
Size–Abundance Relationships of Freshwater Macroinvertebrates in Two Contrasting Floodplain Channels of Rhone River


Abstract
:1. Introduction
2. Materials and Methods
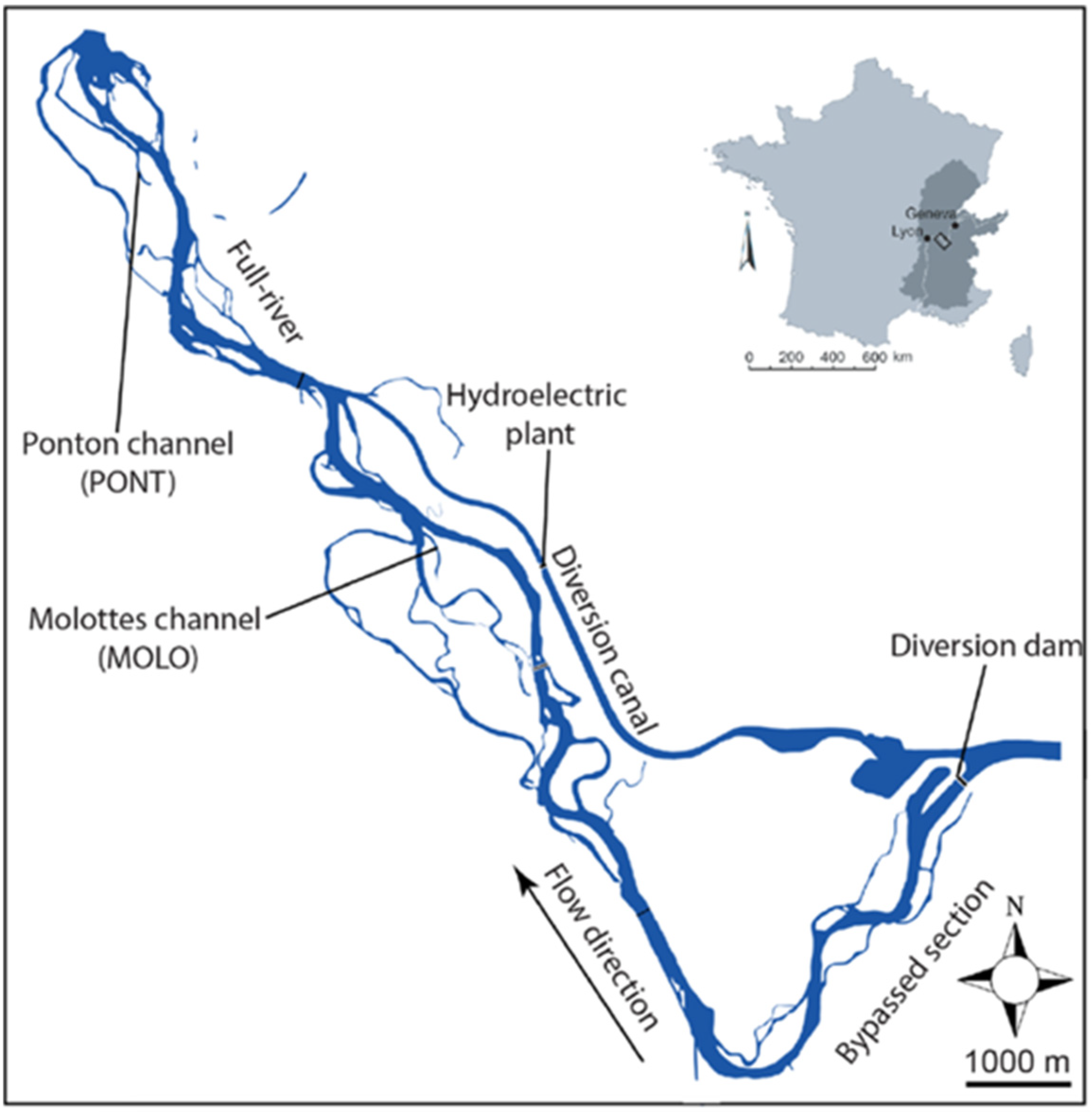
2.1. Studied Sites
2.2. Sampling Method and Ecological Traits
2.3. Statistical Analysis
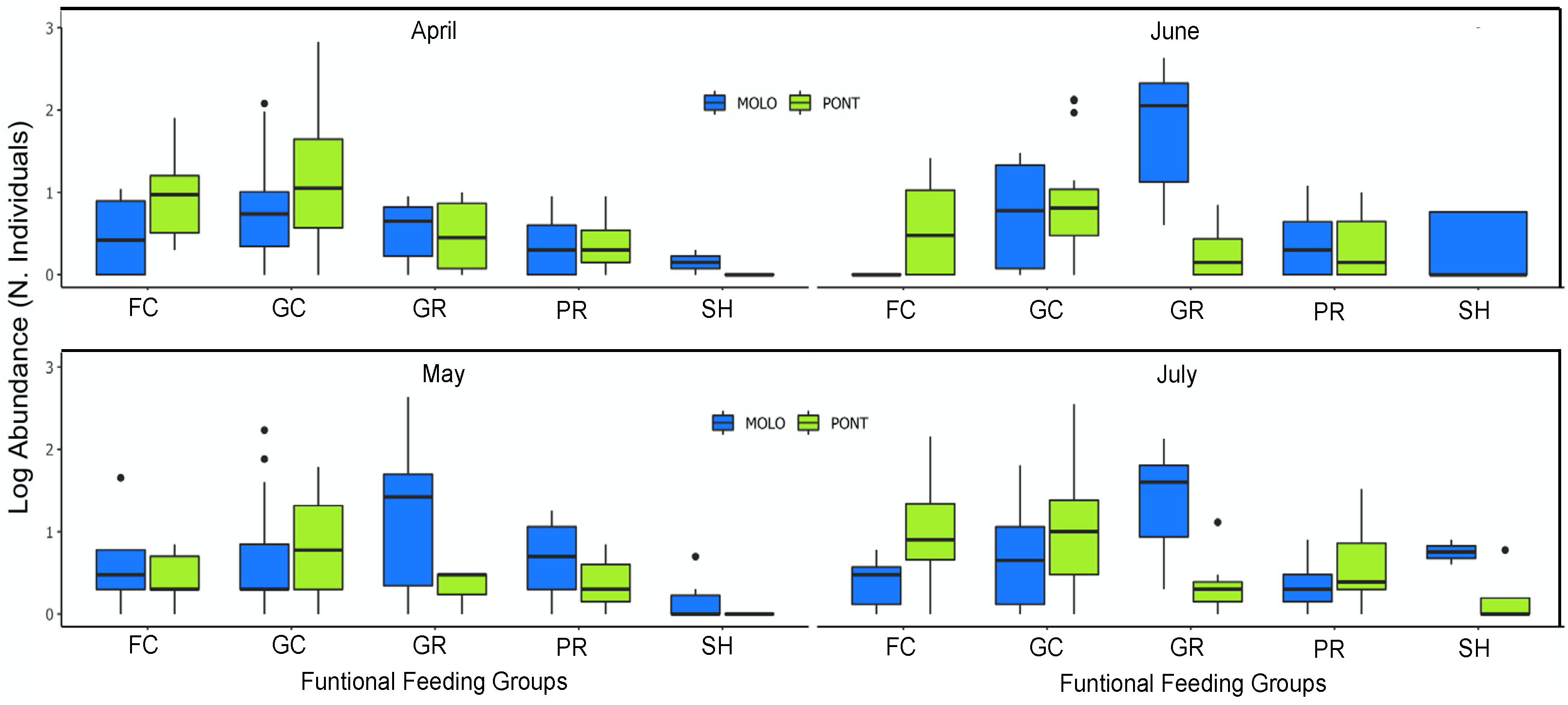
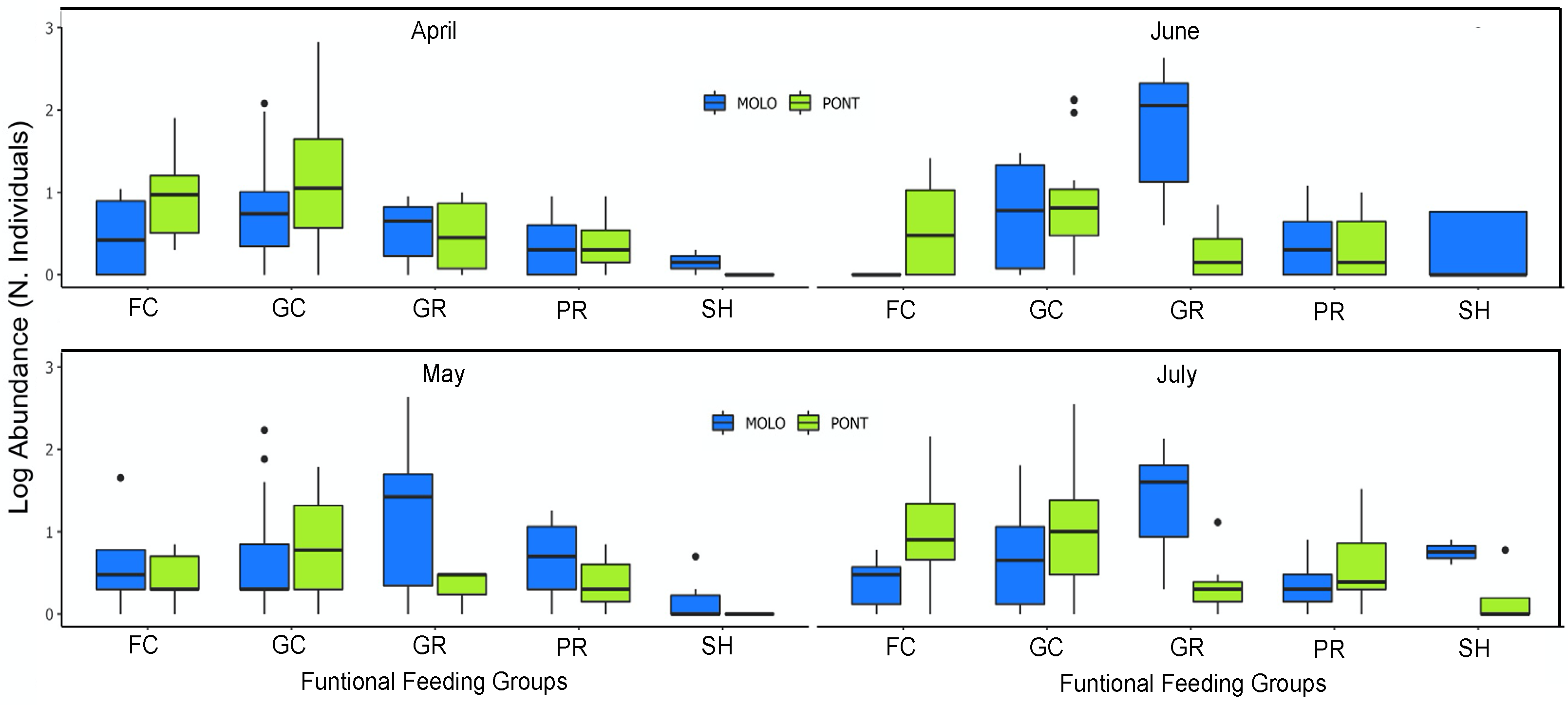
3. Results
4. Discussion
4.1. Patterns of LSDRMOLO and LSDRPONT (All Months Included)
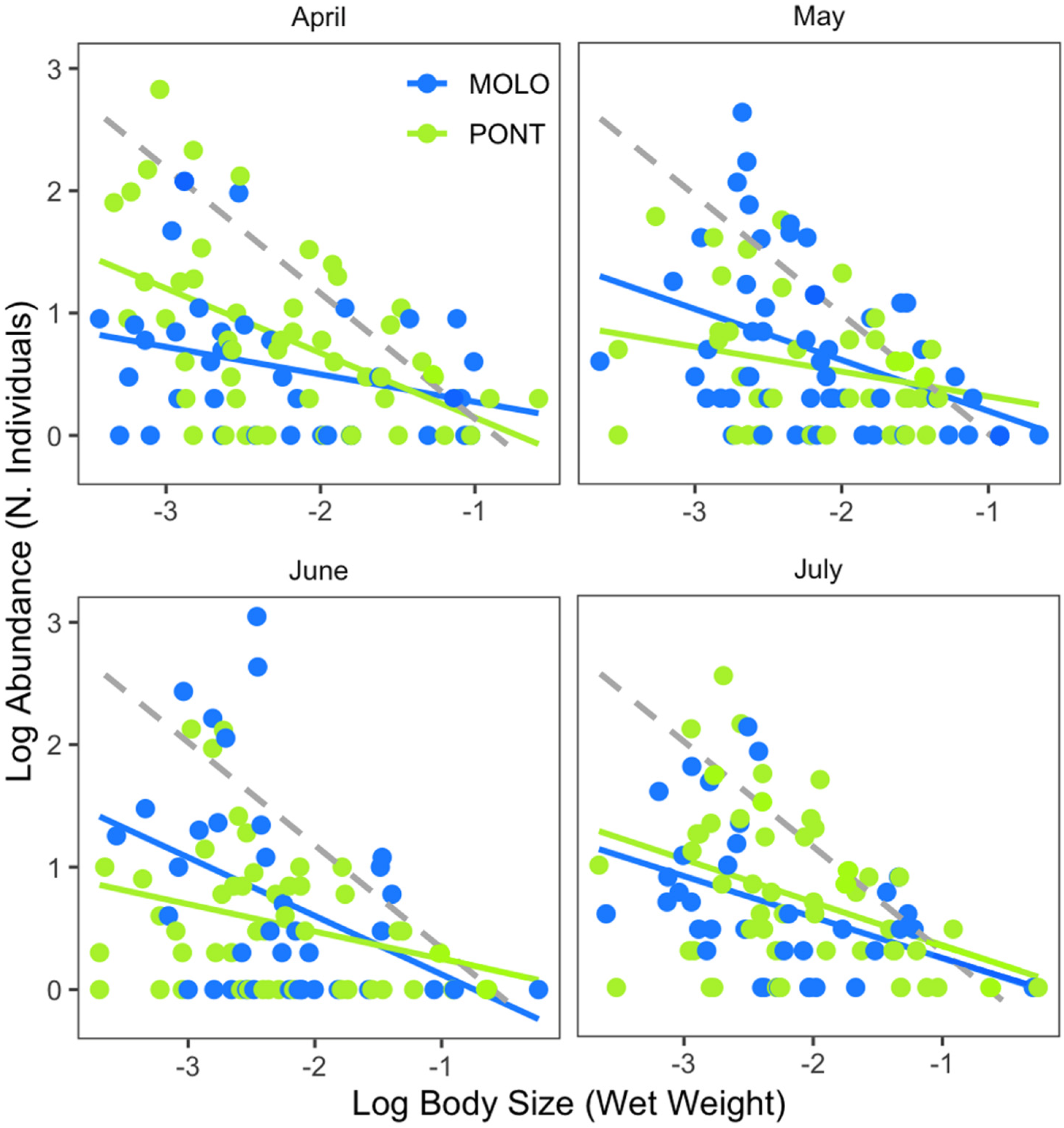
4.2. Patterns of LSDRMOLO and LSDRPONT across Months
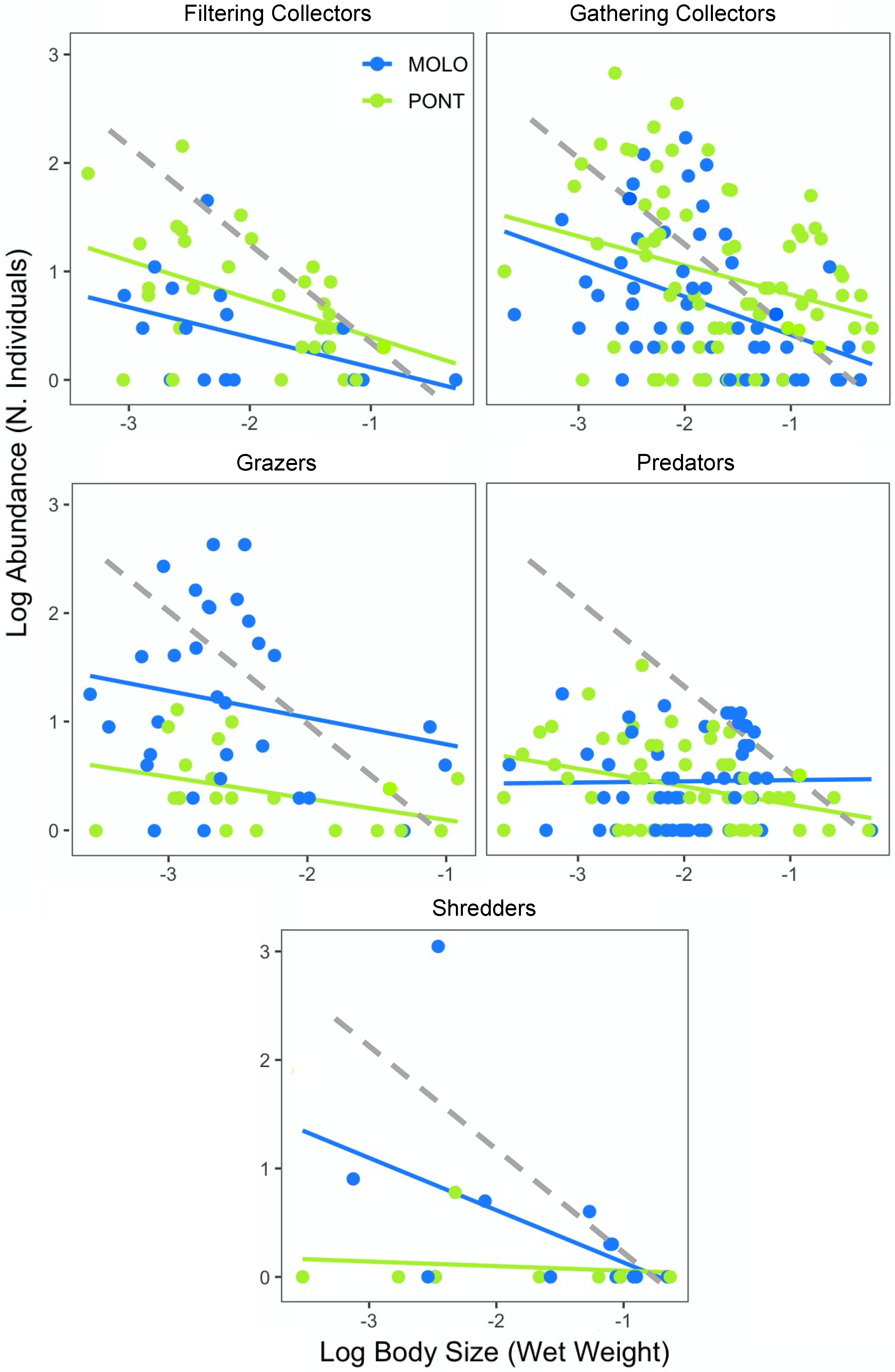
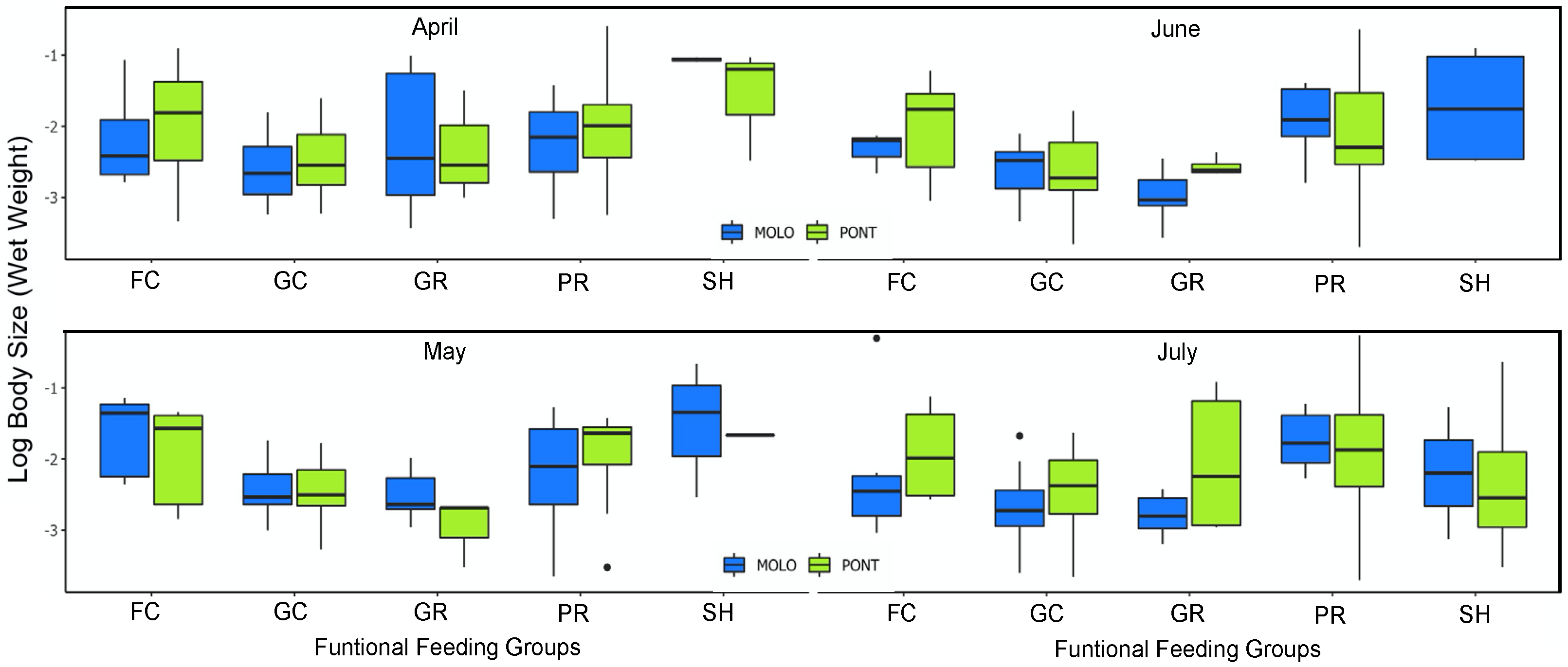
4.3. Patterns of LSDRMOLO and LSDRPONT across Functional Diversity
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Site | p Value for Slope a | p Value for Intercept b | ||
|---|---|---|---|---|
| MOLO | PONT | MOLO | PONT | |
| MOLO | - | ns | - | ns |
| PONT | ns | - | ns | - |
| Month | p Value for Slope a | p Value for Intercept b | ||||||
|---|---|---|---|---|---|---|---|---|
| April | May | June | July | April | May | June | July | |
| April | ** | - | - | - | * | - | - | - |
| May | - | * | - | - | - | - | - | - |
| June | - | - | * | - | - | - | * | - |
| July | - | - | - | ns | - | - | - | ns |
| Feeding Types | p Value for Slope a | p Value for Intercept b | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| FC | GC | GR | PR | SH | FC | GC | GR | PR | SH | |
| Filtering Collectors | ns | - | - | - | - | * | - | - | - | - |
| Gathering Collectors | - | ns | - | - | - | - | * | - | - | - |
| Grazers | - | - | ns | - | - | - | - | * | - | - |
| Predators | - | - | - | ns | - | - | - | - | ns | - |
| Shredders | - | - | - | - | ns | - | - | - | - | ns |
Appendix B
| Channel | Month | Site | Order | Family | Genus | Species | FFG | Abundance |
|---|---|---|---|---|---|---|---|---|
| MOLO | April | 2 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 1 |
| MOLO | April | 2 | Diptera | Ceratopogonidae | Gathering collectors | 5 | ||
| MOLO | April | 2 | Diptera | Chironomidae | Gathering collectors | 47 | ||
| MOLO | April | 2 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 9 |
| MOLO | April | 2 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 4 |
| MOLO | April | 2 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 6 |
| MOLO | April | 2 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 11 |
| MOLO | April | 2 | Odonata | Platycnemididae | Platycnemis | pennipes | Predators | 1 |
| MOLO | April | 2 | Bivalvia | Sphaeridae | Sphaerium | sp. | Filtering collectors | 1 |
| MOLO | April | 2 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 3 |
| MOLO | April | 3 | Diptera | Ceratopogonidae | Gathering collectors | 7 | ||
| MOLO | April | 3 | Diptera | Chironomidae | Gathering collectors | 120 | ||
| MOLO | April | 3 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 5 |
| MOLO | April | 3 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 1 |
| MOLO | April | 3 | Coleoptera | Haliplidae | Haliplus | sp. | Predators | 1 |
| MOLO | April | 3 | Odonata | Coenagrionidae | Ischnura | elegans | Predators | 2 |
| MOLO | April | 3 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 8 |
| MOLO | April | 3 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 7 |
| MOLO | April | 3 | Megaloptera | Sialidae | Sialis | sp. | Predators | 1 |
| MOLO | April | 4 | Trichoptera | Limnephilidae | Anabolia | nervosa | Shredders | 2 |
| MOLO | April | 4 | Crustacea | Asellidae | Asellus | aquaticus | Gathering collectors | 3 |
| MOLO | April | 4 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 2 |
| MOLO | April | 4 | Diptera | Ceratopogonidae | Gathering collectors | 11 | ||
| MOLO | April | 4 | Diptera | Chironomidae | Gathering collectors | 96 | ||
| MOLO | April | 4 | Odonata | Coenagrionidae | Coenagrion | puella | Predators | 9 |
| MOLO | April | 4 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 6 |
| MOLO | April | 4 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 9 |
| MOLO | April | 4 | Coleoptera | Haliplidae | Haliplus | sp. | Predators | 4 |
| MOLO | April | 4 | Trichoptera | Hydroptilidae | Hydroptila | sp. | Grazers | 1 |
| MOLO | April | 4 | Gastropoda | Planorbidae | Menetus | dilatatus | Grazers | 2 |
| MOLO | April | 4 | Odonata | Libellulidae | Orthetrum | albistylum | Predators | 1 |
| MOLO | April | 4 | Odonata | Platycnemididae | Platycnemis | pennipes | Predators | 3 |
| MOLO | April | 4 | Gastropoda | Tateidae | Potamopyrgus | antipodarum | Gathering collectors | 1 |
| MOLO | April | 4 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 1 |
| MOLO | April | 4 | Odonata | Zygoptera | Predators | 8 | ||
| MOLO | May | 1 | Crustacea | Asellidae | Asellus | aquaticus | Gathering collectors | 1 |
| MOLO | May | 1 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 3 |
| MOLO | May | 1 | Diptera | Chironomidae | Gathering collectors | 76 | ||
| MOLO | May | 1 | Crustacea | Crangonyctidae | Crangonyx | pseudogracilis | Shredders | 1 |
| MOLO | May | 1 | Odonata | Coenagrionidae | Erythromma | najas/viridulum | Predators | 12 |
| MOLO | May | 1 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 17 |
| MOLO | May | 1 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 53 |
| MOLO | May | 1 | Odonata | Coenagrionidae | Ischnura | elegans | Predators | 3 |
| MOLO | May | 1 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 2 |
| MOLO | May | 1 | Gastropoda | Valvatidae | Valvata | cristata | Grazers | 2 |
| MOLO | May | 1 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 1 |
| MOLO | May | 1 | Odonata | Zygoptera | Predators | 11 | ||
| MOLO | May | 2 | Trichoptera | Limnephilidae | Anabolia | nervosa | Shredders | 1 |
| MOLO | May | 2 | Crustacea | Asellidae | Asellus | aquaticus | Gathering collectors | 2 |
| MOLO | May | 2 | Diptera | Athericidae | Predators | 1 | ||
| MOLO | May | 2 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 2 |
| MOLO | May | 2 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 2 |
| MOLO | May | 2 | Diptera | Ceratopogonidae | Gathering collectors | 3 | ||
| MOLO | May | 2 | Diptera | Chironomidae | Gathering collectors | 40 | ||
| MOLO | May | 2 | Odonata | Coenagrionidae | Coenagrion | puella | Predators | 14 |
| MOLO | May | 2 | Crustacea | Crangonyctidae | Crangonyx | pseudogracilis | Shredders | 5 |
| MOLO | May | 2 | Odonata | Coenagrionidae | Erythromma | najas/viridulum | Predators | 9 |
| MOLO | May | 2 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 41 |
| MOLO | May | 2 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 432 |
| MOLO | May | 2 | Odonata | Coenagrionidae | Ischnura | elegans | Predators | 1 |
| MOLO | May | 2 | Odonata | Lestidae | Lestes | viridis | Predators | 2 |
| MOLO | May | 2 | Trichoptera | Limnephilidae | Limnephilus | lunatus | Shredders | 2 |
| MOLO | May | 2 | Gastropoda | Planorbidae | Menetus | dilatatus | Grazers | 1 |
| MOLO | May | 2 | Micropterna | lateralis/sequax | Shredders | 1 | ||
| MOLO | May | 2 | Odonata | Platycnemididae | Platycnemis | pennipes | Predators | 5 |
| MOLO | May | 2 | Gastropoda | Tateidae | Potamopyrgus | antipodarum | Gathering collectors | 1 |
| MOLO | May | 2 | Megaloptera | Sialidae | Sialis | sp. | Predators | 2 |
| MOLO | May | 2 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 2 |
| MOLO | May | 3 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 4 |
| MOLO | May | 3 | Diptera | Ceratopogonidae | Gathering collectors | 7 | ||
| MOLO | May | 3 | Diptera | Chironomidae | Gathering collectors | 171 | ||
| MOLO | May | 3 | Heteroptera | Corixidae | Corixinae | sp. | Gathering collectors | 3 |
| MOLO | May | 3 | Dreissena | polymorpha | Filtering collectors | 1 | ||
| MOLO | May | 3 | Odonata | Coenagrionidae | Erythromma | najas/viridulum | Predators | 12 |
| MOLO | May | 3 | Crsutacea | Gammaridae | Shredders | 1 | ||
| MOLO | May | 3 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 41 |
| MOLO | May | 3 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 116 |
| MOLO | May | 3 | Coleoptera | Haliplidae | Haliplus | sp. | Predators | 5 |
| MOLO | May | 3 | Hydracarina | Predators | 18 | |||
| MOLO | May | 3 | Trichoptera | Hydroptilidae | Hydroptila | sp. | Grazers | 2 |
| MOLO | May | 3 | Gastropoda | Planorbidae | Menetus | dilatatus | Grazers | 3 |
| MOLO | May | 3 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 2 |
| MOLO | May | 3 | Bivalvia | Sphaeridae | Musculium | lacustre | Filtering collectors | 6 |
| MOLO | May | 3 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 45 |
| MOLO | May | 3 | Odonata | Platycnemididae | Platycnemis | pennipes | Predators | 1 |
| MOLO | May | 3 | Gastropoda | Tateidae | Potamopyrgus | antipodarum | Gathering collectors | 2 |
| MOLO | May | 3 | Megaloptera | Sialidae | Sialis | sp. | Predators | 4 |
| MOLO | May | 3 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 7 |
| MOLO | June | 1 | Hirudinida | Glossiphonidae | Glossiphonia | sp. | Predators | 3 |
| MOLO | June | 1 | Trichoptera | Limnephilidae | Anabolia | nervosa | Shredders | 1 |
| MOLO | June | 1 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 1 |
| MOLO | June | 1 | Diptera | Ceratopogonidae | Gathering collectors | 1 | ||
| MOLO | June | 1 | Diptera | Chironomidae | Gathering collectors | 22 | ||
| MOLO | June | 1 | Heteroptera | Corixidae | Corixinae | sp. | Gathering collectors | 2 |
| MOLO | June | 1 | Odonata | Coenagrionidae | Erythromma | najas/viridulum | Predators | 12 |
| MOLO | June | 1 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 272 |
| MOLO | June | 1 | Gastropoda | Planorbidae | Haitia | acuta | Shredders | 1114 |
| MOLO | June | 1 | Gastropoda | Planorbidae | Menetus | dilatatus | Grazers | 18 |
| MOLO | June | 1 | Gastropoda | Tateidae | Potamopyrgus | antipodarum | Gathering collectors | 1 |
| MOLO | June | 1 | Megaloptera | Sialidae | Sialis | sp. | Predators | 2 |
| MOLO | June | 1 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 23 |
| MOLO | June | 2 | Diptera | Chironomidae | Gathering collectors | 12 | ||
| MOLO | June | 2 | Crustacea | Crangonyctidae | Crangonyx | pseudogracilis | Shredders | 1 |
| MOLO | June | 2 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 10 |
| MOLO | June | 2 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 164 |
| MOLO | June | 2 | Coleoptera | Dytiscidae | Laccophilus | sp. | Predators | 1 |
| MOLO | June | 2 | Odonata | Lestidae | Lestes | viridis | Predators | 6 |
| MOLO | June | 2 | Bivalvia | Sphaeridae | Musculium | lacustre | Filtering collectors | 1 |
| MOLO | June | 2 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 1 |
| MOLO | June | 2 | Megaloptera | Sialidae | Sialis | sp. | Predators | 1 |
| MOLO | June | 2 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 1 |
| MOLO | June | 3 | Diptera | Chironomidae | Gathering collectors | 20 | ||
| MOLO | June | 3 | Odonata | Coenagrionidae | Coenagrion | puella | Predators | 1 |
| MOLO | June | 3 | Odonata | Coenagrionidae | Erythromma | najas/viridulum | Predators | 10 |
| MOLO | June | 3 | Heteroptera | Gerridae | Gerris | sp. | Predators | 1 |
| MOLO | June | 3 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 113 |
| MOLO | June | 3 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 431 |
| MOLO | June | 3 | Coleoptera | Dytiscidae | Laccophilus | sp. | Predators | 2 |
| MOLO | June | 3 | Odonata | Lestidae | Lestes | viridis | Predators | 3 |
| MOLO | June | 3 | Trichoptera | Limnephilidae | Limnephilus | lunatus | Shredders | 1 |
| MOLO | June | 3 | Gastropoda | Planorbidae | Menetus | dilatatus | Grazers | 4 |
| MOLO | June | 3 | Odonata | Libellulidae | Orthetrum | albistylum | Predators | 1 |
| MOLO | June | 3 | Gastropoda | Tateidae | Potamopyrgus | antipodarum | Gathering collectors | 3 |
| MOLO | June | 3 | Megaloptera | Sialidae | Sialis | sp. | Predators | 5 |
| MOLO | June | 3 | Diptera | Tabanidae | Predators | 1 | ||
| MOLO | June | 3 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 30 |
| MOLO | July | 1 | Diptera | Chironomidae | Gathering collectors | 12 | ||
| MOLO | July | 1 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 48 |
| MOLO | July | 1 | Odonata | Lestidae | Lestes | viridis | Predators | 6 |
| MOLO | July | 1 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 3 |
| MOLO | July | 1 | Megaloptera | Sialidae | Sialis | sp. | Predators | 2 |
| MOLO | July | 1 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 10 |
| MOLO | July | 2 | Crustacea | Asellidae | Asellus | aquaticus | Gathering collectors | 1 |
| MOLO | July | 2 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 6 |
| MOLO | July | 2 | Diptera | Chironomidae | Gathering collectors | 64 | ||
| MOLO | July | 2 | Bivalvia | Corbiculidae | Corbicula | fluminea | Filtering collectors | 1 |
| MOLO | July | 2 | Heteroptera | Corixidae | Corixinae | sp. | Gathering collectors | 1 |
| MOLO | July | 2 | Odonata | Coenagrionidae | Erythromma | najas/viridulum | Predators | 3 |
| MOLO | July | 2 | Crsutacea | Gammaridae | Shredders | 8 | ||
| MOLO | July | 2 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 15 |
| MOLO | July | 2 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 85 |
| MOLO | July | 2 | Coleoptera | Haliplidae | Haliplus | sp. | Predators | 1 |
| MOLO | July | 2 | Hydracarina | Predators | 2 | |||
| MOLO | July | 2 | Coleoptera | Dytiscidae | Laccophilus | sp. | Predators | 1 |
| MOLO | July | 2 | Odonata | Lestidae | Lestes | viridis | Predators | 3 |
| MOLO | July | 2 | Gastropoda | Planorbidae | Menetus | dilatatus | Grazers | 5 |
| MOLO | July | 2 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 1 |
| MOLO | July | 2 | Gastropoda | Tateidae | Potamopyrgus | antipodarum | Gathering collectors | 1 |
| MOLO | July | 2 | Megaloptera | Sialidae | Sialis | sp. | Predators | 2 |
| MOLO | July | 2 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 5 |
| MOLO | July | 4 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 4 |
| MOLO | July | 4 | Diptera | Chironomidae | Gathering collectors | 4 | ||
| MOLO | July | 4 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 40 |
| MOLO | July | 4 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 135 |
| MOLO | July | 4 | Odonata | Lestidae | Lestes | viridis | Predators | 8 |
| MOLO | July | 4 | Gastropoda | Planorbidae | Menetus | dilatatus | Grazers | 2 |
| MOLO | July | 4 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 3 |
| MOLO | July | 4 | Heteroptera | Naucoridae | Naucoris | maculatus | Predators | 1 |
| MOLO | July | 4 | Lepidoptera | Crambidae | Parapoynx | stratiotata | Shredders | 4 |
| MOLO | July | 4 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 3 |
| MOLO | July | 4 | Megaloptera | Sialidae | Sialis | sp. | Predators | 3 |
| MOLO | July | 4 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 22 |
| PONT | April | 2 | Trichoptera | Limnephilidae | Anabolia | nervosa | Shredders | 1 |
| PONT | April | 2 | Crustacea | Asellidae | Asellus | aquaticus | Gathering collectors | 3 |
| PONT | April | 2 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 8 |
| PONT | April | 2 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 19 |
| PONT | April | 2 | Ephemroptera | Caenidae | Caenis | robusta | Gathering collectors | 5 |
| PONT | April | 2 | Diptera | Ceratopogonidae | Gathering collectors | 149 | ||
| PONT | April | 2 | Diptera | Chironomidae | Gathering collectors | 674 | ||
| PONT | April | 2 | Odonata | Coenagrionidae | Coenagrion | puella | Predators | 1 |
| PONT | April | 2 | Odonata | Coenagrionidae | Erythromma | najas/viridulum | Predators | 2 |
| PONT | April | 2 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 10 |
| PONT | April | 2 | Gastropoda | Planorbidae | Hippeutis | complanatus | Grazers | 2 |
| PONT | April | 2 | Hydracarina | Predators | 9 | |||
| PONT | April | 2 | Trichoptera | Hydroptilidae | Hydroptila | sp. | Grazers | 1 |
| PONT | April | 2 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 6 |
| PONT | April | 2 | Bivalvia | Sphaeridae | Musculium | lacustre | Filtering collectors | 18 |
| PONT | April | 2 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 11 |
| PONT | April | 2 | Bivalvia | Sphaeridae | Filtering collectors | 3 | ||
| PONT | April | 2 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 25 |
| PONT | April | 3 | Trichoptera | Limnephilidae | Anabolia | nervosa | Shredders | 1 |
| PONT | April | 3 | Crustacea | Asellidae | Asellus | aquaticus | Gathering collectors | 4 |
| PONT | April | 3 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 11 |
| PONT | April | 3 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 5 |
| PONT | April | 3 | Diptera | Chironomidae | Gathering collectors | 132 | ||
| PONT | April | 3 | Odonata | Coenagrionidae | Coenagrion | puella | Predators | 2 |
| PONT | April | 3 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 4 |
| PONT | April | 3 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 1 |
| PONT | April | 3 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 1 |
| PONT | April | 3 | Odonata | Libellulidae | Orthetrum | albistylum | Predators | 2 |
| PONT | April | 3 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 3 |
| PONT | April | 3 | Bivalvia | Sphaeridae | Filtering collectors | 2 | ||
| PONT | April | 3 | Diptera | Tabanidae | Predators | 1 | ||
| PONT | April | 3 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 3 |
| PONT | April | 4 | Crustacea | Asellidae | Asellus | aquaticus | Gathering collectors | 7 |
| PONT | April | 4 | Trichoptera | Leptoceridae | Athripsodes | sp. | Shredders | 1 |
| PONT | April | 4 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 33 |
| PONT | April | 4 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 34 |
| PONT | April | 4 | Diptera | Ceratopogonidae | Gathering collectors | 98 | ||
| PONT | April | 4 | Diptera | Chironomidae | Gathering collectors | 214 | ||
| PONT | April | 4 | Odonata | Coenagrionidae | Coenagrion | puella | Predators | 1 |
| PONT | April | 4 | Hirudinida | Glossiphonidae | Glossiphonia | sp. | Predators | 2 |
| PONT | April | 4 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 9 |
| PONT | April | 4 | Odonata | Coenagrionidae | Ischnura | elegans | Predators | 2 |
| PONT | April | 4 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 18 |
| PONT | April | 4 | Trichoptera | Leptoceridae | Mystacides | azurea | Gathering collectors | 1 |
| PONT | April | 4 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 80 |
| PONT | April | 4 | Odonata | Platycnemididae | Platycnemis | pennipes | Predators | 6 |
| PONT | April | 4 | Gastropoda | Tateidae | Potamopyrgus | antipodarum | Gathering collectors | 1 |
| PONT | April | 4 | Bivalvia | Sphaeridae | Filtering collectors | 4 | ||
| PONT | April | 4 | Diptera | Tabanidae | Predators | 6 | ||
| PONT | April | 4 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 20 |
| PONT | May | 1 | Crustacea | Asellidae | Asellus | aquaticus | Gathering collectors | 1 |
| PONT | May | 1 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 5 |
| PONT | May | 1 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 5 |
| PONT | May | 1 | Diptera | Ceratopogonidae | Gathering collectors | 20 | ||
| PONT | May | 1 | Diptera | Chironomidae | Gathering collectors | 61 | ||
| PONT | May | 1 | Odonata | Coenagrionidae | Erythromma | najas/viridulum | Predators | 2 |
| PONT | May | 1 | Hirudinida | Glossiphonidae | Glossiphonia | sp. | Predators | 2 |
| PONT | May | 1 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 3 |
| PONT | May | 1 | Trichoptera | Limnephilidae | Limnephilus | lunatus | Shredders | 1 |
| PONT | May | 1 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 1 |
| PONT | May | 1 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 7 |
| PONT | May | 1 | Gastropoda | Tateidae | Potamopyrgus | antipodarum | Gathering collectors | 1 |
| PONT | May | 1 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 9 |
| PONT | May | 2 | Crustacea | Asellidae | Asellus | aquaticus | Gathering collectors | 2 |
| PONT | May | 2 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 5 |
| PONT | May | 2 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 16 |
| PONT | May | 2 | Diptera | Ceratopogonidae | Gathering collectors | 33 | ||
| PONT | May | 2 | Diptera | Chironomidae | Gathering collectors | 57 | ||
| PONT | May | 2 | Odonata | Coenagrionidae | Coenagrion | puella | Predators | 4 |
| PONT | May | 2 | Hirudinida | Glossiphonidae | Glossiphonia | sp. | Predators | 4 |
| PONT | May | 2 | Hydracarina | Predators | 7 | |||
| PONT | May | 2 | Bivalvia | Sphaeridae | Musculium | lacustre | Filtering collectors | 1 |
| PONT | May | 2 | Trichoptera | Leptoceridae | Oecetis | ochracea | Predators | 1 |
| PONT | May | 2 | Megaloptera | Sialidae | Sialis | sp. | Predators | 4 |
| PONT | May | 2 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 6 |
| PONT | May | 4 | Crustacea | Asellidae | Asellus | aquaticus | Gathering collectors | 3 |
| PONT | May | 4 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 2 |
| PONT | May | 4 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 21 |
| PONT | May | 4 | Diptera | Chironomidae | Gathering collectors | 41 | ||
| PONT | May | 4 | Odonata | Coenagrionidae | Erythromma | najas/viridulum | Predators | 4 |
| PONT | May | 4 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 3 |
| PONT | May | 4 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 2 |
| PONT | May | 4 | Bivalvia | Sphaeridae | Musculium | lacustre | Filtering collectors | 1 |
| PONT | May | 4 | Trichoptera | Leptoceridae | Mystacides | azurea | Gathering collectors | 1 |
| PONT | May | 4 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 6 |
| PONT | May | 4 | Odonata | Platycnemididae | Platycnemis | pennipes | Predators | 4 |
| PONT | May | 4 | Megaloptera | Sialidae | Sialis | sp. | Predators | 5 |
| PONT | May | 4 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 6 |
| PONT | June | 1 | Coleoptera | Dytiscidae | Laccophilus | sp. | Predators | 2 |
| PONT | June | 2 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 1 |
| PONT | June | 2 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 4 |
| PONT | June | 2 | Diptera | Ceratopogonidae | Gathering collectors | 6 | ||
| PONT | June | 2 | Diptera | Chironomidae | Gathering collectors | 93 | ||
| PONT | June | 2 | Odonata | Coenagrionidae | Coenagrion | puella | Predators | 3 |
| PONT | June | 2 | Hirudinida | Erpobdellidae | Predators | 1 | ||
| PONT | June | 2 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 2 |
| PONT | June | 2 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 1 |
| PONT | June | 2 | Hydracarina | Predators | 8 | |||
| PONT | June | 2 | Coleoptera | Dytiscidae | Laccophilus | sp. | Predators | 1 |
| PONT | June | 2 | Trichoptera | Leptoceridae | Oecetis | ochracea | Predators | 1 |
| PONT | June | 2 | Gastropoda | Tateidae | Potamopyrgus | antipodarum | Gathering collectors | 2 |
| PONT | June | 2 | Megaloptera | Sialidae | Sialis | sp. | Predators | 9 |
| PONT | June | 2 | Diptera | Tabanidae | Predators | 3 | ||
| PONT | June | 2 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 7 |
| PONT | June | 3 | Odonata | Aeschnidae | Aeschna | sp. | Predators | 1 |
| PONT | June | 3 | Crustacea | Asellidae | Asellus | aquaticus | Gathering collectors | 3 |
| PONT | June | 3 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 3 |
| PONT | June | 3 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 7 |
| PONT | June | 3 | Diptera | Ceratopogonidae | Gathering collectors | 10 | ||
| PONT | June | 3 | Diptera | Chironomidae | Gathering collectors | 131 | ||
| PONT | June | 3 | Heteroptera | Corixidae | Corixinae | sp. | Gathering collectors | 3 |
| PONT | June | 3 | Coleoptera | Dytiscidae | Dytiscus | sp. | Predators | 3 |
| PONT | June | 3 | Odonata | Coenagrionidae | Erythromma | najas/viridulum | Predators | 1 |
| PONT | June | 3 | Hirudinida | Glossiphonidae | Glossiphonia | sp. | Predators | 2 |
| PONT | June | 3 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 7 |
| PONT | June | 3 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 1 |
| PONT | June | 3 | Hydracarina | Predators | 7 | |||
| PONT | June | 3 | Odonata | Libellulidae | Libellula | despressa | Predators | 1 |
| PONT | June | 3 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 26 |
| PONT | June | 3 | Megaloptera | Sialidae | Sialis | sp. | Predators | 10 |
| PONT | June | 3 | Bivalvia | Sphaeridae | Filtering collectors | 6 | ||
| PONT | June | 3 | Diptera | Tabanidae | Predators | 1 | ||
| PONT | June | 3 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 10 |
| PONT | June | 4 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 1 |
| PONT | June | 4 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 3 |
| PONT | June | 4 | Diptera | Ceratopogonidae | Gathering collectors | 2 | ||
| PONT | June | 4 | Diptera | Chironomidae | Gathering collectors | 134 | ||
| PONT | June | 4 | Coleoptera | Dytiscidae | Dytiscus | sp. | Predators | 4 |
| PONT | June | 4 | Odonata | Coenagrionidae | Erythromma | najas/viridulum | Predators | 1 |
| PONT | June | 4 | Hirudinida | Glossiphonidae | Glossiphonia | sp. | Predators | 1 |
| PONT | June | 4 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 14 |
| PONT | June | 4 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 19 |
| PONT | June | 4 | Megaloptera | Sialidae | Sialis | sp. | Predators | 6 |
| PONT | June | 4 | Bivalvia | Sphaeridae | Filtering collectors | 1 | ||
| PONT | June | 4 | Diptera | Tabanidae | Predators | 1 | ||
| PONT | June | 4 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 1 |
| PONT | July | 1 | Trichoptera | Limnephilidae | Anabolia | nervosa | Shredders | 1 |
| PONT | July | 1 | Crustacea | Asellidae | Asellus | aquaticus | Gathering collectors | 17 |
| PONT | July | 1 | Trichoptera | Leptoceridae | Athripsodes | sp. | Shredders | 6 |
| PONT | July | 1 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 143 |
| PONT | July | 1 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 130 |
| PONT | July | 1 | Ephemeroptera | Caenidae | Caenis | luctuosa | Gathering collectors | 1 |
| PONT | July | 1 | Diptera | Ceratopogonidae | Gathering collectors | 10 | ||
| PONT | July | 1 | Diptera | Chironomidae | Gathering collectors | 354 | ||
| PONT | July | 1 | Heteroptera | Corixidae | Corixinae | sp. | Gathering collectors | 24 |
| PONT | July | 1 | Ephemeroptera | Ephemeridae | Ephemera | sp. | Filtering collectors | 24 |
| PONT | July | 1 | Odonata | Coenagrionidae | Erythromma | najas/viridulum | Predators | 2 |
| PONT | July | 1 | Odonata | Coenagrionidae | Erythromma | najas/viridulum | Predators | 7 |
| PONT | July | 1 | Hirudinida | Glossiphonidae | Glossiphonia | sp. | Predators | 33 |
| PONT | July | 1 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 13 |
| PONT | July | 1 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 3 |
| PONT | July | 1 | Coleoptera | Haliplidae | Haliplus | sp. | Predators | 4 |
| PONT | July | 1 | Gastropoda | Planorbidae | Hippeutis | complanatus | Grazers | 2 |
| PONT | July | 1 | Hydracarina | Predators | 18 | |||
| PONT | July | 1 | Trichoptera | Limnephilidae | Limnephilus | lunatus | Shredders | 1 |
| PONT | July | 1 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 50 |
| PONT | July | 1 | Trichoptera | Leptoceridae | Mystacides | azurea | Gathering collectors | 1 |
| PONT | July | 1 | Odonata | Libellulidae | Orthetrumalbistylum | Predators | 1 | |
| PONT | July | 1 | Gastropoda | Tateidae | Potamopyrgus | antipodarum | Gathering collectors | 5 |
| PONT | July | 1 | Megaloptera | Sialidae | Sialis | sp. | Predators | 8 |
| PONT | July | 1 | Bivalvia | Sphaeridae | Filtering collectors | 1 | ||
| PONT | July | 1 | Diptera | Tabanidae | Predators | 1 | ||
| PONT | July | 1 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 3 |
| PONT | July | 3 | Trichoptera | Leptoceridae | Athripsodes | sp. | Shredders | 1 |
| PONT | July | 3 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 22 |
| PONT | July | 3 | Diptera | Chironomidae | Gathering collectors | 54 | ||
| PONT | July | 3 | Heteroptera | Corixidae | Corixinae | sp. | Gathering collectors | 6 |
| PONT | July | 3 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 2 |
| PONT | July | 3 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 1 |
| PONT | July | 3 | Coleoptera | Haliplidae | Haliplus | sp. | Predators | 3 |
| PONT | July | 3 | Coleoptera | Dytiscidae | Laccophilus | sp. | Predators | 1 |
| PONT | July | 3 | Odonata | Lestidae | Lestes | viridis | Predators | 2 |
| PONT | July | 3 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 7 |
| PONT | July | 3 | Bivalvia | Sphaeridae | Pisidium | sp. | Filtering collectors | 7 |
| PONT | July | 3 | Megaloptera | Sialidae | Sialis | sp. | Predators | 4 |
| PONT | July | 3 | Bivalvia | Sphaeridae | Filtering collectors | 8 | ||
| PONT | July | 3 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 17 |
| PONT | July | 4 | Crustacea | Asellidae | Asellus | aquaticus | Gathering collectors | 4 |
| PONT | July | 4 | Gastropoda | Bithynidae | Bithynia | tentaculata | Filtering collectors | 20 |
| PONT | July | 4 | Ephemeroptera | Caenidae | Caenis | horaria | Gathering collectors | 18 |
| PONT | July | 4 | Diptera | Chironomidae | Gathering collectors | 56 | ||
| PONT | July | 4 | Heteroptera | Corixidae | Corixinae | sp. | Gathering collectors | 2 |
| PONT | July | 4 | Gastropoda | Planorbidae | Gyraulus | sp. | Grazers | 2 |
| PONT | July | 4 | Gastropoda | Planorbidae | Haitia | acuta | Grazers | 1 |
| PONT | July | 4 | Coleoptera | Dytiscidae | Laccophilus | sp. | Predators | 2 |
| PONT | July | 4 | Odonata | Lestidae | Lestes | viridis | Predators | 2 |
| PONT | July | 4 | Heteroptera | Corixidae | Micronecta | sp. | Gathering collectors | 1 |
| PONT | July | 4 | Megaloptera | Sialidae | Sialis | sp. | Predators | 9 |
| PONT | July | 4 | Bivalvia | Sphaeridae | Filtering collectors | 3 | ||
| PONT | July | 4 | Gastropoda | Valvatidae | Valvata | piscinalis | Gathering collectors | 2 |
References
- Blackburn, T.M.; Gaston, K.J. Animal body size distributions: Patterns, mechanisms and implications. Trends Ecol. Evol. 1994, 9, 471–474. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Gaston, K.J. The relationship between animal abundance and body size: A review of the mechanisms. Adv. Ecol. Res. 1999, 28, 181–210. [Google Scholar]
- Woodward, G.U.Y.; Hildrew, A.G. Body size determinants of niche overlap and intraguild predation within a complex food web. J. Anim. Ecol. 2002, 71, 1063–1074. [Google Scholar] [CrossRef]
- Woodward, G.; Ebenman, B.; Emmerson, M.; Montoya, J.M.; Olesen, J.M.; Valido, A.; Warren, P.H. Body size in ecological networks. Trends Ecol. Evol. 2005, 20, 402–409. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.H. Macroecology; University of Chicago Press: Chicago, IL, USA, 1995. [Google Scholar]
- Gaston, K.J.; Blackburn, T.M. Pattern and Process in Macroecology; Blackwell Science: Oxford, UK, 2000. [Google Scholar]
- White, E.P.; Ernest, S.M.; Kerkhoff, A.J.; Enquist, B.J. Relationships between body size and abundance in ecology. Trends Ecol. Evol. 2007, 22, 323–330. [Google Scholar] [CrossRef] [Green Version]
- Damuth, J.D. Population density and body size in mammals. Nature 1981, 290, 699–700. [Google Scholar] [CrossRef]
- Damuth, J.D. Of size and abundance. Nature 1991, 351, 268–269. [Google Scholar] [CrossRef]
- Damuth, J.D. Population ecology: Common rules for animals and plants. Nature 1998, 395, 115–116. [Google Scholar] [CrossRef]
- Brown, J.H.; Gilloly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Brown, J.H.; Maurer, B.A. Evolution of species assemblages: Effects of energetic constraints and species dynamics on the diversification of the North American avifauna. Am. Nat. 1987, 130, 1–17. [Google Scholar] [CrossRef]
- Damuth, J. Cope’s rule, the island rule and the scaling of mammalian population density. Nature 1993, 365, 748–750. [Google Scholar] [CrossRef] [PubMed]
- Illius, A.W.; Gordon, I.J. Modelling the nutritional ecology of ungulate herbivores: Evolution of body size and competitive interactions. Oecologia 1992, 89, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Gaston, K.J.; Blackburn, T.M. Global scale macroecology: Interactions between population size, geographic range size and body size in the Anseriformes. J. Anim. Ecol. 1996, 65, 701–714. [Google Scholar] [CrossRef]
- Siemann, E.; Tilman, D.; Haarstad, J. Insect species diversity, abundance and body size relationships. Nature 1996, 380, 704–706. [Google Scholar] [CrossRef]
- Nee, S.; Read, A.F.; Greenwood, J.J.; Harvey, P.H. The relationship between abundance and body size in British birds. Nature 1991, 351, 312–313. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Harvey, P.H.; Pagel, M.D. Species number, population density and body size relationships in natural communities. J. Anim. Ecol. 1990, 59, 335–345. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Brown, V.K.; Doube, B.M.; Greenwood, J.J.; Lawton, J.H.; Stork, N.E. The relationship between abundance and body size in natural animal assemblages. J. Anim. Ecol. 1993, 62, 519–528. [Google Scholar] [CrossRef]
- Cyr, H.; Peters, R.H.; Downing, J.A. Population density and community size structure: Comparison of aquatic and terrestrial systems. Oikos 1997, 80, 139–149. [Google Scholar] [CrossRef]
- Jonsson, T.; Cohen, J.E.; Carpenter, S.R. Food webs, body size, and species abundance in ecological community description. Adv. Ecol. Res. 2005, 36, 1–84. [Google Scholar]
- Cyr, H. Individual energy use and the allometry of population density. In Scaling in Biology; Brown, J.H., West, G.B., Eds.; University Press: Oxford, UK, 2000; pp. 267–283. [Google Scholar]
- Marquet, P.A.; Navarrete, S.A.; Castilla, J.C. Scaling population-density to body size in rocky intertidal communities. Science 1990, 250, 1125–1127. [Google Scholar] [CrossRef]
- Schmid-Araya, J.M.; Schmid, P.E.; Majdi, N.; Traunspurger, W. Biomass and production of freshwater meiofauna: A review and a new allometric model. Hydrobiologia 2020, 847, 2681–2703. [Google Scholar] [CrossRef]
- Arim, M.; Berazategui, M.; Barreneche, J.M.; Ziegler, L.; Zarucki, M.; Abades, S.R. Determinants of density–body size scaling within food webs and tools for their detection. Adv. Ecol. Res. 2011, 45, 1–39. [Google Scholar]
- Gjoni, V.; Cozzoli, F.; Rosati, I.; Basset, A. Size-density relationships: A cross-community approach to benthic macroinvertebrates in Mediterranean and Black Sea lagoons. Estuar. Coast 2017, 40, 1142–1158. [Google Scholar] [CrossRef]
- Lawton, J.H. Species richness and population dynamics of animal assemblages. Patterns in body size: Abundance space. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1990, 330, 283–291. [Google Scholar]
- Cotgreave, P. The relationship between body size and population abundance in animals. Trends Ecol. Evol. 1993, 8, 244–248. [Google Scholar] [CrossRef]
- Marquet, P.A.; Navarrete, S.A.; Castilla, J.C. Body size, population density, and the energetic equivalence rule. J. Anim. Ecol. 1995, 64, 325–332. [Google Scholar] [CrossRef]
- Greve, M.; Gaston, K.J.; Van Rensburg, B.J.; Chown, S.L. Environmental factors, regional body size distributions and spatial variation in body size of local avian assemblages. Glob. Ecol. Biogeogr. 2008, 17, 514–523. [Google Scholar] [CrossRef]
- Kozłowski, J.; Gawelczyk, A.T. Why are species’ body size distributions usually skewed to the right? Funct. Ecol. 2002, 16, 419–432. [Google Scholar] [CrossRef]
- Statzner, B.; Moss, B. Linking ecological function, biodiversity and habitat: A mini-review focusing on older ecological literature. Basic Appl. Ecol. 2004, 5, 97–106. [Google Scholar] [CrossRef]
- Stead, T.K.; Schmid-Araya, J.M.; Schmid, P.E.; Hildrew, A.G. The distribution of body size in a stream community: One system, many patterns. J. Anim. Ecol. 2005, 74, 475–487. [Google Scholar] [CrossRef]
- Schmid, P.E.; Tokeshi, M.; Schmid-Araya, J.M. Scaling in stream communities. Proc. R. Soc. Lond. B 2002, 269, 2587–2594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid, P.E.; Tokeshi, M.; Schmid-Araya, J.M. Relation between population density and body size in stream communities. Science 2000, 289, 1557–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, S.S.; McCoy, E.D.; Mushinsky, H.R. Habitat Structure: The Physical Arrangement of Objects in Space; Chapman and Hall: London, UK, 1991. [Google Scholar]
- Stelzer, P.S.; Mazzuco, A.C.A.; Gomes, L.E.; Martins, J.; Netto, S.; Bernardino, A.F. Taxonomic and functional diversity of benthic macrofauna associated with rhodolith beds in SE Brazil. PeerJ 2021, 9, e11903. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.B.; Webster, J.R. The role of macroinvertebrates in stream ecosystem function. Annu. Rev. Entomol. 1996, 41, 115–139. [Google Scholar] [CrossRef]
- Cummins, K.W.; Merritt, R.W.; Andrade, P.C. The use of invertebrate functional groups to characterize ecosystem attributes in selected streams and rivers in south Brazil. Stud. Neotrop. Fauna Environ. 2005, 40, 69–89. [Google Scholar] [CrossRef]
- Olivier, J.M.; Carrel, G.; Lamouroux, N.; Dole-Olivier, M.J.; Malard, F.; Bravard, J.P.; Amoros, C. Chapter 7—The Rhône River Basin. In Rivers of Europe; Academic Press: Cambridge, MA, USA, 2009; pp. 247–295. [Google Scholar]
- Marle, P.; Riquier, J.; Timoner, P.; Mayor, H.; Slaveykova, V.I.; Castella, E. The interplay of flow processes shapes aquatic invertebrate successions in floodplain channels—A modelling applied to restoration scenarios. Sci. Total Environ. 2021, 750, 142081. [Google Scholar] [CrossRef]
- Blinn, W.; Shannon, J.P.; Stevens, L.E.; Carder, J.P. Consequences of fluctuating discharge for communities. J. N. Am. Benthol. Soc. 1995, 14, 233–248. [Google Scholar] [CrossRef]
- Humphries, P.; King, A.J.; Koehn, J.D. Fish, flows and flood plains: Links between freshwater fishes and their environment in the Murray-Darling River system, Australia. Environ. Biol. Fishes 1999, 56, 129–151. [Google Scholar] [CrossRef]
- King, A.J.; Humphries, P.; Lake, P.S. Fish recruitment on floodplains: The roles of patterns of flooding and life history characteristics. Can. J. Fish. Aquat. Sci. 2003, 60, 773–786. [Google Scholar] [CrossRef]
- Marle, P.; Rabarivelo, S.; Marechal, S.; Castella, E.; Rosset, V.; Roger, M. Light-trapped caddisfly assemblages in two floodplain reaches of the French upper Rhone River [Trichoptera]. Ephemera 2016, 18, 41–59. [Google Scholar]
- Smith, R.J. Use and misuse of the reduced major axis for line-fitting. Am. J. Phys. Anthropol. 2009, 140, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Glazier, D.S. A unifying explanation for diverse metabolic scaling in animals and plants. Biol. Rev. 2010, 85, 111–138. [Google Scholar] [CrossRef] [PubMed]
- Kerkhoff, A.J.; Enquist, B.J. Multiplicative by nature: Why logarithmic transformation is necessary in allometry. J. Theor. Biol. 2009, 257, 519–521. [Google Scholar] [CrossRef]
- Glazier, D.S. Log-transformation is useful for examining proportional relationships in allometric scaling. J. Theor. Biol. 2013, 334, 200–203. [Google Scholar] [CrossRef]
- Gjoni, V.; Ghinis, S.; Pinna, M.; Mazzotta, L.; Marini, G.; Ciotti, M.; Rosati, I.; Vignes, F.; Arima, S.; Basset, A. Patterns of functional diversity of macroinvertebrates across three aquatic ecosystem types, NE Mediterranean. MMS 2019, 20, 703–717. [Google Scholar] [CrossRef] [Green Version]
- Gjoni, V.; Basset, A. A cross-community approach to energy pathways across lagoon macroinvertebrate guilds. Estuar. Coast 2018, 41, 2433–2446. [Google Scholar] [CrossRef]
- Gjoni, V.; Glazier, D.S. A perspective on body size and abundance relationships across ecological communities. Biology 2020, 9, 42. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, J.H.; Gaston, J.F. A critical assessment of the form of the interspecific relationship between abundance and body size in animals. J. Anim. Ecol. 1997, 66, 233–249. [Google Scholar] [CrossRef]
- Pagel, M.D.; Harvey, P.H.; Godfray, H.C.J. Species-abundance, biomass, and resource-use distributions. Am. Nat. 1991, 138, 836–850. [Google Scholar] [CrossRef]
- Brawn, J.D.; Karr, J.R.; Nichols, J.D. Demography of birds in a Neotropical forest: Effects of allometry, taxonomy, and ecology. Ecology 1995, 76, 41–51. [Google Scholar] [CrossRef]
- Russo, S.E.; Robinson, S.K.; Terborgh, J. Size-abundance relationships in an Amazonian bird community: Implications for the energetic equivalence rule. Am. Nat. 2003, 161, 267–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazier, D.S. The 3/4-power law is not universal: Evolution of isometric, ontogenetic metabolic scaling in pelagic animals. BioScience 2006, 56, 325332. [Google Scholar] [CrossRef] [Green Version]
- Shokri, M.; Ciotti, M.; Vignes, F.; Gjoni, V.; Basset, A. Components of standard metabolic rate variability in three species of gammarids. Web. Ecol. 2019, 19, 1–13. [Google Scholar] [CrossRef]
- Cozzoli, F.; Shokri, M.; Ligetta, G.; Ciotti, M.; Gjoni, V.; Marocco, V.; Vignes, F.; Basset, A. Relationship between individual metabolic rate and patch departure behaviour: Evidence from aquatic gastropods. Oikos 2020, 129, 1657–1667. [Google Scholar] [CrossRef]
- Glazier, D.S.; Butler, E.M.; Lombardi, S.A.; Deptola, T.J.; Reese, A.J.; Satterthwaite, E.V. Ecological effects on metabolic scaling: Amphipod responses to fish predators in freshwater springs. Ecol. Monog. 2011, 81, 599–618. [Google Scholar] [CrossRef]
- Glazier, D.S.; Borrelli, J.J.; Hoffman, C.L. Effects of fish predators on the mass-related energetics of a keystone freshwater crustacean. Biology 2020, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Gjoni, V.; Basset, A.; Glazier, D.S. Temperature and predator cues interactively affect ontogenetic metabolic scaling of aquatic amphipods. Biol. Lett. 2020, 16, 20200267. [Google Scholar] [CrossRef]
- Glazier, D.S.; Gring, J.P.; Holsopple, J.R.; Gjoni, V. Temperature effects on metabolic scaling of a keystone freshwater crustacean depend on fish-predation regime. J. Exp. Biol. 2020, 223, jeb232322. [Google Scholar] [CrossRef]
- Gaston, K.J. Patterns in the local and regional dynamics of moth populations. Oikos 1988, 53, 49–57. [Google Scholar] [CrossRef]
- Morse, D.R.; Stork, N.E.; Lawton, J.H. Species number, species abundance and body length relationships of arboreal beetles in Bornean lowland rain forest trees. Ecol. Entomol. 1988, 13, 25–37. [Google Scholar] [CrossRef]
- Verberk, W.C.; Siepel, H.; Esselink, H. Life-history strategies in freshwater macroinvertebrates. Fresh. Biol. 2008, 53, 17221738. [Google Scholar] [CrossRef]
- McHugh, P.A.; Thompson, R.M.; Greig, H.S.; Warburton, H.J.; McIntosh, A.R. Habitat size influences food web structure in drying streams. Ecography 2015, 38, 700–712. [Google Scholar] [CrossRef]
- Tockner, K.; Pennetzdorfer, D.; Reiner, N.; Schiemer, F.; Ward, J.V. Hydrological connectivity, and the exchange of organic matter and nutrients in a dynamic river-floodplain system (Danube, Austria). Fresh. Biol. 1999, 41, 521–535. [Google Scholar] [CrossRef]
- Keckeis, S.; Baranyi, C.; Hein, T.; Holarek, C.; Riedler, P.; Schiemer, F. The significance of zooplankton grazing in a floodplain system of the River Danube. J. Plankton Res. 2003, 25, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Olden, J.D.; Konrad, C.P.; Melis, T.S.; Kennard, M.J.; Freeman, M.C.; Mims, M.C.; Williams, J.G. Are large-scale flow experiments informing the science and management of freshwater ecosystems? Front. Ecol. Environ. 2014, 12, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Poff, N.L.; Schmidt, J.C. How dams can go with the flow. Science 2016, 353, 1099–1100. [Google Scholar] [CrossRef]





| MOLO Channel | PONT Channel | ||||
|---|---|---|---|---|---|
| Physicochemical Factors | Month | Mean | SE | Mean | SE |
| Dissolved oxygen (mg/L) | April | 5.35 | 5.26 | 6.88 | 9.49 |
| May | 5.97 | 4.5 | 5.95 | 8.22 | |
| June | 5.52 | 3.76 | 3.47 | 4.92 | |
| July | 4.85 | 0.73 | 7.38 | 1.12 | |
| Temperature (°C) | April | 13.68 | 0.38 | 13.7 | 0.66 |
| May | 16.58 | 0.76 | 14.86 | 0.55 | |
| June | 19.36 | 0.95 | 18.23 | 0.31 | |
| July | 21.86 | 0.28 | 21.55 | 0.46 | |
| Conductivity (μS/cm) | April | 259.49 | 29.62 | 242.08 | 3.31 |
| May | 293.32 | 31.73 | 295.08 | 17.28 | |
| June | 242.58 | 5.51 | 271.00 | 0.44 | |
| July | 260.8 | 20.9 | 275.02 | 6.01 | |
| Sampling Site | n | Slope | 95% CI | Intercept | r2 | p |
|---|---|---|---|---|---|---|
| MOLO | 162 | −0.39 | −0.53 to −0.24 | −0.19 | 0.19 | *** |
| PONT | 193 | −0.33 | −0.45 to −0.21 | −0.07 | 0.13 | *** |
| Sampling Site | Month | n | Slope | 95% CI | Intercept | r2 | p |
|---|---|---|---|---|---|---|---|
| MOLO | April | 35 | −0.31 | −0.56 to −0.06 | −0.10 | 0.16 | * |
| May | 53 | −0.42 | −0.69 to −0.14 | −0.22 | 0.15 | ** | |
| June | 38 | −0.48 | −0.85 to −0.10 | −0.35 | 0.16 | * | |
| July | 36 | −0.34 | −0.60 to −0.07 | −0.10 | 0.16 | * | |
| PONT | April | 50 | −0.53 | −0.79 to −0.27 | −0.38 | 0.26 | *** |
| May | 41 | −0.28 | −0.55 to −0.01 | −0.04 | 0.10 | * | |
| June | 55 | −0.26 | −0.49 to −0.02 | −0.04 | 0.10 | * | |
| July | 47 | −0.29 | −0.52 to −0.07 | 0.09 | 0.12 | * |
| Sampling Site | Functional Feeding Group | n | Slope | 95% CI | Intercept | r2 | p |
|---|---|---|---|---|---|---|---|
| MOLO | Filterers | 18 | −0.27 | −0.58 to 0.03 | −0.16 | 0.19 | * |
| Gatherers | 51 | −0.60 | −0.98 to −0.22 | −0.82 | 0.15 | *** | |
| Grazers | 31 | −0.31 | −0.80 to 0.17 | −0.42 | 0.06 | ns | |
| Predators | 48 | −0.02 | −0.22 to 0.18 | −0.41 | 0.00 | ns | |
| Shredders | 14 | −0.49 | −1.11 to 0.13 | −0.37 | 0.22 | * | |
| PONT | Filterers | 33 | −0.35 | −0.63 to −0.07 | 0.04 | 0.18 | ** |
| Gatherers | 75 | −0.45 | −0.79 to −0.12 | −0.15 | 0.10 | *** | |
| Grazers | 19 | −0.28 | −0.51 to −0.04 | −0.27 | 0.27 | * | |
| Predators | 58 | −0.16 | −0.29 to −0.04 | −0.07 | 0.11 | * | |
| Shredders | 8 | −0.04 | −0.32 to 0.23 | −0.01 | 0.02 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gjoni, V.; Marle, P.; Ibelings, B.W.; Castella, E. Size–Abundance Relationships of Freshwater Macroinvertebrates in Two Contrasting Floodplain Channels of Rhone River. Water 2022, 14, 794. https://doi.org/10.3390/w14050794
Gjoni V, Marle P, Ibelings BW, Castella E. Size–Abundance Relationships of Freshwater Macroinvertebrates in Two Contrasting Floodplain Channels of Rhone River. Water. 2022; 14(5):794. https://doi.org/10.3390/w14050794
Chicago/Turabian StyleGjoni, Vojsava, Pierre Marle, Bastiaan W. Ibelings, and Emmanuel Castella. 2022. "Size–Abundance Relationships of Freshwater Macroinvertebrates in Two Contrasting Floodplain Channels of Rhone River" Water 14, no. 5: 794. https://doi.org/10.3390/w14050794
APA StyleGjoni, V., Marle, P., Ibelings, B. W., & Castella, E. (2022). Size–Abundance Relationships of Freshwater Macroinvertebrates in Two Contrasting Floodplain Channels of Rhone River. Water, 14(5), 794. https://doi.org/10.3390/w14050794