
Tolerance and Growth of Adult Schizothorax prenanti Exposed and Re-Exposed to Supersaturated Gas Downstream of a Dam




Abstract
:1. Introduction
2. Materials and Methods
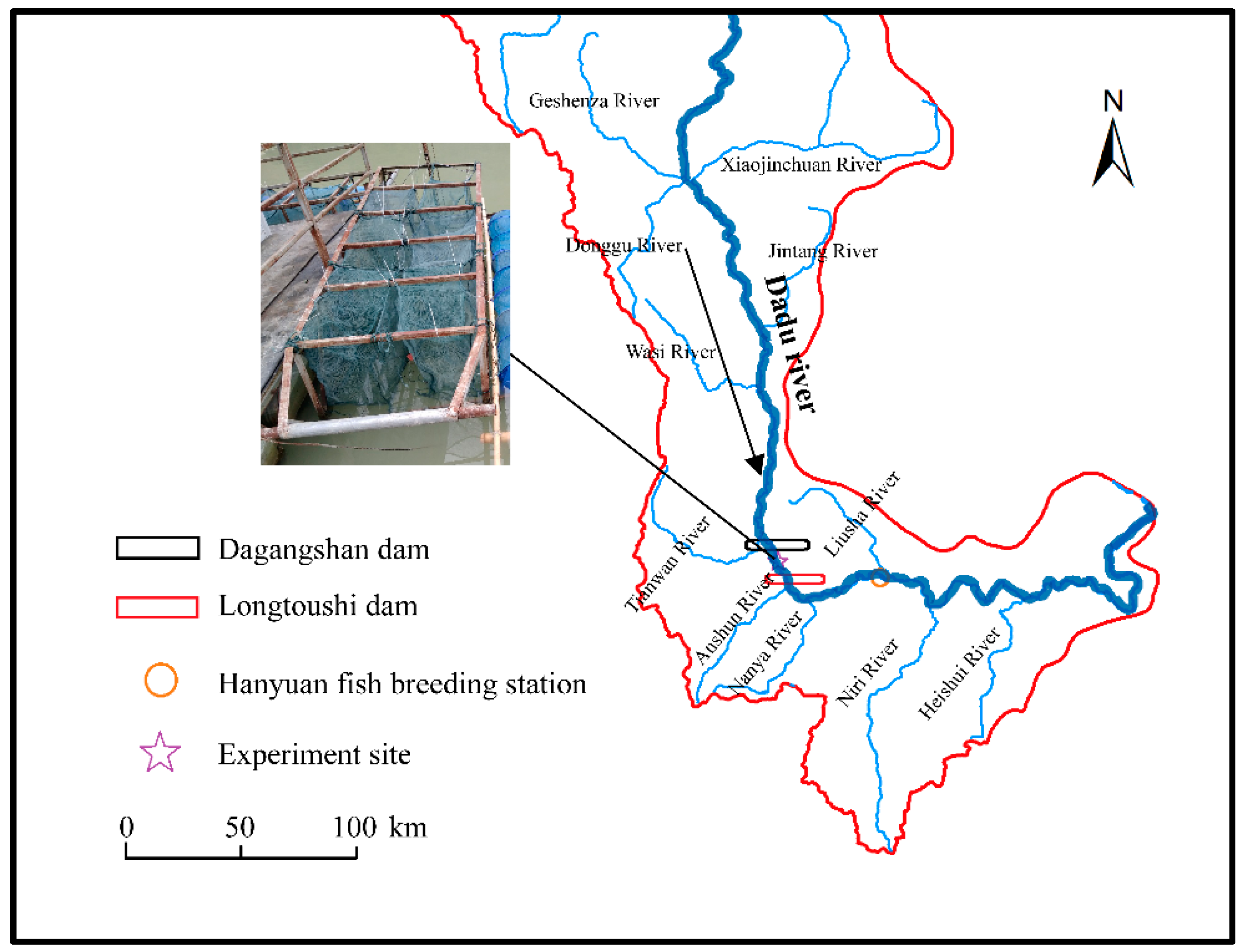
2.1. Study Site and Fish Acclimation
2.2. Experiment Design
2.3. Statistical Analysis
3. Results
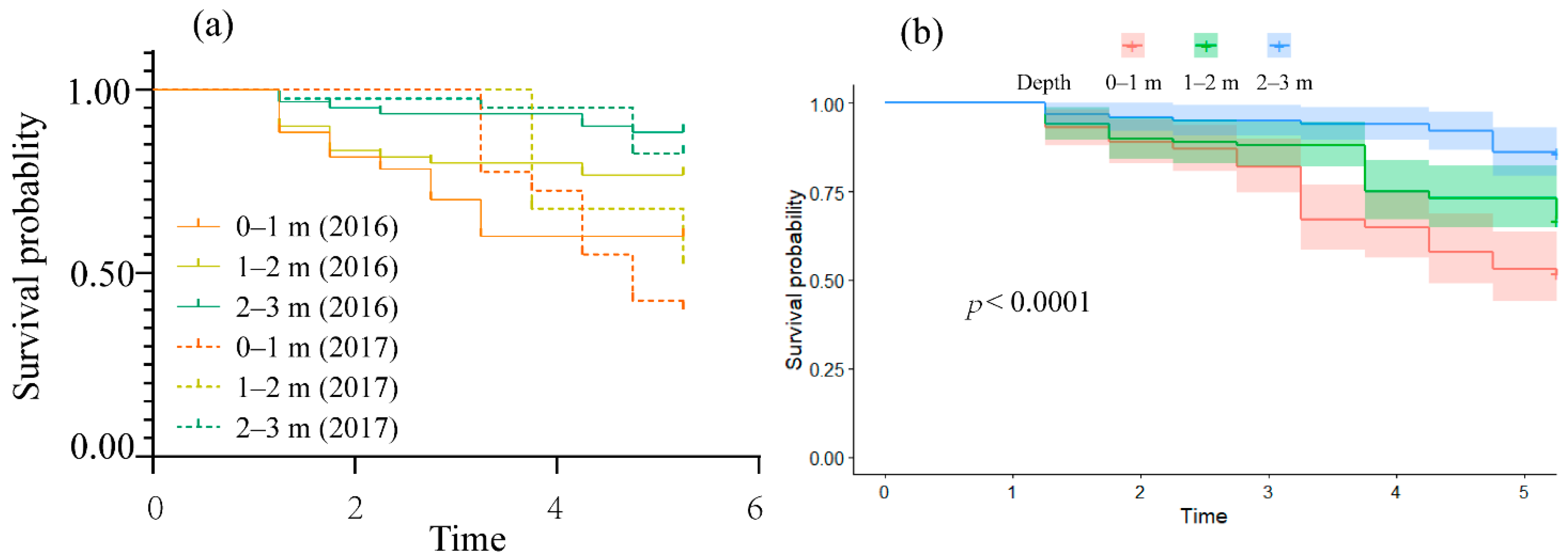
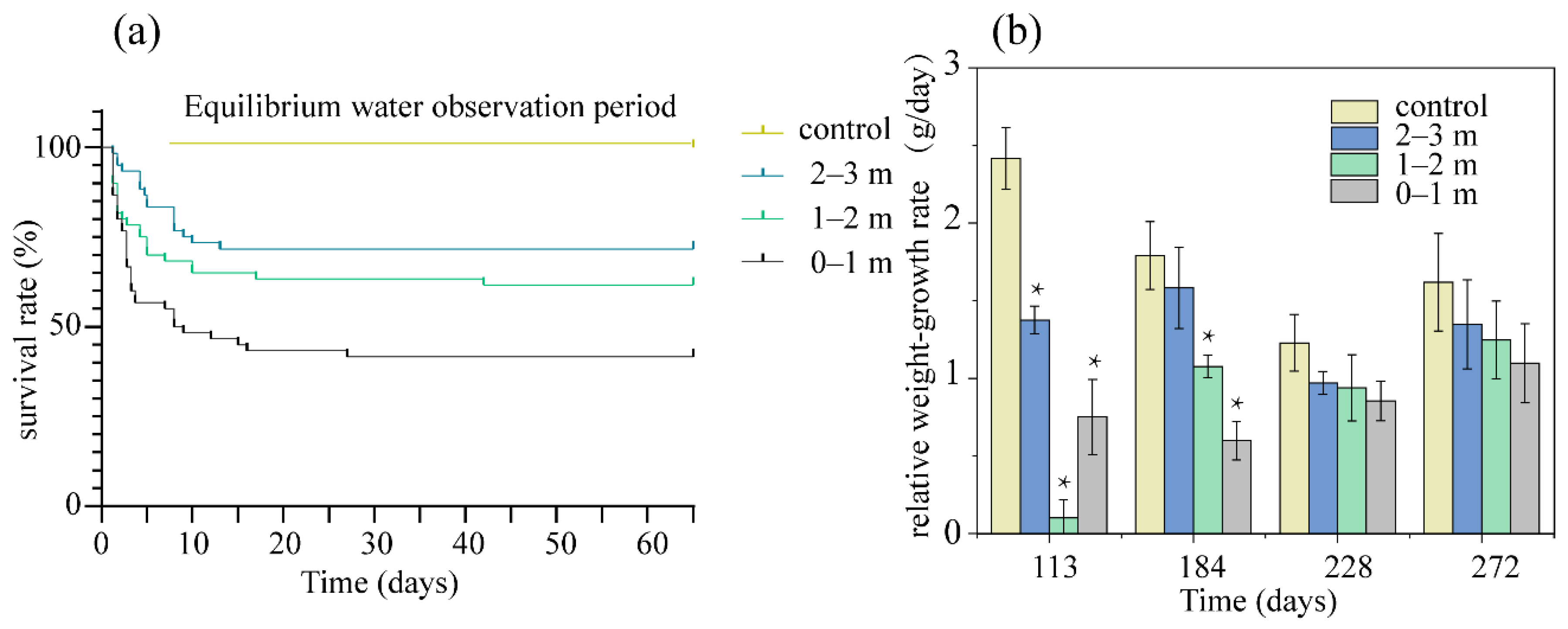
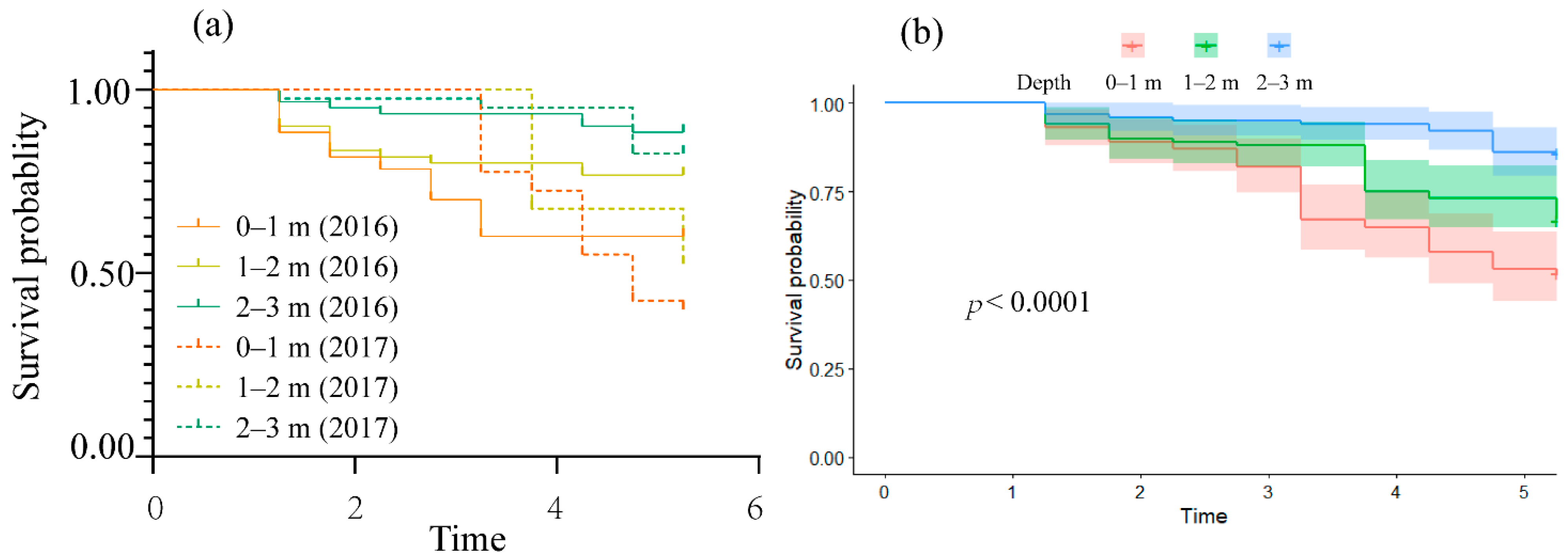
3.1. Survival Characteristics of Fish Exposed to TDG Supersaturation
3.2. Growth Characteristics of Fish after Exposed to TDG Supersaturation
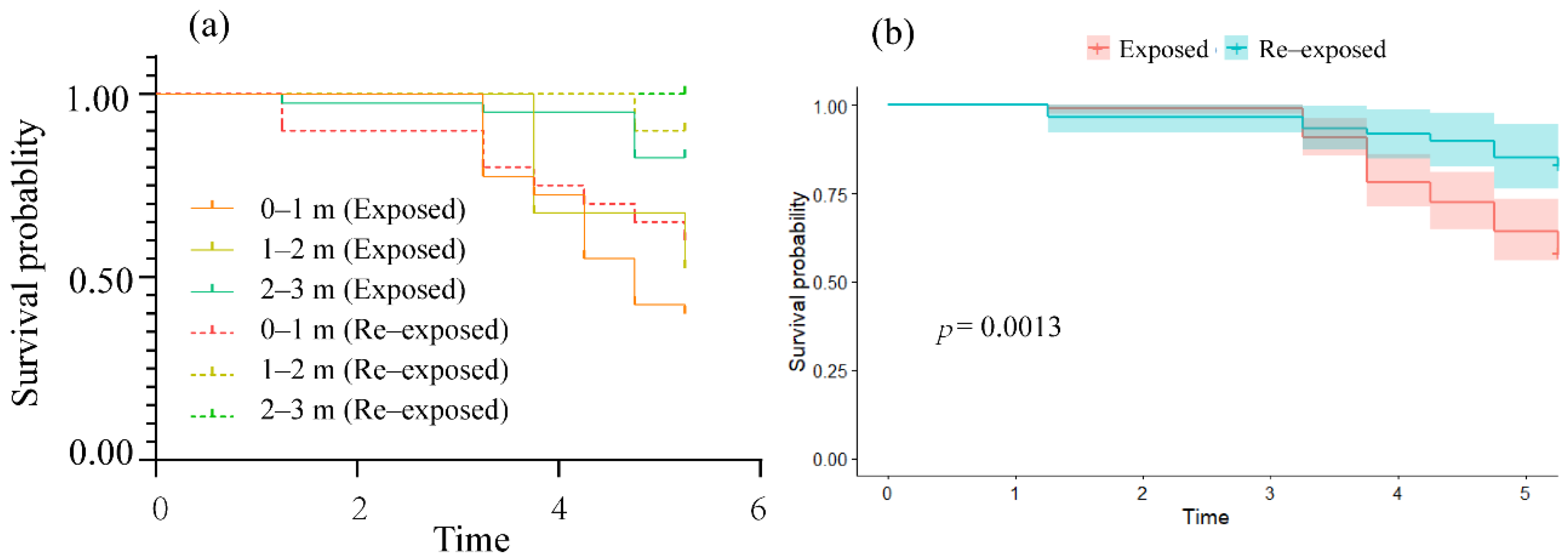
3.3. Survival Characteristics of Fish Re-Exposed to TDG Supersaturation
4. Discussion
4.1. Effect of Water Depth on Fish Survival
4.2. Growth of Fish after Exposure to TDG Supersaturation
4.3. Survival of Fish Re-Exposed to TDG Supersaturation
4.4. Future Study
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuan, Y.; Wei, Q.; Yuan, Q.; Wang, Y.; Liang, R.; Li, K.; Zhu, D.Z. Impact of TDG supersaturation on native fish species under different hydropower flood discharge programs. Aquat. Toxicol. 2021, 237, 105898. [Google Scholar] [CrossRef]
- Xue, S.; Li, K.; Liang, R.; Cao, L.; Wang, Y.; Li, Y.; Feng, J. In situ study on the impact of total dissolved gas supersaturation on endemic fish in the Upper Yangtze River. River Res. Appl. 2019, 35, 1511–1519. [Google Scholar] [CrossRef]
- Cao, C.; Deng, Y.; Yin, Q.; Li, N.; Liu, X.; Shi, H.; Yang, Y.; Xu, L. Effects of continuous acute and intermittent exposure on the tolerance of juvenile yellow catfish (Pelteobagrus fulvidraco) in total dissolved gas supersaturated water. Ecotoxicol. Environ. Saf. 2020, 201, 110855. [Google Scholar] [CrossRef] [PubMed]
- Mesa, M.G.; Weiland, L.K.; Maule, A.G. Progression and severity of gas bubble trauma in juvenile salmonids. Am. Fish. Soc. 2000, 129, 174–185. [Google Scholar] [CrossRef]
- Tsai, J.Y.; Felt, S.A.; Bouley, D.M.; Green, S.L. Acute and chronic outcomes of gas-bubble disease in a colony of African Clawed Frogs (Xenopus laevis). Comp. Med. 2017, 67, 4–10. [Google Scholar] [PubMed]
- Pleizier, N.K.; Algera, D.; Cooke, S.J.; Brauner, C.J. A meta-analysis of gas bubble trauma in fish. Fish Fish. 2020, 21, 1175–1194. [Google Scholar] [CrossRef]
- Yuan, Q.; Wang, Y.; Liang, R.; Feng, J.; Li, Y.; An, R.; Li, K. Field observations of the lethality characteristics of endangered and endemic fish under the stress of total dissolved gas supersaturation. River Res. Appl. 2021, 37, 1156–1167. [Google Scholar] [CrossRef]
- Geist, D.R.; Linley, T.J.; Cullinan, V.; Deng, Z. The effects of total dissolved gas on chum salmon fry survival, growth, gas bubble disease, and seawater tolerance. N. Am. J. Fish. Manag. 2013, 33, 200–215. [Google Scholar] [CrossRef]
- Meekin, T.K.; Turner, B.K. Tolerance of salmonid eggs, juveniles and squawfish to supersaturated nitrogen. In Nitrogen Supersaturation Investigations in the Mid-Columbia River; Washington Department of Fisheries Technical Report: Olympia, WA, USA, 1974; Volume 12, pp. 75–126. [Google Scholar]
- Harvey, H.H.; Cooper, A.C. Origin and treatment of a supersaturated river water. Prog. Rep. 1962, 9, 7. [Google Scholar]
- Nebeker, A.V.; Andros, J.D.; McCrady, J.K.; Stevens, D.G. Survival of steelhead trout (Salmo gairdneri) eggs, embryos, and fry in air-supersaturated water. J. Fish. Board Can. 1978, 35, 261–264. [Google Scholar] [CrossRef]
- Weitkamp, D.E.; Katz, M. A review of dissolved gas supersaturation literature. Trans. Am. Fish. Soc. 1980, 109, 659–702. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, Y.; Zhang, R.; Ouyang, S.; Wang, X.; Liao, X. Integrated optimization of hydroelectric energy in the upper and middle Yangtze River. Renew. Sust. Energ. Rev. 2015, 45, 481–512. [Google Scholar] [CrossRef]
- Ding, R. (Ed.) The Fishes of Sichuan, China; Sichuan Publishing House of Science and Technology: Chengdu, China, 1194; p. 641. (In Chinese) [Google Scholar]
- Liu, J. A quantitative analysis on threat and priority of conservation order of the endemic fishes in upper reaches of the Yangtze River. Environ. Sci. Sin. 2004, 24, 395–399. [Google Scholar]
- Pleizier, N.K.; Nelson, C.; Cooke, S.J.; Brauner, C.J. Understanding gas bubble trauma in an era of hydropower expansion: How do fish compensate at depth? Can. J. Fish. Aquat. Sci. 2020, 77, 556–563. [Google Scholar] [CrossRef]
- Elston, R.; Colt, J.; Abernethy, S.; Maslen, W. Gas bubble reabsorption in Chinook salmon: Pressurization effects. J. Aquat. Anim. Health 1997, 9, 317–321. [Google Scholar] [CrossRef]
- Algera, D.A.; Kamal, R.; Ward, T.D.; Pleizier, N.K.; Brauner, C.J.; Crossman, J.A.; Leake, A.; Zhu, D.Z.; Power, M.; Cooke, S.J. Exposure risk of fish downstream of a hydropower facility to supersaturated total dissolved gas. Water Resour. Res. 2021, 58, e2021WR031887. [Google Scholar] [CrossRef]
- Wang, Y.; Li, K.; Li, J.; Li, R.; Deng, Y. Tolerance and avoidance characteristics of Prenant’s schizothoracin Schizothorax prenanti to total dissolved gas supersaturated water. N. Am. J. Fish. Manag. 2015, 35, 827–834. [Google Scholar] [CrossRef]
- Brege, D.A.; Absolon, R.F.; Graves, R.J. Seasonal and diel passage of juvenile salmonids at John Day Dam on the Columbia River. N. Am. J. Fish. Manage. 1996, 16, 659–665. [Google Scholar] [CrossRef]
- Reddin, D.G.; Downton, P.; Friedland, K.D. Diurnal and nocturnal temperatures for Atlantic salmon postsmolts (Salmo salar L.) during their early marine life. Fish. Bull. 2006, 104, 415–428. [Google Scholar]
- Davidsen, J.G.; Manel-La, N.P.; Økland, F.; Diserud, O.H.; Thorstad, E.B.; Finstad, B.; Sivertsgård, R.; McKinley, R.S.; Rikardsen, A.H. Changes in swimming depths of Atlantic salmon Salmo salar post-smolts relative to light intensity. J. Fish Biol. 2008, 73, 1065–1074. [Google Scholar] [CrossRef]
- Johnson, E.L.; Clabough, T.S.; Peery, C.A.; Bennett, D.H.; Bjornn, T.C.; Caudill, C.C.; Richmond, M.C. Estimating adult Chinook salmon exposure to dissolved gas supersaturation downstream of hydroelectric dams using telemetry and hydrodynamic models. River Res. Appl. 2007, 23, 963–978. [Google Scholar] [CrossRef]
- Beeman, J.W.; Maule, A.G. Migration depths of juvenile Chinook salmon and steelhead relative to total dissolved gas supersaturation in a Columbia River reservoir. Trans. Am. Fish. Soc. 2006, 135, 584–594. [Google Scholar] [CrossRef]
- Keefer, M.L.; Peery, C.A.; Ringe, R.R.; Bjornn, T.C. Regurgitation rates of intragastric radio transmitters by adult Chinook salmon and steelhead during upstream migration in the Columbia and Snake rivers. N. Am. J. Fish. Manag. 2004, 24, 47–54. [Google Scholar] [CrossRef]
- Keefer, M.L.; Caudill, C.C.; Peery, C.A.; Bjornn, T.C. Route selection in a large river during the homing migration of Chinook salmon (Oncorhynchus tshawytscha). Can. J. Fish. Aquat. Sci. 2006, 63, 1752–1762. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, J.; Zhao, J.; Peng, W.; Fu, Y.; Wang, Q.; Zhang, Y. A new method for calculating the downstream ecological flow of diversion-type small hydropower stations. Ecol. Indic. 2021, 125, 107530. [Google Scholar] [CrossRef]
- Wang, Y.; Li, Y.; An, R.; Li, K. Effects of total dissolved gas supersaturation on the swimming performance of two endemic fish species in the upper Yangtze River. Sci. Rep. 2018, 8, 10063. [Google Scholar] [CrossRef]
- Stephenson, J.R.; Gingerich, A.J.; Brown, R.S.; Pflugrath, B.D.; Deng, Z.; Carlson, T.J.; Langeslay, M.J.; Ahmann, M.L.; Johnson, R.L.; Seaburg, A.G. Assessing barotrauma in neutrally and negatively buoyant juvenile salmonids exposed to simulated hydro-turbine passage using a mobile aquatic barotrauma laboratory. Fish. Res. 2010, 106, 271–278. [Google Scholar] [CrossRef]
- Dawley, E.M.; Ebel, W.J. Effects of various concentrations of dissolved atmospheric gas on juvenile chinook salmon and steelhead trout. Fish. Bull. 1975, 73, 777–796. [Google Scholar]
- Gunnarsli, K.S.; Toften, H.; Mortensen, A. Effects of nitrogen gas supersaturation on growth and survival in juvenile Atlantic cod (Gadus morhua L.). Aquaculture 2008, 283, 175–179. [Google Scholar] [CrossRef]
- Liu, X.Q.; Li, K.F.; Du, J.; Li, J.; Li, R. Growth rate, catalase and superoxide dismutase activities in rock carp (Procypris rabaudi Tchang) exposed to supersaturated total dissolved gas. J. Zhejiang Univ. Sci. B 2011, 12, 909–914. [Google Scholar] [CrossRef]
- Jiang, W.; Wu, S. Biochemical responses and survival of rock carp (Procypris rabaudi) to total dissolved gas supersaturated water. Ichthyol. Res. 2015, 62, 171–176. [Google Scholar]
- Huchzermeyer, K.D.A. Clinical and pathological observations on Streptococcus sp. infection on South African trout farms with gas supersaturated water supplies. Onderstepoort J. Vet. Res. 2003, 70, 95–105. [Google Scholar] [PubMed]
- Gray, R.H.; Page, T.L.; Saroglia, M.G.; Bronzi, P. Comparative tolerance to gas supersaturated water of carp, Cyprinus carpio, and black bullhead, Ictalurus melas, from the USA and Italy. J. Fish Biol. 1982, 20, 223–227. [Google Scholar] [CrossRef]
- Dobzhansky, T. Genetics and the Origin of Species; Columbia University Press: New York, NY, USA, 1982. [Google Scholar]
- Grant, P.R.; Grant, B.R. The secondary contact phase of allopatric speciation in Darwin’s finches. Proc. Natl. Acad. Sci. USA 2009, 106, 20141–20148. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, Q.; Wang, Y.; Zhu, D.Z.; Liang, R.; Qin, L.; Li, R.; Ji, Q.; Li, K. River habitat assessment and restoration in high dam flood discharge systems with total dissolved gas supersaturation. Water Res. 2022, 221, 118833. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Li, K.; Wang, Y.; Liang, R.; Feng, J.; Yuan, Q.; Zhang, P.; Zhu, D.Z. Total dissolved gases induced tolerance and avoidance behaviors in pelagic fish in the Yangtze River, China. Ecotoxicol. Environ. Saf. 2021, 216, 112218. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Li, K.; Wang, Y.; Liang, R.; Feng, J.; Li, R.; Zhu, D.Z. Swimming performance of a pelagic species in the Yangtze River under different exposure modes of the total dissolved gas supersaturation. Conserv. Physiol. 2022, 10, coac047. [Google Scholar] [CrossRef]
- Weitkamp, D.E. Total Dissolved Gas Supersaturation Biological Effects, Review of Literature 1980–2007; Parametrix: Bellevue, WA, USA, 2008. [Google Scholar]
- Mathias, J.A.; Barica, J. Gas supersaturation as a cause of early spring mortality of stocked trout. Can. J. Fish. Aquat. Sci. 1985, 42, 268–279. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, N.; Lee, J.Y.; Kang, H.; Sung, M.; Yu, Y.B.; Kim, K.; Je, J.Y.; Kim, H.W.; Kang, J.C.; et al. Case report: Mass mortality of olive flounder (Paralichthys olivaceus) caused by acute gas bubble disease. J. Fish Pathol. 2021, 34, 255–259. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GBD Signs | 2016 | 2017 | ||||
|---|---|---|---|---|---|---|
| 0–1 m | 1–2 m | 2–3 m | 0–1 m | 1–2 m | 2–3 m | |
| Dorsal fin emphysema | 53% | 5% | 5% | 57% | 40% | 3% |
| Tail fin emphysema | 50% | 10% | 3% | 17% | 17% | 3% |
| Tail fin bleeding | 45% | 15% | 13% | 7% | 10% | 0% |
| Pectoral fin bleeding | 43% | 18% | 3% | 13% | 10% | 7% |
| Anal fin emphysema | 43% | 8% | 0% | 17% | 17% | 3% |
| Anal fin bleeding | 38% | 18% | 5% | 3% | 7% | 0% |
| Pelvic fin bleeding | 35% | 8% | 3% | 3% | 3% | 0% |
| Pectoral fin bleeding | 25% | 8% | 3% | 7% | 3% | 0% |
| Dorsal fin bleeding | 25% | 10% | 5% | 3% | 10% | 0% |
| Gill congestion | 3% | 0% | 3% | 3% | 3% | 0% |
| Cutaneous lesions | 0% | 0% | 0% | 3% | 0% | 0% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, Q.; Li, K.; Wang, Y.; Ji, Q.; Liang, R. Tolerance and Growth of Adult Schizothorax prenanti Exposed and Re-Exposed to Supersaturated Gas Downstream of a Dam. Water 2022, 14, 2501. https://doi.org/10.3390/w14162501
Yuan Q, Li K, Wang Y, Ji Q, Liang R. Tolerance and Growth of Adult Schizothorax prenanti Exposed and Re-Exposed to Supersaturated Gas Downstream of a Dam. Water. 2022; 14(16):2501. https://doi.org/10.3390/w14162501
Chicago/Turabian StyleYuan, Quan, Kefeng Li, Yuanming Wang, Qianfeng Ji, and Ruifeng Liang. 2022. "Tolerance and Growth of Adult Schizothorax prenanti Exposed and Re-Exposed to Supersaturated Gas Downstream of a Dam" Water 14, no. 16: 2501. https://doi.org/10.3390/w14162501
APA StyleYuan, Q., Li, K., Wang, Y., Ji, Q., & Liang, R. (2022). Tolerance and Growth of Adult Schizothorax prenanti Exposed and Re-Exposed to Supersaturated Gas Downstream of a Dam. Water, 14(16), 2501. https://doi.org/10.3390/w14162501