
Population Characteristics of the European Lobster, Homarus gammarus in the Adriatic Sea: Implications for Sustainable Fisheries Management



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Data Collection
2.3. Data Analysis
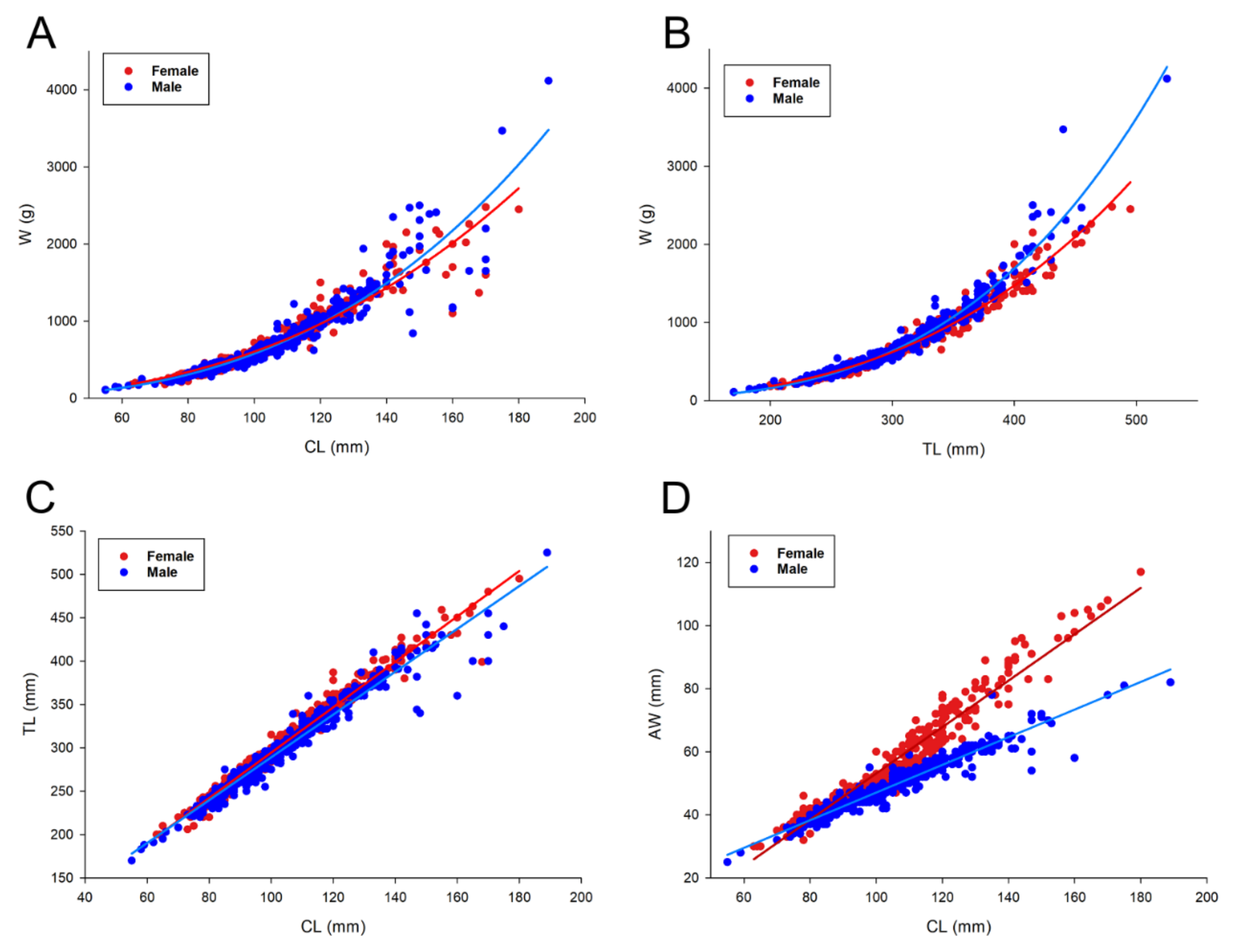
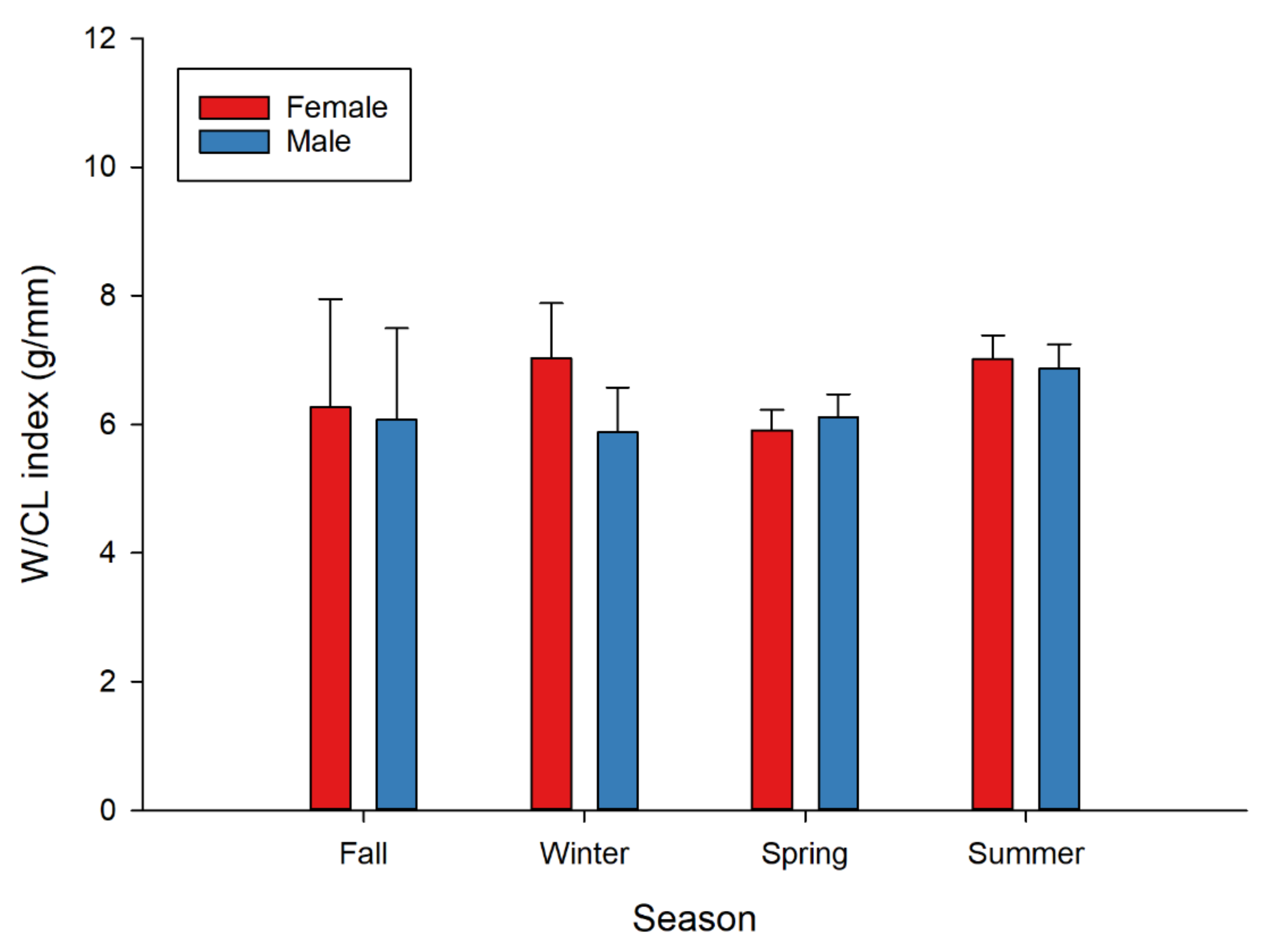
2.3.1. Morphometric Relationships and W/CL Index
2.3.2. Multivariate Analysis
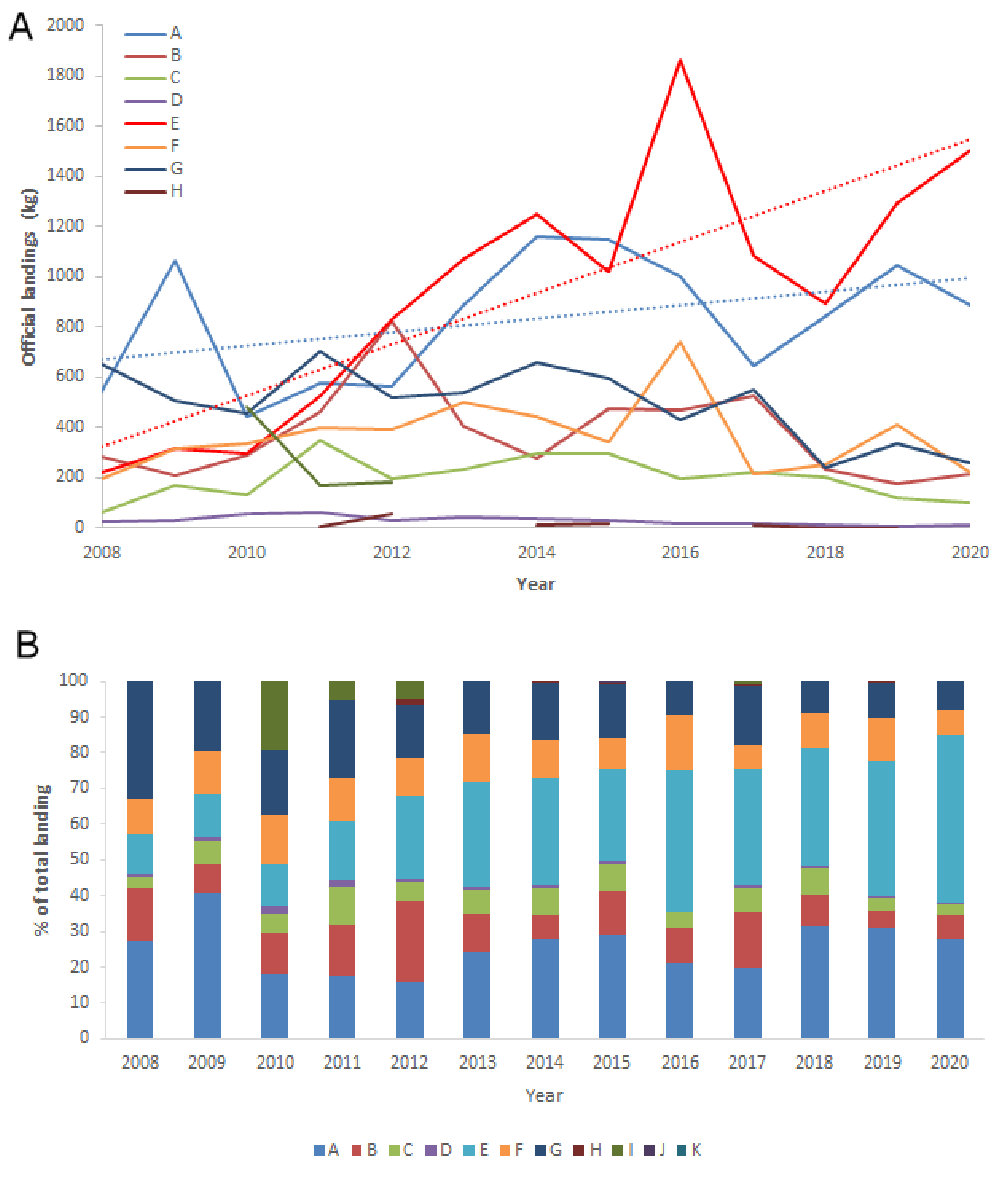
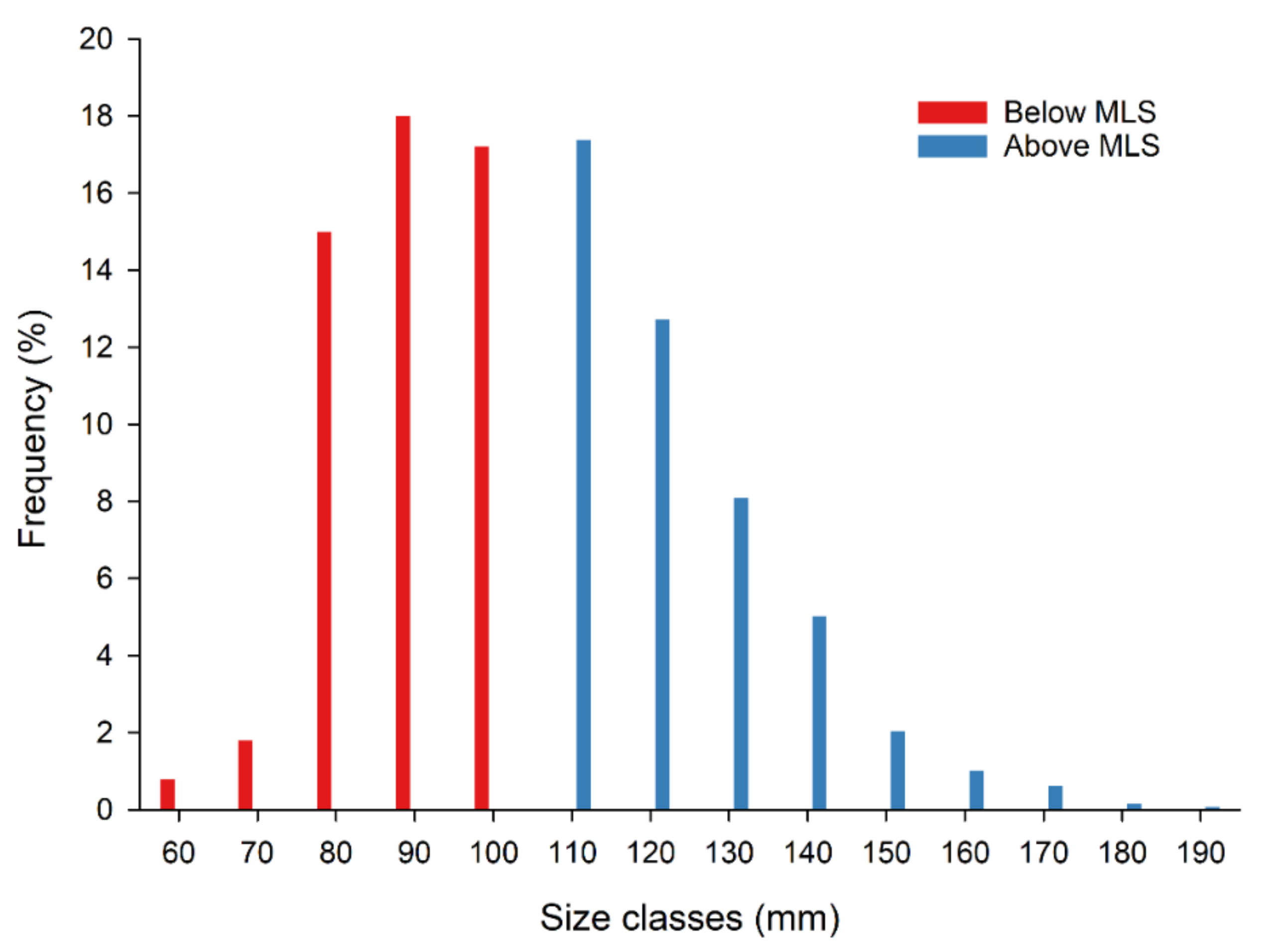
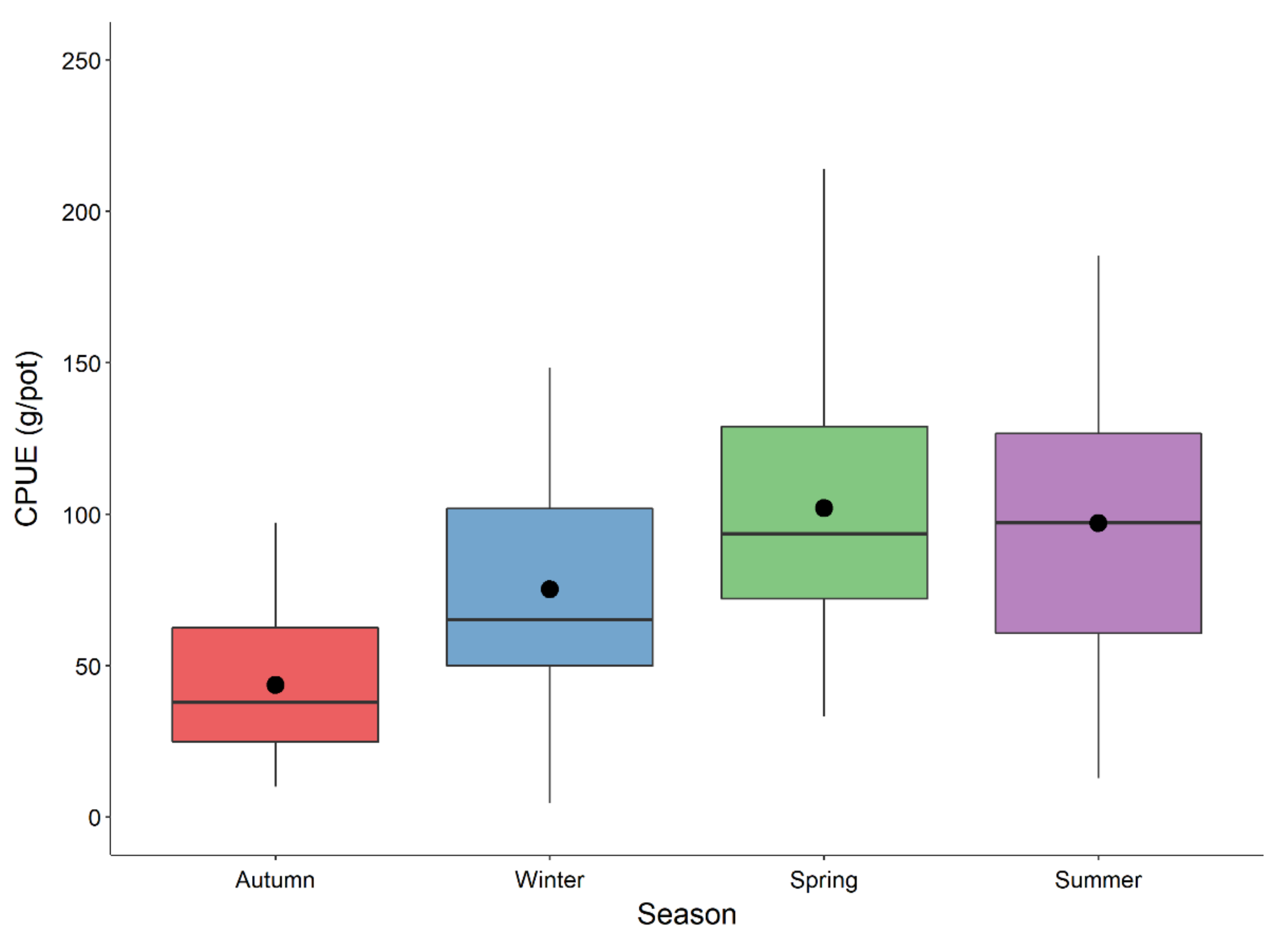
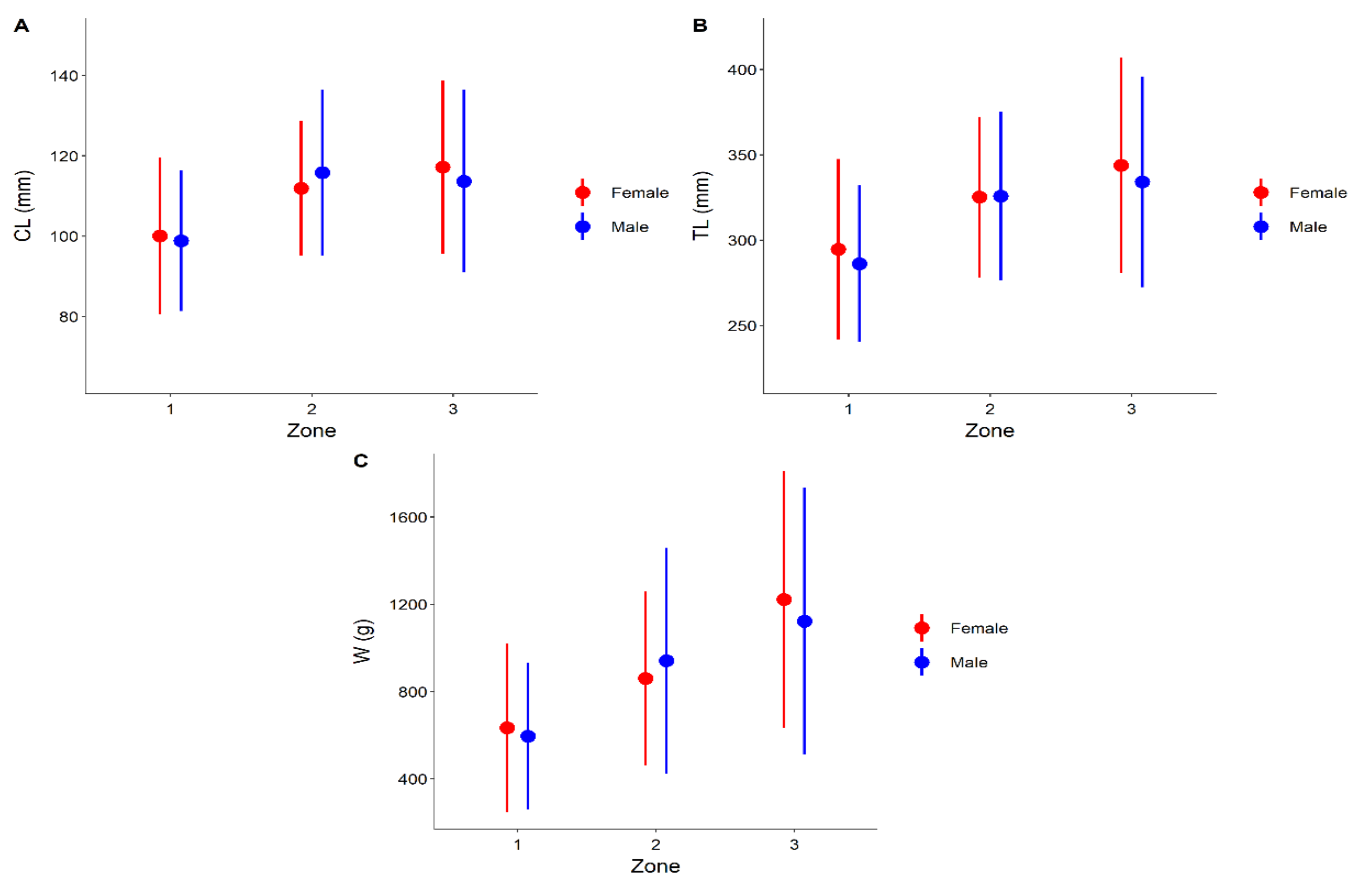
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jennings, S. Indicators to support an ecosystem approach to fisheries. Fish Fish. 2005, 6, 212–232. [Google Scholar] [CrossRef]
- Lleonart, J.; Maynou, F. Fish stock assessments in the Mediterranean: State of the art. Sci. Mar. 2003, 67, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Tzanatos, E.; Somarakis, S.; Tserpes, G.; Koutsikopoulos, C. Identifying and classifying small-scale fisheries métiers in the Mediterranean: A case study in the Patraikos Gulf, Greece. Fish. Res. 2006, 81, 158–168. [Google Scholar] [CrossRef]
- Tzanatos, E.; Castro, J.; Forcada, A.; Matić-Skoko, S.; Gaspar, M.; Koutsikopoulos, C. A Métier-Sustainability-Index (MSI25) to evaluate fisheries components: Assessment of cases from data-poor fisheries from southern Europe. ICES J. Mar. Sci. 2013, 70, 78–98. [Google Scholar] [CrossRef] [Green Version]
- Guyader, O.; Berthou, P.; Koutsikopoulos, C.; Alban, F.; Demanèche, S.; Gaspar, M.B.; Eschbaum, R.; Fahy, E.; Tully, O.; Reynal, L.; et al. Small scale fisheries in Europe: A comparative analysis based on a selection of case studies. Fish. Res. 2013, 140, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lloret, J.; Cowx, I.G.; Cabral, H.; Castro, M.; Font, T.; Gonçalves, J.M.S.; Gordoa, A.; Hoefnagel, E.; Matić-Skoko, S.; Mikkelsen, E.; et al. Small-scale coastal fisheries in European Seas are not what they were: Ecological, social and economic changes. Mar. Policy 2018, 98, 176–186. [Google Scholar] [CrossRef]
- Grati, F.; Aladžuz, A.; Azzurro, E.; Bolognini, L.; Carbonara, P.; Çobani, M.; Domenichetti, F.; Dragičević, B.; Dulčić, J.; Durović, M.; et al. Seasonal dynamics of small-scale fisheries in the Adriatic Sea. Mediterr. Mar. Sci. 2018, 19, 21–35. [Google Scholar] [CrossRef] [Green Version]
- Marin, J. Exploitation, biologie et dymanique du stock de langouste rouge de corse Panulirus elephas Fabricus. Ph.D. Thesis, Université d’Aix-Marseille II, Marseille, France, 1987. [Google Scholar]
- Quetglas, A.; Gaamour, A.; Reñones, O.; Missaoui, H.; Zarrouk, T.; Elabed, A.; Goñi, R. Common spiny lobster (Palinurus elephas Fabricius 1787) fisheries in the western Mediterranean: A comparison of Spanish and Tunisian fisheries. Bolleti Soc. D’hist. Nat. Balear. 2004, 47, 63–80. [Google Scholar]
- Gönülal, O. Spiny lobster (Palinurus elephas Fabricius, 1787) and common lobster (Homarus gammarus Linnaeus, 1758) fishing in the Agean Sea. In The Aegean Sea Marine Biodiversity, Fisheries, Conservation and Governance; Katağan, T., Tokaç, A., Beşiktepe, Ş., Öztürk, B., Eds.; Turkish Marine Research Foundation (TUDAV): Istanbul, Turkey, 2015; p. 714. ISBN 978-975-8825-33-2. [Google Scholar]
- Lotze, H.K.; Coll, M.; Dunne, J.A. Historical Changes in Marine Resources, Food-web Structure and Ecosystem Functioning in the Adriatic Sea, Mediterranean. Ecosystems 2011, 14, 198–222. [Google Scholar] [CrossRef]
- FAO Fisheries & Aquaculture—Species Fact Sheets—Homarus gammarus (Linnaeus, 1758). Available online: http://www.fao.org/fishery/species/2648/en (accessed on 26 February 2020).
- Prodöhl, P.A.; Jørstad, K.E.; Triantafyllidis, A.; Katsares, V.; Triantaphyllidis, C. European lobster-Homarus gammarus. Genetic Impact of Aquaculture Activities on Native Populations. Genimpact Final Scientific Report (EU contract n. RICA-CT-2005-022802). 2007, pp. 91–98. Available online: http://genimpact.imr.no/ (accessed on 9 April 2021).
- Pere, A.; Marengo, M.; Lejeune, P.; Durieux, E.D.H. Evaluation of Homarus gammarus (Crustacea: Decapoda: Nephropidae) catches and potential in a mediterranean small-scale fishery. Sci. Mar. 2019, 83, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Goñi, R.; Latrouite, D. Review of the biology, ecology and fisheries of Palinurus spp. species of European waters: Palinurus elephas (Fabricius, 1787) and Palinurus mauritanicus (Gruvel, 1911). Cah. Biol. Mar. 2005, 46, 127–142. [Google Scholar]
- Lloret, J.; Riera, V. Evolution of a mediterranean coastal zone: Human impacts on the marine environment of Cape Creus. Environ. Manag. 2008, 42, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Pere, A. Déclin des Populations de Langouste Rouge et Baisse de la Ressource Halieutique en Corse: Causes et Perspectives. Ph.D. Thesis, Université de Corse, Corte, France, 2012; p. 478. [Google Scholar]
- Dow, R.L. The Clawed Lobster Fisheries. In The Biology and Management of Lobsters; Elsevier Inc.: Amsterdam, The Netherlands, 1980; Volume 2, pp. 265–316. ISBN 9780080917344. [Google Scholar]
- Cobb, J.S.; Castro, K.M. Homarus Species. In Lobsters: Biology, Management, Aquaculture and Fisheries; Blackwell Publishing Ltd.: Oxford, UK, 2006; pp. 310–339. [Google Scholar]
- Browne, R.M.; Mercer, J.P.; Duncan, M.J. An historical overview of the Republic of Ireland’s lobster (Homarus gammarus Linnaeus) fishery, with reference to European and North American (Homarus americanus Milne Edwards) lobster landings. In Coastal Shellfish A Sustainable Resource; Springer: Amsterdam, The Netherlands, 2001; pp. 49–62. [Google Scholar]
- European Union. Council Regulation (EC) No 1967/2006 of 21 December 2006 Concerning Management Measures for the Sustainable Exploitation of Fishery Resources in the Mediterranean Sea, Amending Regulation (EEC) No 2847/93 and Repealing Regulation (EC) No 1626/94; European Union: Brussels, Belgium, 2006. [Google Scholar]
- Coleman, M.T.; Agnalt, A.-L.; Emmerson, J.; Laurens, M.; Porter, J.S.; Bell, M.C. From the Adriatic to Northern Norway—geographic differences in moult increment and moult probability of the European lobster (Homarus gammarus), across the natural range. ICES J. Mar. Sci. 2020. [Google Scholar] [CrossRef]
- Pavičić, M.; Žužul, I.; Matić-Skoko, S.; Triantafyllidis, A.; Grati, F.; Durieux, E.D.H.; Celić, I.; Šegvić-Bubić, T. Population Genetic Structure and Connectivity of the European Lobster Homarus gammarus in the Adriatic and Mediterranean Seas. Front. Genet. 2020, 11, 1540. [Google Scholar] [CrossRef] [PubMed]
- Debuse, V.J.; Addison, J.T.; Reynolds, J.D. Morphometric variability in UK populations of the European lobster. J. Mar. Biol. Assoc. U. K. 2001, 81, 469–474. [Google Scholar] [CrossRef]
- Galparsoro, I.; Borja, Á.; Bald, J.; Liria, P.; Chust, G. Predicting suitable habitat for the European lobster (Homarus gammarus), on the Basque continental shelf (Bay of Biscay), using Ecological-Niche Factor Analysis. Ecol. Model. 2009, 220, 556–567. [Google Scholar] [CrossRef]
- Lizárraga-Cubedo, H.A.; Tuck, I.; Bailey, N.; Pierce, G.J.; Zuur, A.F.; Bova, D. Scottish lobster fisheries and environmental variability. ICES J. Mar. Sci. 2015, 72, i211–i224. [Google Scholar] [CrossRef] [Green Version]
- Tully, O.; Roantree, V.; Robinson, M. Maturity, fecundity and reproductive potential of the European lobster (Homarus gammarus) in Ireland. J. Mar. Biol. Assoc. U. K. 2001, 81, 61–68. [Google Scholar] [CrossRef]
- Skerritt, D.J.; Robertson, P.A.; Mill, A.C.; Polunin, N.V.C.; Fitzsimmons, C. Fine-scale movement, activity patterns and home-ranges of European lobster Homarus gammarus. Mar. Ecol. Prog. Ser. 2015, 536, 203–219. [Google Scholar] [CrossRef] [Green Version]
- Phillips, B.F. Lobsters: Biology, Management, Aquaculture and Fisheries; Phillips, B.F., Ed.; John Wiley & Sons, Ltd.: Oxford, UK, 2013; ISBN 9781118517444. [Google Scholar]
- Triantafyllidis, A.; Apostolidis, A.P.; Katsares, V.; Kelly, E.; Mercer, J.; Hughes, M.; Jørstad, K.E.; Tsolou, A.; Hynes, R.; Triantaphyllidis, C. Mitochondrial DNA variation in the European lobster (Homarus gammarus) throughout the range. Mar. Biol. 2005, 146, 223–235. [Google Scholar] [CrossRef]
- Ellis, C.D.; Hodgson, D.J.; Daniels, C.L.; Collins, M.; Griffiths, A.G.F. Population genetic structure in European lobsters: Implications for connectivity, diversity and hatchery stocking. Mar. Ecol. Prog. Ser. 2017, 563, 123–137. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, T.L.; Ellis, C.D.; Triantafyllidis, A.; Stevens, J.R. Single nucleotide polymorphisms reveal a genetic cline across the north-east Atlantic and enable powerful population assignment in the European lobster. Evol. Appl. 2019, 12, 1881–1899. [Google Scholar] [CrossRef] [Green Version]
- Devescovi, M.; Lucu, Č. Growth patterns of the lobster Homarus gammarus in the North Adriatic. Period. Biol. 2000, 102, 189–193. [Google Scholar]
- Martínez-Calderón, R.; Lozano-Álvarez, E.; Briones-Fourzán, P. Morphometric relationships and seasonal variation in size, weight, and a condition index of post-settlement stages of the Caribbean spiny lobster. PeerJ 2018, 2018, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Murphy, M.C.; Kruse, G.H. A length-based population model and stock-recruitment relationships for red king crab, Paralithodes camtschaticus, in Bristol Bay, Alaska. Can. J. Fish. Aquat. Sci. 1995, 52, 1229–1246. [Google Scholar] [CrossRef]
- Santos, M.N.; Gaspar, M.B.; Vasconcelos, P.; Monteiro, C.C. Weight-length relationships for 50 selected fish species of the Algarve coast (southern Portugal). Fish. Res. 2002, 59, 289–295. [Google Scholar] [CrossRef]
- Baggio, M. Estimating weight-length relationships without individual weight data: An application to the American lobster (Homarus americanus) fishery of Long Island Sound. Rev. Fish Biol. Fish. 2011, 21, 771–777. [Google Scholar] [CrossRef] [Green Version]
- Lizárraga-Cubedo, H.A.; Tuck, I.; Bailey, N.; Pierce, G.J.; Kinnear, J.A.M. Comparisons of size at maturity and fecundity of two Scottish populations of the European lobster, Homarus gammarus. Fish. Res. 2003, 65, 137–152. [Google Scholar] [CrossRef]
- Conan, G.Y.; Comeau, M.; Moriyasu, M. Are Morphometrical Approaches Appropriate to Establish Size at Maturity for Male American Lobster, Homarus americanus? J. Crustac. Biol. 2001, 21, 937–947. [Google Scholar] [CrossRef] [Green Version]
- Aubry, F.B.; Cossarini, G.; Acri, F.; Bastianini, M.; Bianchi, F.; Camatti, E.; De Lazzari, A.; Pugnetti, A.; Solidoro, C.; Socal, G. Plankton communities in the northern Adriatic Sea: Patterns and changes over the last 30 years. Estuar. Coast. Shelf Sci. 2012, 115, 125–137. [Google Scholar] [CrossRef]
- Djakovac, T.; Degobbis, D.; Supić, N.; Precali, R. Marked reduction of eutrophication pressure in the northeastern Adriatic in the period 2000–2009. Estuar. Coast. Shelf Sci. 2012, 115, 25–32. [Google Scholar] [CrossRef]
- Shaltout, M.; Omstedt, A. Recent sea surface temperature trends and future scenarios for the Mediterranean Sea. Oceanologia 2014, 56, 411–443. [Google Scholar] [CrossRef] [Green Version]
- Vilibić, I.; Zemunik, P.; Šepić, J.; Dunić, N.; Marzouk, O.; Mihanović, H.; Denamiel, C.; Precali, R.; Djakovac, T. Present climate trends and variability in thermohaline properties of the northern Adriatic shelf. Ocean Sci. 2019, 15, 1351–1362. [Google Scholar] [CrossRef] [Green Version]
- Warton, D.I.; Wright, I.J.; Falster, D.S.; Westoby, M. Bivariate line-fitting methods for allometry. Biol. Rev. 2006, 81, 259. [Google Scholar] [CrossRef] [PubMed]
- Ricker, W.E. Linear Regressions in Fishery Research. J. Fish. Res. Board Can. 1973, 30, 409–434. [Google Scholar] [CrossRef]
- Zar, J.H. Biostatistical Analysis, 4th ed.; Prentice Hall: Hoboken, NJ, USA, 1999; ISBN 9780130815422. [Google Scholar]
- Hartnoll, R.G. Growth. In The Biology of Crustacea: Embriology, Morphology and Genetics; Abele, L.G., Ed.; Academic Press: New York, NY, USA, 1982; pp. 11–196. ISBN 0-12-106402-6. [Google Scholar]
- Anderson, M.J. Permutation tests for univariate or multivariate analysis of variance and regression. Can. J. Fish. Aquat. Sci. 2001, 58, 626–639. [Google Scholar] [CrossRef]
- Briones-Fourzán, P.; Baeza-Martínez, K.; Lozano-Álvarez, E. Nutritional indices of juvenile Caribbean spiny lobsters in a Mexican reef lagoon: Are changes over a 10-year span related to the emergence of Panulirus argus Virus 1 (PaV1)? J. Exp. Mar. Biol. Ecol. 2009, 370, 82–88. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial; PRIMER-E Ltd.: Plymouth, UK, 2015. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria. Available online: https://www.r-project.org/ (accessed on 1 July 2019).
- Tully, O. Impact of the v-notch technical conservation measure on reproductive potential in a lobster (Homarus gammarus L.) fishery in Ireland. Mar. Freshw. Res. 2001, 52, 1551–1557. [Google Scholar] [CrossRef]
- Cushing, D.H. Climate and Fisheries; Academic Press: New York, NY, USA, 1982; ISBN 9780121997205. [Google Scholar]
- Matić-Skoko, S.; Stagličić, N.; Pallaoro, A.; Kraljević, M.; Dulčić, J.; Tutman, P.; Dragičević, B. Effectiveness of conventional management in Mediterranean type artisanal fisheries. Estuar. Coast. Shelf Sci. 2011, 91, 314–324. [Google Scholar] [CrossRef]
- Stagličić, N.; Matić-Skoko, S.; Pallaoro, A.; Grgičević, R.; Kraljević, M.; Tutman, P.; Dragičević, B.; Dulčić, J. Long-term trends in the structure of eastern Adriatic littoral fish assemblages: Consequences for fisheries management. Estuar. Coast. Shelf Sci. 2011, 94, 263–271. [Google Scholar] [CrossRef]
- Matić-Skoko, S.; Stagličić, N.; Blažević, D.; Šiljić, J.; Iritani, D. Croatian marine fisheries (Adriatic Sea): 1950-2013. In Global atlas of Marine Fisheries—A Critical Appraisal of Catches and Ecosystem Impacts; Pauly, D., Zeller, D., Eds.; Island Press: Washington, DC, USA, 2016; p. 232. [Google Scholar]
- Caputi, N.; Lestang, S.; Frusher, S.; Wahle, R.A. The Impact of Climate Change on Exploited Lobster Stocks. In Lobsters: Biology, Management, Aquaculture and Fisheries; John Wiley & Sons, Ltd.: Oxford, UK, 2013; pp. 84–112. [Google Scholar]
- Green, B.S.; Gardner, C.; Hochmuth, J.D.; Linnane, A. Environmental effects on fished lobsters and crabs. Rev. Fish Biol. Fish. 2014, 24, 613–638. [Google Scholar] [CrossRef]
- Ben Rais Lasram, F.; Guilhaumon, F.; Albouy, C.; Somot, S.; Thuiller, W.; Mouillot, D. The Mediterranean Sea as a ‘cul-de-sac’ for endemic fishes facing climate change. Glob. Chang. Biol. 2010, 16, 3233–3245. [Google Scholar] [CrossRef]
- Pranovi, F.; Caccin, A.; Franzoi, P.; Malavasi, S.; Zucchetta, M.; Torricelli, P. Vulnerability of artisanal fisheries to climate change in the Venice Lagoon. J. Fish Biol. 2013, 83, 847–864. [Google Scholar] [CrossRef]
- Ministry of Agriculture. Pravilnik o Zaštiti Riba i Drugih Morskih Organizama. Narodne Novine 42/2016. No. 1096. (In Croatian). Available online: https://narodne-novine.nn.hr/eli/sluzbeni/2016/42/1096 (accessed on 9 April 2021).
- Wahle, R.A.; Castro, K.M.; Tully, O.; Cobb, J.S. Homarus. In Lobsters: Biology, Management, Aquaculture and Fisheries; John Wiley & Sons, Ltd.: Oxford, UK, 2013; pp. 221–258. [Google Scholar]
- McCleese, D.W.; Wildner, D.G. Overview of the Inshore Lobster Resources in the Scotia-Fundy Region. In Canadian Atlantic Fisheries Scientific Advisory Committee (CAFSAC) Research Document 85; CAFSAC: Halifax, NS, Canada, 1958; p. 20. [Google Scholar]
- Krouse, J.S. Performance and selectivity of trap fisheries for crustaceans. In Marine Invertebrate Fisheries: Their Assessment and Management; Caddy, J.F., Ed.; Wiley: New York, NY, USA, 1989; pp. 307–325. [Google Scholar]
- Latrouite, D. Le homard (Homarus gammarus) du nord golfe de Gascogne-Manche ouest (divisions VIIe + VIIIa du CIEM). In Contrat Ifremer/MAPA—Réf. 99-I1-03-01 Rapport Final: 1–13; Ifremer: Brest, France, 2001. [Google Scholar]
- Bennet, D.B.; Howard, A.E. Estimates of lobster (Homarus gammarus) fecundity from East and West Britain. ICES Document CM 1987/K: 47; Oxford University Press: Oxford, UK, 1987. [Google Scholar]
- Laurans, M.; Fifas, S.; Demaneche, S.; Brérette, S.; Debec, O. Modelling seasonal and annual variation in size at functional maturity in the European lobster (Homarus gammarus) from self-sampling data. ICES J. Mar. Sci. 2009, 66, 1892–1898. [Google Scholar] [CrossRef] [Green Version]
- Schoeman, D.S.; Cockcroft, A.C.; Van Zyl, D.L.; Goosen, P.C. South African rock lobster Jasus lalandii commercial fishery. S. Afr. J. Mar. Sci. 2010, 24, 37–48. [Google Scholar] [CrossRef] [Green Version]
- Ministry of Agriculture. Pravilnik o Obavljanju Gospodarskog Ribolova na Moru Mrežama Stajaćicama, Klopkastim, Udičarskim i Probodnim Ribolovnim Alatima te Posebnim Načinima Ribolova. Narodne Novine 84/2015. No. 1640. (In Croatian). Available online: https://narodne-novine.nn.hr/eli/sluzbeni/2015/84/1640 (accessed on 9 April 2021).
- Shelton, P.M.J.; Shelton, R.G.J.; Richards, P.R. Eye development in relation to molt stage in the European lobster Homarus gammarus. J. Cons. 1981, 39, 239–243. [Google Scholar] [CrossRef]
- Fütterer, D.; Paul, J. Recent and Pleistocene sediments off the Istrian coast (Northern Adriatic). Senckenbergiana Marit. 1976, 8, 1–21. [Google Scholar]
- Smith, I.P.; Jensen, A.C.; Collins, K.J.; Mattey, E.L. Movement of wild European lobsters Homarus gammarus in natural habitat. Mar. Ecol. Prog. Ser. 2001, 222, 177–186. [Google Scholar] [CrossRef]
- Uglem, I.; Belchier, M.; Svasand, T. Age determination of European lobsters (Homarus gammarus L.) by histological quantification of lipofuscin. J. Crustac. Biol. 2005, 25, 95–99. [Google Scholar] [CrossRef] [Green Version]
- Kilada, R.; Driscoll, J.G. Age determination in crustaceans: A review. Hydrobiologia 2017, 799, 21–36. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Merilä, J. Heritable variation and evolution under favourable and unfavourable conditions. Trends Ecol. Evol. 1999, 14, 96–101. [Google Scholar] [CrossRef]
- Schluter, D. The Ecology of Adaptive Radiation; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Ward, R.D.; Woodwark, M.; Skibinski, D.O.F. A comparison of genetic diversity levels in marine, freshwater, and anadromous fishes. J. Fish Biol. 1994, 44, 213–232. [Google Scholar] [CrossRef]
- Ribeiro, F.B.; Matthews Cascon, H.; Bezerra, L.E.A. Morphometric sexual maturity and allometric growth of the crab Sesarma rectum Randall, 1840 (Crustacea: Sesarmidae) in an impacted tropical mangrove in northeast Brazil. Lat. Am. J. Aquat. Res. 2013, 41, 361–368. [Google Scholar] [CrossRef]
- Glamuzina, L.; Conides, A.; Mancinelli, G.; Dobroslavić, T.; Bartulović, V.; Matić-Skoko, S.; Glamuzina, B. Population Dynamics and Reproduction of Mediterranean Green Crab Carcinus aestuarii in Parila Lagoon (Neretva Estuary, Adriatic Sea, Croatia) as Fishery Management Tools. Mar. Coast. Fish. 2017, 9, 260–270. [Google Scholar] [CrossRef] [Green Version]
- Horsford, I.; Simon, H.; Archibald, M.; Webber, J.; Joseph, T. Biology, status, and current management of the Caribbean Spiny Lobster (Panulirus argus). Gulf Caribb. Fish. Inst. 2014, 66, 423–433. [Google Scholar]
- Kampouris, T.E.; Koutsoubas, D.; Milenkova, D.; Economidis, G.; Tamvakidis, S.; Batjakas, I.E. New data on the biology and fisheries of the threatened Palinurus elephas (Fabricius, 1787) (Decapoda, achelata, Palinuridae) from the north-west Aegean sea, Greece. Water 2020, 12, 2390. [Google Scholar] [CrossRef]
- French McCay, D.; Gibson, M.; Cobb, J. Scaling restoration of American lobsters: Combined demographic and discounting model for an exploited species. Mar. Ecol. Prog. Ser. 2003, 264, 177–196. [Google Scholar] [CrossRef] [Green Version]
- Steinback, S.R.; Allen, R.B.; Thunberg, E. The benefits of rationalization: The case of the American lobster fishery. Mar. Resour. Econ. 2008, 23, 37–63. [Google Scholar] [CrossRef]
- Sheehy, M.R.J.; Bannister, R.C.A.; Wickins, J.F.; Shelton, P.M.J. New perspectives on the growth and longevity of the European lobster (Homarus gammarus). Can. J. Fish. Aquat. Sci. 1999, 56, 1904–1915. [Google Scholar] [CrossRef]
- Radhakrishnan, E.V.; Thangaraja, R.; Vijayakumaran, M. Ontogenetic changes in morphometry of the spiny lobster, Panulirus homarus homarus (Linnaeus, 1758) from southern Indian coast. J. Mar. Biol. Assoc. India 2015, 57, 5–13. [Google Scholar] [CrossRef] [Green Version]
- Ball, B.; Linnane, A.; Munday, B.; Browne, R.; Mercer, J.P. The effect of cover on in situ predation in early benthic phase European lobster Homarus gammarus. J. Mar. Biol. Assoc. U. K. 2001, 81, 639–642. [Google Scholar] [CrossRef]
- Landers, D.F.; Keser, M.; Saila, S.B. Changes in female lobster (Homarus americanus) size at maturity and implications for the lobster resource in Long Island Sound, Connecticut. Mar. Freshw. Res. 2001, 52, 1283–1290. [Google Scholar] [CrossRef]
- Woodruff, J. Stock Assessment—An Evaluation of the Minimum Landing Size and the Pot Limitation Byelaw in the Northumberland Lobster Fishery; NIFCA: Northumberland, UK, 2017. [Google Scholar]
- Baudron, A.R.; Needle, C.L.; Rijnsdorp, A.D.; Tara Marshall, C. Warming temperatures and smaller body sizes: Synchronous changes in growth of North Sea fishes. Glob. Chang. Biol. 2014, 20, 1023–1031. [Google Scholar] [CrossRef] [Green Version]
- Le Bris, A.; Pershing, A.J.; Gaudette, J.; Pugh, T.L.; Reardon, K.M. Multi-scale quantification of the effects of temperature on size at maturity in the American lobster (Homarus americanus). Fish. Res. 2017, 186, 397–406. [Google Scholar] [CrossRef]
- The Shellfish Team. Lobster (Homarus gammarus) Cefas Stock Status Report 2014; Center for Environment Fisheries & Aquaculture Science: Lowestoft, UK, 2014.
- Haarr, M.L.; Sainte-Marie, B.; Comeau, M.; Tremblay, M.J.; Rochette, R. Female American lobster (Homarus americanus) size-at-maturity declined in Canada during the 20th and early 21st centuries. Can. J. Fish. Aquat. Sci. 2018, 75, 908–924. [Google Scholar] [CrossRef] [Green Version]
- Grabowski, J.H.; Clesceri, E.J.; Baukus, A.J.; Gaudette, J.; Weber, M.; Yund, P.O. Use of Herring Bait to Farm Lobsters in the Gulf of Maine. PLoS ONE 2010, 5, e10188. [Google Scholar] [CrossRef]
- Turner, R.; Hardy, M.; Green, J.; Polunin, N. Defining the Northumberland Lobster Fishery; Newcastle University: London, UK, 2009. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Relationship | Intercept a (95% CI) | Slope b (95% CI) | R2 | N | |
|---|---|---|---|---|---|---|
| W vs. CL | Female | W = 0.0004568 × CL2.561 | 0.0004568 (0.003052, 0.006084) | 2.561 (2.492, 2.630) * | 0.919 | 447 |
| Male | W = 0.001311 × CL2.822 | 0.001311 (0.000791, 0.001831) | 2.822 (2.740, 2.904) * | 0.890 | 489 | |
| TL vs. CL | Female | TL = 32.893+2.617 × CL | 32.893 (28.098, 37.689) | 2.617 (2.571, 2.662) * | 0.966 | 447 |
| Male | TL = 42.351+2.467 × CL | 42.351 (37.228, 47.474) | 2.467 (2.419, 2.516) * | 0.953 | 489 | |
| W vs. TL | Female | W = 2.243 × 10−5 × CL3.004 | 0.000022431 (0.0000155, 0.0000293) | 3.004 (2.952, 3.096) * | 0.966 | 447 |
| Male | W = 2.287 × 10−6 × CL3.408 | 0.0000022887 (0.00000151, 0.00000307) | 3.408 (3.350, 3.466) * | 0.958 | 489 | |
| AW vs. CL | Female | AW = −19.327 + 0.724 × CL | −19.327 (−21.133, −17.520) | 0.724 (0.707, 0.742) * | 0.942 | 422 |
| Male | AW = 3.211 + 0.439 × CL | 3.211 (2.089, 4.333) | 0.439 (0.428, 0.449) * | 0.934 | 450 |
| Sex | Intercept a (95% CI) | Slope b (95% CI) | R2 | N | t | p | Allometry | |
|---|---|---|---|---|---|---|---|---|
| Log W vs. Log CL | Female | −3.056 (−3.201, −2.918) | 2.909 (2.839, 2.981) | 0.963 | 447 | 3.494 | <0.001 | Negative |
| Male | −3.134 (−3.295, −2.972) | 2.943 (2.862,3.025) | 0.955 | 489 | 2.017 | 0.044 | Negative | |
| Log TL vs. Log CL | Female | 0.636 (0.596, 0.676) | 0.917 (0.897, 0.937) | 0.974 | 447 | 11.772 | <0.001 | Negative |
| Male | 0.688 (0.646, 0.729) | 0.887 (0.866, 0.908) | 0.966 | 489 | 15.271 | <0.001 | Negative | |
| Log W vs. Log TL | Female | −5.073 (−5.202, −4.943) | 3.172 (3.120, 3.224) | 0.978 | 447 | 7.784 | <0.001 | Positive |
| Male | −5.419 (−5.561, −5.270) | 3.320 (3.259, 3.377) | 0.975 | 489 | 13.436 | <0.001 | Positive | |
| Log AW vs. Log CL | Female | −0.947 (−1.013, 0.884) | 1.334 (1.303, 1.368) | 0.953 | 422 | 23.570 | <0.001 | Positive |
| Male | −0.079 (−0.328, 0.229) | 0.976 (0.951, 1.000) | 0.944 | 450 | 2.223 | 0.027 | Negative |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavičić, M.; Matić-Skoko, S.; Vrdoljak, D.; Vujević, A. Population Characteristics of the European Lobster, Homarus gammarus in the Adriatic Sea: Implications for Sustainable Fisheries Management. Water 2021, 13, 1072. https://doi.org/10.3390/w13081072
Pavičić M, Matić-Skoko S, Vrdoljak D, Vujević A. Population Characteristics of the European Lobster, Homarus gammarus in the Adriatic Sea: Implications for Sustainable Fisheries Management. Water. 2021; 13(8):1072. https://doi.org/10.3390/w13081072
Chicago/Turabian StylePavičić, Mišo, Sanja Matić-Skoko, Dario Vrdoljak, and Ante Vujević. 2021. "Population Characteristics of the European Lobster, Homarus gammarus in the Adriatic Sea: Implications for Sustainable Fisheries Management" Water 13, no. 8: 1072. https://doi.org/10.3390/w13081072
APA StylePavičić, M., Matić-Skoko, S., Vrdoljak, D., & Vujević, A. (2021). Population Characteristics of the European Lobster, Homarus gammarus in the Adriatic Sea: Implications for Sustainable Fisheries Management. Water, 13(8), 1072. https://doi.org/10.3390/w13081072