
Effects of Microalgae Grown in Membrane Treated Distillery Wastewater as Diet on Growth and Survival Rate of Juvenile Pearl Oyster (Pinctada fucata martensii)



Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae Collection and Growth Conditions
2.1.1. Collection and Preparation of Wastewater
2.1.2. Wastewater Nutrient Analysis and Removal Efficiency
2.1.3. Determination of Dry Weight
2.1.4. Carbohydrate, Protein and Lipid Quantification
2.1.5. FAMEs Test and Amino Acids Determination
2.1.6. Experimental Diet and Feeding Procedures
2.1.7. Survival Rate and Growth Rate
3. Results
3.1. Biomass Production and Nutrient Removal Efficiency
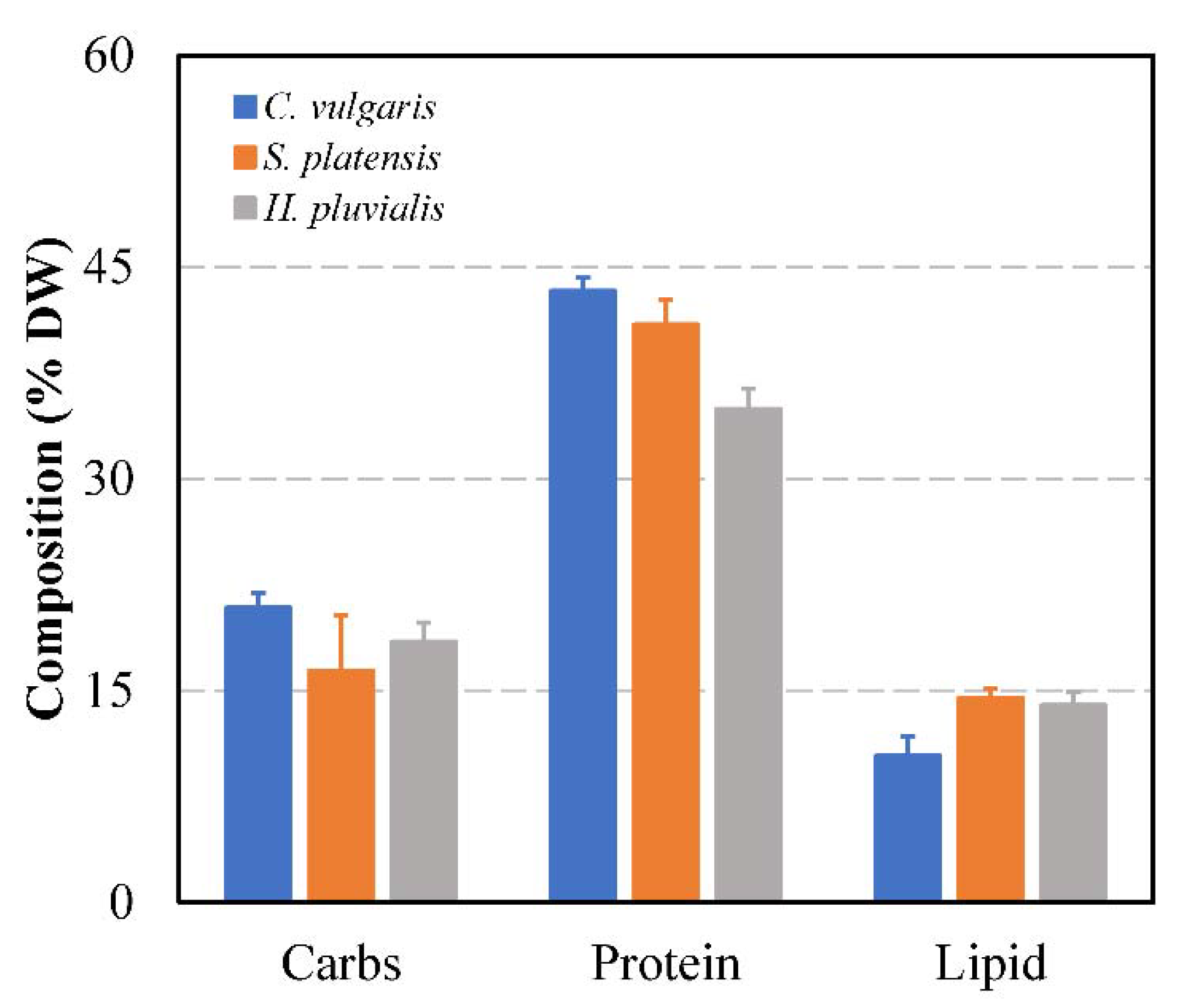
3.2. Biochemical Composition of the Microalgae
3.2.1. Protein, Carbohydrate and Lipid
3.2.2. Amino Acids and Fatty Acids
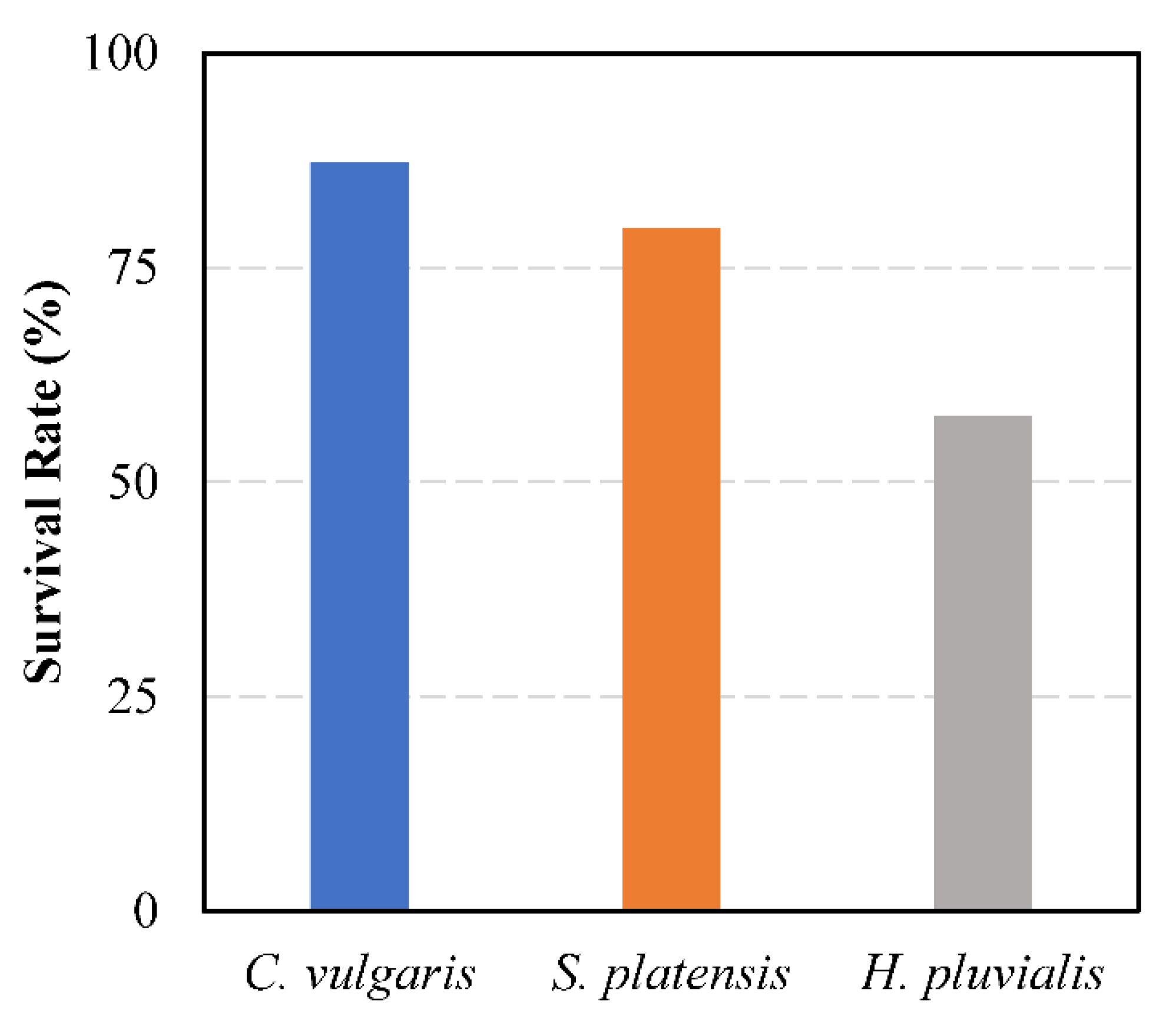
3.3. Growth Performance and Survival Rate of Juvenile Pearl Oyster (Pinctada Fucata Martensii)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mustafa, M.G.; Nakagawa, H. A Review—Dietary Benefits of Algae as an Additive in Fish Feed. Isr. J. Aquac. Bamidgeh 1995, 47, 155–162. [Google Scholar]
- Dineshbabu, G.; Goswami, G.; Kumar, R.; Sinha, A.; Das, D. Microalgae–nutritious, sustainable aqua- and animal feed source. J. Funct. Foods 2019, 62, 103545. [Google Scholar] [CrossRef]
- Madeira, M.S.; Cardoso, C.; Lopes, P.A.; Coelho, D.; Afonso, C.; Bandarra, N.M.; Prates, J.A.M. Microalgae as feed ingredients for livestock production and meat quality: A review. Livest. Sci. 2017, 205, 111–121. [Google Scholar] [CrossRef]
- Patil, V.; Källqvist, T.; Olsen, E.; Vogt, G.; Gislerød, H.R. Fatty acid composition of 12 microalgae for possible use in aquaculture feed. Aquac. Int. 2007, 15, 1–9. [Google Scholar] [CrossRef]
- Kadir, W.N.A.; Lam, M.K.; Uemura, Y.; Lim, J.W.; Lee, K.T. Harvesting and pre-treatment of microalgae cultivated in wastewater for biodiesel production: A review. Energy Convers. Manag. 2018, 171, 1416–1429. [Google Scholar] [CrossRef]
- Slade, R.; Bauen, A. Micro-algae cultivation for biofuels: Cost, energy balance, environmental impacts and future prospects. Biomass Bioenergy 2013, 53, 29–38. [Google Scholar] [CrossRef]
- Leite, L.D.S.; Hoffmann, M.T.; Daniel, L.A. Microalgae cultivation for municipal and piggery wastewater treatment in Brazil. J. Water Process Eng. 2019, 31, 100821. [Google Scholar] [CrossRef]
- Stutter, M.I.; Graeber, D.; Evans, C.D.; Wade, A.J.; Withers, P.J.A. Balancing macronutrient stoichiometry to alleviate eutrophication. Sci. Total Environ. 2018, 634, 439–447. [Google Scholar] [CrossRef]
- Sheng, A.L.K.; Bilad, M.R.; Osman, N.B.; Arahman, N. Sequencing batch membrane photobioreactor for real secondary effluent polishing using native microalgae: Process performance and full-scale projection. J. Clean. Prod. 2017, 168, 708–715. [Google Scholar] [CrossRef]
- Quintero-Dallos, V.; García-Martínez, J.B.; Contreras-Ropero, J.E.; Barajas-Solano, A.F.; Barajas-Ferrerira, C.; Lavecchia, R.; Zuorro, A. Vinasse as a sustainable medium for the production of Chlorella vulgaris UTEX 1803. Water 2019, 11, 1526. [Google Scholar] [CrossRef]
- Olguín, E.J. Phycoremediation: Key issues for cost-effective nutrient removal processes. Biotechnol. Adv. 2003, 22, 81–91. [Google Scholar] [CrossRef]
- Ayala, F.J.; Bravo, R.B. Animal wastes media for Spirulina production. Algological Studies/Archiv für Hydrobiologie. 1984, 67, 349–355. [Google Scholar] [CrossRef]
- Costa, R.H.; Medri, W.; Perdomo, C.C. High-rate pond for treatment of piggery wastes. Water Sci. Technol. 2000, 42, 357–362. [Google Scholar] [CrossRef]
- Jimenez-Perez, M.V.; Sánchez-Castillo, P.; Romera, O.; Fernandez-Moreno, D.; Pérez-Martınez, C. Growth and nutrient removal in free and immobilized planktonic green algae isolated from pig manure. Enzym. Microb. Technol. 2004, 34, 392–398. [Google Scholar] [CrossRef]
- Liu, J.; Hu, Q. Chlorella: Industrial production of cell mass and chemicals. In Handbook of Microalgal Culture: Applied Phycology and Biotechnology, 2nd ed.; Editor Richmond, A., Hu, Q., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 329–338. [Google Scholar]
- Barbarino, E.; Lourenço, S.O. An evaluation of methods for extraction and quantification of protein from marine macro- and microalgae. J. Appl. Phycol. 2005, 17, 447–460. [Google Scholar] [CrossRef]
- Ge, S.; Qiu, S.; Tremblay, D.; Viner, K.; Champagne, P.; Jessop, P.G. Centrate wastewater treatment with Chlorella vulgaris: Simultaneous enhancement of nutrient removal, biomass and lipid production. Chem. Eng. J. 2018, 342, 310–320. [Google Scholar] [CrossRef]
- Ge, S.; Champagne, P. Nutrient removal, microalgal biomass growth, harvesting and lipid yield in response to centrate wastewater loadings. Water Res. 2016, 88, 604–612. [Google Scholar] [CrossRef]
- Yang, C.; Hao, R.; Deng, Y.; Liao, Y.; Wang, Q.; Sun, R.; Jiao, Y.; Du, X. Effects of protein sources on growth, immunity and antioxidant capacity of juvenile pearl oyster Pinctada fucata martensii. Fish Shellfish Immunol. 2017, 67, 411–418. [Google Scholar] [CrossRef]
- Cho, S.; Luong, T.T.; Lee, D.; Oh, Y.K.; Lee, T. Reuse of effluent water from a municipal wastewater treatment plant in microalgae cultivation for biofuel production. Bioresour. Technol. 2011, 102, 8639–8645. [Google Scholar] [CrossRef]
- Mularczyk, M.; Michalak, I.; Marycz, K. Astaxanthin and other nutrients from Haematococcus pluvialis—Multifunctional applications. Mar. Drugs 2020, 18, 459. [Google Scholar] [CrossRef]
- Razzak, S.A.; Hossain, M.M.; Lucky, R.A.; Bassi, A.S.; de Lasa, H. Integrated CO2 capture, wastewater treatment and biofuel production by microalgae culturing—A review. Renew. Sustain. Energy Rev. 2013, 27, 622–653. [Google Scholar] [CrossRef]
- Travieso, L.; Benitez, F.; Dupeyrón, R. Algae growth potential measurement in distillery wastes. Bull. Environ. Contam. Toxicol. 1999, 62, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.E.; Jiménez, J.M.; El Yousfi, F. Influence of phosphorus concentration and temperature on growth and phosphorus uptake by the microalga Scenedesmus obliquus. Bioresour. Technol. 1999, 67, 233–240. [Google Scholar] [CrossRef]
- Tran, H.D.; Do, T.T.; Le, T.L.; Tran Nguyen, M.L.; Pham, C.H.; Melkonian, M. Cultivation of Haematococcus pluvialis for astaxanthin production on angled bench-scale and large-scale biofilm-based photobioreactors. Vietnam J. Sci. Technol. Eng. 2017, 61, 61–70. [Google Scholar] [CrossRef]
- Lorenz, R.T.; Cysewski, G.R. Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends Biotechnol. 2000, 18, 160–167. [Google Scholar] [CrossRef]
- Kim, G.; Mujtaba, G.; Lee, K. Effects of nitrogen sources on cell growth and biochemical composition of marine chlorophyte Tetraselmis sp. for lipid production. Algae. 2016, 31, 257–266. [Google Scholar] [CrossRef]
- Klin, M.; Pniewski, F.; Latała, A. Growth phase-dependent biochemical composition of green microalgae: Theoretical considerations for biogas production. Bioresour. Technol. 2020, 303, 122875. [Google Scholar] [CrossRef]
- Niccolai, A.; Chini Zittelli, G.; Rodolfi, L.; Biondi, N.; Tredici, M.R. Microalgae of interest as food source: Biochemical composition and digestibility. Algal Res. 2019, 42, 101617. [Google Scholar] [CrossRef]
- Pimentel, D. Energy inputs in food crop production in developing and developed nations. Energies 2009, 2, 1–24. [Google Scholar] [CrossRef]
- Lu, W.D.; Lin, M.J.; Guo, X.L.; Lin, Z.Y. Cultivation of spirulina platensis using raw piggery wastewater for nutrients bioremediation and biomass production: Effect of ferrous sulfate supplementation. Desalination Water Treat. 2020, 175, 60–70. [Google Scholar] [CrossRef]
- Craig, S.; Kuhn, D.; Schwarz, M. Understanding Fish Nutrition, Feeds, and Feeding Steven. Virginia Cooperative Extention: Blacksburg, VA, USA, 2017; pp. 1–6. [Google Scholar]
- Adams, C.; Godfrey, V.; Wahlen, B.; Seefeldt, L.; Bugbee, B. Understanding precision nitrogen stress to optimize the growth and lipid content tradeoff in oleaginous green microalgae. Bioresour. Technol. 2013, 131, 188–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwenk, D.; Seppälä, J.; Spilling, K.; Virkki, A.; Tamminen, T.; Oksman-Caldentey, K.M.; Rischer, H. Lipid content in 19 brackish and marine microalgae: Influence of growth phase, salinity and temperature. Aquat. Ecol. 2013, 47, 415–424. [Google Scholar] [CrossRef]
- Tang, B.; Liu, B.; Wang, G.; Zhang, T.; Xiang, J. Effects of various algal diets and starvation on larval growth and survival of Meretrix meretrix. Aquaculture 2006, 254, 526–533. [Google Scholar] [CrossRef]
- Mata, S.N.; de Souza Santos, T.; Cardoso, L.G.; Andrade, B.B.; Duarte, J.H.; Costa, J.A.V.; de Souza, C.O.; Druzian, J.I. Spirulina sp. LEB 18 cultivation in a raceway-type bioreactor using wastewater from desalination process: Production of carbohydrate-rich biomass. Bioresour. Technol. 2020, 311, 123495. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Ghosh, A.K.; Pal, P. Synergy of biofuel production with waste remediation along with value-added co-products recovery through microalgae cultivation: A review of membrane-integrated green approach. Sci. Total Environ. 2020, 698, 134169. [Google Scholar] [CrossRef]
- Zuorro, A.; Malavasi, V.; Cao, G.; Lavecchia, R. Use of cell wall degrading enzymes to improve the recovery of lipids from Chlorella sorokiniana. Chem. Eng. J. 2019, 377, 120325. [Google Scholar] [CrossRef]
- Mehariya, S.; Fratini, F.; Lavecchia, R.; Zuorro, A. Green extraction of value-added compounds from microalgae: A short review on natural deep eutectic solvents (NaDES) and related pre-treatments. J. Environ. Chem. Eng. 2021, 9, 105989. [Google Scholar] [CrossRef]
- Ferreira, G.F.; Ríos Pinto, L.F.; Maciel Filho, R.; Fregolente, L.V. Effects of cultivation conditions on Chlorella vulgaris and Desmodesmus sp. grown in sugarcane agro-industry residues. Bioresour. Technol. 2021, 342, 125949. [Google Scholar] [CrossRef]
- Wang, Y.; Guo, W.; Yen, H.W.; Ho, S.H.; Lo, Y.C.; Cheng, C.L.; Ren, N.; Chang, J.-S. Cultivation of Chlorella vulgaris JSC-6 with swine wastewater for simultaneous nutrient/COD removal and carbohydrate production. Bioresour. Technol. 2015, 198, 619–625. [Google Scholar] [CrossRef]
- Belotti, G.; Bravi, M.; de Caprariis, B.; de Filippis, P.; Scarsella, M. Effect of nitrogen and phosphorus starvations on Chlorella vulgaris lipids productivity and quality under different trophic regimens for biodiesel production. Am. J. Plant Sci. 2013, 04, 44–51. [Google Scholar] [CrossRef]
- Recht, L.; Zarka, A.; Boussiba, S. Patterns of carbohydrate and fatty acid changes under nitrogen starvation in the microalgae Haematococcus pluvialis and Nannochloropsis sp. Appl. Microbiol. Biotechnol. 2012, 94, 1495–1503. [Google Scholar] [CrossRef] [PubMed]
- Uslu, L.; Işik, O.; Koç, K.; Göksan, T. The effects of nitrogen deficiencies on the lipid and protein contents of Spirulina platensis. Afr. J. Biotechnol. 2011, 10, 386–389. [Google Scholar]
- Oslan, S.N.H.; Tan, J.S.; Oslan, S.N.; Matanjun, P.; Mokhtar, R.A.M.; Shapawi, R.; Huda, N. Haematococcus pluvialis as a potential source of astaxanthin with diverse applications in industrial sectors: Current research and future directions. Molecules 2021, 26, 6470. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Cai, C.; Yang, C.; Zheng, Z.; Wang, Q.; Du, X.; Deng, Y. Effect of protein sources in formulated diets on the growth, immune response, and intestinal microflora of pearl oyster Pinctada fucata martensii. Aquac. Rep. 2020, 16, 100253. [Google Scholar] [CrossRef]
- Hua, K.; Cobcroft, J.M.; Cole, A.; Condon, K.; Jerry, D.R.; Mangott, A.; Praeger, C.; Vucko, M.J.; Zeng, C.; Zenger, K.; et al. The future of aquatic protein: Implications for protein sources in aquaculture diets. One Earth 2019, 1, 316–329. [Google Scholar] [CrossRef]
- Kissinger, K.R.; García-Ortega, A.; Trushenski, J.T. Partial fish meal replacement by soy protein concentrate, squid and algal meals in low fish-oil diets containing Schizochytrium limacinum for longfin yellowtail Seriola rivoliana. Aquaculture 2016, 452, 37–44. [Google Scholar] [CrossRef]
- Vizcaíno, A.J.; López, G.; Sáez, M.I.; Jiménez, J.A.; Barros, A.; Hidalgo, L.; Camacho-Rodríguez, J.; Martínez, T.; Cerón-García, M.; Alarcón, F. Effects of the microalga Scenedesmus almeriensis as fishmeal alternative in diets for gilthead sea bream, Sparus aurata, juveniles. Aquaculture 2014, 431, 34–43. [Google Scholar] [CrossRef]
- Yang, C.; Hao, R.; Du, X.; Wang, Q.; Deng, Y.; Sun, R. Response to different dietary carbohydrate and protein levels of pearl oysters (Pinctada fucata martensii) as revealed by GC–TOF/MS-based metabolomics. Sci. Total Environ. 2019, 650, 2614–2623. [Google Scholar] [CrossRef]
- Flaak, A.R.; Epifanio, C.E. Dietary protein levels and growth of the oyster Crassostrea virginica. Mar. Biol. 1978, 45, 157–163. [Google Scholar] [CrossRef]
- Utting, S.D. A preliminary study on growth of Crassostrea gigas larvae and spat in relation to dietary protein. Aquaculture 1986, 56, 123–138. [Google Scholar] [CrossRef]
- Whyte, J.N.C.; Bourne, N.; Hodgson, C.A. Influence of algal diets on biochemical composition and energy reserves in Patinopecten yessoensis (Jay) larvae. Aquaculture 1989, 78, 333–347. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Initial Density (g L−1) | Biomass Productivity (g L−1 day−1) | TN Removal Efficiency (%) | TP Removal Efficiency (%) | |
|---|---|---|---|---|
| CV | 0.031 | 0.069 ± 0.012 b | 79.61 ± 2.61 b | 74.95 ± 4.58 b |
| SP | 0.046 | 0.086 ± 0.001 c | 82.89 ± 0.35 c | 78.21 ± 1.80 c |
| HP | 0.038 | 0.057 ± 0.006 a | 41.73 ± 2.09 a | 29.05 ± 3.75 a |
| C. vulgaris | S. platensis | H. pluvialis | |
|---|---|---|---|
| Pro | 1.2 ± 0.46 | 3.6 ± 1.04 | 0.2 ± 0.63 |
| Leucine * | 1.4 ± 0.24 | 2.6 ± 0.59 | 1.9 ± 0.95 |
| Valine * | 2.3 ± 0.30 | 3.0 ± 0.23 | 0.9 ± 0.13 |
| Lysine * | 3.0 ± 0.72 | 1.4 ± 0.31 | 1.2 ± 0.40 |
| Phenylalanine * | 2.9 ± 0.92 | 3.4 ± 0.62 | 0.6 ± 0.45 |
| Tyrosine | 3.2 ± 0.81 | 3.3 ± 1.01 | 0.5 ± 0.04 |
| Methionine * | 2.1± 0.29 | 2.3 ± 0.56 | 0.3 ± 0.17 |
| Threonine * | 1.8 ± 0.05 | 2.1 ± 0.16 | 0.7 ± 0.33 |
| Cysteine | 0.6 ± 0.09 | 1.0 ± 0.04 | ND |
| Aspartic acid | 2.9 ± 0.03 | 0.8 ± 0.05 | 2.0 ± 0.04 |
| Tryptophan | 2.0 ± 0.63 | 0.4 ± 0.11 | 0.4 ± 0.15 |
| Alanine | 2.4 ± 0.40 | 3.1 ± 0.07 | 1.3 ± 0.09 |
| Arginine | 3.2 ± 0.69 | 4.3 ± 1.05 | 1.0 ± 0.03 |
| Glutamic acid | 3.8 ± 1.08 | 4.8 ± 0.54 | 2.2 ± 0.75 |
| Histidine | 2.0 ± 0.07 | 2.1 ± 0.09 | ND |
| Glycine | 3.9 ± 0.10 | 5.6 ± 1.03 | 1.1 ± 0.05 |
| Isoleucine * | 2.6 ± 0.98 | 1.0 ± 0.82 | 0.6 ± 0.01 |
| Serine | 1.2 ± 0.20 | 3.1 ± 1.08 | 0.2 ± 0.07 |
| Fatty Acid | C. vulgaris | S. platensis | H. pluvialis |
|---|---|---|---|
| Myristic Acid (14:0) | 0.122 ± 0.03 | 0.145 ± 0.06 | 0.093 ± 0.01 |
| Palmitic Acid (16:0) | 0.439 ± 0.52 | 0.521 ± 0.10 | 0.314 ± 0.93 |
| Palmitoleic Acid (16:1) | 0.015 ± 0.07 | 0.012 ± 0.09 | 0.003 ± 0.13 |
| Oleic Acid (18:1) | 0.237 ± 0.11 | 0.265 ± 0.23 | 0.157 ± 0.19 |
| Stearic Acid (18:0) | 0.024 ± 0.04 | 0.034 ± 0.08 | 0.031 ± 0.21 |
| Linoleic Acid (18:2, ω -6) | 0.481 ± 0.28 | 0.460 ± 0.33 | 0.527 ± 0.90 |
| Arachidonic Acid (20:0) | 0.033 ± 0.17 | 0.042 ± 0.54 | 0.045 ± 0.16 |
| γ-Linolenic Acid (18:2) | 0.440 ± 0.80 | 0.361 ± 0.39 | 0.198 ± 0.06 |
| Linoelaidic acid | ND | 0.001 ± 0.02 | 0.003 ± 0.13 |
| Pentadecanoic acid | 0.005 ± 0.12 | 0.003 ± 0.04 | 0.002 ± 0.01 |
| Erucic Acid | ND | ND | ND |
| Docosadienoic Acid (22:6, ω -3) | 0.020 ± 0.01 | 0.018 ± 0.11 | 0.025 ± 0.56 |
| Eicosatrienoic Acid (20:5 ω -3) | ND | 0.003 ± 0.05 | 0.001 ± 0.03 |
| Total saturated | 0.618 | 0.742 | 0.483 |
| Total unsaturated | 1.193 | 1.12 | 0.914 |
| Index | C. vulgaris | S. platensis | H. pluvialis |
|---|---|---|---|
| IW (g) | 2.135 | 2.135 | 2.135 |
| FW (g) | 2.87 ± 0.103 c | 2.77 ± 0.19 bc | 2.45 ± 0.10 a |
| WG (%) | 34.58 ± 4.82 c | 29.89 ± 8.90 bc | 14.91 ± 3.94 a |
| SGR(%/day) | 0.99 ± 0.12 c | 0.87 ± 0.23 bc | 0.46 ± 0.11 a |
| ISL (mm) | 29.39 | 29.39 | 29.39 |
| FSL (mm) | 32.64 ± 1.42 c | 30.91 ± 0.93 abc | 31.65 ± 0.80 abc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amenorfenyo, D.K.; Li, F.; Zhang, Y.; Li, C.; Zhang, N.; Huang, X. Effects of Microalgae Grown in Membrane Treated Distillery Wastewater as Diet on Growth and Survival Rate of Juvenile Pearl Oyster (Pinctada fucata martensii). Water 2022, 14, 2702. https://doi.org/10.3390/w14172702
Amenorfenyo DK, Li F, Zhang Y, Li C, Zhang N, Huang X. Effects of Microalgae Grown in Membrane Treated Distillery Wastewater as Diet on Growth and Survival Rate of Juvenile Pearl Oyster (Pinctada fucata martensii). Water. 2022; 14(17):2702. https://doi.org/10.3390/w14172702
Chicago/Turabian StyleAmenorfenyo, David Kwame, Feng Li, Yulei Zhang, Changling Li, Ning Zhang, and Xianghu Huang. 2022. "Effects of Microalgae Grown in Membrane Treated Distillery Wastewater as Diet on Growth and Survival Rate of Juvenile Pearl Oyster (Pinctada fucata martensii)" Water 14, no. 17: 2702. https://doi.org/10.3390/w14172702
APA StyleAmenorfenyo, D. K., Li, F., Zhang, Y., Li, C., Zhang, N., & Huang, X. (2022). Effects of Microalgae Grown in Membrane Treated Distillery Wastewater as Diet on Growth and Survival Rate of Juvenile Pearl Oyster (Pinctada fucata martensii). Water, 14(17), 2702. https://doi.org/10.3390/w14172702