trout (Salmo Trutta) Encountering Accelerating Flow: Application to Downstream Fish Passage. Ecol. Eng. 2012, 47, 247–253. [Google Scholar] [CrossRef]
- Brown, R. The Potential of Strobe Lighting as a Cost-Effective Means for Reducing Impingement and Entrainment. Environ. Sci. Policy 2000, 3, 405–416. [Google Scholar] [CrossRef]
- Taft, E.P.; Dixon, D.A.; Sullivan, C.W. Electric Power Research Institute’s (EPRI) Research on Behavioral Technologies. In Proceedings of the Behavioral Technologies for Fish Guidance: American Fisheries Society Symposium, Baltimore, MD, USA, 20–23 May 2001. [Google Scholar]
- Fay, R.R.; Popper, A.N. Structure and function in teleost auditory systems. In Comparative Studies of Hearing in Vertebrates; Springer: Berlin, Germany, 1980; pp. 3–42. [Google Scholar]
- Ladich, F.; Fay, R.R. Auditory Evoked Potential Audiometry in Fish. Rev. Fish Biol. Fish. 2013, 23, 317–364. [Google Scholar] [CrossRef]
- Hawkins, A.D.; Roberts, L.; Cheesman, S. Responses of Free-Living Coastal Pelagic Fish to Impulsive Sounds. J. Acoust. Soc. Am. 2014, 135, 3101–3116. [Google Scholar] [CrossRef]
- Neo, Y.Y.; Hubert, J.; Bolle, L.; Winter, H.V.; Ten Cate, C.; Slabbekoorn, H. Sound Exposure Changes European Seabass Behaviour in a Large Outdoor Floating Pen: Effects of Temporal Structure and a Ramp-up Procedure. Environ. Pollut. 2016, 214, 26–34. [Google Scholar] [CrossRef]
- Slabbekoorn, H. Aiming for progress in understanding underwater noise impact on fish: Complementary need for indoor and outdoor studies. In The Effects of Noise on Aquatic Life II; Springer: Berlin, Germany, 2016; pp. 1057–1065. [Google Scholar]
- Akamatsu, T.; Okumura, T.; Novarini, N.; Yan, H.Y. Empirical Refinements Applicable to the Recording of Fish Sounds in Small Tanks. J. Acoust. Soc. Am. 2002, 112, 3073–3082. [Google Scholar] [CrossRef]
- Wysocki, L.E.; Montey, K.; Popper, A.N. The Influence of Ambient Temperature and Thermal Acclimation on Hearing in a Eurythermal and a Stenothermal Otophysan Fish. J. Exp. Biol. 2009, 212, 3091–3099. [Google Scholar] [CrossRef]
- Kastelein, R.A.; Van Der Heul, S.; Verboom, W.C.; Jennings, N.; Van Der Veen, J.; de Haan, D. Startle Response of Captive North Sea Fish Species to Underwater Tones between 0.1 and 64 KHz. Mar. Environ. Res. 2008, 65, 369–377. [Google Scholar] [CrossRef]
- Pottinger, T.G. A Multivariate Comparison of the Stress Response in Three Salmonid and Three Cyprinid Species: Evidence for Inter-Family Differences. J. Fish Biol. 2010, 76, 601–621. [Google Scholar] [CrossRef]
- Douglas, R.H.; Hawryshyn, C.W. Behavioural studies of fish vision: An analysis of visual capabilities. In The Visual System of Fish; Springer: Berlin, Germany, 1990; pp. 373–418. [Google Scholar]
- Li, L.; Maaswinkel, H. Visual Sensitivity and Signal Processing in Teleosts. Fish Physiol. 2006, 25, 179–241. [Google Scholar]
- Stenkamp, D.L. The Rod Photoreceptor Lineage of Teleost Fish. Prog. Retin. Eye Res. 2011, 30, 395–404. [Google Scholar] [CrossRef]
- Sullivan, B.G.; Wilson, A.D.; Gutowsky, L.F.; Patrick, P.H.; Sills, M.; Cooke, S.J. The Behavioral Responses of a Warmwater Teleost to Different Spectra of Light-Emitting Diodes. N. Am. J. Fish. Manag. 2016, 36, 1000–1005. [Google Scholar] [CrossRef]
- Banks, M.S.; Sprague, W.W.; Schmoll, J.; Parnell, J.A.; Love, G.D. Why Do Animal Eyes Have Pupils of Different Shapes? Sci. Adv. 2015, 1, e1500391. [Google Scholar] [CrossRef] [PubMed]
- Brischoux, F.; Pizzatto, L.; Shine, R. Insights into the Adaptive Significance of Vertical Pupil Shape in Snakes. J. Evol. Biol. 2010, 23, 1878–1885. [Google Scholar] [CrossRef] [PubMed]
- Land, M.F.; Fernald, R.D. The Evolution of Eyes. Annu. Rev. Neurosci. 1992, 15, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Schwassmann, H.O.; Kruger, L. Experimental Analysis of the Visual System of the Four-Eyed Fish Anableps Microlepis. Vision Res. 1965, 5, 269-IN1. [Google Scholar] [CrossRef]
- Vinterstare, J.; Hulthen, K.; Nilsson, D.E.; Nilsson, A.; Brönmark, C. More than Meets the Eye: Predator-induced Pupil Size Plasticity in a Teleost Fish. J. Anim. Ecol. 2020, 89, 2258–2267. [Google Scholar] [CrossRef]
- Ruetz III, C.R.; Hurford, A.L.; Vondracek, B. Interspecific Interactions between Brown Trout and Slimy Sculpin in Stream Enclosures. Trans. Am. Fish. Soc. 2003, 132, 611–618. [Google Scholar] [CrossRef]
- McIntosh, A.R.; Townsend, C.R. Contrasting Predation Risks Presented by Introduced Brown Trout and Native Common River Galaxias in New Zealand Streams. Can. J. Fish. Aquat. Sci. 1995, 52, 1821–1833. [Google Scholar] [CrossRef]
- Bachman, R.A.; Reynolds, W.W.; Casterlin, M.E. Diel Locomotor Activity Patterns of Wild Brown Trout (Salmo Trutta L.) in an Electronic Shuttlebox. Hydrobiologia 1979, 66, 45–47. [Google Scholar] [CrossRef]
- Gregory, R.S.; Levings, C.D. Turbidity Reduces Predation on Migrating Juvenile Pacific Salmon. Trans. Am. Fish. Soc. 1998, 127, 275–285. [Google Scholar] [CrossRef]
- Lehtiniemi, M.; Engström-Öst, J.; Viitasalo, M. Turbidity Decreases Anti-Predator Behaviour in Pike Larvae, Esox Lucius. Environ. Biol. Fishes 2005, 73, 1–8. [Google Scholar] [CrossRef]
- de Oliveira Mesquita, F.; Godinho, H.P.; de Azevedo, P.G.; Young, R.J. A Preliminary Study into the Effectiveness of Stroboscopic Light as an Aversive Stimulus for Fish. Appl. Anim. Behav. Sci. 2008, 111, 402–407. [Google Scholar] [CrossRef]
- Taylor, R.M.; Pegg, M.A.; Chick, J.H. Response of Bighead Carp to a Bioacoustic Behavioural Fish Guidance System. Fish. Manag. Ecol. 2005, 12, 283–286. [Google Scholar] [CrossRef]
- Brevik, I.; Kristiansen, Ø. The Flow in and around Air-Bubble Plumes. Int. J. Multiph. Flow 2002, 28, 617–634. [Google Scholar] [CrossRef]
- Soga, C.L.; Rehmann, C.R. Dissipation of Turbulent Kinetic Energy near a Bubble Plume. J. Hydraul. Eng. 2004, 130, 441–449. [Google Scholar] [CrossRef]
- Leighton, T.G.; Walton, A.J. An Experimental Study of the Sound Emitted from Gas Bubbles in a Liquid. Eur. J. Phys. 1987, 8, 98. [Google Scholar] [CrossRef]
- Leighton, T.G. Acoustic Bubble Detection-I. The Detection of Stable Gas Bodies. Environ. Eng. 1994, 7, 9–16. [Google Scholar]
- Lin, J.N.; Banerji, S.K.; Yasuda, H. Role of Interfacial Tension in the Formation and the Detachment of Air Bubbles. 1. A Single Hole on a Horizontal Plane Immersed in Water. Langmuir 1994, 10, 936–942. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).