Abstract
Permafrost peatland is a unique ecosystem that represents a huge carbon terrestrial pool. Soil quality has a relatively high level of variation at small scale in this ecosystem and is closely related to the carbon cycle. To quantify this variability, we analyzed total organic carbon, total nitrogen, total phosphorus, nutrient, and element (sodium, magnesium, potassium, and calcium), and microbial respiration activities (microbial biomass carbon, soil basal respiration, metabolic, and microbial quotients) in the humus layer to deeper soil layer of 6 plots at small scale (meters). For all samples, the coefficients of variation (CV) values of total carbon concentrations were lowest and these values of sodium, magnesium, potassium, and calcium concentrations were higher than those of total carbon, nitrogen, and phosphorus concentrations. The largest variations of total phosphorus, ash, and Na were in the 10–20 cm layer with soil depths. The litter decomposition and water table may cause this variation at small-scale. The CV values of microbial respiration activities were largest compared with soil properties. There were different correlations between basal respiration and soil properties among the plots. Our results showed that soil properties and microbial respiration activities in permafrost peatland exhibits considerable variability at small scale. This variability indicates that sampling location and number are very important in peatland studies if we want to accurately estimate the biogeochemistry in a peatland.
1. Introduction
Permafrost peatland represents a huge global C (carbon) pool, which accumulate of organic matter, resulting from low microbial decomposition [1,2]. However, climate warming will be most severe in the boreal region [3]. The response of permafrost peatland to warming may include permafrost degradation, reduction of soil water table, increasing of peat aeration, and changes of plant species [4,5]. The features of permafrost peatlands are higher heterogeneity and permafrost degradation in peatlands results in peat surfaces collapse, hydrological process revision, and increased nutrient availability during thaw [6], which could cause more complex and heterogeneity at small-scale environment. The greenhouse gas flux, such as methane, strongly depends on these changes of small-scale environments [7,8]. Thus, the effects of small-scale heterogeneities on C cycle become a especial issue in northern peatlands and changes of small scale processes could have larger scale effects.
The spatial variability is apparent in peatlands at different scales. Soil quality also varies strongly from small scale to large scale, which affects the peatland functions [9,10]. For example, temperature and hydrology mainly influence peatland C dynamics at regional and global scales [11,12], while water-table level and litter quality are dominantly impacted factors at local scales in the peatland [13,14]. At an ecosystem scale, the vegetation composition of peatland is controlled by water level and pH [12]. The difference of environmental factors at small scale are common and lead to the changes of biogeochemical characteristics in peatland [15]. Vegetation composition, litter decomposition, soil moisture content, and topographic position cause these variabilities of soil properties [15,16]. Moreover, the ecological processes of plant litter inputs and decomposition, cation exchange mechanisms, and microbial decomposition and mineralization usually occur at small-scale [17,18,19]. Small-scale changes highlight local heterogeneities and can have larger-scale impacts in peatlands, which should be crucial for future predictions [7,20,21].
Although temperature and vegetation can be similar on a local scale in peatland, characteristics of permafrost, microtopography, and nutrient availability are often spatially variable at small scale [22,23]. These variabilities have an impact on substrate quality, biological processes, and microbial activities in peatland soil [24,25,26]. Therefore, field experiments should quantify the small-scale variability of soil quality and elevate the applicability of studies in peatlands [19,27]. If the study does not take into account the small-scale heterogeneity, results maybe not accurate [21]. Ulanowski and Branfireun (2013) found that under-sampling could lead to erroneous conclusions of pore-water biogeochemistry at small scale (1 m) in northern peatland [19]. Although assessment at small-scale is important, information is still unavailable on difference of soil quality at small-scale (meters) in permafrost peatlands.
To evaluate soil quality at small-scale (meters), we collected soil samples from 6 plots in the permafrost peatland of the Great Hing’an Mountains. With the exception of measurements of soil biochemical properties, we used soil basal respiration to identify potential differences in microbial activities, which is considered as a good indicator of soil quality [28]. The objectives of our study were: (1) to quantify the variations of the soil quality at small-scale (meters) through characterizing soil properties and soil microbial respiration activities from the humus layer to the deeper soil layer; (2) to evaluate the effects of the physical and chemical properties on soil basal respiration activities among all plots. The hypotheses of our study were evaluated: first, the variations of soil properties have similar trends with depth because all samples were collected in hummocks with similar vegetation types. Second, the factors that influenced soil microbial respiration would be different among all plots.
2. Materials and Methods
2.1. Study Area
The study was conducted in the permafrost peatland (52°55′59″ N, 122°46′26″ E) of the Great Hing’an Mountains, Heilongjiang Province, Northeast China. The study area is located at the southern boundary of the Eurasian continuous permafrost, which belongs to continental monsoon climate in cold temperate zone, the mean annual air temperature was about –5.5 °C, and the annual precipitation was about 400 mm [29]. The dominant species in this peatland are Betula fruticosa, Ledum palustre, Vaccinium uliginosum, Vaccinium uliginosum, Eriophorum vaginatum, and Sphagnum spp. The peatlands are mainly distributed in the wide valley and the active layer ranges from 50 to 60 cm above the permafrost layer [30].
2.2. Samples Treatment
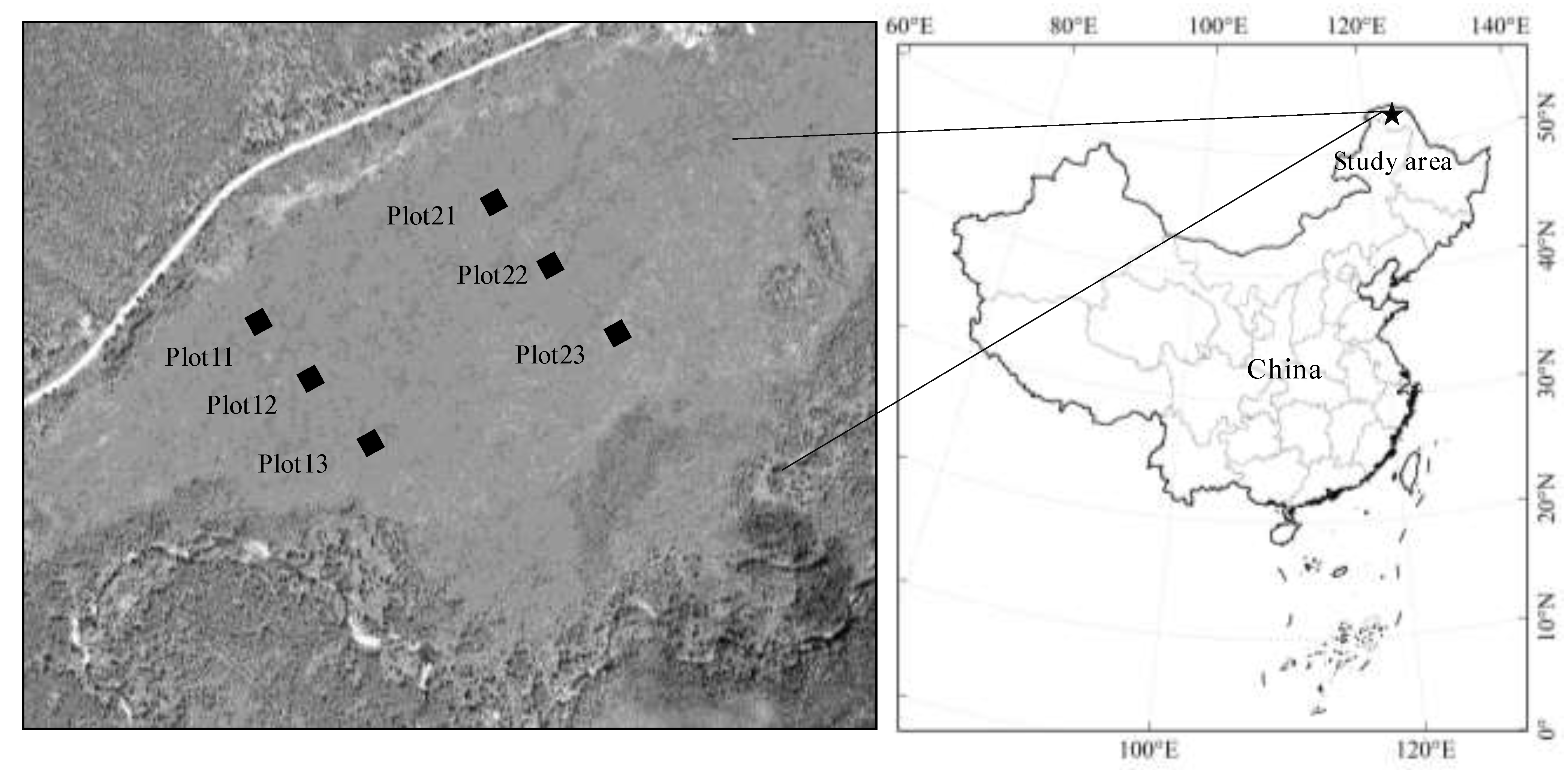
In September 2012, we established 2 small-scale (meters) transects, which were about 250 m apart and altitude ranges from 466 to 468 m. In each transect, 3 plots were about 50–100 m (Figure 1). In each plot, 5 layers samples (humus, 0–10, 10–20, 20–30, and 30–40 cm layers) were collected in hummocks with similar vegetation types. After removing the litter, living roots, and fauna by hand, we collected the samples of the modern humus layer, which was about 0–10 cm and consisted of hardly or not recognizable plant parts (Figure S1).
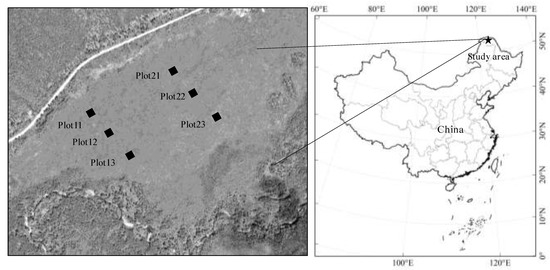
Figure 1.
Location of the study site and sampling plots.
Samples were placed in Ziplock bags with headspace removed, then transported to the laboratory within 48–72 h. Part of the samples were air-dried, crushed, and sieved through 0.25-mm mesh for chemical measurement. The remaining part of each sample was mixed and homogenized, with coarse roots moved by hand. All samples (about 0.25 kg) were moistened up to 70–80% water-holding capacity (WHC) and pre-incubated in aerated plastic at 22 °C for 7 days. WHC was following the method described by Zhang [31].
2.3. Soil Microbial Carbon (Cmic), Basal Respiration (BR), and Metabolic Quotient (qCO2)
Cmic of samples was determined by substrate-induced respiration (SIR) [32,33]. SIR was estimated by the rate of initial maximal respiration of microorganisms after amendment of all subsamples with glucose. Samples (1 g) were taken from preincubated soil and were placed in a vial (15 mL). Glucose solution was added to the samples (10 mg glucose g−1 soil, the volume was 0.1 mL). The vials were incubated for 3–5 h at 22 °C and the CO2 emission rate was measured by gas chromatography (Agilent 7820A). Cmic was calculated by the SIR rate according to the equation: Cmic (μg C g−1 sample) = (μL CO2 g−1 sample h−1) × 40.04 + 0.37. BR was determined by the CO2 emission rate from samples incubated for 24 h at 22 °C, which was carried out as described for SIR. In this treatment, water (0.1 mL g−1 sample) was added to samples instead of a glucose solution. The BR was expressed as mg CO2-C g−1 sample h−1. The specific respiration activity (qCO2) was calculated as BR/Cmic = qCO2 (μg CO2-C mg−1 Cmic h−1) [32].
2.4. Soil Properties Analyses
The volumetric samples was dried to subsequent weighing at 105 °C and determined bulk density (BD). The soil pH was measured in a water slurry (1:10 solid:liquid ratio). The samples were sieved through a 0.25 mm mesh for measuring the ash content, because there were lots of Sphagnum spp. debirs in the samples. Pre-drying at 100 °C was carried out to correct the dry mass and to evaporate the residual moisture. Then, the temperature was applied at 130 °C for 90 min. The samples were heated at 600 °C for 4 h and the remained material after heating was defined as the ash content [31,34]. The total carbon (TC) concentration was measured with the Multi N/C 2100 Analyzer (containing an HT 1500 Solid Module, Analytik Jena AG, Jena, Germany). The concentrations of total N (TN) and total P (TP) were analyzed by the Kjeldahl digestion method using a seal analytical analyzer (Seal AA3 Analytical, Norderstedt, Germany). The concentrations of sodium (Na), magnesium (Mg), potassium (K), and calcium (Ca) were analyzed by the HNO3-HClO4-HF extraction method using inductively coupled plasma emission spectrometer (ICP-AES, American).
2.5. Statistical Date Analyses
Physical and chemical analyses of all samples were performed in three replications, while SIR and BR were measured in four replications. One-way analysis of variance (ANOVA) was used to assess the mean values of sample properties, Cmic, Cmic/Corg, BR, and qCO2. Significance of differences from least square means was tested at a 5% significance level. Statistical analyses were performed using SPSS 20.0 (SPSS Inc., Chicago, IL, USA) and all graphs were prepared using the Origin 8.0 package (Origin Lab Corporation, Northampton, MA, USA). To examine the variation of soil properties and microbial respiration activities at small scale among plots, we used coefficients of variation (CV: standard deviation/mean ×100) based on Griffiths et al. [35]. Data were log-transformed for correlation analysis. The relationship between microbial respiration activities and all soil property variables was analyzed using Spearman’s rank correlation coefficient with R package [36].
3. Results
3.1. Sample Properties Variation
For properties of all samples at small-scale in permafrost peatland, variation values (as CV) of TC were smaller than the other properties (Table 1). CV values of TN were higher than the values of TC and TP, and CV values of C/P ratios were lower than those of C/N ratios and N/P ratios. It means that the variation of TN was larger at small-scale in this permafrost peatlands, which caused the changes of stoichiometry. The variation of pH values was also relatively lower, while the ash content and bulk density were relatively larger (Table S1). CV values of Na concentration was larger than the nutrient values (Mg, K, and Ca). These values of Mg and Ca were similar (Table S2).

Table 1.
Soil properties and microbial respiration activities of all samples as mean, standard deviation, and coefficients of variation (in percentage: CV).
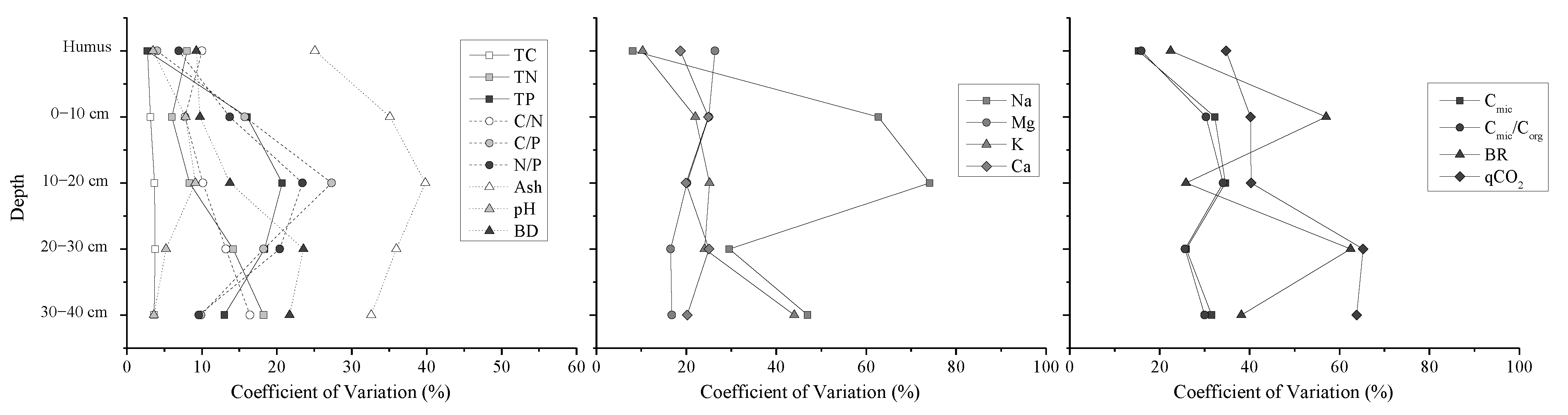
Most properties of humus layer varied smaller among plots than those of the different layers of peat. Variability of TN and TP in peat depth was greater than TC. CV values of Mg at humus layer were larger than these values of the other peat layers. But, this trend in the depth was different from the other elements. The variability in the peat Na concentration was greater in the depth than the other elements (Figure 2). CV values of TP, ash, pH, and Na in 10–20 cm layer were largest than those in the other layers, which did not support our first hypothesis.
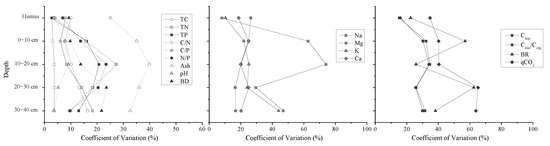
Figure 2.
Coefficients of variation (CVs) (%) with depth for sample properties and microbial respiration activities. The CVs reflect variation at small scale for all plots, with higher CVs indicative of greater plot-to-plot variability (TC: total carbon concentration, TN: total nitrogen concentration, TP: total phosphors concentration, C/N: carbon to nitrogen ratio, C/P: carbon to phosphors ration, Ash: ash content, pH: pH value, BD: bulk density, Na: sodium concentration, Mg: magnesium concentration, K: potassium concentration, Ca: calcium concentration, Cmic: microbial biomass carbon, Cmic/Corg: microbial entropy, BR: basal respiration rate, and qCO2: metabolic entropy).
3.2. Microbial Respiration Activities at Small-Scale
There was considerable variation in the microbial respiration activities among all plots. The values of Cmic, Cmic/Corg, BR, and qCO2 in humus layer were significantly higher than those in the other layers (p < 0.05). The Cmic was significantly decreased in soil depths (p < 0.05). The Cmic/Corg and BR was also decreased with soil depths, but Cmic/Corg was no significant difference between 20–30 cm and 30–40 cm layers (p > 0.05), BR was no significant difference between 10–20 cm and 20–30 cm layers (p > 0.05) (Table S3). For all samples at small scale, microbial respiration activities had higher CV values than soil properties (Table 1). The soil microbial activities were more complex at small scale in the permafrost peatland. The variation of microbial respiration activities in the humus layer was smaller than the other peat layers, and the variability of Cmic with depth was similar to the BR (Figure 2).
3.3. Relationships between Microbial Respiration Activities and Sample Properties
Considering all samples, there were significant correlations with TC, TN, ash, and bulk density (p < 0.05); the TC concentration positively correlated with Mg concentration; the TN concentration significantly correlated with TP and K concentrations (p < 0.05); the Na concentration was significantly related to bulk density (p < 0.05); there was significant correlation between Ca concentration and N/P ratio, pH value, Mg and K concentrations (p < 0.05); the K concentration also positively correlated with C/N ratio (Table S4). The correlations were different between sample nutrients and soil properties for all samples.
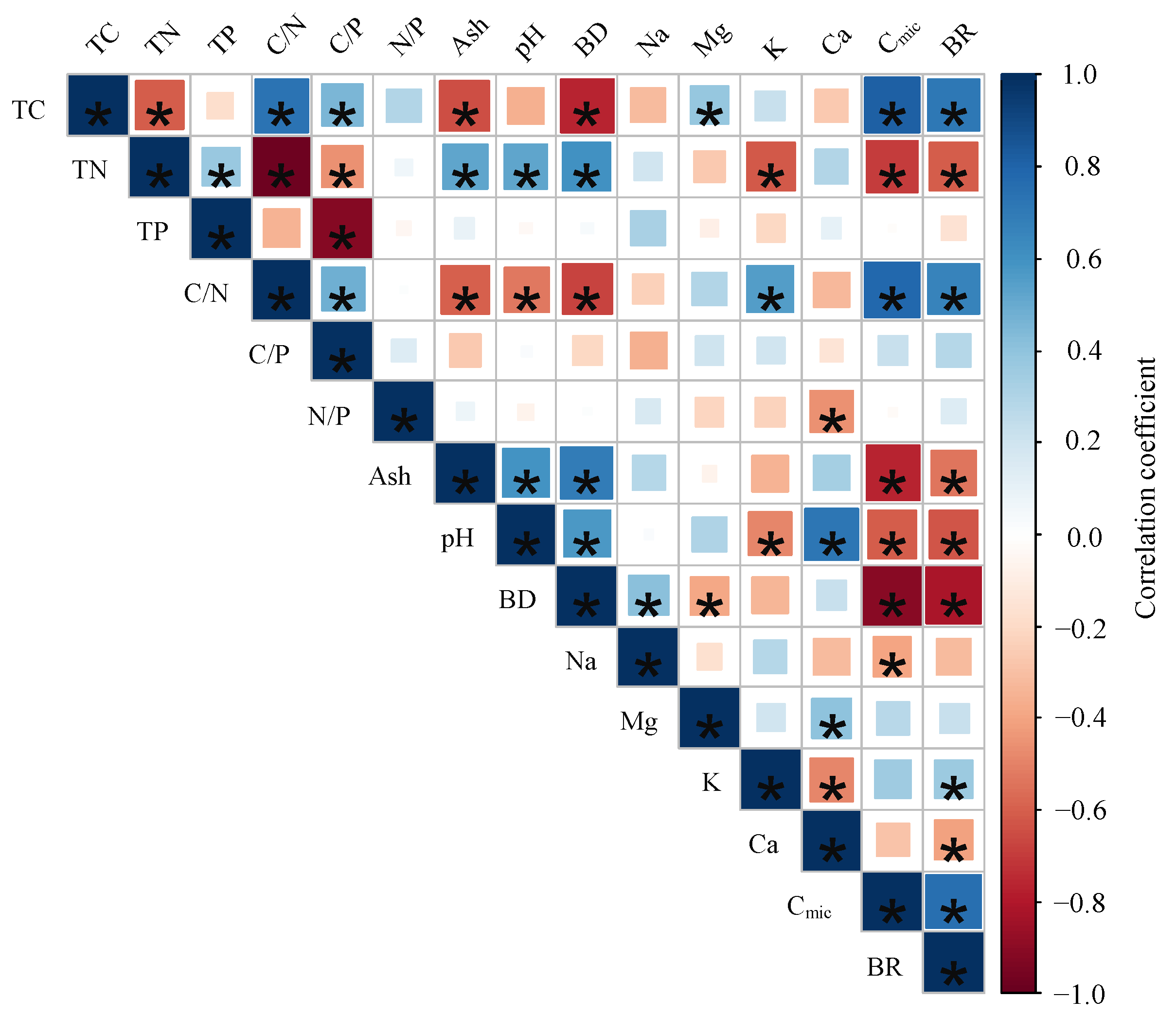
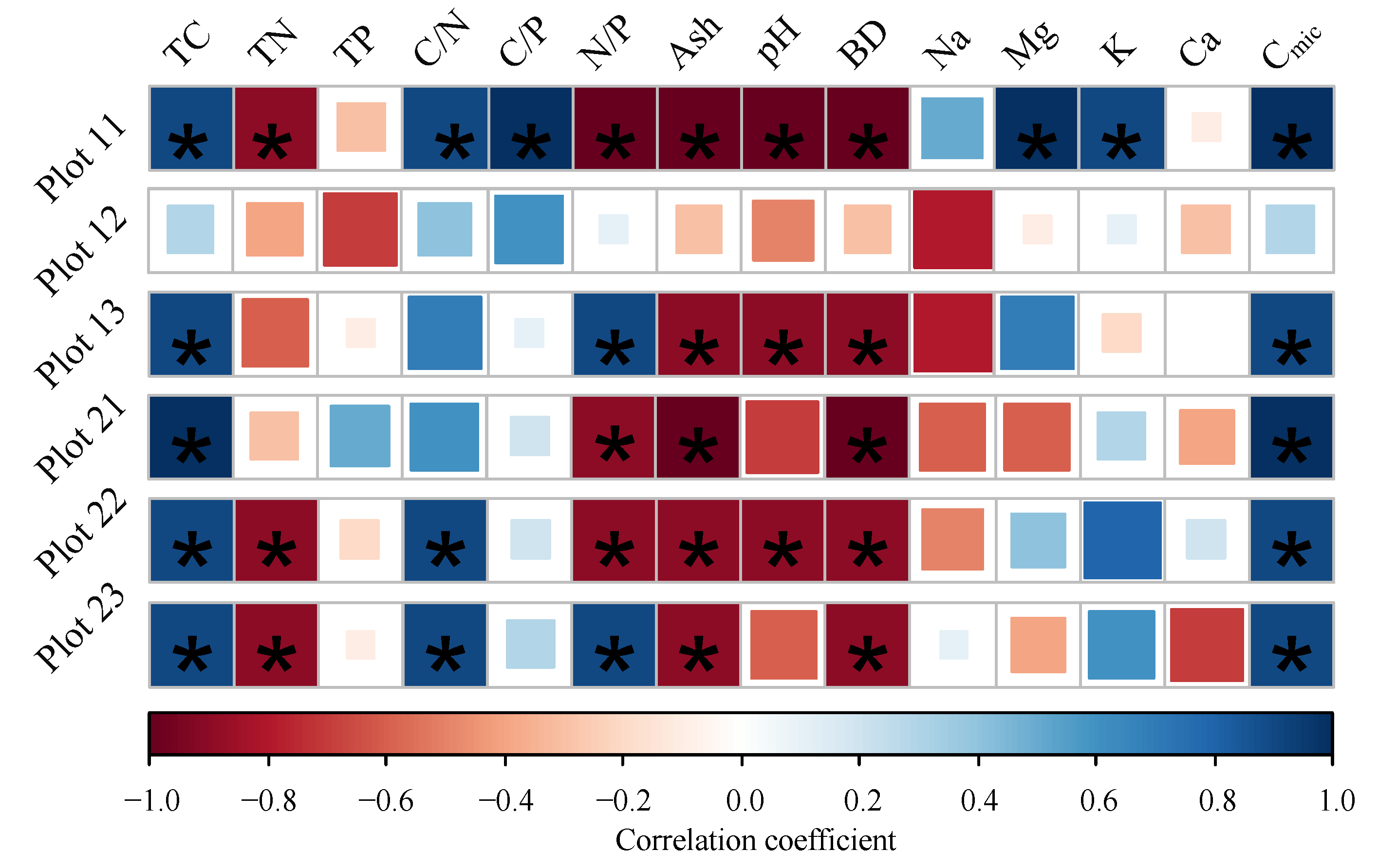
The significantly relationships were found between Cmic and TC, TN, C/N, ash, pH, bulk density, and Na (p < 0.05); the BR significantly correlated with TC, TN, C/N, pH, bulk density, K, Ca, and Cmic (p < 0.05) (Figure 3). However, The correlations between BR and soil properties were different among the plots (Table S5). There was no significant relationship between BR and soil properties in plot 12 (p > 0.05). Except for P12, BR significantly correlated with TC, ash, bulk density, and Cmic in the other plots (p < 0.05). There were strong negative correlations between BR and pH in plot 11, plot 13, and plot 22. Furthermore, BR was only significantly related to Mg and K in P 11 (p < 0.05) (Figure 4).
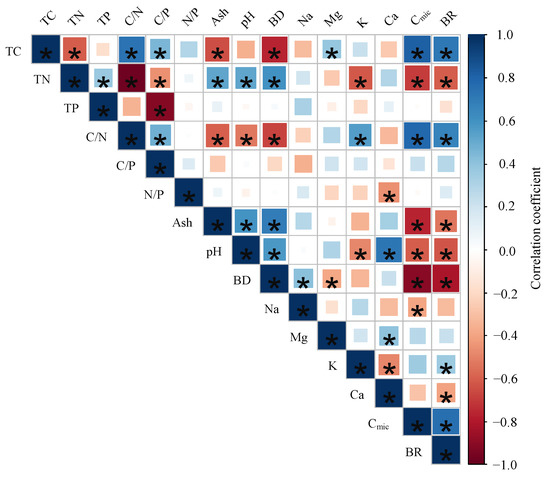
Figure 3.
Relationship between soil basal respiration rates (BR) and soil properties (TC: total carbon concentration, TN: total nitrogen concentration, TP: total phosphors concentration, C/N: carbon to nitrogen ratio, C/P: carbon to phosphors ration, Ash: ash content, pH: pH value, BD: bulk density, Na: sodium concentration, Mg: magnesium concentration, K: potassium concentration, Ca: calcium concentration, Cmic: microbial biomass carbon, Cmic/Corg: microbial entropy, BR: basal respiration rate, and qCO2: metabolic entropy). Significant (p < 0.05) correlation coefficients after Spearman (significant correlations (p < 0.05) are marked with asterisks.
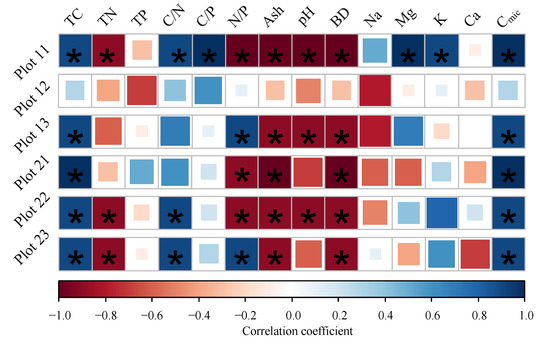
Figure 4.
Correlation coefficients after Spearman (significant correlations (p < 0.05) are marked with asterisks) for the soil basal respiration rates separated into the plots (TC: total carbon concentration, TN: total nitrogen concentration, TP: total phosphors concentration, C/N: carbon to nitrogen ratio, C/P: carbon to phosphors ration, Ash: ash content, pH: pH value, BD: bulk density, Na: sodium concentration, Mg: magnesium concentration, K: potassium concentration, Ca: calcium concentration, Cmic: microbial biomass carbon, Cmic/Corg: microbial entropy, BR: basal respiration rate, and qCO2: metabolic entropy).
4. Discussion
4.1. Soil Properties at Small Scale in the Permafrost Peatland
Peatland is a type of ecosystem in which the production exceeds decomposition, resulting in the accumulation of soil organic matter [14]. Vegetation composition in peatland varies on a spatial scale of several meters [37], which causes the variability of soil organic C and N stocks at small scales [10]. In our study, soil properties varied at small scale of meters. Peat properties can also vary at small scale in a tropical peatland, between both plant species [38], but the C/N values in our study area were higher than this tropical peatland. Different litter tissue types and plant species in peatland show greatly differences in litter decomposition rates and mass loss [11,39]. In the permafrost peatland of our study, the litter decomposition is also different among the different plants [40,41]. This might cause the variation of the properties in humus layer and peat layers. The accumulation of C, N, and P in peatland is related to litter resorption and decomposition, which affect the C:N:P stoichiometry in peatland biogeochemistry [41,42]. The variability of C/N ratio in depth was different to C/P and N/P rations at small-scale in our study and the largest variation of C/P and N/P rations of were in 10–20 cm layer, which was consistent to the variations of TP, pH, ash, and Na. The P and C chemistry is related to the aerobic and anaerobic environments in peatland [43]. The water table mostly ranged from 10 to 30 cm during the growing season in our study area [44]. The water table influences the vegetation composition, litter decomposition, and biogeochemical processes in the peatlands [12,13]. Changes in aerobic and anaerobic environments in soil may cause this variation at small scale.
Soil nutrient and biochemical element concentrations are the precise characterization of peat property and indicators of soil quality [43,45]. N, P, Mg, K, and Ca are macronutrients, which are uptaken by vegetation and released from decaying litter [14,46]. Ca concentrations in our studied area were higher than those in soils of the forest [46]. Our results showed that the variations of Mg, K, Ca, and Na were larger than those of TC, TN, and TP at small scale, and the variation of Mg was greater than those of K and Ca in the humus layer. The spatial variability pattern of soil chemical properties has been presented at small scale in a rewetted degraded peatland [18]. Adumitroaei et al. reported the different depositions of Ca, Mg, and other elements at local scale in peatland [47]. On the one hand, peat chemical characteristics are partly controlled by peatland nutrient status and vegetation types [13,14]; on the other hand, the variability of nutrient elements in peat could also affect the plant existing at small-scale in peatland. The larger Mg concentration in environments may be beneficial to the growing of the deciduous species, and moss is limited by K concentration [42]. The local environments exhibit heterogeneity in terms of plant community composition, hydrological regime, and active layer depth in permafrost peatland [20,48]. Our results showed that the relatively larger variation of soil nutrient and element concentrations in a permafrost peatland. Under climate change, the variation of soil nutrient and element should be contributed to plant change at small scale.
For all samples in our study, the TN concentration was negatively correlated to K concentration, the Mg concentration was positively correlated to TC concentration, and the Ca concentration was positively correlated to pH values. Peatland is N limitation or N and P co-limitation [43,49]. Some studies showed the effects of the N, P, and K additions on peatland [42,50,51]. K is a major factor that determine the effect of N deposition on Sphagnum growth in peatland [52], and the pH/Ca values could reflect the nutrient gradient in peatland [53]. However, few studies concern the interactive effect of N, P, K and other nutrients, such as Mg and Ca. We found the correlations were different between soil nutrients and soil properties. Quantification of soil nutrients and properties at small scale can provide a database of biochemical properties to inform our understanding of the biogeochemical processes under climate change.
4.2. Microbial Respiration Activities at Small Scale in the Permafrost Peatland
Peat properties are influenced by peatland nutrient status and vegetation types, which control microbial activities [11,14]. Ecosystem-level, such as primary productivity and parent material, controls Cmic [54]. The microbial respiration activities were different across nutrient gradient in peatland and peat types [54,55,56,57] and changed with soil depths at local and landscape scale in peatland [55,58]. Hence, soil microbial parameters are considered to be a good indicator of soil quality that leads to the changes of all soil properties [28]. Moreover, we observed that the variability of microbial respiration activities was higher than soil properties at small-scale. For the microbial study in peatland, sampling location should receive more attention at different scales.
For all samples, soil BR rates were positively correlated to soil TC and Cmic in our study. However, soil base respiration rates were also significantly correlated to soil TC and Cmic for all plots, except plot 12. Soil carbon properties still determine soil microbial respiration activities at small-scale in peatland. Soil organic carbon content determines the microbial activity in peatland [25,59]. However, there was no significant relationship between BR and soil properties in plot 12. The higher ash concentrations in this plot might cause this special result. This also indicates the great differences of soil quality at small scale.
Although peat chemical properties could control microbial activities [11,14], it is still difficult to establish general trends when relating soil microbial respiration activity to the soil properties. Some studies found the soil pH was one of the most influential factors to control microbial respiration activities across peat types [54,58,60]. There was also no strong correlations between soil BR and soil properties in heavily disturbed fens [61]. Our results showed the same results in plot 12. However, the BR rates were correlated negatively to total nitrogen concentrations in plot 11 and plot 12, and to soil pH values in plot 11, plot 13, and plot 22, but positively to K and Ca concentrations in plot 11. It indicated that there were different correlation relationships between soil microbial activities and soil properties at small scale among the plots, which is consistent with our second hypothesis. The microbial activities may be determined by the different peat quality in different studied regions [25,57]. For example, BR was positively correlated to N concentrations in undisturbed peatland [61], which was consistent with our studies in plot 11 and plot 22. Furthermore, it is difficult to disentangle soil properties under field conditions [61]. The sampling location and limitation of sampling number at small scale may cause these results.
4.3. Implications of Variation at Small Scale in Peatland
Permafrost peatlands are mostly found in boreal and subarctic regions [4,5], and the spatial variability at small scale would be caused by variations in hydrology, plant community, and micro-topography [24,37,62]. Peat properties predominantly derive from different plant species and vary at small scale [8,38]. Soil quality directly affects the substrate stability and indirectly affects microhabitat conditions [27,38,61]. C fluxes from peatlands have been shown to vary at small scale and microtopographic variability of hummock-hollow [63,64]. Climate warming and consequent permafrost degradation cause the changes of plant composition, soil environment, and hydrological processes, which have high spatial variability at different scales [18,24,65,66]. Small-scale local changes can have larger scale impacts in peatlands [19,22]. Therefore, the accuracy of experimental results in space scales becomes an issue in ecology, which should be crucial for future predictions [12,48]. The results of this study have shown that soil properties and microbial respiration activities in permafrost peatland exhibits considerable variability at small scale (meters). This variability indicates that sampling location is very important in peatland biogeochemical studies if we want to accurately estimate the geochemistry in a peatland.
5. Conclusions
Climate change and permafrost degradation create small-scale mosaics of plant and soil types in peatlands. This study examined the variability of soil quality at small scale by determining the soil properties and microbial respiration activities. We found that the soil quality presented high variation at small-scale, especially for microbial respiration activities. Total phosphorus, ash, and Na in the 10–20 cm layer had the largest variability with soil depth. Hence, it is very important to monitor soil properties and microbial respiration at small scale, which present an objective understanding of soil status, quality, and a way to determine the response of peatland to climate change. Although our results showed that carbon properties still determined the BR, soil properties were differently related to soil basal respiration among all plots. Sampling location and number may lead to these results, indicating that small-scale sampling is required to fully understand the soil quality in permafrost peatland.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/w14172597/s1, Figure S1: Soil profile in the permafrost peatland of the Great Hing’an Mountains; Table S1: Mean and standard deviation of samples characters among all plots; Table S2: Mean and standard deviation of element concentrations of samples among all plots; Table S3: Mean and standard deviation of microbial respiration activities among all plots; Table S4: Correlation coefficients after Spearman for basal respiration rates (BR) with soil properties (TC: total carbon concentration, TN: total nitrogen concentration, TP: total phosphors concentration, C/N: carbon to nitrogen ratio, C/P: carbon to phosphors ration, Ash: ash content, pH: pH value, BD: bulk density, Na: sodium concentration, Mg: magnesium concentration, K: potassium concentration, Ca: calcium concentration, and Cmic: microbial biomass carbon); Table S5: Correlation coefficients after Spearman for basal respiration rates (BR) separated into the different plots with soil properties (TC: total carbon concentration, TN: total nitrogen concentration, TP: total phosphors concentration, C/N: carbon to nitrogen ratio, C/P: carbon to phosphors ration, Ash: ash content, pH: pH value, BD: bulk density, Na: sodium concentration, Mg: magnesium concentration, K: potassium concentration, Ca: calcium concentration, and Cmic: microbial biomass carbon).
Author Contributions
Conceptualization, X.W. and L.S.; methodology, X.S. and L.S.; investigation, Y.D.; data curation, N.C.; writing—original draft preparation, X.W.; funding acquisition, X.W. and X.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China, grant number 41471056, 41971143, and 41861134029, and National Science and Technology Fundamental Resources Investigation Program of China grant number 2019FY100600.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We wish to acknowledge Zhenling Gao for the field work, and Jinli Gao for the laboratory analyses.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gorham, E. Northern peatlands: Role in the carbon cycle and probable responses to climatic warming. Ecol. Appl. 1991, 1, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Tarnocai, C.; Canadell, J.G.; Schuur, E.A.G.; Kuhry, P.; Mazhitova, G.; Zimov, S. Soil organic carbon pools in the northern circumpolar permafrost region. Glob. Biogeochem. Cycles 2009, 23, GB2023. [Google Scholar] [CrossRef]
- Räisänen, J.; Hansson, U.; Ullerstig, A.; Döscher, R.; Graham, L.; Jones, C.; Meier, H.; Samuelsson, P.; Willén, U. European climate in the late twenty-first century: Regional simulations with two driving global models and two forcing scenarios. Clim. Dynam. 2004, 22, 13–31. [Google Scholar] [CrossRef]
- Dorrepaal, E.; Toet, S.; van Logtestijn, R.S.; Swart, E.; van de Weg, M.J.; Callaghan, T.V.; Aerts, R. Carbon respiration from subsurface peat accelerated by climate warming in the subarctic. Nature 2009, 460, 616–619. [Google Scholar] [CrossRef]
- Pengerud, A.; Dignac, M.-F.; Certini, G.; Strand, L.T.; Forte, C.; Rasse, D.P. Soil organic matter molecular composition and state of decomposition in three locations of the European Arctic. Biogeochemistry 2017, 135, 277–292. [Google Scholar] [CrossRef]
- Turetsky, M.R.; Wieder, R.; Vitt, D.; Evans, R.; Scott, K. The disappearance of relict permafrost in boreal north America: Effects on peatland carbon storage and fluxes. Glob. Chang. Biol. 2007, 13, 1922–1934. [Google Scholar] [CrossRef]
- Cresto Aleina, F.; Runkle, B.R.K.; Kleinen, T.; Kutzbach, L.; Schneider, J.; Brovkin, V. Modeling micro-topographic controls on boreal peatland hydrology and methane fluxes. Biogeosciences 2015, 12, 5689–5704. [Google Scholar] [CrossRef] [Green Version]
- Wright, E.L.; Black, C.R.; Turner, B.L.; Sjögersten, S. Environmental controls of temporal and spatial variability in CO2 and CH4 fluxes in a neotropical peatland. Glob. Chang. Biol. 2013, 19, 3775–3789. [Google Scholar] [CrossRef]
- Jenerette, G.D.; Wu, J. Interactions of ecosystem processes with spatial heterogeneity in the puzzle of nitrogen limitation. Oikos 2004, 107, 273–282. [Google Scholar] [CrossRef]
- Evgrafova, A.; de la Haye, T.R.; Haase, I.; Shibistova, O.; Guggenberger, G.; Tananaev, N.; Sauheitl, L.; Spielvogel, S. Small-scale spatial patterns of soil organic carbon and nitrogen stocks in permafrost-affected soils of northern Siberia. Geoderma 2018, 329, 91–107. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Schimel, J.P.; Trumbore, S.E.; Randerson, J.R. Controls over carbon storage in high-latitude soils. Glob. Chang. Biol. 2000, 6, 196–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limpens, J.; Berendse, F.; Blodau, C.; Canadell, J.; Freeman, C.; Holden, J.; Roulet, N.; Rydin, H.; Schaepman-Strub, G. Peatlands and the carbon cycle: From local processes to global implications—A synthesis. Biogeosciences 2008, 5, 1475–1491. [Google Scholar] [CrossRef] [Green Version]
- Laiho, R. Decomposition in peatlands: Reconciling seemingly contrasting results on the impacts of lowered water levels. Soil Biol. Biochem. 2006, 38, 2011–2024. [Google Scholar] [CrossRef]
- Moore, T.R.; Basiliko, N. Decomposition in boreal peatlands. In Boreal Peatland Ecosystems; Wieder, R.K., Vitt, D.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 125–143. [Google Scholar]
- Mathijssen, P.J.H.; Galka, M.; Borken, W.; Knorr, K.H. Plant communities control long term carbon accumulation and biogeochemical gradients in a Patagonian bog. Sci. Total Environ. 2019, 684, 670–681. [Google Scholar] [CrossRef]
- Baldrian, P. Distribution of extracellular enzymes in soils: Spatial heterogeneity and determining factors at various scales. Soil Sci. Soc. Am. J. 2014, 78, 11–18. [Google Scholar] [CrossRef]
- Drollinger, S.; Knorr, K.-H.; Knierzinger, W.; Glatzel, S. Peat decomposition proxies of Alpine bogs along a degradation gradient. Geoderma 2020, 369, 114331. [Google Scholar] [CrossRef]
- Negassa, W.; Baum, C.; Schlichting, A.; Müller, J.; Leinweber, P. Small-scale spatial variability of soil chemical and biochemical properties in a rewetted degraded peatland. Front. Environ. Sci. 2019, 7, 166. [Google Scholar] [CrossRef] [Green Version]
- Ulanowski, T.A.; Branfireun, B.A. Small-scale variability in peatland pore-water biogeochemistry, Hudson Bay Lowland, Canada. Sci. Total Environ. 2013, 454–455, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Holden, J. Peatland hydrology and carbon release: Why small-scale process matters. Philos. Trans. R. Soc. A 2005, 363, 2891–2913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sachs, T.; Giebels, M.; Boike, J.; Kutzbach, L. Environmental controls on CH4 emission from polygonal tundra on the microsite scale in the Lena river delta, Siberia. Glob. Chang. Biol. 2010, 16, 3096–3110. [Google Scholar] [CrossRef]
- Macrae, M.; Devito, K.; Strack, M.; Waddington, J. Effect of water table drawdown on peatland nutrient dynamics: Implications for climate change. Biogeochemistry 2013, 112, 661–676. [Google Scholar] [CrossRef]
- Nungesser, M.K. Modelling microtopography in boreal peatlands: Hummocks and hollows. Ecol. Model. 2003, 165, 175–207. [Google Scholar] [CrossRef]
- Cannone, N.; Guglielmin, M.; Gerdol, R. Relationships between vegetation patterns and periglacial landforms in northwestern Svalbard. Polar Biol. 2004, 27, 562–571. [Google Scholar] [CrossRef] [Green Version]
- Blaško, R.; Holm Bach, L.; Yarwood, S.A.; Trumbore, S.E.; Högberg, P.; Högberg, M.N. Shifts in soil microbial community structure, nitrogen cycling and the concomitant declining N availability in ageing primary boreal forest ecosystems. Soil Biol. Biochem. 2015, 91, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.W.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kogel-Knabner, I.; Lehmann, J.; Manning, D.A.; et al. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drzymulska, D. Peat decomposition-shaping factors, significance in environmental studies and methods of determination; a literature review. Geologos 2016, 22, 61–69. [Google Scholar] [CrossRef] [Green Version]
- Romaniuk, R.; Giuffré, L.; Costantini, A.; Bartoloni, N.; Nannipieri, P. A comparison of indexing methods to evaluate quality of soils: The role of soil microbiological properties. Soil Res. 2011, 49, 733–741. [Google Scholar] [CrossRef]
- Jin, H.; Yu, Q.; Lü, L.; Guo, D.; He, R.; Yu, S.; Sun, G.; Li, Y. Degradation of permafrost in the Xing’anling Mountains, Northeastern China. Permafr. Periglac. Processes 2007, 18, 245–258. [Google Scholar] [CrossRef]
- Wang, X.; Li, X.; Hu, Y.; Lü, J.; Sun, J.; Li, Z.; He, H. Potential carbon mineralization of permafrost peatlands in Great Hing’an Mountains, China. Wetlands 2010, 30, 747–756. [Google Scholar] [CrossRef]
- Zhang, Z. Development and Utilization of Peat Resources; Chinese Jilin Science and Technology Press: Changchun, China, 2000; pp. 185–193. [Google Scholar]
- Anderson, J.; Domsch, K. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Susyan, E.A.; Wirth, S.; Ananyeva, N.D.; Stolnikova, E.V. Forest succession on abandoned arable soils in European Russia–Impacts on microbial biomass, fungal-bacterial ratio, and basal CO2 respiration activity. Eur. Soil Biol. 2011, 47, 169–174. [Google Scholar] [CrossRef]
- Leifeld, J.; Gubler, L.; Grünig, A. Organic matter losses from temperate ombrotrophic peatlands: An evaluation of the ash residue method. Plant Soil 2011, 341, 349–361. [Google Scholar] [CrossRef]
- Griffiths, N.A.; Sebestyen, S.D.; Oleheiser, K.C. Variation in peatland porewater chemistry over time and space along a bog to fen gradient. Sci. Total Environ. 2019, 697, 134152. [Google Scholar] [CrossRef] [PubMed]
- Harrell, F. Hmisc: Harrell Miscellaneous. R Package Version 3.17-4. 2016. Available online: http://CRAN.R-project.org/package=Hmisc (accessed on 1 December 2019).
- Eppinga, M.B.; Rietkerk, M.; Wassen, M.J.; De Ruiter, P.C. Linking habitat modification to catastrophic shifts and vegetation patterns in bogs. Plant Ecol. 2009, 200, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Girkin, N.T.; Vane, C.H.; Cooper, H.V.; Moss-Hayes, V.; Craigon, J.; Turner, B.L.; Ostle, N.; Sjögersten, S. Spatial variability of organic matter properties determines methane fluxes in a tropical forested peatland. Biogeochemistry 2018, 142, 231–245. [Google Scholar] [CrossRef]
- Hoyos-Santillan, J.; Lomax, B.H.; Large, D.; Turner, B.L.; Boom, A.; Lopez, O.R.; Sjögersten, S. Getting to the root of the problem: Litter decomposition and peat formation in lowland Neotropical peatlands. Biogeochemistry 2015, 126, 115–129. [Google Scholar] [CrossRef]
- Mao, R.; Zhang, X.; Song, C.; Wang, X.; Finnegan, P.M. Plant functional group controls litter decomposition rate and its temperature sensitivity: An incubation experiment on litters from a boreal peatland in northeast China. Sci. Total Environ. 2018, 626, 678–683. [Google Scholar] [CrossRef]
- Xu, J.; Lin, G.; Liu, B.; Mao, R. Linking leaf nutrient resorption and litter decomposition to plant mycorrhizal associations in boreal peatlands. Plant Soil 2020, 448, 413–424. [Google Scholar] [CrossRef]
- Wang, M.; Larmola, T.; Murphy, M.T.; Moore, T.R.; Bubier, J.L. Stoichiometric response of shrubs and mosses to long-term nutrient (N, P and K) addition in an ombrotrophic peatland. Plant Soil 2016, 400, 403–416. [Google Scholar] [CrossRef]
- Basiliko, N.; Blodau, C.; Roehm, C.; Bengtson, P.; Moore, T.R. Regulation of decomposition and methane dynamics across natural, commercially mined, and restored northern peatlands. Ecosystems 2007, 10, 1148–1165. [Google Scholar] [CrossRef]
- Miao, Y.; Song, C.; Sun, L.; Wang, X.; Meng, H.; Mao, R. Growing season methane emission from a boreal peatland in the continuous permafrost zone of Northeast China: Effects of active layer depth and vegetation. Biogeosciences 2012, 9, 4455–4464. [Google Scholar] [CrossRef] [Green Version]
- Gil-Sotres, F.; Trasar-Cepeda, C.; Leirós, M.C.; Seoane, S. Different approaches to evaluating soil quality using biochemical properties. Soil Biol. Biochem. 2005, 37, 877–887. [Google Scholar] [CrossRef]
- Viers, J.; Prokushkin, A.S.; Pokrovsky, O.S.; Auda, Y.; Kirdyanov, A.V.; Beaulieu, E.; Zouiten, C.; Oliva, P.; Dupré, B. Seasonal and spatial variability of elemental concentrations in boreal forest larch foliage of Central Siberia on continuous permafrost. Biogeochemistry 2012, 113, 435–449. [Google Scholar] [CrossRef]
- Adumitroaei, M.V.; Iancu, G.O.; Rățoi, B.G.; Doru, C.S.; Sandu, C.M. Spatial distribution and geochemistry of major and trace elements from Mohoș peatland, Harghita Mountains, Romania. Holocene 2018, 28, 1936–1947. [Google Scholar] [CrossRef]
- Siewert, M.B. High-resolution digital mapping of soil organic carbon in permafrost terrain using machine learning: A case study in a sub-Arctic peatland environment. Biogeosciences 2018, 15, 1663–1682. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Moore, T.R.; Talbot, J.; Richard, P.J.H. The cascade of C:N:P stoichiometry in an ombrotrophic peatland: From plants to peat. Environ. Res. Lett. 2014, 9, 024003. [Google Scholar] [CrossRef] [Green Version]
- Chiwa, M.; Sheppard, L.J.; Leith, I.D.; Leeson, S.R.; Tang, Y.S.; Neil Cape, J. Long-term interactive effects of N addition with P and K availability on N status of Sphagnum. Environ. Pollut. 2018, 237, 468–472. [Google Scholar] [CrossRef] [Green Version]
- Juutinen, S.; Moore, T.R.; Bubier, J.L.; Arnkil, S.; Humphreys, E.; Marincak, B.; Roy, C.; Larmola, T. Long-term nutrient addition increased CH4 emission from a bog through direct and indirect effects. Sci. Rep. 2018, 8, 3838. [Google Scholar] [CrossRef]
- Limpens, J.; Holmgren, M.; Jacobs, C.M.; Van der Zee, S.E.; Karofeld, E.; Berendse, F. How does tree density affect water loss of peatlands? A mesocosm experiment. PLoS ONE 2014, 9, e91748. [Google Scholar]
- Hájek, M.; Hekera, P.; Hájková, P. Spring fen vegetation and water chemistry in the Western Carpathian flysch zone. Folia Geobot. 2002, 37, 205–224. [Google Scholar] [CrossRef]
- Fisk, M.C.; Ruether, K.F.; Yavitt, J.B. Microbial activity and functional composition among northern peatland ecosystems. Soil Biol. Biochem. 2003, 35, 591–602. [Google Scholar] [CrossRef]
- Brouns, K.; Keuskamp, J.A.; Potkamp, G.; Verhoeven, J.T.A.; Hefting, M.M. Peat origin and land use effects on microbial activity, respiration dynamics and exo-enzyme activities in drained peat soils in the Netherlands. Soil Biol. Biochem. 2016, 95, 144–155. [Google Scholar] [CrossRef]
- Myers, B.; Webster, K.L.; McLaughlin, J.W.; Basiliko, N. Microbial activity across a boreal peatland nutrient gradient: The role of fungi and bacteria. Wetl. Ecol. Manag. 2011, 20, 77–88. [Google Scholar] [CrossRef]
- Preston, M.D.; Smemo, K.A.; McLaughlin, J.W.; Basiliko, N. Peatland microbial communities and decomposition processes in the James Bay Lowlands, Canada. Front. Microbiol. 2012, 3, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grodnitskaya, I.D.; Karpenko, L.V.; Knorre, A.A.; Syrtsov, S.N. Microbial activity of peat soils of boggy larch forests and bogs in the permafrost zone of central Evenkia. Eurasian Soil Sci. 2013, 46, 61–73. [Google Scholar] [CrossRef]
- Kraigher, B.; Stres, B.; Hacin, J.; Ausec, L.; Mahne, I.; Vanelsas, J.; Mandicmulec, I. Microbial activity and community structure in two drained fen soils in the Ljubljana Marsh. Soil Biol. Biochem. 2006, 38, 2762–2771. [Google Scholar] [CrossRef]
- Ausec, L.; Kraigher, B.; Mandic-Mulec, I. Differences in the activity and bacterial community structure of drained grassland and forest peat soils. Soil Biol. Biochem. 2009, 41, 1874–1881. [Google Scholar] [CrossRef]
- Säurich, A.; Tiemeyer, B.; Don, A.; Fiedler, S.; Bechtold, M.; Amelung, W.; Freibauer, A. Drained organic soils under agriculture-The more degraded the soil the higher the specific basal respiration. Geoderma 2019, 355, 113911. [Google Scholar] [CrossRef]
- Waddington, J.M.; Roulet, N.T. Carbon balance of a boreal patterned peatland. Glob. Chang. Biol. 2008, 6, 87–97. [Google Scholar] [CrossRef]
- Pelletier, L.; Garneau, M.; Moore, T.R. Variation in CO2 exchange over three summers at microform scale in a boreal bog, Eastmain region, Québec, Canada. J. Geophys. Res. 2011, 116, G03019. [Google Scholar]
- Pullens, J.W.M.; Sottocornola, M.; Kiely, G.; Toscano, P.; Gianelle, D. Carbon fluxes of an alpine peatland in Northern Italy. Agr. For. Meteorol. 2016, 220, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Dieleman, C.M.; Branfireun, B.A.; McLaughlin, J.W.; Lindo, Z. Climate change drives a shift in peatland ecosystem plant community: Implications for ecosystem function and stability. Glob. Chang. Biol. 2015, 21, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Treat, C.C.; Marushchak, M.E.; Voigt, C.; Zhang, Y.; Tan, Z.; Zhuang, Q.; Virtanen, T.A.; Rasanen, A.; Biasi, C.; Hugelius, G.; et al. Tundra landscape heterogeneity, not interannual variability, controls the decadal regional carbon balance in the Western Russian Arctic. Glob. Chang. Biol. 2020, 24, 5188–5204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).