
Distribution of the Macrophyte Communities in the Danube Reflects River Serial Discontinuity

 , ,
, , 
 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Macrophyte Survey
2.3. Assessment of Habitat Parameters
2.4. Data Treatment
2.5. Similarity of Macrophyte Communities
2.6. The Influence of Environmental Factors on the Composition of the Macrophyte Community
3. Results
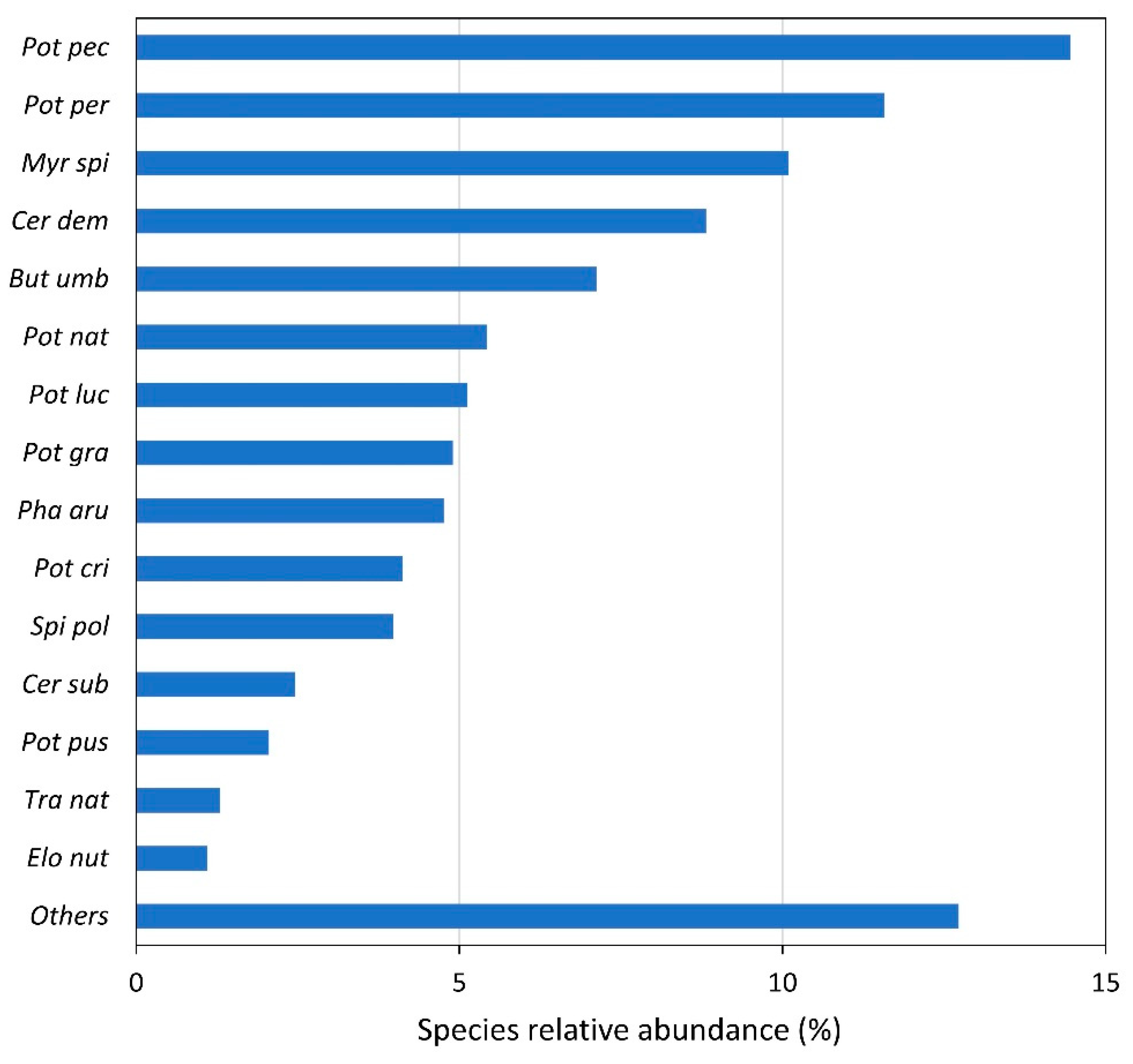
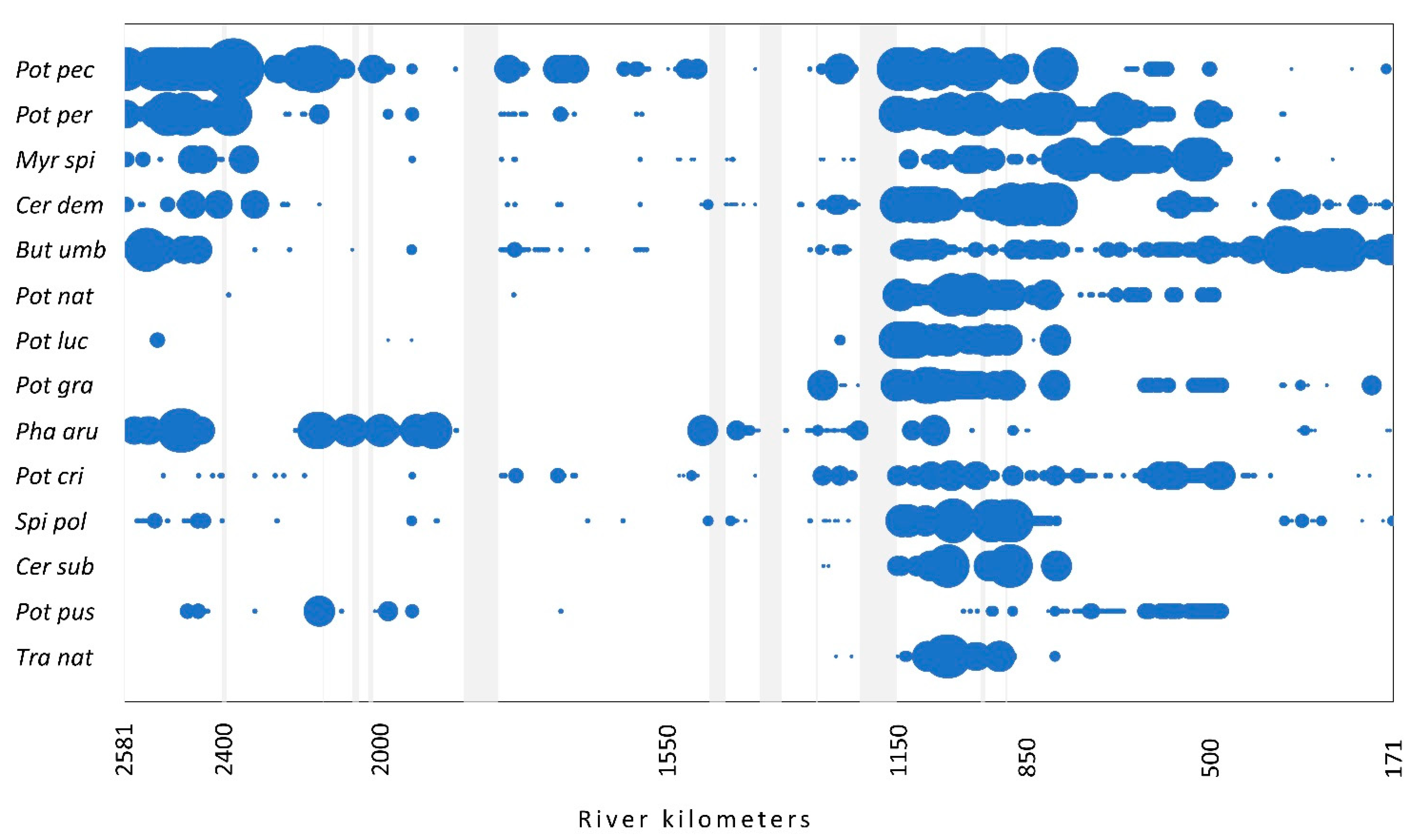
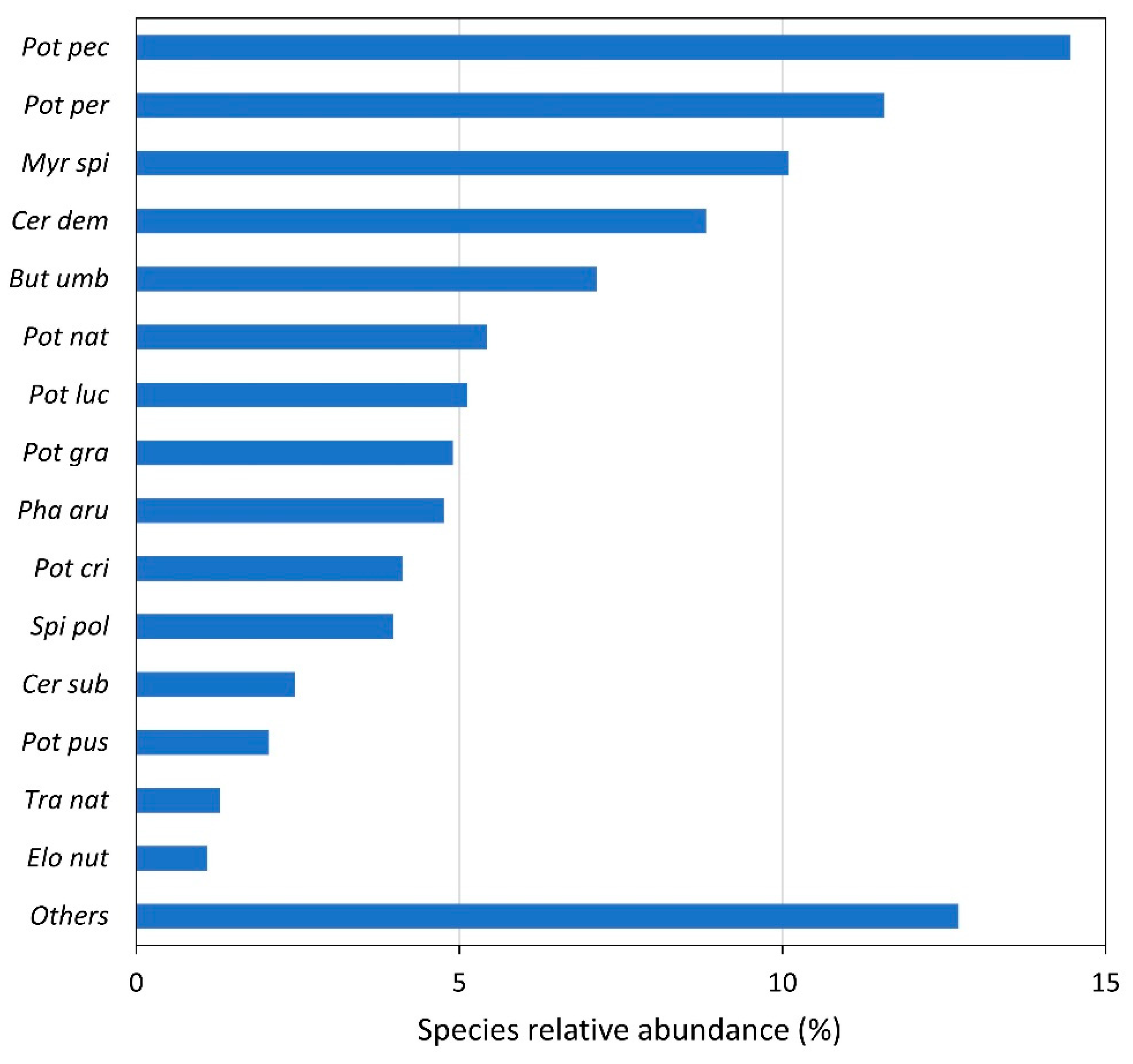
3.1. Species Presence and Abundance
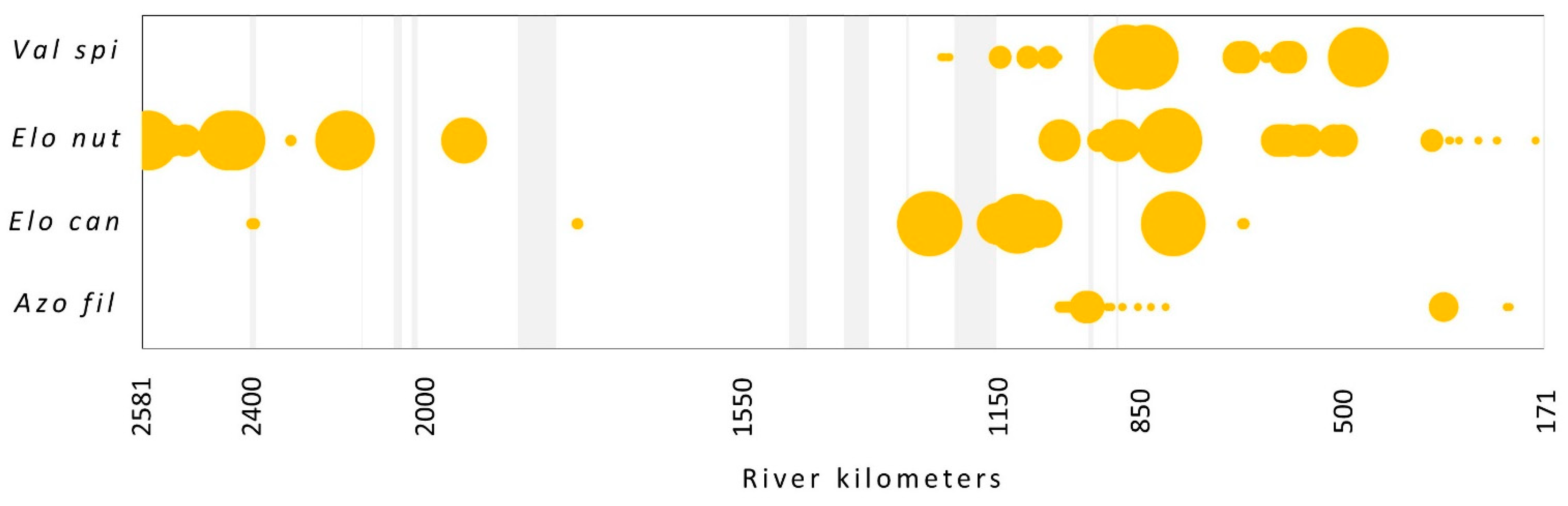
3.2. Neophyte Species Presence and Abundance
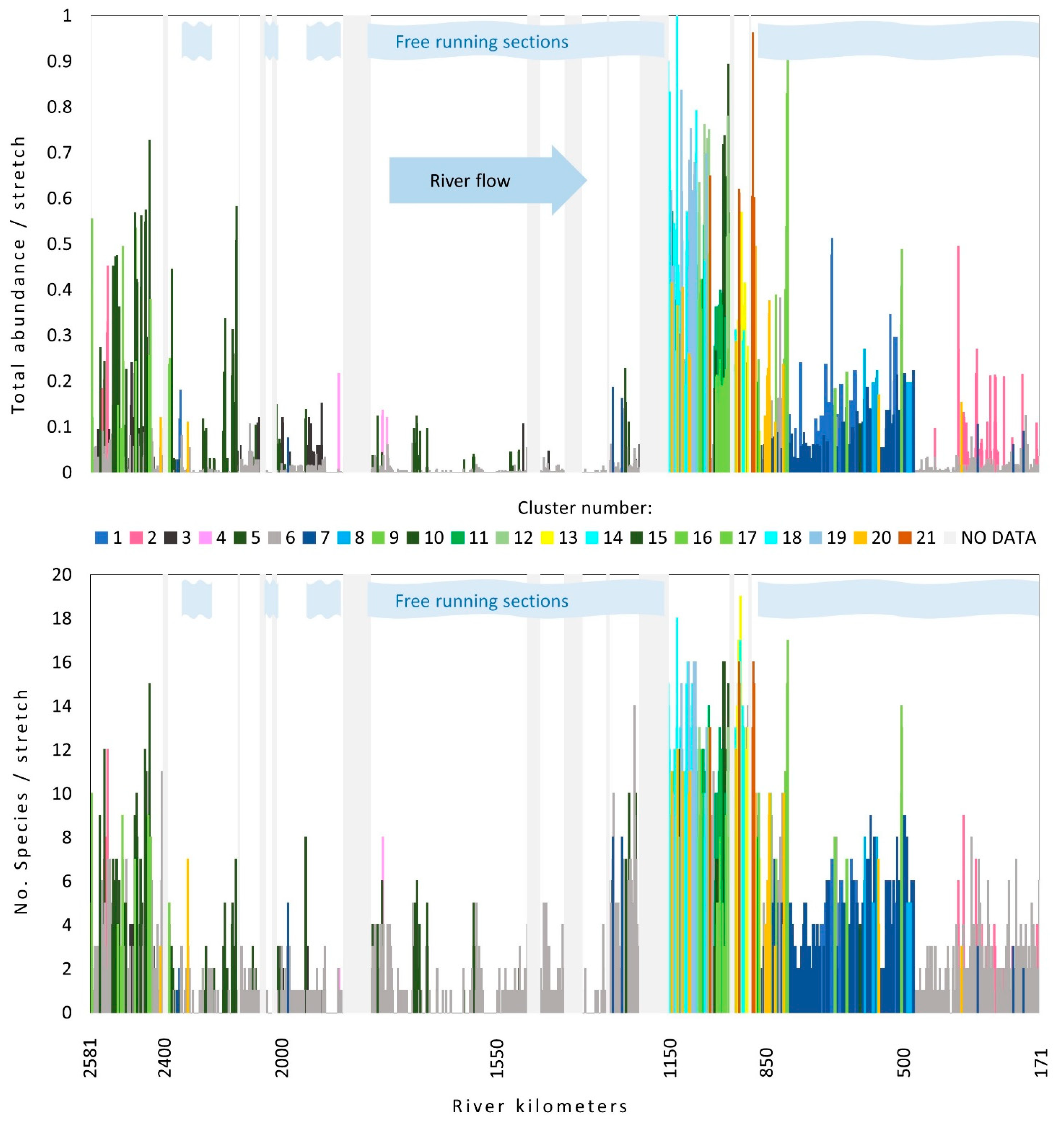
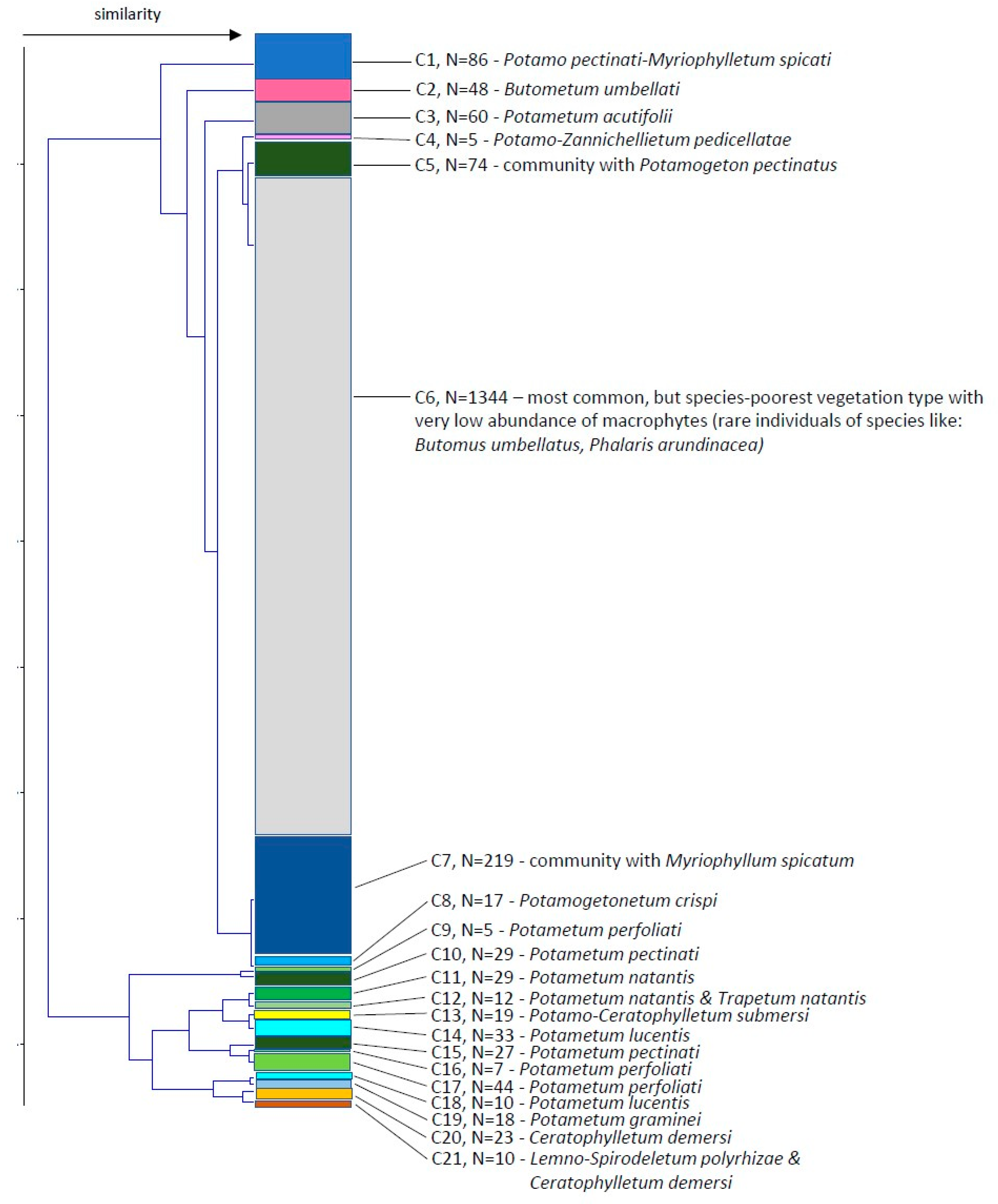
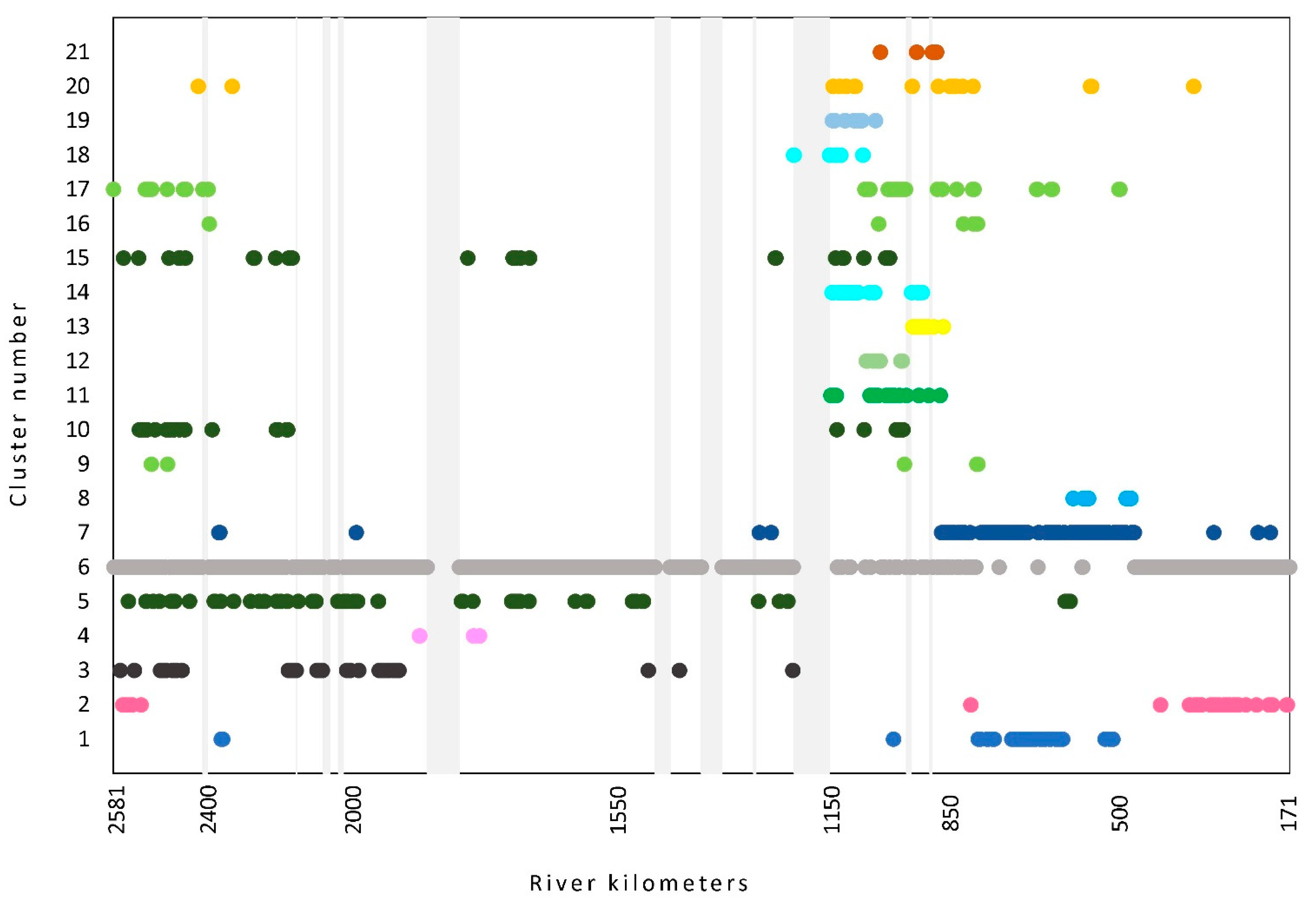
3.3. Plant Communities along the Course
3.3.1. Order Potamogetonetalia Koch 1926
- Alliance Potamogetonion Libbert 1931
Association Potamo pectinati-Myriophylletum spicati Rivas Goday 1964(cluster 1, 86 stretches)
Plant community with Myriophyllum spicatum(cluster 7, 219 stretches)
Association Potametum acutifolii Segal ex Šumberová et Hrivnák in Chytrý 2011(cluster 3, 60 stretches)
Association Parvo-Potamo-Zannichellietum pedicellatae Soó 1947(cluster 4, 5 stretches)
Association Potametum crispi Soo 1927(cluster 8, 17 stretches)
Association Potametum pectinati Carstensen ex Hilbig 1971(cluster 10, 29 stretches; cluster 15, 27 stretches)
Plant community with Potamogeton pectinatus(cluster 5, 74 stretches)
Association Potametum perfoliati Miljan 1933(cluster 9, 5 stretches; cluster 16, 7 stretches; cluster 17, 44 stretches)
Association Potametum natantis Hild 1959(cluster 11, 29 stretches; cluster 12, 4 of 12 stretches)
Association Potametum lucentis Hueck 1931(cluster 14, 33 stretches; cluster 18, 10 stretches)
Association Potametum graminei Lang 1967(cluster 19, 18 stretches)
Species-poorest vegetation type in sections with the lowest abundance of macrophytes.(cluster 6, 1345 stretches)
- Alliance Nymphaeion albae Oberdorfer 1957
Association Trapetum natantis Kárpáti 1963(cluster 12, 8 of 12 stretches)
3.3.2. Order Lemnetalia minoris O. de Bolòs et Masclans 1955
- Alliance Stratiotion Den Hartog et Segal 1964
Association Potamo-Ceratophylletum submersi Pop 1962(cluster 13, 19 stretches)
Association Ceratophylletum demersi Corillion 1957(cluster 20, 23 stretches)
- Alliance Lemnion minoris O. de Bolòs et Masclans 1955
Association Lemno-Spirodeletum polyrhizae Koch 1954(cluster 21, 8 of 10 stretches)
3.3.3. Order Phragmitetalia
Association Butometum umbellati Philippi 1973(cluster 2, 48 stretches)
3.4. The Influence of Environmental Factors on the Composition of Macrophyte Communities
4. Discussion
4.1. Species Distribution and Abundance
4.2. Neophytes in the Danube Main Channel
4.3. The Distribution of Macrophyte Communities along the Course
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sommerwerk, N.; Hein, T.; Schneider-Jacoby, M.; Baumgartner, C.; Ostojić, A.; Siber, R.; Bloesch, J.; Paunović, M.; Tockner, K. The Danube River Basin. In Rivers of Europe; Elsevier: Amsterdam, The Netherlands, 2009; pp. 59–112. [Google Scholar]
- Lenhardt, M.; Smederevac-Lalić, M.; Hegediš, A.; Skorić, S.; Cvijanović, G.; Višnjić-Jeftić, Ž.; Djikanović, V.; Jovičić, K.; Jaćimović, M.; Jarić, I. Human Impacts on Fish Fauna in the Danube River in Serbia: Current Status and Ecological Implications. In Geobotany Studies; Springer: Berlin/Heidelberg, Germany, 2020; pp. 257–279. [Google Scholar]
- Haslam, S.M. The Historic River: Rovers and Cultures Down the Ages; Cobden of Cambridge Press: Cambridge, UK, 1999; p. 324. [Google Scholar]
- Winiwarter, V.; Schmid, M.; Hohensinner, S.; Haidvogl, G. The Environmental History of the Danube River Basin as an Issue of Long-Term Socio-ecological Research. In Long Term Socio-Ecological Research; Springer: Dordrecht, The Netherlands, 2013; pp. 103–122. [Google Scholar]
- Hynes, H.B.N. The Ecology of Running Waters; University of Toronto Press: Toronto, ON, Canada, 1970; p. 555. [Google Scholar]
- Odum, H.T. Trophic Structure and Productivity of Silver Springs, Florida. Ecol. Monogr. 1957, 27, 55–112. [Google Scholar] [CrossRef]
- Illies, J.; Botosaneanu, L. Problèmes et méthodes de la classification et de la zonation écologique des eaux courantes, considerées surtout du point de vue faunistique. Int. Ver. Theor. Angew. Limnol. Mitt. 1963, 12, 1–57. [Google Scholar] [CrossRef]
- Hawkes, H.A. River zonation and classification. In River Ecology; Whitton, B.A., Ed.; Blackwell Scientific Publishing: Oxford, UK, 1975; pp. 312–374. [Google Scholar]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The River Continuum Concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Ward, J.V.; Stanford, J. The serial discontinuity concept of lotic ecosystems. In Dynamics of Lotic Ecosystems; Fontane, T.D., Bartell, S.M., Eds.; Ann Arbor Scientific Publications: Ann Arbor, MI, USA, 1983; pp. 29–42. [Google Scholar]
- Ward, J.V.; Stanford, J.A. The serial discontinuity concept: Extending the model to floodplain rivers. Regul. Rivers Res. Manag. 1995, 10, 159–168. [Google Scholar] [CrossRef]
- Bornette, G.; Amoros, C.; Lamouroux, N. Aquatic plant diversity in riverine wetlands: The role of connectivity. Freshw. Biol. 1998, 39, 267–283. [Google Scholar] [CrossRef]
- Moog, O.; Sommerhäuser, M.; Robert, S.; Battisti, T.; Birk, S.; Hering, D.; Ofenböck, T.; Schmedtje, U.; Schmidt-Kloiber, A.; Vogel, B. Typology of the Danube River based on “top-down” and “bottom-up” approaches. In Proceedings of the Proceedings 36th International Conference of IAD. Austrian Committee DanubeResearch/IAD, Vienna, Austria, 4–8 September 2006; pp. 260–264. [Google Scholar]
- Laszloffy, W. Die Hydrographie der Donau. Der Fluss als Lebensraum. In Limnologie der Donau—Eine Monographische Darstellung, Kapitel II; Liepolt, R., Ed.; Schweizerbart: Stuttgart, Germany, 1965; pp. 16–57. [Google Scholar]
- Illies, J. Limnofauna Europaea: Eine Zusammenstellung aller die Europäischen Binnengewässer Bewohnenden Mehrzelligen Tierarten mit Angaben über ihre Verbreitung und Ökologie; Fischer: Stuttgart, Germany, 1978. [Google Scholar]
- Olson, K.R.; Krug, E. The Danube, an Empire Boundary River: Settlements, Invasions, Navigation, and Trade Pathway. J. Water Resour. Prot. 2020, 12, 884–897. [Google Scholar] [CrossRef]
- Stoica, C.; Camejo, J.; Banciu, A.; Nita-Lazar, M.; Paun, I.; Cristofor, S.; Pacheco, O.R.; Guevara, M. Water quality of Danube Delta systems: Ecological status and prediction using machine-learning algorithms. Water Sci. Technol. 2016, 73, 2413–2421. [Google Scholar] [CrossRef]
- Takić, L.; Mladenović-Ranisavljević, I.; Vasović, D.; Đorđević, L. The Assessment of the Danube River Water Pollution in Serbia. Water Air Soil Pollut. 2017, 228, 380. [Google Scholar] [CrossRef]
- Hein, T.; Funk, A.; Pletterbauer, F.; Graf, W.; Zsuffa, I.; Haidvogl, G.; Schinegger, R.; Weigelhofer, G. Management challenges related to long-term ecological impacts, complex stressor interactions, and different assessment approaches in the Danube River Basin. River Res. Appl. 2019, 35, 500–509. [Google Scholar] [CrossRef]
- Janauer, G.A.; Gaberščik, A.; Květ, J.; Germ, M.; Exler, N. Macrophytes of the River Danube Basin; Academia: Praha, Czech Republic, 2018; p. 408. ISBN 978-80-200-2743-6. [Google Scholar]
- Wilzbach, M.A.; Cummins, K.W. Rivers and Streams: Physical Setting and Adapted Biota. In Encyclopedia of Ecology; Elsevier: Amsterdam, The Netherlands, 2019; pp. 594–606. [Google Scholar]
- Neuhold, B.; Janauer, J.D.; Janauer, G.A. Within-weed bed architectural adaptation of branching pattern in Myriophyllum spicatum L. Acta Biol. Slov. 2018, 61, 55–63. [Google Scholar]
- Breugnot, E.; Dutartre, A.; Laplace-Treyture, C.; Haury, J. Local distribution of macrophytes and consequences for sampling methods in large rivers. Hydrobiologia 2008, 610, 13–23. [Google Scholar] [CrossRef]
- Baattrup-Pedersen, A.; Riis, T. Macrophyte diversity and composition in relation to substratum characteristics in regulated and unregulated Danish streams. Freshw. Biol. 1999, 42, 375–385. [Google Scholar] [CrossRef]
- Clarke, S.J. Vegetation growth in rivers: Influences upon sediment and nutrient dynamics. Prog. Phys. Geogr. Earth Environ. 2002, 26, 159–172. [Google Scholar] [CrossRef]
- Chambers, P.A.; Lacoul, P.; Murphy, K.J.; Thomaz, S.M. Global diversity of aquatic macrophytes in freshwater. Hydrobiologia 2008, 595, 9–26. [Google Scholar] [CrossRef]
- Preiner, S.; Dai, Y.; Pucher, M.; Reitsema, R.E.; Schoelynck, J.; Meire, P.; Hein, T. Effects of macrophytes on ecosystem metabolism and net nutrient uptake in a groundwater fed lowland river. Sci. Total Environ. 2020, 721, 137620. [Google Scholar] [CrossRef]
- Gibbs, H.; Gurnell, A.; Heppell, K.; Spencer, K. Macrophytes: Ecosystem engineers in UK urban rivers. Geophys. Res. Abstr. 2012, 14, 490. [Google Scholar]
- Sand-Jensen, K. Macrophytes as biological engineers in the ecology of Danish streams. In Freshwater Biology. Priorities and Development in Danish Research; Gad: København, Denmark, 1997; pp. 74–101. [Google Scholar]
- Rolland, D.C.; Haury, J.; Marmonier, P.; Lagadeuc, Y. Effect of Macrophytes on Flow Conditions and Deposition of Suspended Particles in Small Streams: An Experimental Study Using Artificial Vegetation. Rev. Sci. l’eau/J. Water Sci. 2015, 28, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Mechora, Š.; Germ, M.; Stibilj, V. Selenium compounds in selenium-enriched cabbage. Pure Appl. Chem. 2012, 84, 259–268. [Google Scholar] [CrossRef]
- Rejmankova, E. The role of macrophytes in wetland ecosystems. J. Ecol. Environ. 2011, 34, 333–345. [Google Scholar] [CrossRef] [Green Version]
- Bornette, G.; Amoros, C.; Chessel, D. Effect of allogenic processes on successional rates in former river channels. J. Veg. Sci. 1994, 5, 237–246. [Google Scholar] [CrossRef]
- Šraj-Kržič, N.; Germ, M.; Urbanc-Berčič, O.; Kuhar, U.; Janauer, G.A.; Gaberščik, A. The quality of the aquatic environment and macrophytes of karstic watercourses. Plant Ecol. 2007, 192, 107–118. [Google Scholar] [CrossRef]
- Jusik, S.; Szoszkiewicz, K.; Kupiec, J.M.; Lewin, I.; Samecka-Cymerman, A. Development of comprehensive river typology based on macrophytes in the mountain-lowland gradient of different Central European ecoregions. Hydrobiologia 2015, 745, 241–262. [Google Scholar] [CrossRef] [Green Version]
- Graham, S.E.; O’Brien, J.M.; Burrell, T.K.; McIntosh, A.R. Aquatic macrophytes alter productivity-richness relationships in eutrophic stream food webs. Ecosphere 2015, 6, 1–18. [Google Scholar] [CrossRef]
- Pip, E. Water temperature and freshwater macrophyte distribution. Aquat. Bot. 1989, 34, 367–373. [Google Scholar] [CrossRef]
- Haslam, S.M. River plants of Western Europe: The Macrophytic Vegetation of Watercourses of the European Economic Community; Cambridge University Press: Cambridge, UK, 1987; p. 512. [Google Scholar]
- Haslam, S.M. River Plants: The Macrophytic Vegetation of Watercourses; Forrest Text: Ceredigion, UK, 2006; p. 450. [Google Scholar]
- Kuhar, U.; Germ, M.; Gaberščik, A.; Urbanič, G. Development of a River Macrophyte Index (RMI) for assessing river ecological status. Limnologica 2011, 41, 235–243. [Google Scholar] [CrossRef]
- Mechora, Š.; Germ, M.; Stibilj, V. Monitoring of selenium in macrophytes—The case of Slovenia. Chemosphere 2014, 111, 464–470. [Google Scholar] [CrossRef]
- Kohler, A.; Schneider, S. Macrophytes as bioindicators. River Syst. 2003, 14, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Kuhar, U.; Germ, M.; Gaberščik, A. Macrophytes of watercourses in the Slovenian Danube Basin. In Macrophytes of Watercourses in the Slovenian Danube Basin; Academia: Praha, Czech Republic, 2018; pp. 234–251. [Google Scholar]
- Kuhar, U.; Gregorc, T.; Renčelj, M.; Šraj-Kržič, N.; Gaberščik, A. Distribution of macrophytes and condition of the physical environment of streams flowing through agricultural landscape in north-eastern Slovenia. Limnologica 2007, 37, 146–154. [Google Scholar] [CrossRef] [Green Version]
- Demars, B.O.L.; Edwards, A.C. Distribution of aquatic macrophytes in contrasting river systems: A critique of compositional-based assessment of water quality. Sci. Total Environ. 2009, 407, 975–990. [Google Scholar] [CrossRef]
- Sedell, J.R.; Richey, J.E.; Swanson, F.J. The river continuum concept: A basis for the expected ecosystem behavior of very large rivers? In Proceedings of the International Large River Symposium, Honey Harbour, ON, Canada, 14–19 September 1986. [Google Scholar]
- Hasenbichler, H.-P. Manual on Danube Navigation; Thomas, H., Vera, H., Eds.; via donau–Österreichische Wasserstraßen-Gesellschaft mbH: Vienna, Austria, 2013; p. 218. [Google Scholar]
- Vukov, D.; Ilic, M.; Cuk, M.; Igic, R.; Janauer, G. The relationship between habitat factors and aquatic macrophyte assemblages in the Danube river in Serbia. Arch. Biol. Sci. 2017, 69, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Fuller, M.R.; Doyle, M.W.; Strayer, D.L. Causes and consequences of habitat fragmentation in river networks. Ann. N. Y. Acad. Sci. 2015, 1355, 31–51. [Google Scholar] [CrossRef]
- ISTER. Donau-Stromkilometer-Stromauf-Stromab.pdf. Available online: http://www.ister.at/Ister1/wp-content/uploads/2019/06 (accessed on 8 January 2021).
- HPP-Bavaria. Available online: http://www.bew-augsburg.de/CBP/Wasserkraft/Daten/ODK.asp (accessed on 7 January 2021).
- Danube Commission. List of Locks. Available online: https://de.wikipedia.org/wiki/Liste_der_Schleusen_in_der_Donau (accessed on 7 January 2021).
- Kohler, A.; Janauer, G.A. Zur Methodik der Untersuchung von aquatischen Makrophyten in Fließgewässern. Handb. Angew. Limnol. Ecomed-Verlag 1995, VIII, 1–22. [Google Scholar] [CrossRef]
- Atlas Florae Europaeae (AFE)—Distribution of Vascular Plants in Europe. Available online: https://www.luomus.fi/en/atlas-florae-europaeae-afe-distribution-vascular-plants-europe (accessed on 23 December 2020).
- Corine Land Cover. Copernicus Land Monitoring Service—High Resolution Layer Water and Wetness; European Environment Agency: Copenhagen, Denmark, 2018; Available online: http://land.copernicus.eu (accessed on 22 December 2020).
- Schaumburg, J.; Schranz, C.; Foerster, J.; Gutowski, A.; Hofmann, G.; Meilinger, P.; Schneider, S.; Schmedtje, U. Ecological classification of macrophytes and phytobenthos for rivers in Germany according to the Water Framework Directive. Limnologica 2004, 34, 283–301. [Google Scholar] [CrossRef] [Green Version]
- Braun-Blanquet, J. Pflanzensoziologie, Grundzüge der Vegetationskunde, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 1964; p. 631. [Google Scholar]
- Lukács, B.A.; Tóthmérész, B.; Borics, G.; Várbíró, G.; Juhász, P.; Kiss, B.; Müller, Z.; G-Tóth, L.; Eros, T. Macrophyte diversity of lakes in the Pannon Ecoregion (Hungary). Limnologica 2015, 53, 74–83. [Google Scholar] [CrossRef]
- Pall, K.; Gecheva, G.; Soaru-Minea, A.; Lukacs, B.S.P. Intercalibration of the National Classifications of Ecological Status for Eastern Continental Lakes; Publications Office of the European Union: Luxembourg, 2018; p. 81. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Murtagh, F.; Legendre, P. Ward’s Hierarchical Agglomerative Clustering Method: Which Algorithms Implement Ward’s Criterion? J. Classif. 2014, 31, 274–295. [Google Scholar] [CrossRef] [Green Version]
- Swaine, M.D.; Adomako, J.; Ameka, G.; de Graft-Johnston, K.A.A.; Cheek, M. Forest river plants and water quality in Ghana. Aquat. Bot. 2006, 85, 299–308. [Google Scholar] [CrossRef]
- Wiegleb, G.; Bröring, U.; Filetti, M.; Brux, H.; Herr, W. Long-term dynamics of macrophyte dominance and growth-form types in two north-west German lowland streams. Freshw. Biol. 2014, 59, 1012–1025. [Google Scholar] [CrossRef]
- Zelnik, I. Vegetation of the Meadows from the Order Molinietalia W. Koch 1926 and Contact Sites in Slovenia. Ph.D. Thesis, University of Ljubljana (Slovenia), Ljubljana, Slovenia, 2005; p. 196. [Google Scholar]
- Hrivnák, R.; Bubíková, K.; Oťaheľová, H.; Šumberová, K. Formalised classification of aquatic vegetation in Slovakia. Phytocoenologia 2019, 49, 107–133. [Google Scholar] [CrossRef]
- Landucci, F.; Tichý, L.; Šumberová, K.; Chytrý, M. Formalized classification of species-poor vegetation: A proposal of a consistent protocol for aquatic vegetation. J. Veg. Sci. 2015, 26, 791–803. [Google Scholar] [CrossRef]
- Chytrý, M. Vegetation of the Czech Republic 3. Aquatic and wetland vegetation; Academia: Praha, Czech Republic, 2011; p. 828. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. CANOCO Reference Manual and CanoDraw for Windows User’s Guide: Software for Canonical Community Ordination (Version 4.5); www.canoco.com: Ithaca, NY, USA, 2002. [Google Scholar]
- Hudon, C.; Gagnon, P.; Amyot, J.P.; Létourneau, G.; Jean, M.; Plante, C.; Rioux, D.; Deschênes, M. Historical changes in herbaceous wetland distribution induced by hydrological conditions in Lake Saint-Pierre (St. Lawrence River, Quebec, Canada). In Proceedings of the Hydrobiologia; Springer: Berlin/Heidelberg, Germany, 2005; Volume 539, pp. 205–224. [Google Scholar]
- Kornijów, R.; Strayer, D.L.; Caraco, N.F. Macroinvertebrate communities of hypoxic habitats created by an invasive plant (Trapa natans) in the freshwater tidal Hudson River. Fundam. Appl. Limnol. 2010, 176, 199–207. [Google Scholar] [CrossRef]
- Skliar, I.L.; Skliar, V.H. Trapa natans L. s.l. growth features in Desna River basin. Ukr. J. Ecol. 2017, 7, 239–245. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, R.G. Land–water interfaces: Larger plants. In Limnology; Elsevier: Amsterdam, The Netherlands, 2001; pp. 527–575. [Google Scholar]
- Preston, C. Pondweeds of Great Britain and Ireland; Botanical Society of the British Isles: London, UK, 1995; p. 352. [Google Scholar]
- Germ, M.; Kuhar, U.; Gaberščik, A. Abundance and Diversity of Taxa Within the Genus Potamogeton in Slovenian Watercourses. In Natural and Constructed Wetlands; Vymazal, J., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 283–291. [Google Scholar]
- Kaplan, Z.; Jarolímová, V.; Fehrer, J. Revision of Chromosome Numbers of Potamogetonaceae: A New Basis for Taxonomic and Evolutionary Implications. Preslia 2013, 85, 421–482. [Google Scholar]
- Case, M.L.; Madsen, J.D. Factors limiting the growth of Stuckenia pectinata (sago pondweed) in heron lake, minnesota. J. Freshw. Ecol. 2004, 19, 17–23. [Google Scholar] [CrossRef]
- Kantrud, H.A. Sago Pondweed (Potamogeton pectinatus L.): A Literature Review; U.S. Fish and Wildlife Service: Washington, DC, USA, 1990; p. 89. [Google Scholar]
- Chambers, P.A.; Prepas, E.E.; Hamilton, H.R.; Bothwell, M.L. Current Velocity and Its Effect on Aquatic Macrophytes in Flowing Waters. Ecol. Appl. 1991, 1, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Hussner, A. Alien aquatic plant species in European countries. Weed Res. 2012, 52, 297–306. [Google Scholar] [CrossRef]
- Zehnsdorf, A.; Hussner, A.; Eismann, F.; Rönicke, H.; Melzer, A. Management options of invasive Elodea nuttallii and Elodea canadensis. Limnologica 2015, 51, 110–117. [Google Scholar] [CrossRef]
- Hérault, B.; Bornet, A.; Trémolières, M. Redundancy and niche differentiation among the European invasive Elodea species. Biol. Invasions 2008, 10, 1099–1107. [Google Scholar] [CrossRef]
- Kuhar, U.; Germ, M.; Gaberščik, A. Habitat characteristics of an alien species Elodea canadensis in Slovenian watercourses. Hydrobiologia 2010, 656, 205–212. [Google Scholar] [CrossRef]
- Thiébaut, G. Does competition for phosphate supply explain the invasion pattern of Elodea species? Water Res. 2005, 39, 3385–3393. [Google Scholar] [CrossRef] [PubMed]
- Grudnik, Z.M.; Jelenko, I.; Germ, M. Influence of abiotic factors on invasive behaviour of alien species Elodea nuttallii in the Drava River (Slovenia). Ann. Limnol. Int. J. Limnol. 2014, 50, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Simpson, D.A. Displacement of Elodea canadensis Michx by Elodea nuttallii (Planch.) H. St John in the British Isles. Watsonia 1990, 18, 173–177. [Google Scholar]
- Thiébaut, G. Invasion success of non-indigenous aquatic and semi-aquatic plants in their native and introduced ranges. A comparison between their invasiveness in North America and in France. Biol. Invasions 2006, 9, 1–12. [Google Scholar] [CrossRef]
- Barrat-Segretain, M.H.; Lemoine, D.G. Can snail herbivory influence the outcome of competition between Elodea species? Aquat. Bot. 2007, 86, 157–162. [Google Scholar] [CrossRef]
- Erhard, D.; Gross, E.M. Allelopathic activity of Elodea canadensis and Elodea nuttallii against epiphytes and phytoplankton. Aquat. Bot. 2006, 85, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Barrat-Segretain, M.H.; Cellot, B. Response of invasive macrophyte species to drawdown: The case of Elodea sp. Aquat. Bot. 2007, 87, 255–261. [Google Scholar] [CrossRef]
- Lumpkin, T.A.; Plucknett, D.L. Azolla: Botany, physiology, and use as a green manure. Econ. Bot. 1980, 34, 111–153. [Google Scholar] [CrossRef]
- Szczęśniak, E.; Błachuta, J.; Krukowski, M.; Picińska-Fałtynowicz, J. Distribution of Azolla filiculoides Lam. (Azollaceae) in Poland. Acta Soc. Bot. Pol. 2011, 78, 241–246. [Google Scholar] [CrossRef]
- Sabetraftar, K.; Zarkami, R.; Sadeghi, R.; Van Damme, P. A review of some ecological factors affecting the growth of Azolla spp. Casp. J. Environ. Sci. 2013, 11, 65–76. [Google Scholar]
- Wagner, G.M. Azolla: A Review of Its Biology and Utilization. Bot. Rev. 1997, 63, 1–26. [Google Scholar] [CrossRef]
- Paolacci, S.; Jansen, M.A.K.; Harrison, S. Competition Between Lemna minuta, Lemna minor, and Azolla filiculoides. Growing Fast or Being Steadfast? Front. Chem. 2018, 6, 207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolpagni, R.; Laini, A.; Soana, E.; Tomaselli, M.; Nascimbene, J. Growth performance of Vallisneria spiralis under oligotrophic conditions supports its potential invasiveness in mid-elevation freshwaters. Weed Res. 2015, 55, 185–194. [Google Scholar] [CrossRef]
- Anačkov, G.T.; Rat, M.M.; Radak, B.D.; Igić, R.S.; Vukov, D.M.; Rućando, M.M.; Krstivojević, M.M.; Radulović, S.B.; Cvijanović, D.L.; Milić, D.M.; et al. Alien invasive neophytes of the Southeastern part of the Pannonian Plain. Cent. Eur. J. Biol. 2013, 8, 1032–1043. [Google Scholar] [CrossRef]
- Illies, J. Versuch einer allgemeinen biozönotischen Gliederung der Fließgewässer. Int. Rev. Gesamten Hydrobiol. Hydrogr. 1961, 46, 205–213. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in Tropical Rain Forests and Coral Reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grime, J.P. Competitive Exclusion in Herbaceous Vegetation. Nature 1973, 242, 344–347. [Google Scholar] [CrossRef]
- Webster, J.R. Spiraling down the river continuum: Stream ecology and the U-shaped curve. J. N. Am. Benthol. Soc. 2007, 26, 375–389. [Google Scholar] [CrossRef]
- The Mosaic-Cycle Concept of Ecosystems; Remmert, H. (Ed.) Ecological Studies; Springer: Berlin/Heidelberg, Germany, 1991; Volume 85, p. 168. [Google Scholar]
- Engloner, A.; Szalma, E.; Sipos, K.; Dinka, M. Occurrence and habitat preference of aquatic macrophytes in a large river channel. Community Ecol. 2013, 14, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Oťaheľová, H.; Valachovic˘, M.; Hrivnák, R. The impact of environmental factors on the distribution pattern of aquatic plants along the Danube River corridor (Slovakia). Limnologica 2007, 37, 290–302. [Google Scholar] [CrossRef] [Green Version]
- Markovics, R.; Kanduč, T.; Szramek, K.; Golobočanin, D.; Milačič, R.; Ogrinc, N. Chemical dynamics of the Sava riverine system—A stable isotopic approach. J. Environ. Monit. 2010, 12, 2165–2176. [Google Scholar] [CrossRef] [PubMed]
- Chmara, R.; Banaś, K.; Szmeja, J. Changes in the structural and functional diversity of macrophyte communities along an acidity gradient in softwater lakes. Flora-Morphol. Distrib. Funct. Ecol. Plants 2015, 216, 57–64. [Google Scholar] [CrossRef]
- Riis, T.; Sand-Jensen, K. Historical changes in species composition and richness accompanying perturbation and eutrophication of Danish lowland streams over 100 years. Freshw. Biol. 2001, 46, 269–280. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster No. | N | Mean Secchi Depth (cm) | Mean No. of Species | Dominant Species | Adj. Mean Abundance Values | Species Frequency (%) | Association |
|---|---|---|---|---|---|---|---|
| 1 | 86 | 126 ± 45 | 4.5 ± 1.7 | Myr spi | 32.8 | 100 | Potamo pectinati-Myriophylletum spicati Rivas Goday 1964 |
| Pot per | 6.5 | 78 | |||||
| Ali pla | 3.5 | 78 | |||||
| 2 | 48 | 72 ± 19 | 3.6 ± 2.4 | But umb | 28.7 | 100 | Butometum umbellati Philippi 1973 |
| Agr sto | 4.8 | 42 | |||||
| Spa ere | 2.1 | 23 | |||||
| 3 | 60 | 66 ± 23 | 1.8 ± 1.2 | Pot acu | 25.1 | 100 | Potametum acutifolii Segal ex Šumberová et Hrivnák in Chytrý 2011 |
| Aco cal | 1.8 | 100 | |||||
| 4 | 5 | 66 ± 2 | 3.8 ± 2.5 | Zan pal | 41.8 | 100 | Parvo-Potamo-Zannichellietum pedicellatae Soó 1947 |
| But umb | 3.4 | 60 | |||||
| Pot pec | 2.0 | 60 | |||||
| 5 | 74 | 48 ± 28 | 3.3 ± 2.2 | Pot pec | 8.4 | 97 | Species-poor and abundance low community with Potamogeton pectinatus |
| Elo nut | 1.6 | 19 | |||||
| Pot cri | 1.5 | 34 | |||||
| Pha aru | 1.4 | 32 | |||||
| 6 | 1345 | 70 ± 25 | 1.5± 2.2 | But umb | 0.42 | 21 | Species-poorest and abundance lowest sections. |
| Pha aru | 0.36 | 15 | |||||
| Pot per | 0.34 | 6 | |||||
| 7 | 219 | 89 ± 40 | 4.7 ± 2.1 | Myr spi | 6.6 | 86 | suboptimal community with Myriophyllum spicatum |
| Cer dem | 3.7 | 42 | |||||
| Pot per | 3.4 | 52 | |||||
| Pot cri | 3.3 | 61 | |||||
| 8 | 17 | / | 5.4 ± 1.0 | Pot cri | 26.4 | 100 | Potametum crispi Soo 1927 |
| Myr spi | 9.1 | 100 | |||||
| Pot pus | 7.5 | 94 | |||||
| 9 | 5 | 116 ± 18 | 12.0 ± 4.1 | Pot pec | 63.3 | 100 | Potametum perfoliati Miljan 1933 |
| Pot per | 63.2 | 100 | |||||
| Cer dem | 16.1 | 80 | |||||
| 10 | 29 | 78 ± 21 | 8.1 ± 3.9 | Pot pec | 65.1 | 100 | Potametum pectinati Carstensen ex Hilbig 1971 |
| Pot per | 14.7 | 85 | |||||
| Myr spi | 6.8 | 45 | |||||
| 11 | 29 | 100 ± 21 | 10.0 ± 2.3 | Pot nat | 26.2 | 100 | Potametum natantis Hild 1959 |
| Pot per | 22.1 | 97 | |||||
| Pot gra | 17.3 | 97 | |||||
| 12 | 12 | 96 ± 17 | 11.5 ± 1.3 | Pot nat | 29.8 | 100 | Potametum natantis Hild 1959and Trapetum natantis Kárpáti 1963 (6 out of 12) |
| Pot pec | 25.5 | 100 | |||||
| Tra nat | 20.7 | 83 | |||||
| Pot per | 17.8 | 75 | |||||
| 13 | 19 | 130 ± 10 | 12.2 ± 2.8 | Cer sub | 21.3 | 89 | Potamo-Ceratophylletum submersi Pop 1962 |
| Spi pol | 18.0 | 100 | |||||
| Pot pec | 5.9 | 95 | |||||
| Pot luc | 4.7 | 95 | |||||
| Lem min | 2.1 | 89 | |||||
| 14 | 33 | 84 ± 23 | 11.8 ± 2.9 | Pot luc | 23.5 | 100 | Potametum lucentis Hueck 1931 |
| Pot pec | 16.1 | 97 | |||||
| Cer dem | 12.7 | 97 | |||||
| Spi pol | 8.7 | 100 | |||||
| 15 | 27 | 69 ± 23 | 6.0 ± 3.2 | Pot pec | 31.6 | 100 | Potametum pectinati Carstensen ex Hilbig 1971 |
| Pot nat | 3.8 | 25.9 | |||||
| Pot per | 3.7 | 44.4 | |||||
| 16 | 7 | 123 ± 19 | 9.6 ± 3.3 | Pot per | 62.3 | 100 | Potametum perfoliati Miljan 1933 |
| Cer dem | 20.8 | 86 | |||||
| Pot gra | 6.6 | 43 | |||||
| 17 | 44 | 109 ± 32 | 7.2 ± 2.8 | Pot per | 26.1 | 93 | Potametum perfoliati Miljan 1933 |
| Pot pec | 26.2 | 75 | |||||
| Myr spi | 13.8 | 75 | |||||
| 18 | 10 | 68 ± 9 | 12.4 ± 2.9 | Pot pec | 47.4 | 100 | Potametum lucentis Hueck 1931 |
| Pot luc | 41.9 | 100 | |||||
| Cer dem | 23.4 | 100 | |||||
| Pot gra | 17.8 | 100 | |||||
| 19 | 18 | 78 ± 7 | 12.9 ± 2.5 | Pot pec | 32.7 | 100 | Potametum graminei Lang 1967 |
| Cer dem | 29.6 | 100 | |||||
| Pot gra | 27.2 | 100 | |||||
| Pot luc | 19.4 | 94 | |||||
| 20 | 23 | 70 ± 9 | 8.8 ± 3.8 | Cer dem | 30.6 | 100 | Ceratophylletum demersi Corillion 1957 |
| Spi pol | 13.5 | 57 | |||||
| Pot luc | 7.8 | 26 | |||||
| 21 | 10 | 125 ± 18 | 13.2 ± 2.5 | Spi pol | 56.8 | 100 | Lemno-Spirodeletum polyrhizae Koch 1954 (6) and Ceratophylletum demersi Corillion 1957 (4) |
| Cer dem | 29.3 | 90 | |||||
| Cer sub | 19.8 | 90 | |||||
| Wol arr | 14.5 | 60 |
| Analysis * | Data Set | Total Explained Variance (%) | Significant Parameters/Explained Variance |
|---|---|---|---|
| RDA | Whole course (all species) | 8 | All, with the exception of land use |
| CCA | Whole course; only species with total abundance more than 1% | 21 | All parameters, with exception of flow velocity Secchi depth (R) (6%, p = 0.001) Distance from the source (4%, p = 0.001) Current velocity (R) (3%, p = 0.001) Other parameters (8%, p ≤ 0.014) |
| CCA | Neophyte species only | 45 | Current velocity (L) (17%, p = 0.001) Secchi depth (L) (12%, p = 0.001) Sediment type (L) (7%, p = 0.002) No of species per stretch (5%, p = 0.001) Bank structure (R) (3%, p = 0.001) Current velocity (R) (1%, p = 0.022) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janauer, G.A.; Exler, N.; Anačkov, G.; Barta, V.; Berczik, Á.; Boža, P.; Dinka, M.; Georgiev, V.; Germ, M.; Holcar, M.; et al. Distribution of the Macrophyte Communities in the Danube Reflects River Serial Discontinuity. Water 2021, 13, 918. https://doi.org/10.3390/w13070918
Janauer GA, Exler N, Anačkov G, Barta V, Berczik Á, Boža P, Dinka M, Georgiev V, Germ M, Holcar M, et al. Distribution of the Macrophyte Communities in the Danube Reflects River Serial Discontinuity. Water. 2021; 13(7):918. https://doi.org/10.3390/w13070918
Chicago/Turabian StyleJanauer, Georg A., Norbert Exler, Goran Anačkov, Veronika Barta, Árpád Berczik, Pal Boža, Mária Dinka, Valeri Georgiev, Mateja Germ, Matej Holcar, and et al. 2021. "Distribution of the Macrophyte Communities in the Danube Reflects River Serial Discontinuity" Water 13, no. 7: 918. https://doi.org/10.3390/w13070918
APA StyleJanauer, G. A., Exler, N., Anačkov, G., Barta, V., Berczik, Á., Boža, P., Dinka, M., Georgiev, V., Germ, M., Holcar, M., Hrivnák, R., Igić, R., Ozimec, S., Sârbu, A., Schmidt, B., Schmidt-Mumm, U., Schütz, W., Sipos, K., Szalma, E., ... Gaberščik, A. (2021). Distribution of the Macrophyte Communities in the Danube Reflects River Serial Discontinuity. Water, 13(7), 918. https://doi.org/10.3390/w13070918