
Advances in the Fate of Rare Earth Elements, REE, in Transitional Environments: Coasts and Estuaries





{kind=link}
{kind=link}
Abstract
:1. Introduction
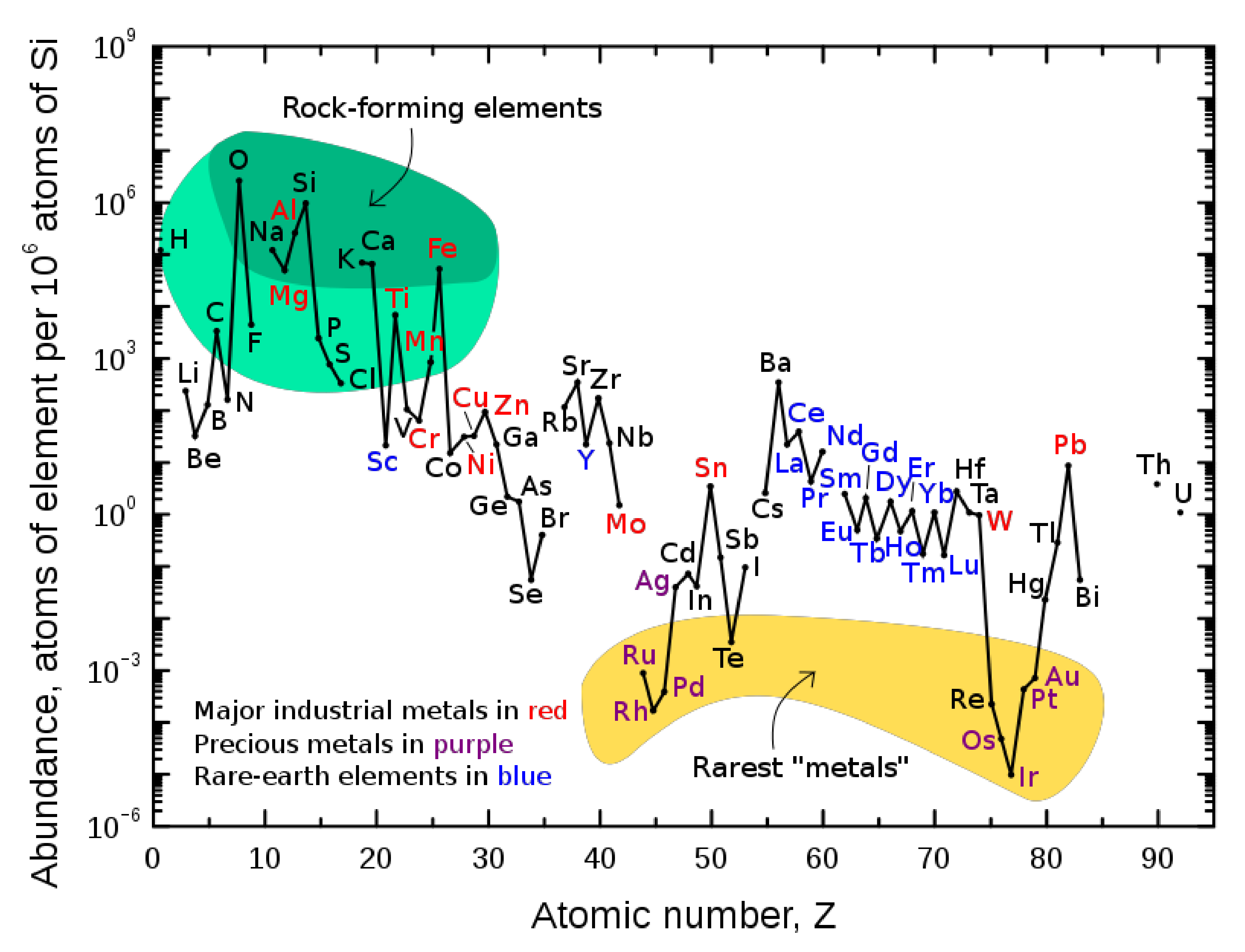
1.1. The Natural Abundance
1.2. The Sources of REE Contamination
2. Fate of REE in Riverine and Estuary System
2.1. Fractionation
2.1.1. Factors Controlling Fractionation of REE
2.1.2. Environmental Application of REE as Tracers of Pollution
3. The Bioaccumulation and Bioavailability of REE
3.1. Fish Species as Bioindicator of REE Pollution
3.2. Mechanisms of Bioavailability
3.3. Factors Influencing Bioavailability
4. The Toxicity of REE
4.1. The Hormetic Effect and the Weight of pH
4.2. REE Detoxification Routes
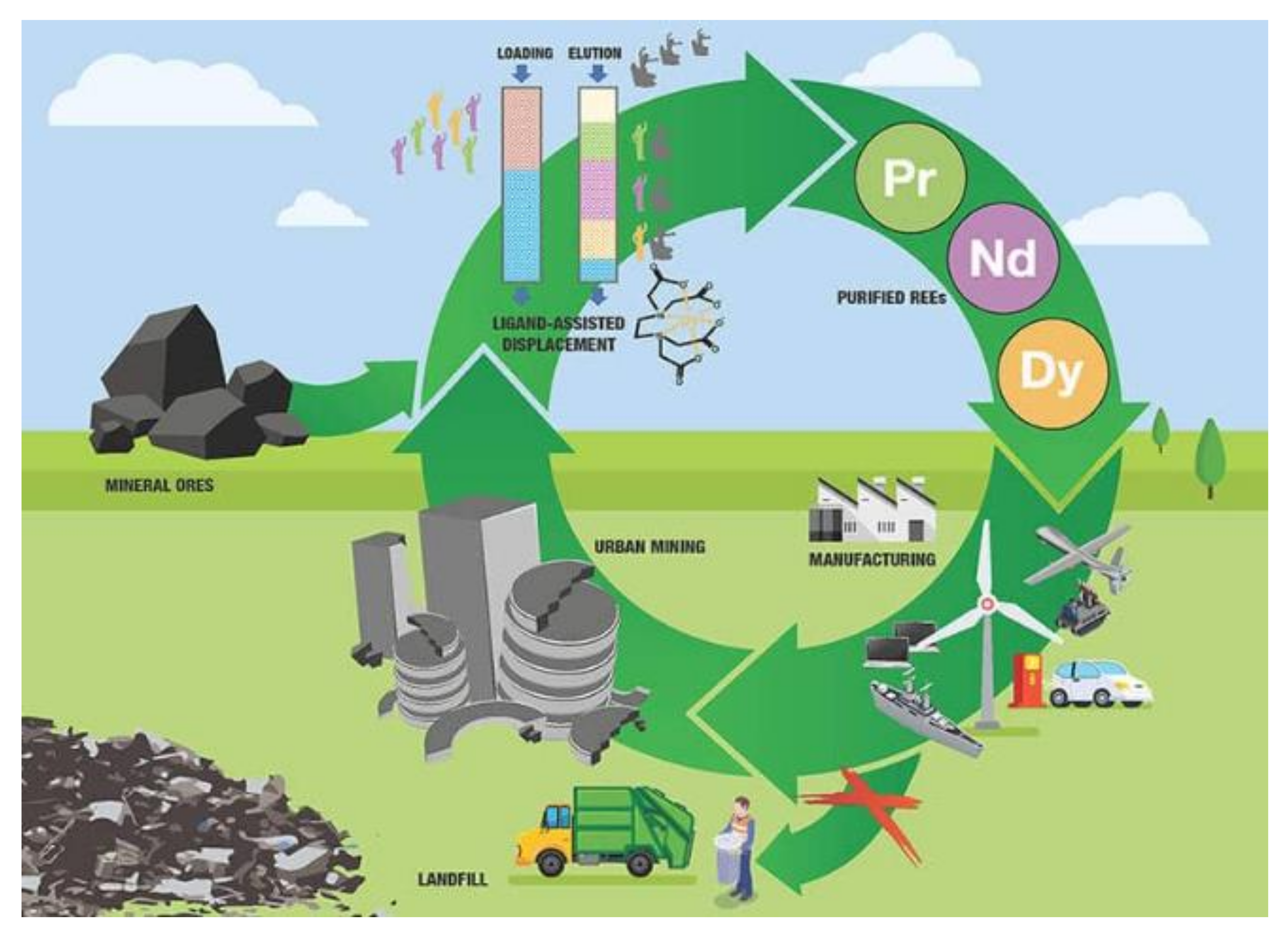
5. Reducing REE Fluxes
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klinger, J.M. A historical geography of rare earth elements: From discovery to the atomic age. Extrac. Ind. Soc. 2015, 2, 572–580. [Google Scholar] [CrossRef]
- Tan, Q.; Li, J.; Zeng, X. Rare earth elements recovery from waste fluorescent lamps: A review. Crit. Rev. Environ. Sci. Technol. 2015, 457, 749–776. [Google Scholar] [CrossRef]
- Ramos, S.J.; Dinali, G.S.; Oliveira, C.; Martins, G.C.; Moreira, C.G.; Siqueira, J.O.; Guilherme, L.R.G. Rare earth elements in the soil environment. Curr. Pollut. Rep. 2016, 21, 28–50. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.H. Rare Earth Coordination Chemistry: Fundamentals and Applications; John Wiley & Sons: Singapore, 2010; p. 592. [Google Scholar]
- Wood, S.A. The aqueous geochemistry of the rare-earth elements and yttrium: 1. Review of available low-temperature data for inorganic complexes and the inorganic REE speciation of natural waters. Chem. Geol. 1990, 82, 159–186. [Google Scholar] [CrossRef]
- Sneller, F.E.; Kalf, D.; Weltje, L.; Van Wezel, A. Maximum Permissible Concentrations and Negligible Concentrations for Rare Earth Elements (REEs); Report 601; Rivm: Bilthoven, The Netherlands, 2000; p. 66. [Google Scholar]
- Nockemann, P.; Thijs, B.; Postelmans, N.; Van Hecke, K.; Van Meervelt, L.; Binnemans, K. Anionic rare-earth thiocyanate complexes as building blocks for low-melting metal-containing ionic liquids. J. Am. Chem. Soc. 2006, 128, 13658–13659. [Google Scholar] [CrossRef] [PubMed]
- IUPAC. Nomenclature of Inorganic Chemistry. IUPAC Recommendations, 2005 (“Red Book”); RSC Publishing: Cambridge, UK, 2005; p. 366. [Google Scholar]
- Gwenzi, W.; Mangori, L.; Danha, C.; Chaukura, N.; Dunjana, N.; Sanganyado, E. Sources, behaviour, and environmental and human health risks of high-technology rare earth elements as emerging contaminants. Sci. Total Environ. 2018, 636, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Bru, K.; Christmann, P.; Labbé, J.F.; Lefebvre, G. Panorama 2014 du Marché des Terres Rares; BRGM/RP-65330; Cedex: Orleans, France, 2015. [Google Scholar]
- Atwood, D.A. The Rare Earth Elements. Fundamentals and Applications; John Wiley & Sons: West Sussex, UK, 2012; p. 624. [Google Scholar]
- Australian Industry Commission. New and Advanced Materials; Australian Government Publishing Service: Melbourne, Australia, 1995; p. 401. [Google Scholar]
- U.S. Geological Survey. Minerals Yearbook-Rare Earths. Available online: https://minerals.usgs.gov/minerals/pubs/commodity/rare_earths/mcs-2015-raree.pdf (accessed on 28 October 2021).
- International Union for Pure and Applied Chemistry (IUPAC). Nomenclature of Inorganic Chemistry: IUPAC Recommendations 2005. IUPAC Red Book; RSC Publishing: London, UK, 2005; ISBN 0-85404-438-8. [Google Scholar]
- de Baar, H.J.W.; German, C.R.; Elderfield, H.; van Gaans, P. Rare earth element distributions in anoxic waters of the Cariaco Trench. Geochem. Cosmochim. Acta 1988, 52, 1203–1219. [Google Scholar] [CrossRef]
- Elderfield, H.; Greaves, M.J. The rare earth elements in seawater. Nature 1982, 296, 214–219. [Google Scholar] [CrossRef]
- Deng, Y.; Ren, J.; Guo, Q.; Cao, J.; Wang, H.; Liu, C. Rare earth element geochemistry characteristics of seawater and porewater from deep sea in western Pacific. Sci. Rep. 2017, 7, 16539. [Google Scholar] [CrossRef] [Green Version]
- Massari, S.; Ruberti, M. Rare earth elements as critical raw materials: Focus on international markets and future strategies. Res. Policy 2013, 381, 36–43. [Google Scholar] [CrossRef]
- Metalpedia. Available online: http://metalpedia.asianmetal.com/metal/rare_earth/application.shtml (accessed on 15 November 2021).
- Li, C.R.; Zhuang, Z.Y.; Huang, F.; Wu, Z.C.; Hong, Y.P.; Lin, Z. Recycling rare earth elements from industrial wastewater with flowerlike nano-Mg(OH)2. ACS Appl. Mater. Interf. 2013, 5, 9719–9725. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, Z.; Chen, Z.; Zhang, Y. A human health risk assessment of rare earth elements in soil and vegetables from a mining area in Fujian Province, Southeast China. Chemosphere 2013, 93, 1240–1246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulick, H.; Machacek, E. The global rare earth element exploration boom: An analysis of resources outside of China and discussion of development perspectives. Resour. Policy 2017, 52, 134–153. [Google Scholar] [CrossRef]
- Tepe, N.; Romero, M.; Bau, M. High-technology metals as emerging contaminants: Strong increase of anthropogenic gadolinium levels in tap water of Berlin, Germany, from 2009 to 2012. Appl. Geochem. 2014, 45, 191–197. [Google Scholar] [CrossRef]
- Zhang, Y.; Gao, X. Rare earth elements in surface sediments of a marine coast under heavy anthropogenic influence: The Bohai Bay, China. Estuar. Coast. Shelf Sci. 2015, 164, 86–93. [Google Scholar] [CrossRef]
- Anastopoulos, I.; Bhatnagar, A.; Lima, E.C. Adsorption of rare earth metals: A review of recent literature. J. Mol. Liq. 2016, 221, 954–962. [Google Scholar] [CrossRef]
- Zhou, B.; Li, Z.; Zhao, Y.; Zhang, C.; Wei, Y. Rare earth elements supply vs. clean energy technologies: New problems to solve. Gospod. Surowcami Miner. 2016, 32, 29–44. [Google Scholar] [CrossRef] [Green Version]
- Alonso, E.; Sherman, A.M.; Wallington, T.J.; Everson, M.P.; Field, F.R.; Roth, R.; Kirchain, R.E. Evaluating rare earth element availability: A case with revolutionary demand from clean technologies. Environ. Sci. Technol. 2012, 46, 3406–3414. [Google Scholar] [CrossRef]
- Qu, K.M.; Xin, F.Y. Enhancement effects of three rare earth elements on the growth of Chaetoeeros mulleri. Mar. Sci. Bull. 2001, 3, 83–88. [Google Scholar]
- Cai, L.; Park, Y.S.; Seong, S.I.; Yoo, S.W.; Kim, I.H. Effects of rare earth elements-enriched yeast on growth performance, nutrient digestibility, meat quality, relative organ weight and excreta microflora in broiler chickens. Livest. Sci. 2015, 172, 43–49. [Google Scholar] [CrossRef]
- Migaszewski, Z.M.; Galuszka, A. The characteristics, occurrence, and geochemical behavior of rare earth elements in the environment: A review. Crit. Rev. Environ. Sci. Technol. 2015, 45, 429–471. [Google Scholar] [CrossRef]
- Liang, T.; Zhang, S.; Wang, L.; Kung, H.T.; Wang, Y.; Hu, A.; Ding, S. Environmental biogeochemical behaviors of rare earth elements in soil-plant systems. Environ. Geochem. Health 2005, 27, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Li, D.; Peng, A. Application of rare-earth elements in the agriculture of China and its environmental behavior in soil. Environ. Sci. Pollut. Res. 2002, 92, 143–148. [Google Scholar] [CrossRef] [PubMed]
- US-EPA. Rare Earth Elements: A Review of Production, Processing, Recycling, and Associated Environmental Issues; EPA/600/R-12/572; United States Environmental Protection Agency: Cincinnati, OH, USA, 2012. [Google Scholar]
- Adeel, M.; Lee, J.Y.; Zain, M.; Rizwan, M.; Nawab, A.; Ahmad, M.A.; Shafiq, M.; Yi, H.; Jilani, G.; Javed, R.; et al. Cryptic footprints of rare earth elements on natural resources and living organisms. Environ. Int. 2019, 127, 785–800. [Google Scholar] [CrossRef]
- King, H.M. REE-Rare Earth Elments and Their Uses. Available online: https://geology.com/articles/rare-earth-elements/ (accessed on 15 November 2021).
- MacMillan, G.A.; Chételat, J.; Heath, J.; Mickpegak, R.; Amyot, M. Rare earth elements (REE) in freshwater, marine, and terrestrial ecosystems in the eastern Canadian Arctic. Environ. Sci. Processes Impacts 2017, 19, 1336–1345. [Google Scholar] [CrossRef]
- Souza, I.C.; Morozesk, M.; Azevedo, V.C.; Mendes, V.A.S.; Duarte, I.D.; Rocha, L.D.; Matsumoto, S.T.; Elliott, M.; Baroni, M.V.; Wunderlin, D.A.; et al. Trophic transfer of emerging metallic contaminants in a neotropical mangrove ecosystem food web. J. Hazard. Mater. 2021, 408, 124424. [Google Scholar] [CrossRef]
- Lortholarie, M.; Zalouk-Vergnoux, A.; Couderc, M.; Kamari, A.; François, Y.; Herrenknecht, C. Rare earth element bioaccumulation in the yellow and silver European eel (Anguilla anguilla): A case study in the Loire estuary (France) Laurence Poirier. Sci. Total Environ. 2020, 719, 134938. [Google Scholar] [CrossRef]
- Négrel, P.; Guerrot, C.; Cocherie, A.; Azaroual, M.; Brach, M.; Fouillac, C. Rare earth elements, neodymium and strontium isotopic systematics in mineral waters: Evidence from the Massif Central, France. Appl. Geochem. 2000, 15, 1345–1367. [Google Scholar] [CrossRef]
- Vrel, A. Reconstitution de L’historique des Apports en Radionucléides et Contaminants Métalliques à L’estuaire Fluvial de la Seine Par L’analyse de Leur Enregistrement Sédimentaire. Ph.D. Thesis, University of Caen, Caen, France, 2012. [Google Scholar]
- Akagi, T.; Edanami, K. Sources of rare earth elements in shells and soft tissues of bivalves from Tokyo Bay. Mar. Chem. 2017, 194, 55–62. [Google Scholar] [CrossRef]
- Johannesson, K.H.; Palmore, C.D.; Fackrell, J.; Prouty, N.G.; Swarzenski, P.W.; Chevis, D.A.; Telfeyan, K.; White, C.D.; Burdige, D.J. Rare earth element behavior during groundwater–seawater mixing along the Kona Coast of Hawaii. Geochim. Cosmochim. Acta 2017, 198, 229–258. [Google Scholar] [CrossRef] [Green Version]
- Trifuoggi, M.; Donadio, C.; Ferrara, L.; Stanislao, C.; Toscanesi, M.; Arienzo, M. Levels of pollution of rare earth elements in the surface sediments from the Gulf of Pozzuoli (Campania, Italy). Mar. Poll. Bull. 2018, 136, 374–384. [Google Scholar] [CrossRef] [PubMed]
- Folens, K.; Du Laing, G. Dispersion and solubility of Ln, Tl, Ta and Nb in the aquatic environment and intertidal sediments of the Scheldt estuary (Flanders, Belgium). Chemosphere 2017, 183, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Censi, P.; Mazzola, S.; Sprovieri, M.; Bonanno, A.; Patti, B.; Punturo, R.; Spoto, S.E.; Saiano, F.; Alonzo, G. Rare earth elements distribution in seawater and suspended particulate of the central Mediterranean Sea. Chem. Ecol. 2004, 20, 323–343. [Google Scholar] [CrossRef]
- Khadijeh, R.S.E.; Saion, E.; Wood, A.K.; Reza, A.M. Rare earth elements distribution in marine sediments of Malaysia coasts. J. Rare Earths 2009, 27, 1066–1071. [Google Scholar] [CrossRef]
- Oliveri, E.; Neri, R.; Bellanca, A.; Riding, R. Carbonate stromatolites from a Messinian hypersaline setting in the Caltanissetta Basin, Sicily: Petrographic evidence of microbial activity and related stable isotope and rare earth element signatures. Sedimentology 2010, 57, 142–161. [Google Scholar] [CrossRef]
- Lerat-Hardy, A.; Coynel, A.; Dutruch, L.; Pereto, C.; Bossy, C.; Gil-Diaz, T.; Capdeville, M.J.; Blanc, G.; Schäfe, J. Rare Earth Element fluxes over 15 years into a major European Estuary (Garonne-Gironde, SW France): Hospital effluents as a source of increasing gadolinium anomalies. Sci. Total Environ. 2019, 656, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Kulaksız, S.; Bau, M. Anthropogenic dissolved and colloid/nanoparticle-bound samarium, lanthanum and gadolinium in the Rhine River and the impending destruction of the natural rare earth element distribution in rivers. Earth Planet. Sci. Lett. 2013, 362, 43–50. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Z.Z.; Wu, Q.; An, Y.; Jia, H.; Shen, Y. Anthropogenic rare earth elements: Gadolinium in small catchment in Guizhou Province, Southwwest China. Int. J. Environ. Res. Publ. Health 2019, 16, 4052. [Google Scholar] [CrossRef] [Green Version]
- Sholkovitz, E.R.; Szymczak, R. The estuarine chemistry of rare earth elements: Comparison of the Amazon, Fly, Sepik and the Gulf of Papua systems. Earth Planet. Sci. Lett. 2000, 179, 299–309. [Google Scholar] [CrossRef]
- Sholkovitz, E.R. Chemical evolution of rare earth elements: Fractionation between colloidal and solution phases of filtered river water. Earth Planet. Sci. Lett. 1992, 114, 77–84. [Google Scholar] [CrossRef]
- Evans, C.H. (Ed.) Episodes from the History of the Rare Earth Elements; Springer Science: Dordrecht, The Netherlands, 1996; p. 268. [Google Scholar]
- Zhou, B.; Li, Z.; Chen, C. Global potential of rare earth resources and rare earth demand from clean technologies. Minerals 2017, 7, 203. [Google Scholar] [CrossRef] [Green Version]
- Henderson, P. (Ed.) General geochemical properties, and abundances of the rare earth elements. In Rare Earth Element Geochemistry; Elsevier: Amsterdam, The Netherlands, 2011; pp. 1–29. [Google Scholar]
- Goodenough, K.M.; Schilling, J.; Jonsson, E.; Kalvig, P.; Charles, N.; Tuduri, J.; Deady, E.A.; Sadeghi, M.; Schiellerup, H.; Müller, A.; et al. Europe’s rare earth element resource potential: An overview of REE metallogenetic provinces and their geodynamic setting. Ore Geol. Rev. 2016, 72, 838–856. [Google Scholar] [CrossRef]
- Kamenopoulos, S.N.; Shields, D.; Agioutantis, Z. Sustainable development criteria and indicators for the assessment of rare earth element mining projects. In Rare Earths Industry; De Lima, I.B., Filho, W.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 87–109. [Google Scholar]
- Zepf, V. (Ed.) Rare earth elements: What and where they are. In Rare Earth Elements; Springer: Berlin/Heidelberg, Germany, 2013; pp. 11–39. [Google Scholar]
- Rudnick, R.L.; Gao, S. Composition of the continental crust. In Treatise on Geochemistry; Rudnick, R.L., Holland, H.D., Turekian, K.K., Eds.; Elsevier Pergamon: Oxford, UK, 2003; Volume 3, pp. 1–7. [Google Scholar]
- Wikipedia. Available online: https://en.wikipedia.org/wiki/Abundance_of_the_chemical_elements (accessed on 15 November 2021).
- Kanazawa, Y.; Kamitani, M. Rare earth minerals and resources in the world. J. Alloys Compd. 2006, 408, 1339–1343. [Google Scholar] [CrossRef]
- Trapasso, G.; Chiesa, S.; Freitas, R.; Pereira, E. What do we know about the ecotoxicological implications of the rare earth element gadolinium in aquatic ecosystems? Sci. Total Environ. 2021, 781, 146273. [Google Scholar] [CrossRef] [PubMed]
- Castor, S.B.; Hendrik, J.B. Rare earth elements. In Industrial Minerals and Rocks: Commodities, Markets, and Uses; Kogel, J.E., Trivedi, N.C., Barker, J.M., Krukowski, S.T., Eds.; Society for Mining Mineralogy: Englewood, CO, USA, 2006; Volume 7. [Google Scholar]
- Schüler, D.; Buchert, M.; Liu, R.; Dittrich, S.; Merz, C. Study on Rare Earths and Their Recycling. Final Report for the Greens/EFA Group in the European Parliament; The Greens/European Free Alliance, Öko-Institut e.V. Darmstadt: Frelburg, Germany, 2011; p. 140. [Google Scholar]
- Binnemans, K.; Jones, P.T.; Blanpain, B.; Van Gerven, T.; Yang, Y.; Walton, A.; Buchert, M. Recycling of rare earths: A critical review. J. Clean. Prod. 2013, 51, 1–22. [Google Scholar] [CrossRef]
- Cao, S.; Duan, X.; Ma, Y.; Zhao, X.; Qin, Y.; Liu, Y.; Li, S.; Zheng, B.; Wei, F. Health benefit from decreasing exposure to heavy metals and metalloid after strict pollution control measures near a typical river basin area in China. Chemosphere 2017, 184, 866–878. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-Gutiérrez, S.C.; Coulon, F.; Jiang, Y.; Wagland, S. Rare earth elements and critical metal content of extracted landfilled material and potential recovery opportunities. Waste Manag. 2015, 42, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Resende, L.V.; Morais, C.A. Study of the recovery of rare earth elements from computer monitor scraps–leaching experiments. Miner. Eng. 2010, 233, 277–280. [Google Scholar] [CrossRef]
- Ikhlayel, M. An integrated approach to establish e-waste management systems for developing countries. J. Clean. Prod. 2018, 170, 119–130. [Google Scholar] [CrossRef]
- Kim, T.; Kim, H.; Kim, G. Tracing River water versus wastewater sources of trace elements using rare earth elements in the Nakdong River estuarine waters. Mar. Poll. Bull. 2020, 160, 111589. [Google Scholar] [CrossRef] [PubMed]
- Pagano, G.; Aliberti, F.; Guida, M.; Oral, R.; Siciliano, A.; Trifuoggi, M.; Tommasi, F. Rare earth elements in human and animal health: State of art and research priorities. Environ. Res. 2015, 142, 215–220. [Google Scholar] [CrossRef]
- Pagano, G.; Guida, M.; Tommasi, F.; Oral, R. Health effects and toxicity mechanisms of rare earth elements—Knowledge gaps and research prospects. Ecotoxicol. Environ. Saf. 2015, 115, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Aide, M. Lanthanide soil chemistry and its importance in understanding soil pathways: Mobility, plant uptake, and soil health. Lanthanides. In Lanthanides, 23rd ed.; Awwad, N.S., Mubarak, A.T., Eds.; IntechOpen: London, UK, 2019. [Google Scholar]
- Caccia, V.G.; Milero, J.F. Distribution of yttrium and rare earths in Florida Bay sediments. Mar. Chem. 2007, 104, 171–185. [Google Scholar] [CrossRef]
- Sholkovitz, E.R. The aquatic chemistry of rare earth elements in rivers and estuaries. Aquat. Geochem. 1995, 1, 1–34. [Google Scholar] [CrossRef]
- Marmolejo-Rodríguez, A.J.; Prego, R.; Meyer-Willerer, A.; Shumilin, E.; Sapozhnikov, D. Rare earth elements in iron oxy-hydroxide rich sediments from the Marabasco River-Estuary System (Pacific coast of Mexico), REE affinity with iron and aluminium. J. Geochem. Explor. 2007, 94, 43–51. [Google Scholar] [CrossRef]
- Elderfield, H.; Upstill-Goddard, R.; Sholkovitz, E.R. The rare earth elements in rivers, estuaries, and coastal seas and their significance to the composition of ocean waters. Geochim. Cosmochim. Acta 1990, 54, 971–991. [Google Scholar] [CrossRef]
- Moermond, C.T.A.; Tijink, J.; van Wezel, A.P.; Koelmans, A.A. Distribution, speciation, and bioavailability of lanthanides in the Rhine-Meuse estuary, The Netherlands. Environ. Toxicol. Chem. 2001, 20, 1916–1926. [Google Scholar] [CrossRef]
- Goldstein, S.J.; Jacobsen, S.B. The Nd and Sr isotopic systematics of river-water dissolved material: Implications for the sources of Nd and Sr in seawater. Chem. Geol. 1987, 46, 245–272. [Google Scholar] [CrossRef]
- Goldstein, S.J.; Jacobsen, S.B. Rare earth elements in river waters. Earth Planet. Sci. Lett. 1988, 89, 35–47. [Google Scholar] [CrossRef]
- Kulaksiz, S.; Bau, M. Rare earth elements in the Rhine River, Germany: First case of anthropogenic lanthanum as a dissolved microcontaminant in the hydrosphere. Environ. Int. 2011, 37, 973–979. [Google Scholar] [CrossRef]
- Bau, M.; Dulski, P. Anthropogenic origin of positive gadolinium anomalies in river waters. Earth Planet. Sci. Lett. 1996, 143, 245–255. [Google Scholar] [CrossRef]
- Tang, J.; Johannesson, K.H. Speciation of rare earth elements in natural terrestrial waters: Assessing the role of dissolved organic matter from the modeling approach. Geochim. Cosmochim. Acta 2003, 67, 2321–2339. [Google Scholar] [CrossRef]
- Pourret, O.; Tuduri, J. Continental shelves as potential resource of rare earth elements. Sci. Rep. 2017, 7, 5857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johannesson, K.H.; Lyons, W.B.; Bird, D.A. Rare earth element concentrations and speciation in alkaline lakes from the western USA. Geophys. Res. Lett. 1994, 21, 773–776. [Google Scholar] [CrossRef]
- Wilde, P.; Quinby-Hunt, M.S.; Erdtmann, B.D. The whole-rock cerium anomaly: A potential indicator of eustatic sea-level changes in shales of the anoxic facies. Sediment. Geol. 1996, 101, 43–53. [Google Scholar] [CrossRef]
- Ramesh, R.; Ramanathan, A.L.; James, R.A.; Subramaniam, V.; Jacobsen, S.B.; Holland, H.D. Rare earth elements and heavy metal distribution in estuarine sediments of east coast of India. Hydrobiologia 1999, 397, 89–99. [Google Scholar] [CrossRef]
- Borrego, J.; López-González, N.; Carro, B.; Lozano-Soria, O. Origin of the anomalies in light and middle REE in sediments of an estuary affected by phosphogypsum wastes (south-western Spain). Mar. Pollut. Bull. 2004, 49, 1045–1053. [Google Scholar] [CrossRef]
- Prego, R.; Caetano, M.; Bernárdez, P.; Brito, P.; Ospina-Alvarez, N.; Vale, C. Rare Earth Elements in coastal sediments of the Northern Galician Shelf: Influence of geological features. Cont. Shelf Res. 2012, 35, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Olmez, I.; Sholkovltz, E.R.; Hermann, D.; Eganhouse, R.P. Rare earth elements in sediments off Southern California: A new anthropogenic indicator. Environ. Sci. Technol. 1991, 25, 310–316. [Google Scholar] [CrossRef]
- Sholkovitz, E.R. REE’s in marine sediments and geochemical standards. Chem. Geol. 1990, 88, 333–347. [Google Scholar] [CrossRef]
- Zhu, W.; Kennedy, M.; de Leer, E.W.B.; Zhou, H.; Alaerts, G.J.F.R. Distribution and modelling of rare earth elements in Chinese river sediments. Sci. Total Environ. 1997, 204, 233–243. [Google Scholar] [CrossRef]
- Nozaki, Y.; Lerche, D.; Alibo, D.S.; Snidvongs, A. The estuarine geochemistry of rare earth elements and indium in the Chao Phraya River, Thailand. Geochim. Cosmochim. Acta 2000, 64, 3983–3994. [Google Scholar] [CrossRef]
- Singh, P.; Rajamani, V. Geochemistry of the floodplain sediments of the Kaveri River, Southern India. J. Sed. Res. 2001, 71, 50–60. [Google Scholar] [CrossRef]
- Chaillou, G.; Anschutz, P.; Lavaux, G.; Blanc, G. Rare earth elements in the modern sediments of the Bay of Biscay (France). Mar. Chem. 2006, 100, 39–52. [Google Scholar] [CrossRef]
- Lawrence, M.G.; Kamber, B.S. The behavior of the rare earth elements during estuarine mixing revisited. Mar. Chem. 2006, 100, 147–161. [Google Scholar] [CrossRef]
- Censi, P.; Sprovieri, M.; Saiano, F.; Di Geronimo, S.I.; Larocca, D.; Placenti, F. The behaviour of REEs in Thailand’s Mae Klong estuary: Suggestions from the Y/Ho ratios and lanthanide tetrad effects. Estuar. Coast. Shelf Sci. 2007, 71, 569–579. [Google Scholar] [CrossRef]
- Hannigan, R.; Dorval, E.; Jones, C. The rare earth element chemistry of estuarine surface sediments in the Chesapeake Bay. Chem. Geol. 2010, 272, 20–30. [Google Scholar] [CrossRef]
- Deepulal, P.M.; Gireesh Kumar, T.R.; Sujatha, C.H. Behaviour of REEs in a tropical estuary and adjacent continental shelf of southwest coast of India: Evidence from anomalies. J. Earth Syst. Sci. 2012, 121, 1215–1227. [Google Scholar] [CrossRef] [Green Version]
- López-González, N.; Borrego, G.; Carro, B.; Grande, J.A. Rare-earth-element fractionation patterns in estuarine sediments as a consequence of acid mine drainage: A case study in SW Spain. Bol. Geol. Min. 2012, 123, 55–64. [Google Scholar]
- Suja, S.; Fernandes, L.L.; Rao, V.P. Distribution and fractionation of rare earth elements and Yttrium in suspended and bottom sediments of the Kali estuary, western India. Environ. Earth Sci. 2017, 76, 174. [Google Scholar] [CrossRef]
- Laukert, G.; Frank, M.; Bauch, D.; Hathorne, E.C.; Rabe, B.; von Appen, W.J.; Wegner, C.; Zieringer, M.; Kassens, H. Ocean circulation and freshwater pathways in the Arctic Mediterranean based on a combined Nd isotope, REE and oxygen isotope section across Fram Strait. Geochem. Cosmochim. Acta 2017, 202, 285–309. [Google Scholar] [CrossRef] [Green Version]
- Osborne, A.H.; Haley, B.; Hathorne, E.C.; Flögel, S.; Frank, M. Neodymium isotopes and concentrations in Caribbean seawater: Tracing water mass mixing and continental input in a semi-enclosed ocean basin. Earth Planet. Sci. Lett. 2014, 406, 174–186. [Google Scholar] [CrossRef]
- Bayon, G.; German, C.R.; Burton, K.W.; Nesbitt, R.W.; Rogers, N. Sedimentary Fe–Mn oxy-hydroxides as paleoceanographic archives and the role of aeolian flux in regulating oceanic dissolved REE. Earth Planet. Sci. Lett. 2004, 224, 477–492. [Google Scholar] [CrossRef]
- Canet, C.; Prol-Ledesma, R.M.; Proensa, J.A.; Rubio-Ramos, M.A.; Forrest, M.J.; Torres-Vera, M.A.; Rodríguez-Díaz, A.A. Mn-Ba–Hg mineralization at shallow submarine hydrothermal vents in Bahía, Baja California Sur, México. Chem. Geol. 2005, 224, 96–112. [Google Scholar] [CrossRef]
- Sifeta, K.; Roser, B.P.; Kimura, J.I. Geochemistry, provenance, and tectonic setting of Neoproterozoic metavolcanic and metasedimentary units, Werri area, Northern Ethiopia. J. Afr. Earth Sci. 2005, 41, 212–234. [Google Scholar] [CrossRef]
- Brookins, D. Aqueous geochemistry of rare earth elements. Rev. Miner. Geochem. 1989, 21, 201–225. [Google Scholar]
- Saha, N.; Webb, G.E.; Zhao, J.X.; Lewis, S.E.; Nguyen, A.D.; Feng, Y. Spatiotemporal variation of rare earth elements from river to reef continuum aids monitoring of terrigenous sources in the Great Barrier Reef. Geoch. Cosm. Acta 2021, 299, 85–112. [Google Scholar] [CrossRef]
- Wilke, C.; Barkleit, A.; Stumpf, T.; Ikeda-Ohno, A. Speciation of the trivalent elements Eu(III) and Cm(III) in digestive media. J. Inorg. Biochem. 2017, 175, 248–258. [Google Scholar] [CrossRef]
- Sklyarova, O.A.; Sklyarov, E.V.; Och, L.; Pastukhov, M.V.; Zagorulko, N.A. Rare earth elements in tributaries of lake Baikal (Siberia, Russia). Appl. Geochem. 2017, 82, 164–176. [Google Scholar] [CrossRef] [Green Version]
- Sholkovitz, E. Flocculation of dissolved organic and inorganic matter during the mixing of river water and seawater. Geochim. Cosmochim. Acta 1976, 40, 831–845. [Google Scholar] [CrossRef]
- Delgado, J.; Pérez-López, R.; Galván, L.; Nieto, J.M.; Boski, T. Enrichment of rare earth elements as environmental tracers of contamination by acid mine drainage in salt marshes: A new perspective. Mar. Pollut. Bull. 2012, 64, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Schijf, J.; Christenson, E.A.; Byrne, R.H. YREE scavenging in seawater: A new look at an old model. Mar. Chem. 2015, 177, 460–471. [Google Scholar] [CrossRef] [Green Version]
- Borrego, J.; López-González, N.; Carro, B.; Lozano-Soria, O. Geochemistry of rare-earth elements in Holocene sediments of an acidic estuary: Environmental markers (Tinto River estuary, South-western Spain). J. Geochem. Explor. 2005, 86, 119–129. [Google Scholar] [CrossRef]
- Byrne, R.H.; Kim, K.H. Rare-Earth element scavenging in seawater. Geochim. Cosmochim. Acta 1990, 54, 2645–2656. [Google Scholar] [CrossRef]
- Sholkovitz, E.R.; Landing, W.M.; Lewis, B.L. Ocean particle chemistry: The fractionation of rare earth elements between suspended particles and seawater. Geochim. Cosmochim. Acta 1994, 58, 1567–1579. [Google Scholar] [CrossRef]
- Bau, M. Controls on the fractionation of isovalent trace elements in magmatic and aqueous systems: Evidence from Y/Ho, Zr/Hf, and lanthanide tetrad effect. Contrib. Mineral. Petrol. 1996, 123, 323–333. [Google Scholar] [CrossRef]
- Sholkovitz, E.R. Artifacts associated with the chemical leaching of sediments for rare-earth elements. Chem. Geol. 1989, 77, 47–51. [Google Scholar] [CrossRef]
- Andrade, R.L.B.; Hatje, V.; Pedreira, R.M.A.; Böning, P.; Pahnke, K. REE fractionation and human Gd footprint along the continuum between Paraguaçu River to coastal South Atlantic waters. Chem. Geol. 2020, 532, 119303. [Google Scholar] [CrossRef]
- Merschel, G.; Bau, M.; Dantas, E.L. Contrasting impact of organic and inorganic nanoparticles and colloids on the behavior of particle-reactive elements in tropical estuaries: An experimental study. Geochim. Cosmochim. Acta 2017, 197, 1–13. [Google Scholar] [CrossRef]
- Chakraborty, P.; Raghunadh Babu, P.V.; Sarma, V.V. A multi-method approach for the study of lanthanum speciation in coastal and estuarine sediments. J. Geochem. Explor. 2011, 110, 225–231. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, L.; Zhang, S.; Li, X. Geochemistry of rare earth elements in the mainstream of the Yangtze River, China. Appl. Geochem. 1998, 13, 451–462. [Google Scholar] [CrossRef]
- Rezaee, K.; Saion, E.B.; Wood, A.K.; Abdi, M.R. Rare earth elements determination and distribution patterns in surface marine sediments of the South China Sea by INAA. Malays. J. Radioanal. Nucl. Chem. 2010, 283, 823–829. [Google Scholar] [CrossRef]
- Shynu, R.; Purnachandra Rao, V.; Kessarkar, P.M.; Rao, T.G. Rare earth elements in suspended and bottom sediments of the Mandovi estuary, central west coast of India: Influence of mining Estuarine. Coast. Shelf Sci. 2011, 94, 355–368. [Google Scholar] [CrossRef]
- Hu, G.; Lin, C.; Yang, Q.; Yan, Y.; Huang, H.; Yu, R.; Cui, J.; Yan, Y. Distribution and ratioointment of rare earth elements in offshore sediments of Western Xiamen Bay, Southeast China. J. Geochem. Expl. 2019, 201, 31–39. [Google Scholar] [CrossRef]
- Rivas-Sanchez, M.L.; Alva-Valdivia, L.M.; Arenas-Alatorre, J.; Urrutia-Fucugauchi, J.; Ruiz-Sandoval, M.; Ramos-Molina, M.A. Berthierine and chamosite hydrothermal: Genetic guides in the Peña Colorada magnetite-bearing ore deposit, Mexico. Earth Planets Space 2006, 58, 1389–1400. [Google Scholar] [CrossRef] [Green Version]
- Weltje, L.; Heidenreich, H.; Zhu, W.; Wolterbeek, H.T.; Korhammer, S.; Goeij, J.J.M.; Markert, B. Lanthanide concentrations in freshwater plants and molluscs, related to those in surface water, pore water and sediment. A case study in The Netherlands. Sci. Total Environ. 2002, 286, 191–214. [Google Scholar] [CrossRef]
- Zhang, S.; Shan, X.Q. Speciation of rare earth elements in soil and accumulation by wheat with rare earth fertilizer application Environ. Pollut. 2001, 112, 395–405. [Google Scholar] [CrossRef]
- Strady, E.; Kim, I.; Radakovitch, O.; Kim, G. Rare earth element distributions and fractionation in plankton from the northwestern Mediterranean Sea. Chemosphere 2015, 119, 72–82. [Google Scholar] [CrossRef]
- Figueiredo, C.; Grilo, T.F.; Lopes, C.; Brito, P.; Diniz, M.; Caetano, M.; Rosa, R.; Raimundo, J. Accumulation, elimination and neuro-oxidative damage under lanthanum exposure in glass eels (Anguilla anguilla). Chemosphere 2018, 206, 414–423. [Google Scholar] [CrossRef]
- Censi, P.; Randazzo, L.A.; D’Angelo, S.; Saiano, F.; Zuddas, P.; Mazzola, S.; Cuttitta, A. Relationship between lanthanide contents in aquatic turtles and environmental exposures. Chemosphere 2013, 91, 1130–1135. [Google Scholar] [CrossRef]
- Mayfield, D.B.; Fairbrother, A. Examination of rare earth element concentration patterns in freshwater fish tissues. Chemosphere 2015, 120, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wang, X.; Nie, H.; Shao, L.; Wang, G.; Liu, Y. Residual levels of rare earth elements in freshwater and marine fish and their health risk assessment from Shandong, China. Mar. Pollut. Bull. 2016, 107, 393–397. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.M.; Behkami, S.; Yusoff, I.; Md Zain, S.B.; Bakar, N.K.A.; Bakar, A.F.A.; Alias, Y. Geochemical characteristics of rare earth elements in different types of soil: A chemometric approach. Chemosphere 2017, 184, 673–678. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, P.; Miramand, P. Subcellular and body distributions of 17 trace elements in the variegated scallop Chlamys varia from the French coast of the Bay of Biscay. Sci. Total Environ. 2005, 337, 59–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, A.S.; Snape, I.; Stark, J.S.; Johnstone, G.J.; Townsend, A.T. Baseline metal concentrations in Paramoera walkeri from East Antarctica. Mar. Pollut. Bull. 2006, 52, 1441–1449. [Google Scholar] [CrossRef]
- Riget, F.; Johansen, P.; Asmund, G. Influence of length on element concentrations in blue mussels (Mytilus edulis). Mar. Pollut. Bull. 1996, 32, 745–751. [Google Scholar] [CrossRef]
- Nørregaard, R.D.; Kaarsholm, H.; Bach, L.; Nowak, B.; Geertz-Hansen, O.; Søndergaard, J.; Sonne, C. Bioaccumulation of rare earth elements in juvenile arctic char (Salvelinus alpinus) under field experimental conditions. Sci. Total Environ. 2019, 688, 529–535. [Google Scholar] [CrossRef]
- Ma, L.; Dang, D.H.; Wang, W.; Evans, R.D.; Wang, W.X. Rare earth elements in the Pearl River Delta of China: Potential impacts of the REE industry on water, suspended particles, and oysters. Environ. Pollut. 2019, 244, 190–201. [Google Scholar] [CrossRef]
- Jiao, Y.; Yang, L.; Kong, Z.; Shao, L.; Wang, G.; Ren, X.; Liu, Y. Evaluation of trace metals and rare earth elements in mantis shrimp Oratosquilla oratoria collected from Shandong Province, China, and its potential risks to human health. Mar. Poll. Bull. 2021, 162, 111815. [Google Scholar] [CrossRef]
- Menegario, A.A.; Yabuki, L.N.M.; Luko, K.S.; Williams, P.N.; Blackburn, D.M. Use of diffusive gradient in thin films for in situ measurements: A review on the progress in chemical fractionation, speciation, and bioavailability of metals in waters. Anal. Chim. Acta 2017, 983, 54–66. [Google Scholar] [CrossRef] [Green Version]
- Cánovas, C.R.; Basallote, M.D.; Macías, F. Distribution and availability of rare earth elements and trace elements in the estuarine waters of the Ría of Huelva (SW Spain). Environ. Poll. 2020, 267, 115506. [Google Scholar] [CrossRef] [PubMed]
- Linde, A.R.; Arribas, P.; Sanchez-Galan, S.; Garcia-Vazquez, F. Eel (Anguilla anguilla) and brown trout (Salmo trutta) target species to assess the biological impact of trace metal pollution in freshwater ecosystems. Arch. Environ. Contam. Toxicol. 1996, 31, 297–302. [Google Scholar] [CrossRef]
- Durrieu, G.; Maury-Brachet, R.; Girardin, M.; Rochard, E.; Boudou, A. Contamination by heavy metals (Cd, Zn, Cu, and Hg) of eight fish species in the Gironde estuary (France). Estuaries 2005, 28, 581–591. [Google Scholar] [CrossRef]
- Lortholarie, M.; Poirier, L.; Kamari, A.; Herrenknecht, C.; Zalouk-Vergnoux, A. Rare earth element organotropism in European eel (Anguilla anguilla). Sci. Total Environ. 2021, 766, 142513. [Google Scholar] [CrossRef]
- Pernice, M.; Boucher, J.; Boucher-Rodoni, R.; Joannot, P.; Bustamante, P. Comparative bioaccumulation of trace elements between Nautilus pompilius and Nautilus macromphalus (Cephalopoda: Nautiloidea) from Vanuatu and NewCaledonia. Ecotoxicol. Environ. Saf. 2009, 72, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Ichihashi, H.; Kohno, H.; Kannan, K.; Tsumura, A.; Yamasaki, S.I. Multi-elemental analysis of purple back flying squid using high resolution inductively coupled plasma-mass spectrometry (HR ICP-MS). Environ. Sci. Technol. 2001, 35, 3103–3108. [Google Scholar] [CrossRef] [PubMed]
- Chassard-Bouchard, C.; Hallegot, P. Bioaccumulation de lanthane par desmoulesMytilus edulis(L) récoltées sur les côtes françaises. Microanalyse parspectrographie des rayons X et par émission ionique secondaire. Acad. Sci. III 1984, 298, 567–572. [Google Scholar]
- Lavezzo, B.; Kinoshita, A.; Figueiredo, A.M.G.; Faita Pinheiro, M.M.; Santana, W. Detection of rare-earth elements using fiddler crabs Leptuca leptodactyla (Crustacea: Ocypodidae) as bioindicators in mangroves on the coast of São Paulo, Brazil. Sci. Total Environ. 2020, 738, 139787. [Google Scholar] [CrossRef] [PubMed]
- Natálio, L.F.; Pardo, J.C.; Machado, G.B.; Fortuna, M.D.; Gallo, D.G.; Costa, T.M. Potential effect of fiddler crabs on organic matter distribution: A combined laboratory and field experimental approach. Estuar. Coast. Shelf Sci. 2017, 184, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Ng, J.S.; Lui, K.K.; Lai, C.H.; Leung, K.M. Harpiosquilla harpax (Crustacea, Stomatopoda) as a biomonitor of trace metal contamination in benthic sediments in Hong Kong waters. Mar. Pollut. Bull. 2007, 54, 1523–1529. [Google Scholar] [CrossRef]
- Bonnail, E.; Perez-López, R.; Sarmiento, A.M.; Nieto, J.M.; Del Valls, T.A. A novel approach for acid mine drainage pollution biomonitoring using rare earth elements bioaccumulated in the freshwater clam Corbicula fluminea. J. Hazard. Mater. 2017, 338, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Dang, D.H.; Schaefer, J.; Brach-Papa, C.; Lenoble, V.; Durrieu, G.; Dutruch, L.; Chiffoleau, J.F.; Gonzalez, J.L.; Blanc, G.; Mullot, J.U.; et al. Evidencing the impact of coastal contaminated sediments on mussels. Environ. Sci. Technol. 2015, 49, 11438–11448. [Google Scholar] [CrossRef] [PubMed]
- Bridges, C.C.; Zalups, R.K. Molecular and ionic mimicry and the transport of toxic metals. Toxicol. Appl. Pharmacol. 2005, 204, 274–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, C.H. Interesting and useful biochemical properties of lanthanides. Trends Biochem. Sci. 1983, 8, 445–449. [Google Scholar] [CrossRef]
- Wang, S.C. PCNA: A silent housekeeper or a potential therapeutic target? Trends Pharmacol. Sci. 2014, 35, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, Z.; Jiang, W.; Yu, M.; Zhou, Y.; Zhao, Y.; Chai, Z. Direct measurement of lanthanum uptake and distribution in internodal cells of Chara. Plant Sci. 2008, 174, 496–501. [Google Scholar] [CrossRef]
- Palasz, A.; Czekaj, P. Toxicological and cytophysiological aspects of lanthanides action. Acta Biochim. Pol. 2000, 47, 1107–1114. [Google Scholar] [CrossRef] [Green Version]
- Barry, M.J.; Meehan, B.J. The acute and chronic toxicity of lanthanum to Daphnia carinata. Chemosphere 2000, 41, 1669–1674. [Google Scholar] [CrossRef]
- Borgmann, U.; Couillard, Y.; Doyle, P.; Dixon, D.G. Toxicity of sixty-three metals and metalloids to Hyalella Azteca at two levels of water hardness. Environ. Toxicol. Chem. 2005, 24, 641–652. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Directive of the European Parliament and of the Council on Environmental Quality Standards in the Field of Water Policy and Amending Directive 2000/60/EC; COM 2006 398 final; European Commission: Brussels, Belgium, 2000; p. 25. [Google Scholar]
- Tang, J.; Johannesson, K.H. Ligand extraction of rare earth elements from aquifer sediments: Implications for rare earth element complexation with organic matter in natural waters. Geochim. Cosmochim. Acta 2010, 74, 6690–6705. [Google Scholar] [CrossRef]
- Zhao, C.M.; Wilkinson, K.J. Biotic ligand model does not predict the bioavailability of rare earth elements in the presence of organic ligands. Environ. Sci. Technol. 2015, 49, 2207–2214. [Google Scholar] [CrossRef] [PubMed]
- Amyot, M.; Clayden, M.G.; Macmillan, G.A.; Perron, T.; Arscott-Gauvin, A. Fate and trophic transfer of rare earth elements in temperate lake food webs. Environ. Sci. Technol. 2017, 51, 6009–6017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reindl, A.R.; Falkowska, L. Trace elements in the muscle, ova and seminal fluid of key clupeid representatives from the Gdansk Bay (South Baltic Sea) and Iberian Peninsula (North-East Atlantic). J. Trace Elem. Med. Biol. 2021, 68, 126803. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Saion, E.; Halimah, M.K.; Yap, C.K.; Hamzah, M.S. Rare earth element (REE) in surface mangrove sediment by instrumental neutron activation analysis. J. Radioanal. Nucl. Chem. 2014, 301, 667–676. [Google Scholar] [CrossRef]
- Li, J.-X.; Zheng, L.; Sun, C.-J.; Jiang, F.-H.; Yin, X.-F.; Chen, J.-H.; Han, B.; Wang, X.-R. Study on ecological and chemical properties of rare earth elements in tropical marine organisms. Chin. J. Anal. Chem. 2016, 44, 1539–1546. [Google Scholar] [CrossRef]
- Carpenter, D.; Boutin, C.; Allison, J.E.; Parsons, J.L.; Ellis, D.M. Uptake and effects of six rare earth elements (REEs) on selected native and crop species growing in contaminated soils. PLoS ONE 2015, 10, e0129936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyler, G. Rare earth elements in soil and plant systems—A review. Plant Soil 2004, 267, 191–206. [Google Scholar] [CrossRef]
- Liu, D.; Wang, Z. Influence of rare earth elements on chemical transformation of nitrogen in agricultural soil. J. Appl. Ecol. 2001, 124, 545–548. [Google Scholar]
- Herrmann, H.; Nolde, J.; Berger, S.; Heise, S. Aquatic ecotoxicity of lanthanum—A review and an attempt to derive water and sediment quality criteria. Ecotoxicol. Environ. Saf. 2016, 124, 213–238. [Google Scholar] [CrossRef] [Green Version]
- González, V.; Vignati, D.A.L.; Leyval, C.; Giamberini, L. Environmental fate and ecotoxicity of lanthanides: Are they a uniform group beyond chemistry? Environ. Int. 2014, 71, 148–157. [Google Scholar] [CrossRef]
- Trifuoggi, M.; Pagano, G.; Guida, M.; Palumbo, A.; Siciliano, A.; Gravina, M.; Lyons, D.M.; Burić, P.; Levak, M.; Thomas, P.J.; et al. Comparative toxicity of seven rare earth elements in sea urchin early life stages. Environ. Sci. Pollut. Res. 2017, 24, 20803–20810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martino, C.; Costa, C.; Roccheri, M.C.; Koop, D.; Scudiero, R.; Byrne, M. Gadolinium perturbs expression of skeletogenic genes, calcium uptake and larval development in phylogenetically distant sea urchin species. Aquat. Toxicol. 2018, 194, 57–66. [Google Scholar]
- Blaise, C.; Gagné, F.; Harwood, M.; Quinn, B.; Hanana, H. Ecotoxicity responses of the freshwater cnidarian Hydra attenuata to 11 rare earth elements. Ecotoxicol. Environ. Saf. 2018, 163, 486–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freitas, R.; Cardoso, C.; Costa, S.; Morais, T.; Moleiro, P.; Lima, A.F.D.; Soares, M.; Figueiredo, S.; Águeda, T.L.; Rocha, P.; et al. New insights on the impacts of e-waste towards marine bivalves: The case of the rare earth element dysprosium. Environ. Pollut. 2020, 260, 113859. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.; Costa, S.; Cardoso, C.E.; Morais, T.; Moleiro, P.; Matias, A.C.; Pereira, A.F.; Machado, J.; Correia, B.; Pinheiro, D.; et al. Toxicological effects of the rare earth element neodymium in Mytilus galloprovincialis. Chemosphere 2020, 244, 125457. [Google Scholar] [CrossRef]
- Tseng, M.T.; Lu, X.; Duan, X.; Hardas, S.S.; Sultana, R.; Wu, P.; Unrine, J.M.; Graham, U.; Butterfield, D.A.; Grulke, E.A.; et al. Alteration of hepatic structure and oxidative stress induced by intravenous nanoceria. Toxicol. Appl. Pharmacol. 2012, 260, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Luo, X.; Tian, Y.; Xie, Y.; Wang, S.; Li, Y.; Tian, L.; Wang, X. Biphasic effects of lanthanum on Vicia faba L. seedlings under cadmium stress, implicating finite antioxidation and potential ecological risk. Chemosphere 2012, 86, 530–537. [Google Scholar] [CrossRef]
- Zhao, H.; Hong, J.; Yu, X.; Zhao, X.; Sheng, L.; Ze, Y.; Sang, X.; Gui, S.; Sun, Q.; Wang, L.; et al. Oxidative stress in the kidney injury of mice following exposure to lanthanides trichloride. Chemosphere 2013, 93, 875–884. [Google Scholar] [CrossRef]
- Hongyan, G.; Liang, C.; Xiaorong, W.; Ying, C. Physiological responses of Carassius auratus to ytterbium exposure. Ecotoxicol. Environ. Saf. 2002, 53, 312–316. [Google Scholar] [CrossRef]
- Oral, R.; Bustamante, P.; Warnau, M.; D’Ambra, A.; Guida, M.; Pagano, G. Cytogenetic and developmental toxicity of cerium and lanthanum to sea urchin embryos. Chemosphere 2010, 81, 194–198. [Google Scholar] [CrossRef] [Green Version]
- González, V.; Vignati, D.A.L.; Pons, M.N.; Montarges-Pelletier, E.; Bojic, C.; Giamberini, L. Lanthanide ecotoxicity: First attempt to measure environmental risk for aquatic organisms. Environ. Pollut. 2015, 199, 139–147. [Google Scholar] [PubMed]
- Hua, D.; Wang, J.; Yu, D.; Liu, J. Lanthanum exerts acute toxicity and histopathological changes in gill and liver tissue of rare minnow (Gobiocypris rarus). Ecotoxicology 2017, 26, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Mestre, N.C.; Serrão Sousam, V.; Lopes Rocha, T.; Bebianno, M.J. Ecotoxicity of rare earths in the marine mussel Mytilus galloprovincialis and a preliminary approach to assess environmental risk. Ecotoxicology 2019, 28, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Campbell, P.G.C. Interactions between trace metals and aquatic organisms: A critique of the free-ion activity model. In Metal Speciation and Bioavailability in Aquatic Systems; Tessier, A., Turner, D.R., Eds.; John Wiley & Sons: London, UK, 1995; pp. 45–102. [Google Scholar]
- Huang, Q.; Bu, Q.; Zhong, W.; Shi, K.; Cao, Z.; Yu, G. Derivation of aquatic predicted no-effect concentration (PNEC) for ibuprofen and sulfamethoxazole based on various toxicity endpoints and the associated risks. Chemosphere 2018, 193, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Aharchaou, I.; Beaubien, C.; Campbell, P.G.C.; Fortin, F. Lanthanum and cerium toxicity to the freshwater green alga chlorella fusca: Applicability of the biotic ligand model. Environ. Toxicol. Chem. 2020, 39, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- El-Akl, P.; Smith, S.; Wilkinson, K.J. Linking the chemical speciation of cerium to its bioavailability in water for a freshwater alga. Environ. Toxicol. Chem. 2015, 34, 1711–1719. [Google Scholar] [CrossRef]
- Lürling, M.; Tolman, Y. Effects of lanthanum and lanthanum-modified clay on growth, survival, and reproduction of Daphnia magna. Water Res. 2010, 44, 309–319. [Google Scholar] [CrossRef]
- Joonas, E.; Aruoja, V.; Olli, K.; Syvertsen-Wiig, G.; Vija, H.; Kahru, A. Potency of (doped) rare earth oxide particles and their constituent metals to inhibit algal growth and induce direct toxic effects. Sci. Total Environ. 2017, 593, 478–486. [Google Scholar] [CrossRef]
- Mebane, C.A.; Chowdhury, M.J.; De Schamphelaere, K.A.C.; Lofts, S.; Paquin, P.R.; Santore, R.C.; Wood, C.M. Metal bioavailability models: Current status, lessons learned, considerations for regulatory use, and the path forward. Environ. Toxicol. Chem. 2020, 39, 60–84. [Google Scholar] [CrossRef] [Green Version]
- Blinova, I.; Lukjanova, A.; Muna, M.; Vija, H.; Kahru, A. Evaluation of the potential hazard of lanthanides to freshwater microcrustaceans. Sci. Total Environ. 2018, 642, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Weltje, L.; Verhoof, L.R.C.W.; Verweij, W.; Hamers, T. Lutetium speciation and toxicity in a microbial bioassay: Testing the free-ion model for lanthanides. Environ. Sci. Technol. 2004, 38, 6597–6604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Q.G.; Yang, G.; Wilkinson, K.J. Biotic ligand model explains the effects of competition but not complexation for Sm biouptake by Chlamydomonas reinhardtii. Chemosphere 2017, 168, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Walton, A.; Sheridan, R.; Güth, K.; Gauß, R.; Gutfleisch, O.; Buchert, M.; Steenari, B.M.; Van Gerven, T.; Jones, P.T.; et al. REE recovery from end-of-life NdFeB permanent magnet scrap: A critical review. J. Sustain. Metall. 2016, 3, 122–149. [Google Scholar] [CrossRef]
- Tai, P.; Zhao, Q.; Su, D.; Li, P.; Stagnitti, F. Biological toxicity of lanthanide elements on algae. Chemosphere 2010, 80, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- Atibu, E.K.; Devarajan, N.; Laffite, A.; Giuliani, G.; Salumu, J.A.; Muteb, R.C.; Mulaji, C.K.; Otamonga, J.P.; Elongo, V.; Mpiana, P.T.; et al. Assessment of trace metaland rare earth elements contamination in rivers around abandoned and activemine areas. The case of Lubumbashi River and Tshamilemba Canal, Katanga, Democratic Republic of the Congo. Geochemistry 2016, 76, 353–362. [Google Scholar] [CrossRef]
- Lompré, J.S.; Moleiro, P.; De Marchi, L.; Soares, A.M.V.M.; Pretti, C.; Chielini, F.; Pereira, E.; Freitas, R. Bioaccumulation and ecotoxicological responses of clams exposed to terbium and carbon nanotubes: Comparison between native (Ruditapes decussatus) and invasive (Ruditapes philippinarum) species. Sci. Total Environ. 2021, 784, 146914. [Google Scholar] [CrossRef]
- Rabiet, M.; Letouzet, M.; Hassanzadeh, S.; Simon, S. Transmetallation of Gd-DTPA by Fe3+, Cu2+ and Zn2+ in water: Batch experiments and coagulation–flocculation simulations. Chemosphere 2014, 95, 639–642. [Google Scholar] [CrossRef]
- Möller, P.; Morteani, G.; Dulski, P. Anomalous gadolinium, cerium, and Yttrium contents in the Adige and Isarco River waters and in the water of their tributaries (Provinces Trento and Bolzano/Bozen, NE Taly). Acta Hydrochim. Hydrobiol. 2003, 31, 225–239. [Google Scholar] [CrossRef]
- Dulski, P.; Möller, P.; Pekdeger, A. Comparison of gadopentetic acid (Gd-DTPA) and bromide in a dual-tracer filed experiment. Hydrogeol. J. 2011, 19, 823–834. [Google Scholar] [CrossRef]
- Telgmann, L.; Wehe, C.A.; Künnemeyer, J.; Bülter, A.-C.; Sperling, M.; Karst, U. Speciation of Gd-based MRI contrast agents and potential products of transmetalation with iron ions or parenteral iron supplements. Anal. Bioanal. Chem. 2012, 404, 2133–2141. [Google Scholar] [CrossRef]
- Nozaki, Y.; Lerche, D.; Alibo, D.S.; Tsutsumi, M. Dissolved indium and rare earth elements in three Japanese rivers and Tokyo Bay: Evidence for anthropogenic Gd and In. Geochem. Cosmochim. Acta 2000, 64, 3975–3982. [Google Scholar] [CrossRef]
- Bau, M.; Knappe, A.; Dulski, P. Anthropogenic gadolinium as a micropollutant in river waters in Pennsylvania and in Lake Erie, northeastern United States. Chem. Erde-Geochem. 2006, 66, 143–152. [Google Scholar] [CrossRef]
- Lawrence, M.G. Detection of anthropogenic gadolinium in the Brisbane River plume in Moreton Bay, Queensland, Australia. Mar. Pollut. Bull. 2010, 60, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Hatje, V.; Bruland, K.W.; Flegal, A.R. Increases in anthropogenic gadolinium anomalies and rare earth element concentrations in San Francisco Bay over a 20 year record. Environ. Sci. Technol. 2016, 50, 4159–4168. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, T.C.; Sonke, J.E.; Chmeleff, J.; Van Beek, P.; Souhaut, M.; Boaventura, G.; Seyler, P.; Jeandel, C. Rapid neodymium release to marine waters from lithogenic sediments in the Amazon estuary. Nat. Commun. 2015, 6, 7592. [Google Scholar] [CrossRef] [PubMed]
- Kulaksız, S.; Bau, M. Contrasting behaviour of anthropogenic gadolinium and natural rare earth elements in estuaries and the gadolinium input into the North Sea. Earth Plan. Sci. Lett. 2007, 260, 361–371. [Google Scholar] [CrossRef]
- Schijf, J.; Christy, I.J. Effect of Mg and Ca on the stability of the MRI contrast agent Gd–DTPA in seawater. Front. Mar. Sci. 2018, 5, 111. [Google Scholar] [CrossRef] [Green Version]
- Henriques, B.; Coppola, F.; Monteiro, R.; Pinto, J.; Viana, T.; Pretti, C.; Soares, A.; Freitas, R.; Pereira, E. Toxicological assessment of anthropogenic Gadolinium in seawater: Biochemical effects in mussels Mytilus galloprovincialis. Sci. Total Environ. 2019, 664, 626–634. [Google Scholar] [CrossRef]
- Fujita, Y.; Walton, M.; Das, G.; Dohnalkova, A.; Vanzin, G.; Anderko, A. Impacts of anthropogenic gadolinium on the activity of the ammonia oxidizing bacterium Nitrosomonas europaea. Chemosphere 2020, 257, 127250. [Google Scholar] [CrossRef]
- Martino, C.; Bonaventura, R.; Byrne, M.; Roccheri, M.; Matranga, V. Effects of exposure to gadolinium on the development of geographically and phylogenetically distant sea urchin’s species. Mar. Environ. Res. 2017, 128, 98–106. [Google Scholar] [CrossRef] [Green Version]
- Martino, C.; Chiarelli, R.; Bosco, L.; Roccheri, M.C. Induction of skeletal abnormalities and autophagy in Paracentrotus lividus sea urchin embryos exposed to gadolinium. Mar. Environ. Res. 2017, 130, 12–20. [Google Scholar] [CrossRef]
- Merschel, G.; Bau, M. Rare earth elements in the aragonitic shell of freshwater mussel Corbicula fluminea and the bioavailability of anthropogenic lanthanum, samarium and gadolinium in river water. Sci. Total Environ. 2015, 533, 91–101. [Google Scholar] [CrossRef]
- Merschel, G.; Bau, M.; Baldewein, L.; Dantas, E.L.; Walde, D.; Bühn, B. Tracing and tracking wastewater-derived substances in freshwater lakes and reservoirs: Anthropogenic gadolinium and geogenic in Lake Paranoá, Brasilia. Compt. Rendus Geosci. 2015, 3475, 284–293. [Google Scholar] [CrossRef]
- Olias, M.; Ceron, J.C.; Fernandez, I.; De La Rosa, J. Distribution of rare earth elements in an alluvial aquifer affected by acid mine drainage: The Guadiamar aquifer (SW Spain). Environ. Pollut. 2005, 135, 53–64. [Google Scholar] [PubMed]
- Åström, M.E.; Österholm, P.; Gustafsson, J.P.; Nystrand, M.; Peltola, P.; Nordmyr, L.; Boman, A. Attenuation of rare earth elements in a boreal estuary. Geochim. Cosmochim. Acta 2012, 96, 105–119. [Google Scholar] [CrossRef]
- Das, T.; Sharma, A.; Talukder, G. Effects of lanthanum in cellular systems.Biolo. Trace Elem. Res. 1988, 18, 201–228. [Google Scholar] [CrossRef] [PubMed]
- Pinto, J.; Costa, M.; Leite, C.; Borges, C.; Coppola, F.; Henriques, B.; Monteiro, R.; Russo, T.; Di Cosmo, A.; Soares, A.M.V.M.; et al. Ecotoxicological effects of lanthanum in Mytilus galloprovincialis: Biochemical and histopathological impacts. Aquat. Toxicol. 2019, 211, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, G.; Mutus, B.; McLennan, I.; Mooibroek, M.J. Activation and in-activation of purified acetylcholinesterase from Electrophorus electricus by lanthanum(III). Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 1982, 703, 142–148. [Google Scholar] [CrossRef]
- Hong, F.; Wang, L.; Liu, C.; Su, M.; Huang, H.; Chen, L. Prevention of La3þonDNA Damage Caused by Hg2þfrom Fish Intestines. J. Rare Earths 2007, 25, 243–248. [Google Scholar] [CrossRef]
- Verbost, P.M.; Flik, G.; Lock, R.A.; Wendelaar Bonga, S.E. Cadmium inhibition of Ca2+ uptake in rainbow trout gills. Am. J. Physiol. 1987, 253, 216–221. [Google Scholar]
- Block, M.; Pärt, P. Uptake of 109Cd by cultured gill epithelial cells from rain-bow trout (Oncorhynchus mykiss). Aquat. Toxicol. 1992, 23, 137–151. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, H.; Deng, S.; Chen, Y.; Yu, Y. Effect of neodymium on growth and physiological characteristics of Microcystis aeruginosa. J. Rare Earths 2011, 29, 388–395. [Google Scholar] [CrossRef]
- Wang, C.; Zhu, W.; Wang, Z.; Guicherit, R. Rare earth elements and other metals in atmospheric particulate matter in the western part of the Netherlands. Water Air Soil Pollut. 2000, 121, 109–118. [Google Scholar] [CrossRef]
- Jenkins, W.; Perone, P.; Walker, K.; Bhagavathula, N.; Aslam, M.N.; Da Silva, M.; Dame, M.K.; Varani, J. Fibroblast response to lanthanoid metalion stimulation: Potential contribution to fibrotic tissue injury. Biol. Trace Elem. Res. 2011, 144, 621–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, L.M.; Norstrom, R.J.; Hobson, K.A.; Muir, D.C.G.; Backus, S.; Fisk, A.T. Mercury and other trace elements in a pelagic Arctic marine food web (North water Polynya, Baffin Bay). Sci. Total Environ. 2005, 351, 247–263. [Google Scholar] [CrossRef]
- Constantinides, S. The Demand for Rare Earth Materials in Permanent Magnets. In Proceedings of the 51st Annual Conference of Metallurgists COM Niagara Falls, Niagara Falls, NY, USA, 30 September–3 October 2012. [Google Scholar]
- Haque, N.; Hughes, A.; Lim, S.; Vernon, C. Rare earth elements: Overview of mining, mineralogy, uses, sustainability, and environmental impact. Resources 2014, 34, 614–635. [Google Scholar] [CrossRef] [Green Version]
- Stegen, K.S. Heavy rare earths, permanent magnets, and renewable energies: An imminent crisis. Energy Policy 2015, 79, 1–8. [Google Scholar] [CrossRef]
- PV Magazine. Available online: https://www.pv-magazine.com/2021/05/07/recycling-rare-earth-elements-in-dead-lithium-batteries/. (accessed on 15 November 2021).
- Chakhmouradian, A.R.; Wall, F. Rare Earth Elements: Minerals, mines, magnets (and more). Elements 2012, 8, 333–340. [Google Scholar] [CrossRef]
- Bonawandt, C. Recycling Rare Earth Metals Presents Challenges, Opportunities. Engineering.com 2013. Available online: https://www.engineering.com/Blogs/tabid/3207/ArticleID/5693/Recycling-Rare-Earth-Metals-Presents-Challenges-Opportunities.aspx?e_src=relart (accessed on 26 October 2021).
- Hono, K.; Sepehri-Amin, H. Prospect for HRE-free high coercivity Nd-Fe-B permanent magnets. Scr. Mater. 2018, 151, 6–13. [Google Scholar] [CrossRef]
- Abrenica, G.H.A.; Ocon, J.D.; Lee, J. Dip-coating synthesis of high-surface area nanostructured FeB for direct usage as anode in metal/metalloid-air battery. Curr. Appl. Phys. 2016, 16, 1075–1080. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arienzo, M.; Ferrara, L.; Trifuoggi, M.; Toscanesi, M. Advances in the Fate of Rare Earth Elements, REE, in Transitional Environments: Coasts and Estuaries. Water 2022, 14, 401. https://doi.org/10.3390/w14030401
Arienzo M, Ferrara L, Trifuoggi M, Toscanesi M. Advances in the Fate of Rare Earth Elements, REE, in Transitional Environments: Coasts and Estuaries. Water. 2022; 14(3):401. https://doi.org/10.3390/w14030401
Chicago/Turabian StyleArienzo, Michele, Luciano Ferrara, Marco Trifuoggi, and Maria Toscanesi. 2022. "Advances in the Fate of Rare Earth Elements, REE, in Transitional Environments: Coasts and Estuaries" Water 14, no. 3: 401. https://doi.org/10.3390/w14030401
APA StyleArienzo, M., Ferrara, L., Trifuoggi, M., & Toscanesi, M. (2022). Advances in the Fate of Rare Earth Elements, REE, in Transitional Environments: Coasts and Estuaries. Water, 14(3), 401. https://doi.org/10.3390/w14030401