
The Toxic Effects of Lindane via the Impact of pH and Dissolved Organic Matter on Microcystis aeruginosa

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
2.2. Algal Cell Density Measurements
2.3. Acute and Chronic Toxicity Test
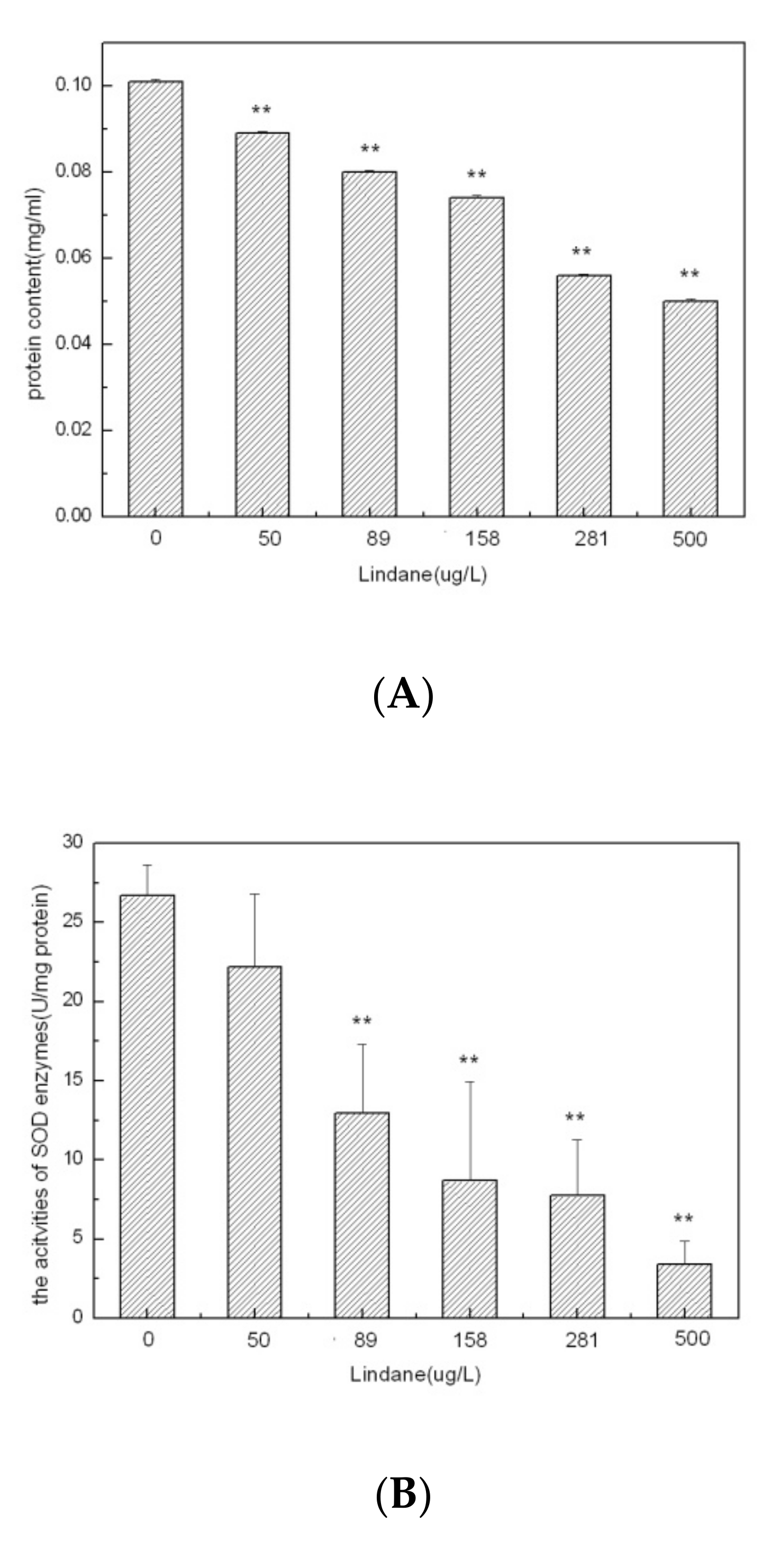
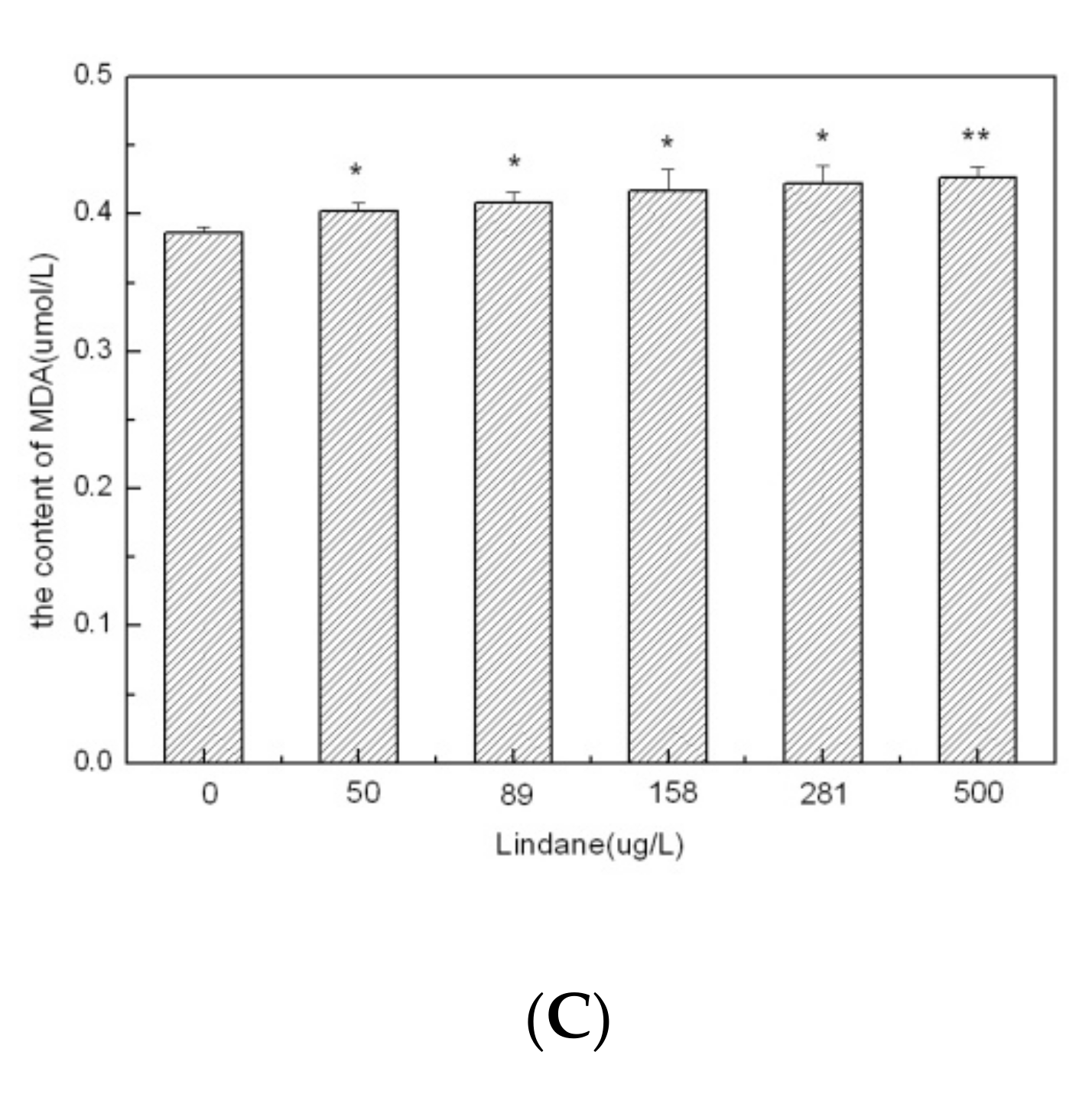
2.4. Determination of the Protein, SOD and MDA Levels in Algal Cells
2.5. Determination of Lindane Concentration in Algal Cells
2.6. The Influence of pH and DOM
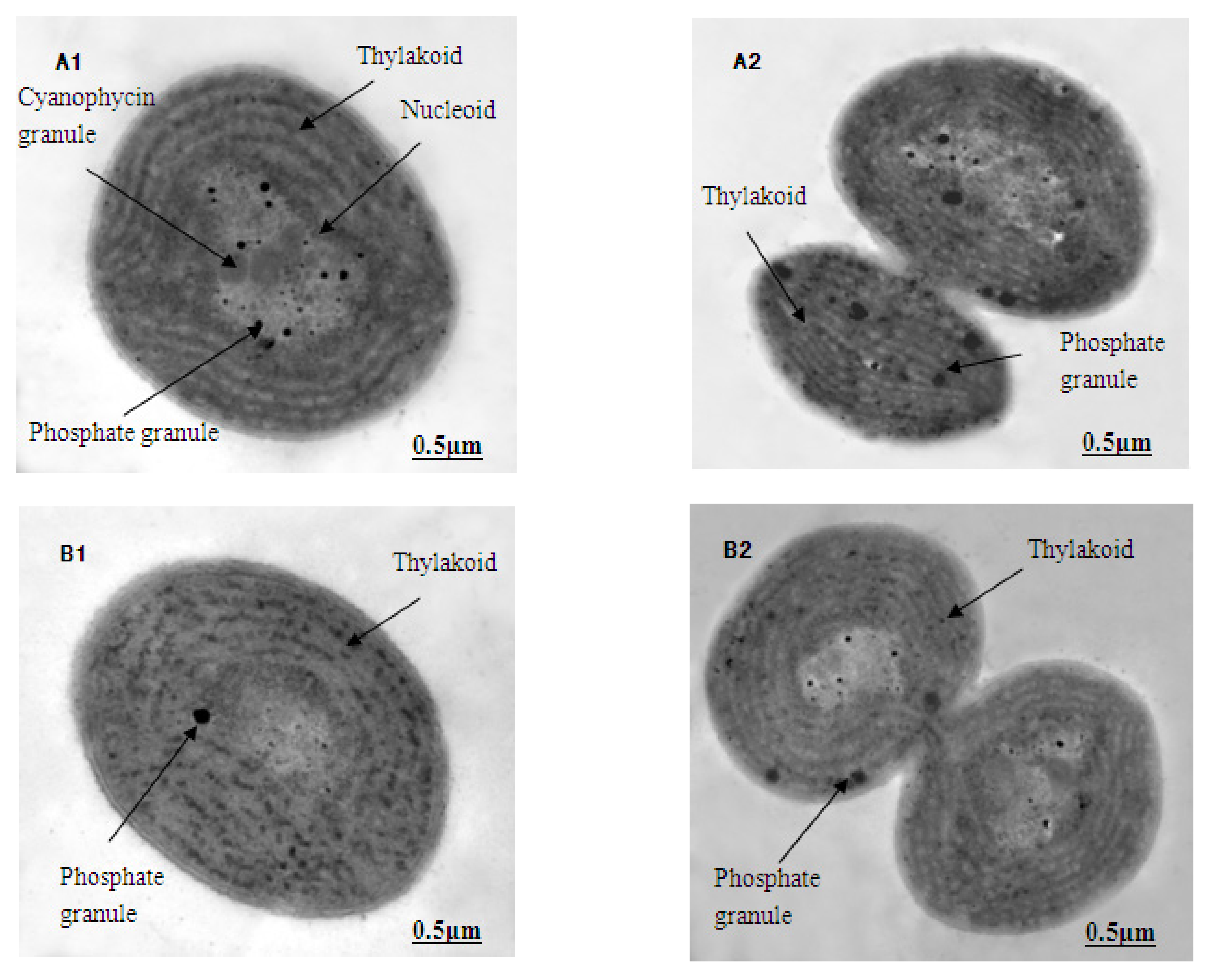
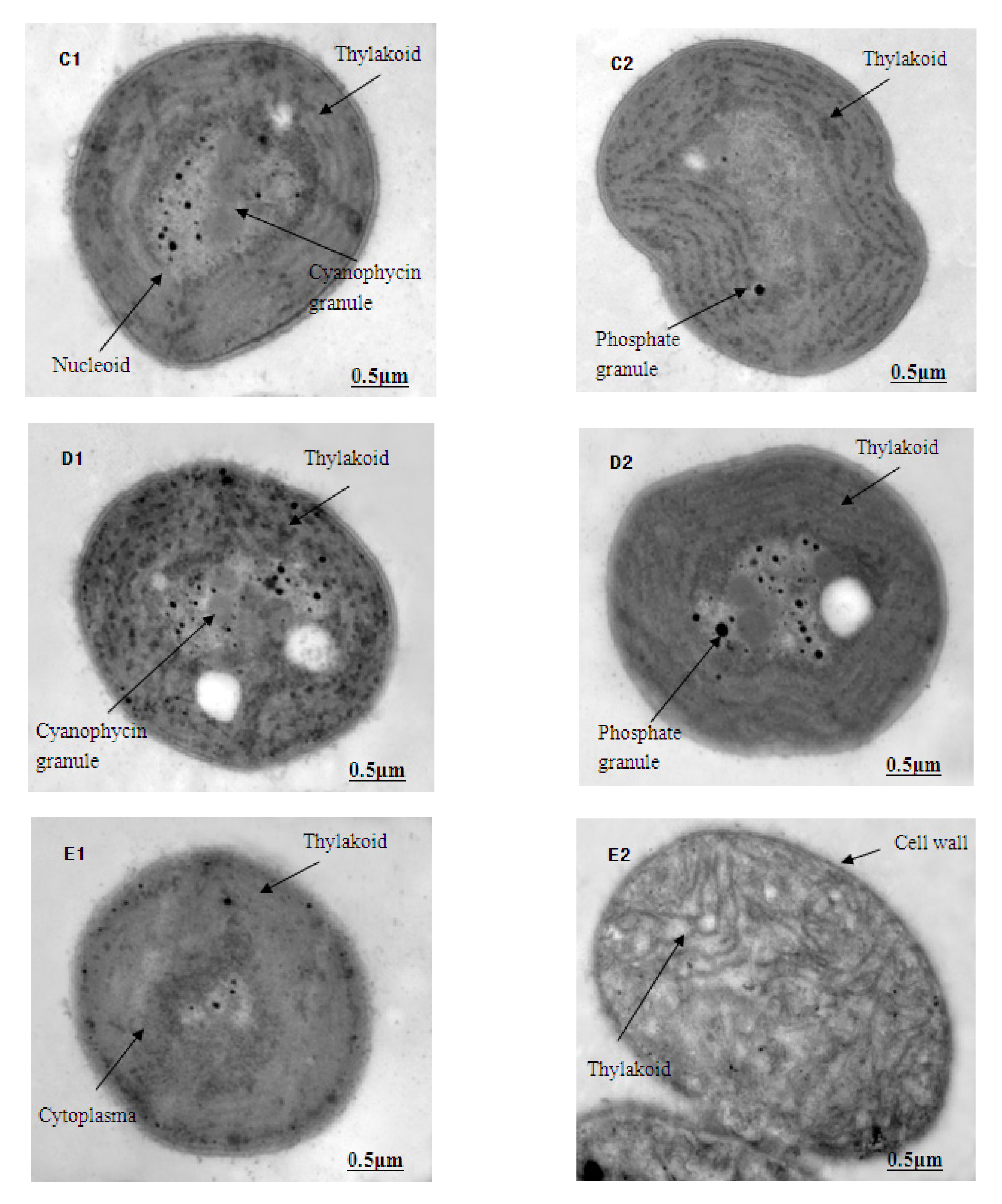
2.7. The Ultrastructural Observations of M. aeruginosa
2.8. Statistical Analysis
3. Results and Discussion
3.1. Growth Inhibition of M. aeruginosa by Lindane
3.2. Cellular Effects of Lindane Exposure in M. aeruginosa
3.3. Bioaccumulation of Lindane in M. aeruginosa
3.4. The Influence of pH and DOM on Lindane Toxicity
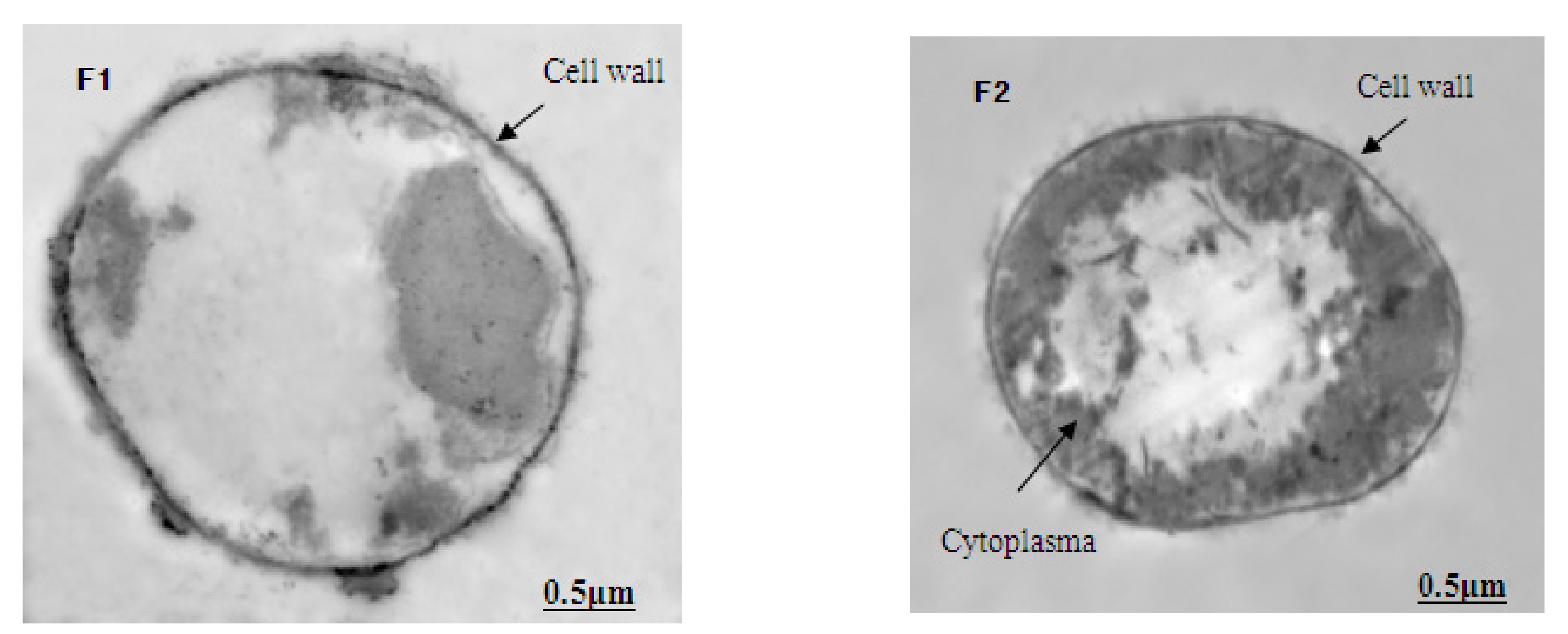
3.5. The Ultrastructure of M. aeruginosa
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Saez, J.M.; Benimeli, C.S.; Amoroso, M.J. Lindane removal by pure and mixed cultures of immobilized actinobacteria. Chemosphere 2012, 89, 982–987. [Google Scholar] [CrossRef] [PubMed]
- Bidlan, R.; Afsar, M.; Manonmani, H.K. Bioremediation of HCH-contaminated soil: Elimination of inhibitory effects of the insecticide on radish and green gram seed germination. Chemosphere 2004, 56, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Phillips, T.M.; Seech, A.G.; Lee, H.; Trevors, J.T. Biodegradation of hexachlorocyclohexane (HCH) by microorganisms. Biodegradation 2005, 16, 363–392. [Google Scholar] [CrossRef] [PubMed]
- Arnot, J.A.; Gobas, F.A.P.C. A review of bioconcentration factor (BCF) and bioaccumulation factor (BAF) assessments for organic chemicals in aquatic organisms. Environ. Rev. 2006, 14, 257–297. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Carvalho, P.N.; Rodrigues, P.N.R.; Basto, M.; Vasconcelos, M. Organochlorine pesticides levels in Portuguese coastal areas. Chemosphere 2009, 75, 595–600. [Google Scholar] [CrossRef]
- Cid, F.D.; Anton, R.I.; Caviedes-Vidal, E. Organochlorine pesticide contamination in three bird species of the Embalse La Florida water reservoir in the semiarid midwest of Argentina. Sci. Total Environ. 2007, 385, 86–96. [Google Scholar] [CrossRef]
- Concha-Grana, E.; Turnes-Carou, M.I.; Muniategui-Lorenzo, S.; Lopez-Mahia, P.; Prada-Rodriguez, D.; Fernandez-Fernandez, E. Evaluation of HCH isomers and metabolites in soils, leachates, river water and sediments of a highly contaminated area. Chemosphere 2006, 64, 588–595. [Google Scholar] [CrossRef]
- Vega, F.A.; Covelo, E.F.; Andrade, M.L. Accidental organochlorine pesticide contamination of soil in Porrino, Spain. J. Environ. Qual. 2007, 36, 272–279. [Google Scholar] [CrossRef]
- Kobraei, M.E.; White, D.S. Effects of 2, 4-dichlorophenoxyacetic acid on Kentucky algae: Simultaneous laboratory and field toxicity testings. Arch. Environ. Contam. Toxicol. 1996, 31, 571–580. [Google Scholar] [CrossRef]
- Olvera-Ramirez, R.; Coria-Cedillo, M.; Canizares-Villanueva, R.O.; Martinez-Jeronimo, F.; Ponce-Noyola, T.; Rios-Leal, E. Growth evaluation and bioproducts characterization of Calothrix sp. Bioresour. Technol. 2000, 72, 121–124. [Google Scholar] [CrossRef]
- Hoekstra, J.A. Estimation of the LC50, a review. Environmetrics 1991, 2, 139–152. [Google Scholar] [CrossRef]
- Saker, M.L.; Neilan, B.A. Varied diazotrophies, morphologies, and toxicities of genetically similar isolates of Cylindrospermopsis raciborskii (nostocalss, cyanophyceae) from northern Australia. Appl. Environ. Microbiol. 2001, 67, 1839–1845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Pollio, A.; Pinto, G.; Ligrone, R.; Aliotta, G. Effects of the potential allelochemical α-asarone on growth, physiology and ultrastructure of two unicellular green algae. J. Appl. Phycol. 1993, 5, 395–403. [Google Scholar] [CrossRef]
- Hans, R.K.; Singh, J.; Viswanathan, P.N.; Joshi, P.C. Effect of lindane on the growth and metabolic activities of cyanobacteria. Ecotoxicol. Environ. Saf. 2001, 48, 219–221. [Google Scholar]
- Scandalios, J.G. Oxygen stress and superoxide dismutases. Plant Physiol. 1993, 101, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Chaoui, A.; Mazhoudi, S.; Ghorbal, M.H.; El Ferjani, E. Cadmium and zinc induction of lipid peroxidation and effects on antioxidant enzyme activities in bean (Phaseolus vulgaris L.). Plant Sci. 1997, 127, 139–147. [Google Scholar] [CrossRef]
- Thybaud, E.; Le Bras, S. Absorption and elimination of lindane byAsellus aquaticus (Crustacea, Isopoda). Bull. Environ. Contam. Toxicol. 1988, 40, 731–735. [Google Scholar] [CrossRef]
- Vigano, L.; Galassi, S.; Gatto, M. Factors affecting the bioconcentration of hexachlorocyclohexanes in early life stages of Oncorhynchus mykiss. Environ. Toxicol. Chem. 1992, 11, 535–540. [Google Scholar] [CrossRef]
- Jin, X.C.; Li, Z.C.; Zheng, S.F.; Yang, S.W.; Hu, X.Z.; Chu, Z.S. Studies on the Growth Characteristics of Microcystis aeruginosa. Res. Environ. Sci. S 2004, 17, 52–54, 61. [Google Scholar]
- Ngabe, B.; Bidleman, T.F.; Falconer, R.L. Base hydrolysis of .alpha.- and .gamma.-hexachlorocyclohexanes. Environ. Sci. Technol. 1993, 27, 1930–1933. [Google Scholar] [CrossRef]
- Saleh, F.Y.; Dickson, K.L.; Rodgers, J.H. Fate of lindane in the aquatic environment: Rate constants of physical and chemical processes. Environ. Toxicol. Chem. 1982, 1, 289–297. [Google Scholar] [CrossRef]
- Liu, X.M.; Peng, P.A.; Huang, W.L.; Sheng, G.Y.; Fu, J.M. Influence of acidity on Lindane hydrolysis. China Environ. Sci. 2002, 22, 485–489. [Google Scholar]
- Boggs, S.; Livermore, D.G.; Seitz, M.G. Humic macromolecules in natural waters. J. Macromol. Sci.-Rev. Macromol. Chem. Phys. 1985, 25, 599–657. [Google Scholar] [CrossRef]
- Edwards, R.; Cole, D.J. Glutathione Transferases in Wheat (Triticum) Species with Activity toward Fenoxaprop-Ethyl and Other Herbicides. Pestic. Biochem. Physiol. 1996, 54, 96–104. [Google Scholar] [CrossRef]
- Thevenot, M.; Dousset, S.; Hertkorn, N.; Schmitt-Kopplin, P.; Andreux, F. Interactions of diuron with dissolved organic matter from organic amendments. Sci. Total Environ. 2009, 407, 4297–4302. [Google Scholar] [CrossRef]
- Haitzer, M.; Hoss, S.; Traunspurger, W.; Steinberg, C. Effects of dissolved organic matter (DOM) on the bioconcentration of organic chemicals in aquatic organisms—A review. Chemosphere 1998, 37, 1335–1362. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | HA (mg/L) | The Regression Equation | R2 | EC50 (μg/L) | LOEC (μg/L) | NOEC (μg/L) | MATC (μg/L) |
|---|---|---|---|---|---|---|---|
| 5 | 0 | y = 0.6766 x + 0.9149 | 0.9923 | 419 | 121 | 63 | 87 |
| 7 | y = 0.6435 x + 1.0806 | 0.9964 | 442 | 120 | 60 | 85 | |
| 9 | y = 0.6607 x + 0.9533 | 0.9787 | 457 | 128 | 66 | 92 | |
| 7 | 2.5 | y = 0.6330 x + 1.0018 | 0.9988 | 554 | 147 | 73 | 104 |
| 5 | y = 0.6330 x + 0.8135 | 0.9966 | 745 | 198 | 99 | 140 | |
| 7.5 | y = 0.6140 x + 0.8358 | 0.9756 | 882 | 225 | 110 | 157 | |
| 10 | y = 0.6505 x + 0.5610 | 0.9813 | 920 | 238 | 129 | 175 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, X.; Jiang, Y.; Xian, Q. The Toxic Effects of Lindane via the Impact of pH and Dissolved Organic Matter on Microcystis aeruginosa. Water 2022, 14, 1915. https://doi.org/10.3390/w14121915
Deng X, Jiang Y, Xian Q. The Toxic Effects of Lindane via the Impact of pH and Dissolved Organic Matter on Microcystis aeruginosa. Water. 2022; 14(12):1915. https://doi.org/10.3390/w14121915
Chicago/Turabian StyleDeng, Xiru, Yingnan Jiang, and Qiming Xian. 2022. "The Toxic Effects of Lindane via the Impact of pH and Dissolved Organic Matter on Microcystis aeruginosa" Water 14, no. 12: 1915. https://doi.org/10.3390/w14121915
APA StyleDeng, X., Jiang, Y., & Xian, Q. (2022). The Toxic Effects of Lindane via the Impact of pH and Dissolved Organic Matter on Microcystis aeruginosa. Water, 14(12), 1915. https://doi.org/10.3390/w14121915