
Strawberry Growth under Current and Future Rainfall Scenarios

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Methods
2.1. Experimental Design
2.2. Volumetric Water Content, Soil Temperature, Leachate and Nitrate-Leachate Concentration
2.3. Disease
2.4. Leaf Chlorophyll Fluorescence and Chlorophyll Concentration
2.5. Biomass
2.6. Analyses
3. Results
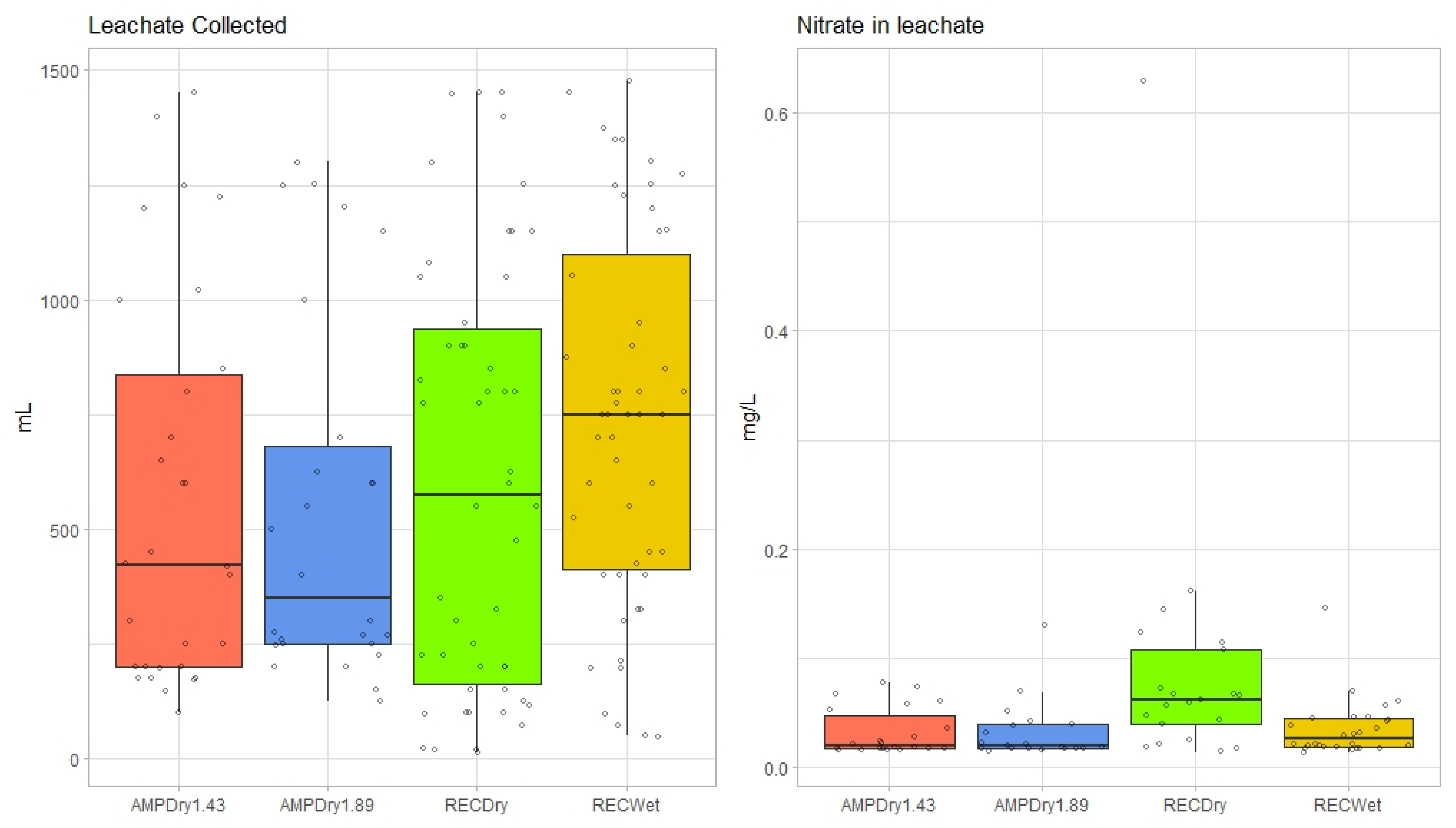
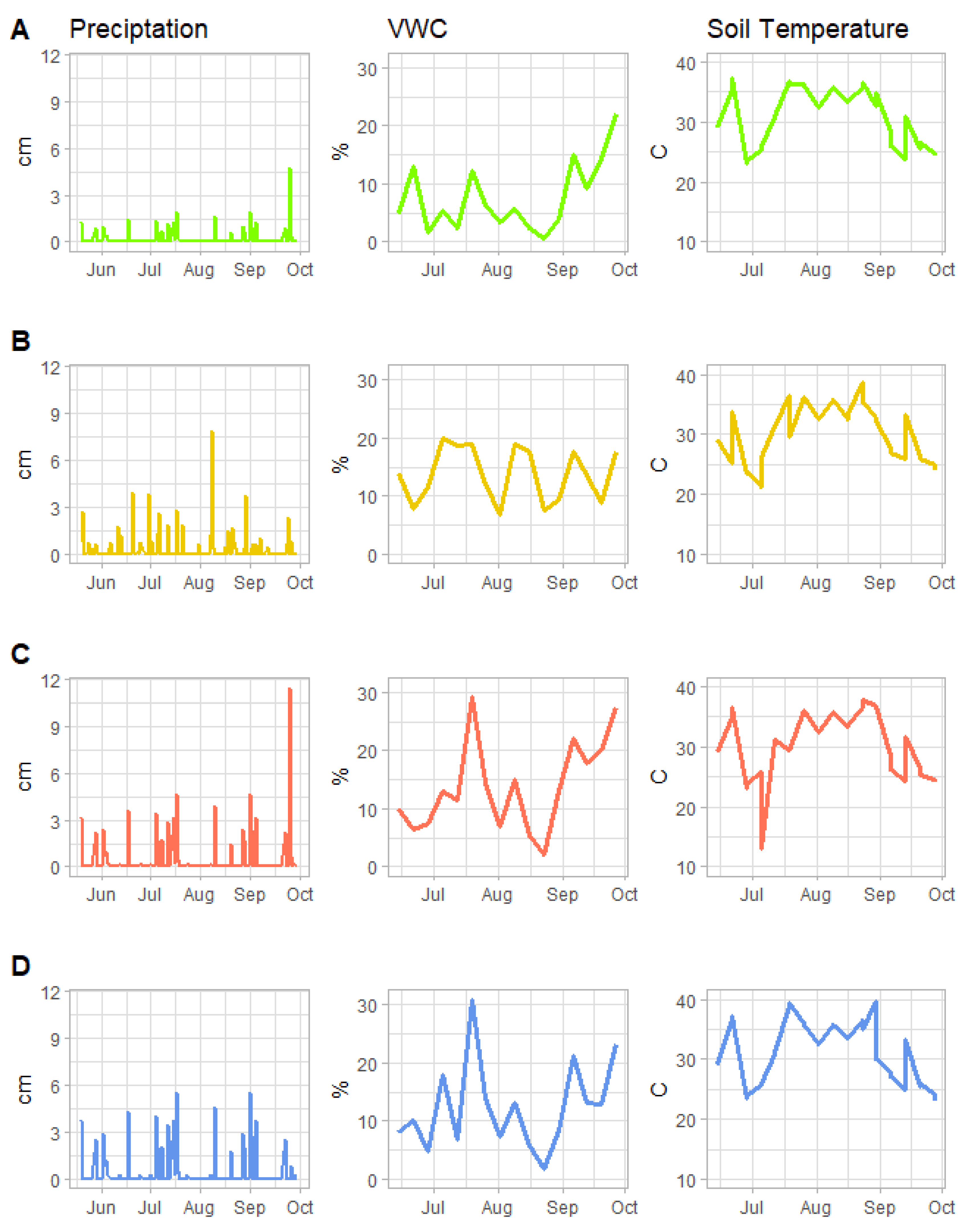
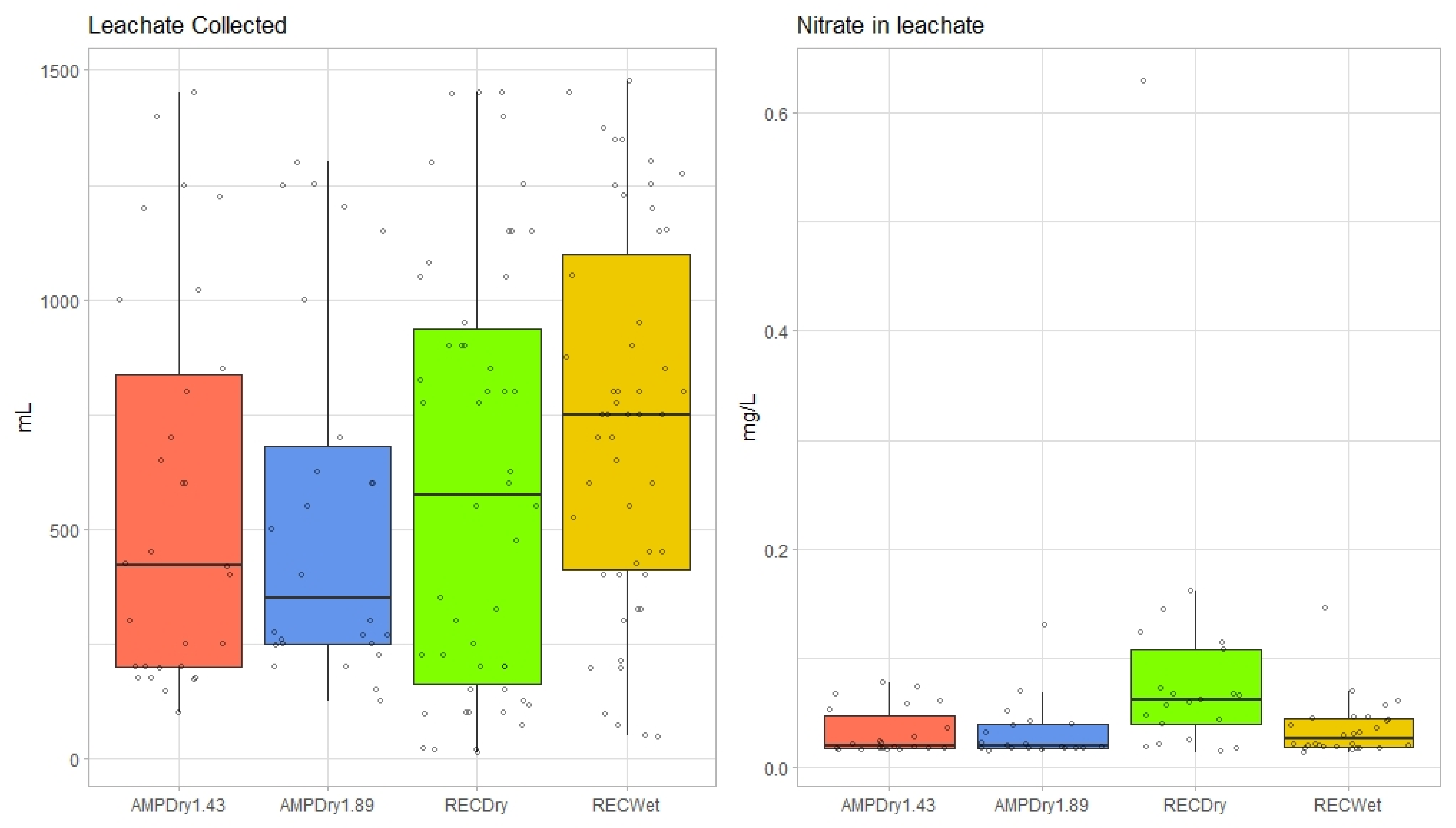
3.1. Volumetric Water Content, Soil Temperature, Leachate and Nitrate-Leachate Concentration
3.2. Disease
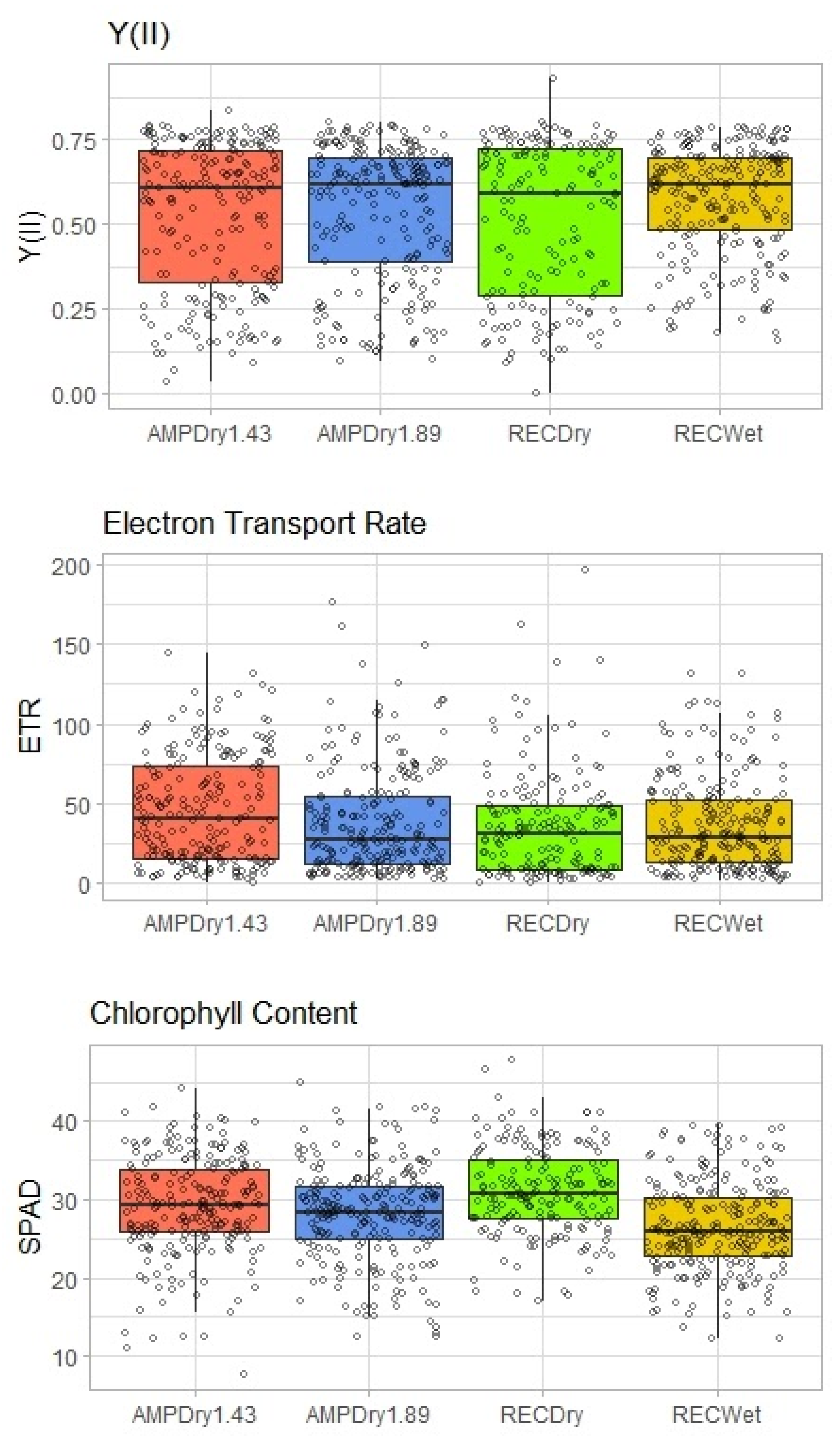
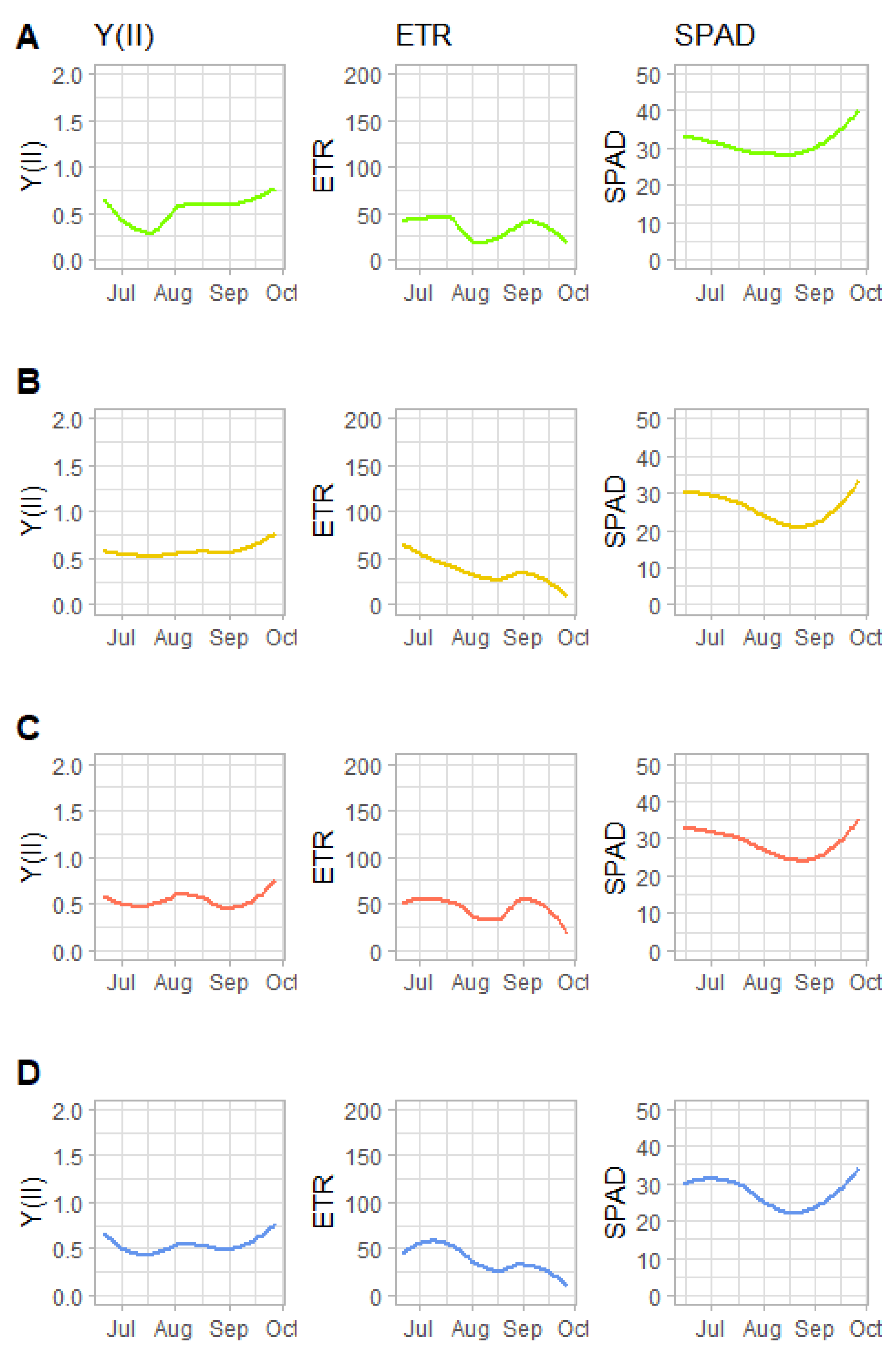
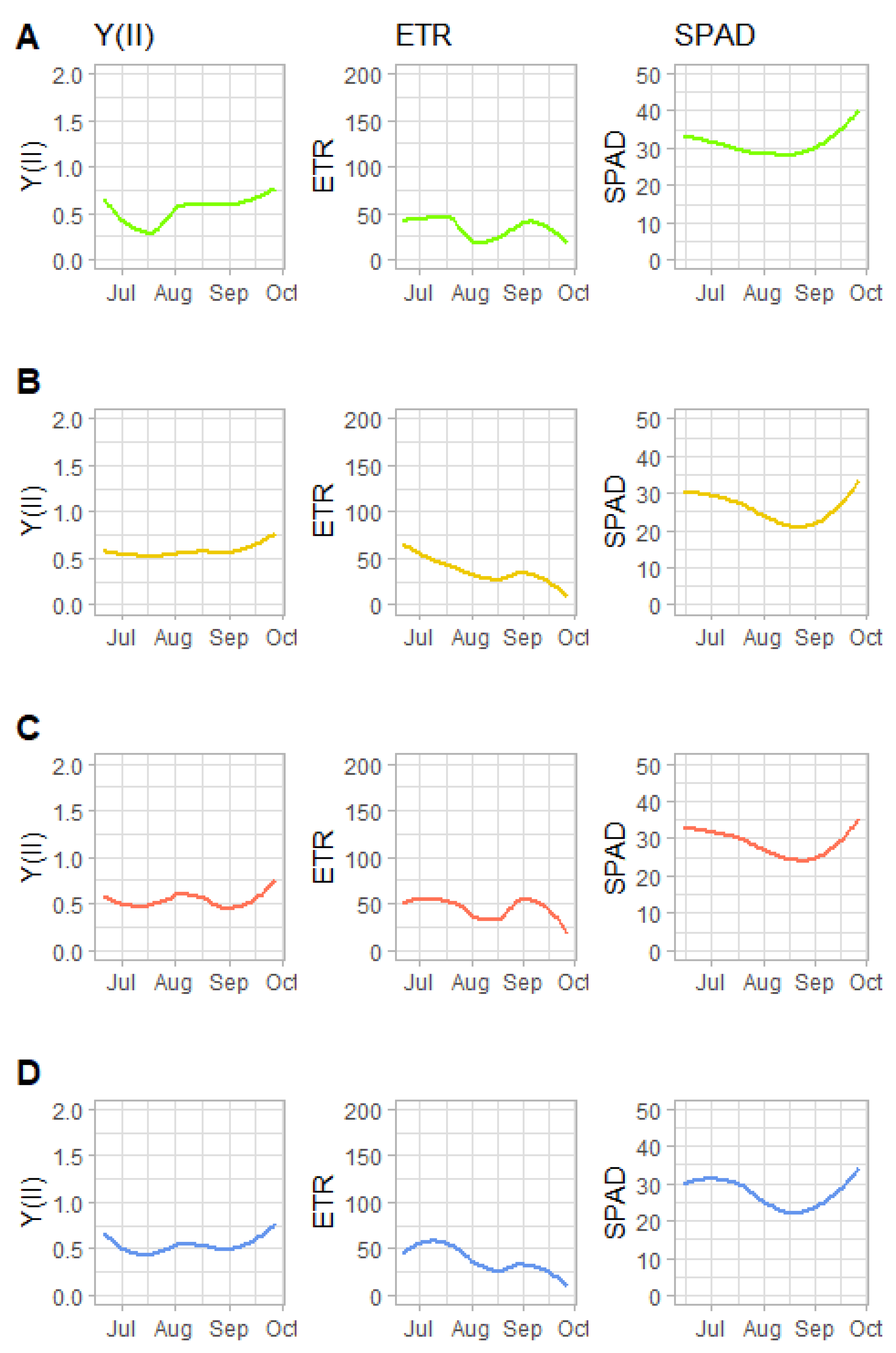
3.3. Leaf Chlorophyll Fluorescence and Chlorophyll Concentration
3.4. Biomass
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Samtani, J.B.; Rom, C.R.; Friedrich, H.; Fennimore, S.A.; Finn, C.E.; Petran, A.; Wallace, R.W.; Pritts, M.P.; Fernandez, G.; Chase, C.A.; et al. The status and future of the strawberry industry in the United States. HortTechnology 2019, 29, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Palencia, P.; Martínez, F.; Medina, J.J.; López-Medina, J. Strawberry yield efficiency and its correlation with temperature and solar radiation. Hortic. Bras. 2013, 31, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Ariza, M.T.; Miranda, L.; Gómez-Mora, J.A.; Medina, J.J.; Lozano, D.; Gavilán, P.; Soria, C.; Martínez-Ferri, E. Yield and Fruit Quality of strawberry cultivars under different irrigation regimes. Agronomy 2021, 11, 261. [Google Scholar] [CrossRef]
- Sidemane, B. (Ed.) New England Small Fruit Management Guide, 2019–2020th ed.; University of Massachusetts Extension: Amherst, MA, USA, 2019. [Google Scholar]
- Orde, K.; Sideman, B.; Pritts, M.; Demchak, K. Low Tunnel Strawberry Production Guide; University of New Hampshire Cooperative Extension: Durham, NH, USA, 2018. [Google Scholar]
- IPCC. Climate Change 2021: The Physical Science Basis; United Nations Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2021.
- Kundzewicz, Z.W.; Mata, L.J.; Arnell, N.W.; Döll, P.; Jimenez, B.; Miller, K.; Oki, T.; Şen, Z.; Shiklomanov, I. The implications of projected climate change for freshwater resources and their management. Hydrol. Sci. J. 2008, 53, 3–10. [Google Scholar] [CrossRef]
- Döll, P. Vulnerability to the impact of climate change on renewable groundwater resources: A global-scale assessment. Environ. Res. Lett. 2009, 4, 035006. [Google Scholar] [CrossRef]
- Kundzewicz, Z.W.; Döll, P. Will groundwater ease freshwater stress under climate change? Hydrol. Sci. J. 2009, 54, 665–675. [Google Scholar] [CrossRef] [Green Version]
- Kunkel, K.E.; Stevens, L.E.; Stevens, S.E.; Sun, L.; Janssen, E.; Wuebbles, D.; Rennells, J.; DeGaetano, A.; Dobson, J.G. Regional Climate Trends and Scenarios for the U.S. National Climate Assessment. Part 1. Climate of the Northeast U.S.; U.S. Department of Commerce: Washington, DC, USA, 2013; pp. 1–87. [Google Scholar]
- Huang, H.; Winter, J.M.; Osterberg, E.C.; Horton, R.M.; Beckage, B. Total and extreme precipitation changes over the Northeastern United States. J. Hydrometeorol. 2017, 18, 1783–1798. [Google Scholar] [CrossRef] [PubMed]
- Easterling, D.R.; Arnold, J.R.; Knutson, T.; Kunkel, K.E.; LeGrande, A.N.; Leung, L.R.; Vose, R.S.; Waliser, D.E.; Wehner, M.F. Chapter 7: Precipitation Change in the United States. Climate Science Special Report: Fourth National Climate Assessment, Volume I; U.S. Global Change Research Program: Washington, DC, USA, 2017. [Google Scholar]
- Walsh, J.; Wuebbles, D.; Hayhoe, K. Our Changing Climate; Melillo, J., Richmont, T., Yohe, G., Eds.; U.S. Global Change Research Program: Washington, DC, USA, 2014; p. 67. [Google Scholar]
- Wolfe, D.W.; DeGaetano, A.T.; Peck, G.M.; Carey, M.; Ziska, L.H.; Lea-Cox, J.; Kemanian, A.R.; Hoffmann, M.P.; Hollinger, D.Y. Unique challenges and opportunities for Northeastern US crop production in a changing climate. Clim. Chang. 2018, 146, 231–245. [Google Scholar] [CrossRef] [Green Version]
- Schattman, R.E.; Niles, M.T.; Aitken, H.M. Water use governance in a temperate region: Implications for agricultural climate change adaptation in the Northeastern United States. Ambio 2021, 50, 942–955. [Google Scholar] [CrossRef] [PubMed]
- Slatyer, R.O. Physiological significance of internal water relations to crop yield. In Physiological Aspects of Crop Yield; Eastin, J.D., Haskins, F.A., Sullivan, C.Y., van Bavel, C.H.M., Eds.; American Society of Agronomy, Crop Science Society of America: Madison, WI, USA, 1969; pp. 53–83. ISBN 978-0-89118-571-0. [Google Scholar]
- Palencia, P.; Martínez, F.; Medina-M’inguez, J.; Vázquez, E.; Flores, F.; López-Medina, J. Effects of climate change on strawberry production. Acta Hortic. 2009, 838, 51–53. [Google Scholar] [CrossRef]
- Barai, K.; Tasnim, R.; Hall, B.; Rahimzadeh-Bajgiran, P.; Zhang, Y.-J. Is drought increasing in maine and hurting wild blueberry production? Climate 2021, 9, 178. [Google Scholar] [CrossRef]
- Ahmed, S.; Stepp, J.R. Beyond yields: Climate change effects on specialty crop quality and agroecological management. Elem. Sci. Anthropocene 2016, 4, 000092. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Dey, P. Effects of different mulches and irrigation methods on root growth, nutrient uptake, water-use efficiency and yield of strawberry. Sci. Hortic. 2011, 127, 318–324. [Google Scholar] [CrossRef]
- May, G.M.; Pritts, M.P.; Kelly, M.J. Seasonal patterns of growth and tissue nutrient content in strawberries. J. Plant Nutr. 1994, 17, 1149–1162. [Google Scholar] [CrossRef]
- Silva-Pérez, V.; De Faveri, J.; Molero, G.; Deery, D.M.; Condon, A.G.; Raynolds, M.P.; Evans, J.R.; Furbank, R.T. Genetic variation for photosynthetic capacity and efficiency in spring wheat. J. Exp. Bot. 2020, 71, 2299–2311. [Google Scholar] [CrossRef]
- Ghimire, B.; Timsina, D.; Nepal, J. Analysis of chlorophyll content and its correlation with yield attributing traits on early varieties of maize (Zea mays L.). J. Maize Res. Dev. 2015, 1, 134–145. [Google Scholar] [CrossRef]
- Datt, B. A new reflectance index for remote sensing of chlorophyll content in higher plants: Tests using eucalyptus leaves. J. Plant Physiol. 1999, 154, 30–36. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Gritz, Y.; Merzlyak, M.N. Relationships between leaf chlorophyll content and spectral reflectance and algorithms for non-destructive chlorophyll assessment in higher plant leaves. J. Plant Physiol. 2003, 160, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Yıldırım, M.; Kılıç, H.; Kendal, E.; Karahan, T. Applicability of chlorophyll meter readings as yield predictor in durum wheat. J. Plant Nutr. 2010, 34, 151–164. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Babani, F. Detection of photosynthetic activity and water stress by imaging the red chlorophyll fluorescence. Plant Physiol. Biochem. 2000, 38, 889–895. [Google Scholar] [CrossRef]
- Fierer, N.; Schimel, J.P. Effects of drying–rewetting frequency on soil carbon and nitrogen transformations. Soil Biol. Biochem. 2002, 34, 777–787. [Google Scholar] [CrossRef]
- Cabrera, M.L. Modeling the flush of nitrogen mineralization caused by drying and rewetting soils. Soil Sci. Soc. Am. J. 1993, 57, 63–66. [Google Scholar] [CrossRef]
- Hess, L.J.T.; Hinckley, E.-L.S.; Robertson, G.P.; Matson, P.A. Rainfall intensification increases nitrate leaching from tilled but not no-till cropping systems in the U.S. Midwest. Agric. Ecosyst. Environ. 2020, 290, 106747. [Google Scholar] [CrossRef]
- Van Gestel, M.; Merckx, R.; Vlassak, K. Microbial biomass responses to soil drying and rewetting: The fate of fast-and slow-growing microorganisms in soils from different climates. Soil Biol. Biochem. 1993, 25, 109–123. [Google Scholar] [CrossRef]
- Sosnowski, M.R.; Ayres, M.R.; Scott, E.S. The Influence of water deficit stress on the grapevine trunk disease pathogens eutypa lata and diplodia seriata. Plant Dis. 2021, 105, 2217–2221. [Google Scholar] [CrossRef]
- Rossi, F.G.; Asalf, B.; Grieu, C.; Onofre, R.B.; Peres, N.A.; Gadoury, D.M.; Stensvand, A. Effect of water stress on reproduction and colonization of podosphaera aphanis of strawberry. Plant Dis. 2020, 104, 2973–2978. [Google Scholar] [CrossRef]
- Freeman, S.; Gnayem, N. Use of plasticulture for strawberry plant production. Small Fruits Rev. 2005, 4, 21–32. [Google Scholar] [CrossRef]
- Tasnim, R.; Zhang, Y.J. Are wild blueberries a crop with low photosynthetic capacity? Chamber-size effects in measuring photosynthesis. Agronomy 2021, 11, 1572. [Google Scholar] [CrossRef]
- Hirte, J.; Leifeld, J.; Abiven, S.; Oberholzer, H.-R.; Hammelehle, A.; Mayer, J. Overestimation of crop root biomass in field experiments due to extraneous organic matter. Front. Plant Sci. 2017, 8, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- USDA-NRCS. Above-Ground Biomass (Plant) Determinations; United States Department of Agriculture Natural Resources Conservation Service, National Soil Survey Center: Lincoln, NE, USA, 1997; pp. 1–2.
- McCullagh, P.; Nelder, J.A. Generalized Linear Models, 2nd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 1989. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern applied statistics with S. In Statistics and Computing, 4th ed.; Springer: New York, NY, USA, 2002; ISBN 978-0-387-95457-8. [Google Scholar]
- Mekis, E.; Hogg, W.D. Rehabilitation and analysis of Canadian daily precipitation time series. Atmos.-Ocean 1999, 37, 53–85. [Google Scholar] [CrossRef]
- Ng, V.K.Y.; Cribbie, R.A. Using the gamma generalized linear model for modeling continuous, skewed and heteroscedastic outcomes in psychology. Curr. Psychol. 2017, 36, 225–235. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Tjosvold, S.A. Soil Mixes Part 3: How Much Air and Water? University of California, Agriculture and Natural Resources: Berkeley, CA, USA, 2019. [Google Scholar]
- Yahdjian, L.; Sala, O.E. Size of precipitation pulses controls nitrogen transformation and losses in an arid patagonian ecosystem. Ecosystems 2010, 13, 575–585. [Google Scholar] [CrossRef]
- Hess, L.J.T.; Hinckley, E.-L.S.; Robertson, G.P.; Hamilton, S.K.; Matson, P.A. Rainfall intensification enhances deep percolation and soil water content in tilled and no-till cropping systems of the US Midwest. Vadose Zone J. 2018, 17, 180128. [Google Scholar] [CrossRef] [Green Version]
- Van Metre, P.C.; Frey, J.W.; Musgrove, M.; Nakagaki, N.; Qi, S.; Mahler, B.J.; Wieczorek, M.E.; Button, D.T. High nitrate concentrations in some Midwest United States streams in 2013 after the 2012 drought. J. Environ. Qual. 2016, 45, 1696–1704. [Google Scholar] [CrossRef] [Green Version]
- Razavi, F.; Pollet, B.; Steppe, K.; van Labeke, M.C. Chlorophyll fluorescence as a tool for evaluation of drought stress in strawberry. Photosynthetica 2008, 46, 631–633. [Google Scholar] [CrossRef]
- Flexas, J.; Escalona, J.M.; Medrano, H. Water stress induces different levels of photosynthesis and electron transport rate regulation in grapevines. Plant Cell Environ. 1999, 22, 39–48. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Escalona, J.M.; Sampol, B.; Medrano, H. Effects of drought on photosynthesis in grapevines under field conditions: An evaluation of stomatal and mesophyll limitations. Funct. Plant Biol. 2002, 29, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaderi, N.; Siosemardeh, A. Response to drought stress of two strawberry cultivars (cv. Kurdistan and Selva). Hortic. Environ. Biotechnol. 2011, 52, 6–12. [Google Scholar] [CrossRef]
- Mann, C.E.T. Studies in the root and shoot growth of the strawberry. V. The origin, development, and function of the roots of the cultivated strawberry (Fragaria Virginiana × Chiloensis). Ann. Bot. 1930, 44, 55–86. [Google Scholar] [CrossRef]
- Brouder, S.M.; Volenec, J.J. Impact of climate change on crop nutrient and water use efficiencies. Physiol. Plant. 2008, 133, 705–724. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ferri, E.; Soria, C.; Ariza, M.T.; Medina, J.J.; Miranda, L.; Domíguez, P.; Muriel, J.L. Water relations, growth and physiological response of seven strawberry cultivars (Fragaria × ananassa Duch.) to different water availability. Agric. Water Manag. 2016, 164, 73–82. [Google Scholar] [CrossRef]
- Savé, R.; Peñuelas, J.; Marfà, O.; Serrano, L. Changes in leaf osmotic and elastic properties and canopy structure of strawberries under mild water stress. HortScience 1993, 28, 925–927. [Google Scholar] [CrossRef] [Green Version]
- Tasnim, R.; Drummond, F.; Zhang, Y.J. Climate change patterns of wild blueberry fields in Downeast, maine over the past 40 years. Water 2021, 13, 594. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Description of Treatment Development |
|---|---|
| RECDry | Based on 2001 observations—infrequent rainfall separated by prolonged periods (2–5 weeks) of dryness. Daily rainfall above 2.54 cm (1 inch) occurred only once in an event of 4.70 cm (1.85 inches) on 25 September. Total rainfall April–September rainfall = 29.95 cm (11.79 inches). |
| RECWet | Based on 2019 observations—frequent, moderate-intensity rainfall events distributed throughout the growing season without any prolonged dryness. Rainfall exceeding 2.54 cm (1 inch) occurred on eight separate days. Total April–September rainfall = 72.85 cm (28.68 inches). |
| AMPDry1.43 | Modified from observed 2001 precipitation—daily values multiplied by 1.43 to increase total growing season rainfall to match that observed in 2019 (72.85 cm; 28.68 inches), while preserving the seasonal distribution observed in 2001. |
| AMPDry1.89 | Modified from observed 2001 precipitation—similar to AMPDry1.43, except that the large magnitude rainfall event 25 September was removed and the daily precipitation values were subsequential multiplied by 1.89 to attain the same season total in AMPDry1.43 and observed in 2019. |
| Model 1: Soil Moisture (kPa; Measured Weekly) | |||||
|---|---|---|---|---|---|
| Estimate (β) | Standard Error | t-Value | p-Value | CI (2.5%; 97.5%) | |
| RECDry | 0.111 | 0.007 | 14.95 | <0.001 | 0.098; 0.127 |
| RECWet | 0.073 | 0.005 | 15.81 | <0.001 | 0.063; 0.080 |
| AMPDry1.43 | 0.065 | 0.004 | 15.65 | <0.001 | 0.058; 0.074 |
| AMPDry1.89 | 0.073 | 0.005 | 15.81 | <0.001 | 0.064; 0.083 |
| Dispersion parameter = 0.416; null deviance = NaN, on 398 degrees of freedom; residual deviance = 191.47, on 394 degrees of freedom; AIC = 2718.9; number of Fisher scoring iterations = 6; R2 = 0.047. Note that kPa = 0 indicates soil water saturation and dryness increases as kPa increases. | |||||
| Model 2: Soil Temperature (Measured Weekly) | |||||
| Estimate (β) | Standard Error | t-Value | p-Value | CI (2.5%; 97.5%) | |
| RECDry | 0.032 | 0.001 | 34.16 | <0.001 | 0.031; 0.034 |
| RECWet | 0.033 | 0.001 | 34.16 | <0.001 | 0.031; 0.035 |
| AMPDry1.43 | 0.033 | 0.001 | 34.16 | <0.001 | 0.031; 0.035 |
| AMPDry1.89 | 0.032 | 0.001 | 34.16 | <0.001 | 0.030; 0.034 |
| Dispersion parameter = 0.027; null deviance = NaN, on 128 degrees of freedom; residual deviance = 3.712, on 124 degrees of freedom; AIC = 793.79; number of Fisher scoring iterations = 4; R2 = −0.026. | |||||
| Model 3: N-Leachate (Collected Weekly; mg/L) | |||||
|---|---|---|---|---|---|
| Estimate (β) | Standard Error | t-Value | p-Value | CI (2.5%; 97.5%) | |
| RECDry | 10.720 | 22.208 | 44.855 | <0.001 | 5.628; 15.838 |
| RECWet | 28.543 | 5.092 | 5.608 | <0.001 | 17.657; 39.554 |
| AMPDry1.43 | 31.030 | 6.970 | 4.970 | <0.001 | 17.366; 44.992 |
| AMPDry1.89 | 31.397 | 6.627 | 4.738 | <0.001 | 16.803; 46.230 |
| Dispersion parameter = 0.866; null deviance = NaN, on 91 degrees of freedom; residual deviance = 44.913, on 87 degrees of freedom; AIC = −409.81; number of Fisher scoring iterations = 7; adjusted R2 = 0.075. | |||||
| Model 4: Leachate (Collected at Each Precipitation Event; mL) | |||||
| Estimate (β) | Standard Error | t-Value | p-Value | CI (2.5%; 97.5%) | |
| RECDry | 0.001 | 0.001 | 5.415 | <0.001 | 0.001; 0.001 |
| RECWet | < 0.000 | < 0.000 | 5.982 | <0.001 | 0.001; 0.001 |
| AMPDry1.43 | < 0.000 | < 0.000 | 5.552 | <0.001 | 0.001; 0.001 |
| AMPDry1.89 | < 0.000 | < 0.000 | 5.322 | <0.001 | <0.000; 0.001 |
| Dispersion parameter = 1.436; null deviance = NaN, on 250 degrees of freedom; residual deviance = 379.51, on 246 degrees of freedom; AIC = 4342.5; number of Fisher scoring iterations = 7; adjusted R2 = < −0.000. | |||||
| Model 5: Y(II) Measured Weekly | |||||
|---|---|---|---|---|---|
| Estimate (β) | Standard Error | t-Value | p-Value | CI (2.5%; 97.5%) | |
| RECDry | 1.937 | 0.057 | 34.33 | <0.001 | 1.829; 2.050 |
| RECWet | 1.734 | 0.043 | 40.44 | <0.001 | 1.652; 1.820 |
| AMPDry1.43 | 1.893 | 0.048 | 39.19 | <0.001 | 1.800; 2.000 |
| AMPDry1.89 | 1.863 | 0.046 | 40.17 | <0.001 | 1.774; 1.955 |
| Dispersion parameter = 0.140; null deviance = NaN, on 835 degrees of freedom; residual deviance = 170.48 on 831 degrees of freedom; AIC = −115.11; number of Fisher scoring iterations = 5; adjusted R2 = 0.004. | |||||
| Model 6: ETR Measured Weekly | |||||
| Estimate (β) | Standard Error | t-Value | p-Value | CI (2.5%; 97.5%) | |
| RECDry | 0.030 | 0.002 | 15.69 | <0.001 | 0.024; 0.030 |
| RECWet | 0.027 | 0.001 | 18.48 | <0.001 | 0.025; 0.030 |
| AMPDry1.43 | 0.022 | 0.001 | 17.91 | <0.001 | 0.019; 0.024 |
| AMPDry1.89 | 0.026 | 0.001 | 18.48 | <0.001 | 0.023; 0.029 |
| Dispersion parameter = 0.671; null deviance = NaN, on 835 degrees of freedom; residual deviance = 656.95, on 831 degrees of freedom; AIC = 7758.5; number of Fisher scoring iterations = 6; adjusted R2 = 0.006. | |||||
| Model 7: Chlorophyll Concentration (SPAD) Measured Weekly | |||||
| Estimate (β) | Standard Error | t-Value | p-Value | CI (2.5%; 97.5%) | |
| RECDry | 0.032 | 0.001 | 60.73 | <0.001 | 0.032; 0.034 |
| RECWet | 0.038 | 0.001 | 70.80 | <0.001 | 0.038; 0.040 |
| AMPDry1.43 | 0.035 | 0.001 | 68.61 | <0.001 | 0.035; 0.037 |
| AMPDry1.89 | 0.036 | 0.001 | 70.80 | <0.001 | 0.038; 0.040 |
| Dispersion parameter = 0.671; null deviance = NaN, on 899 degrees of freedom; residual deviance = 5757.425, on 895 degrees of freedom; AIC = 66,049; number of Fisher scoring iterations = 44; adjusted R2 = 0.070070. | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schattman, R.E.; Smart, A.; Birkel, S.; Jean, H.; Barai, K.; Zhang, Y.-J. Strawberry Growth under Current and Future Rainfall Scenarios. Water 2022, 14, 313. https://doi.org/10.3390/w14030313
Schattman RE, Smart A, Birkel S, Jean H, Barai K, Zhang Y-J. Strawberry Growth under Current and Future Rainfall Scenarios. Water. 2022; 14(3):313. https://doi.org/10.3390/w14030313
Chicago/Turabian StyleSchattman, Rachel E., Alicyn Smart, Sean Birkel, Haley Jean, Kallol Barai, and Yong-Jiang Zhang. 2022. "Strawberry Growth under Current and Future Rainfall Scenarios" Water 14, no. 3: 313. https://doi.org/10.3390/w14030313
APA StyleSchattman, R. E., Smart, A., Birkel, S., Jean, H., Barai, K., & Zhang, Y.-J. (2022). Strawberry Growth under Current and Future Rainfall Scenarios. Water, 14(3), 313. https://doi.org/10.3390/w14030313