
How Do Specific Environmental Conditions in Canals Affect the Structure and Variability of the Zooplankton Community?


Abstract
:1. Introduction
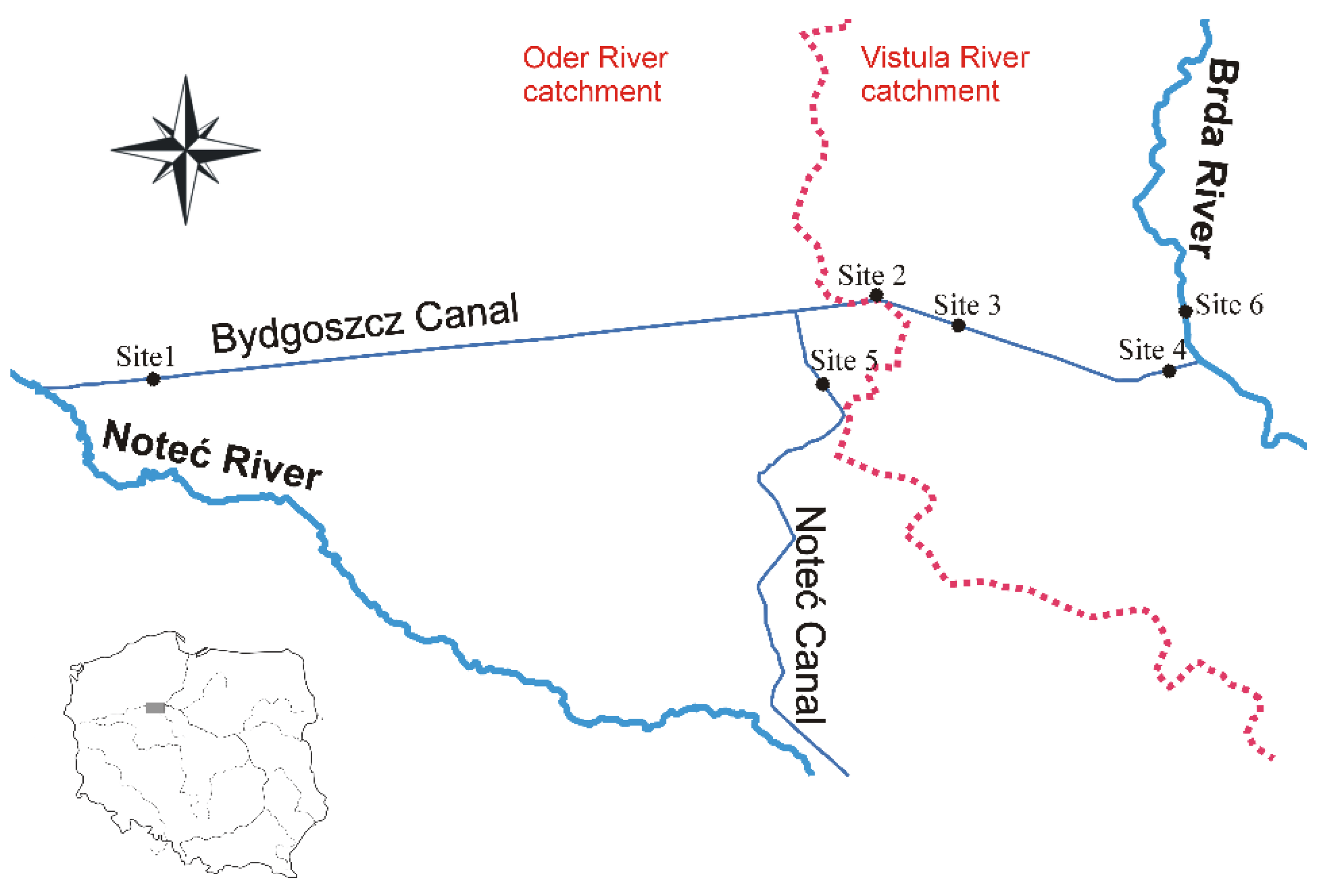
2. Materials and Methods
3. Results
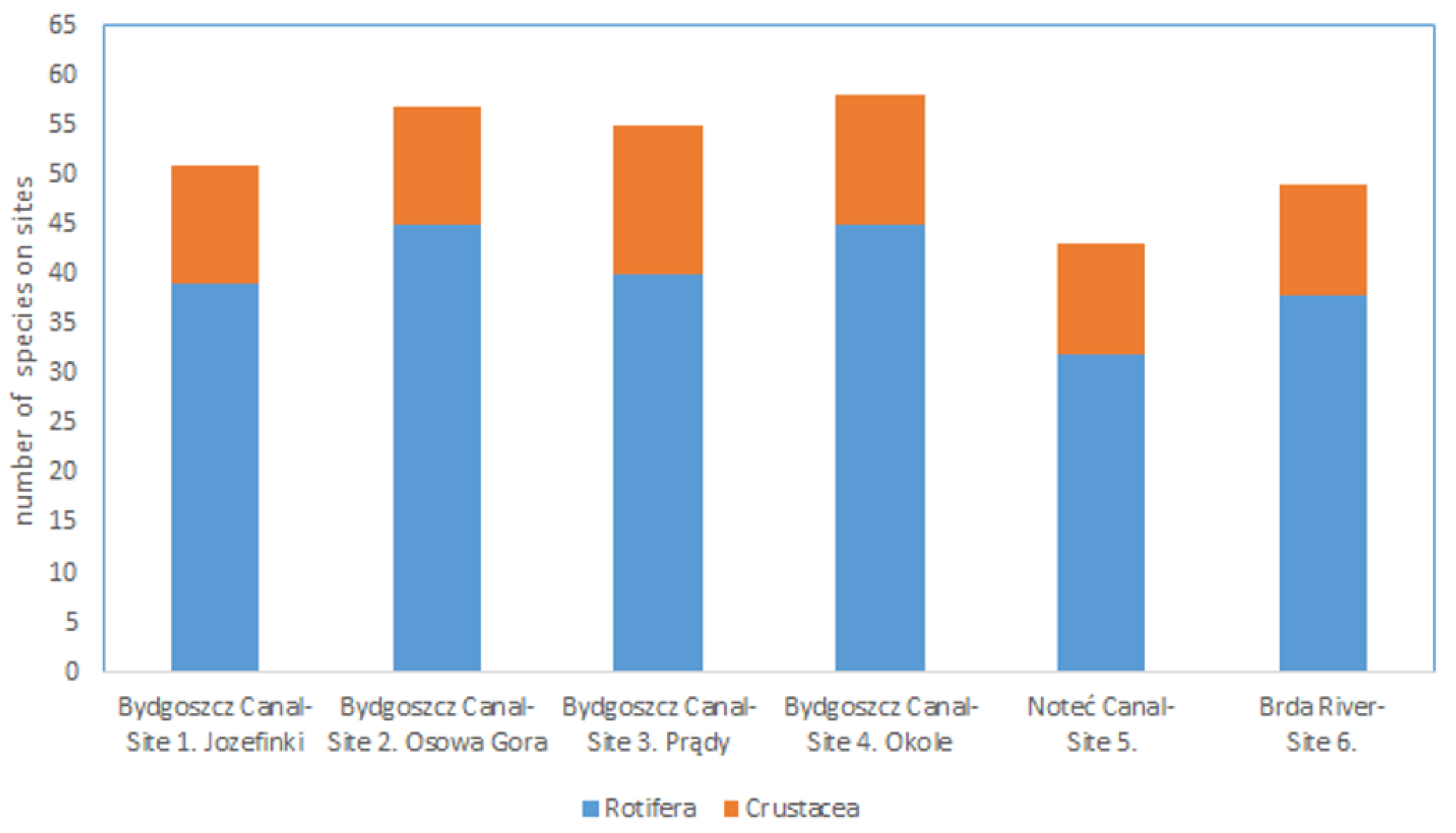
3.1. Taxonomic Diversity and Density
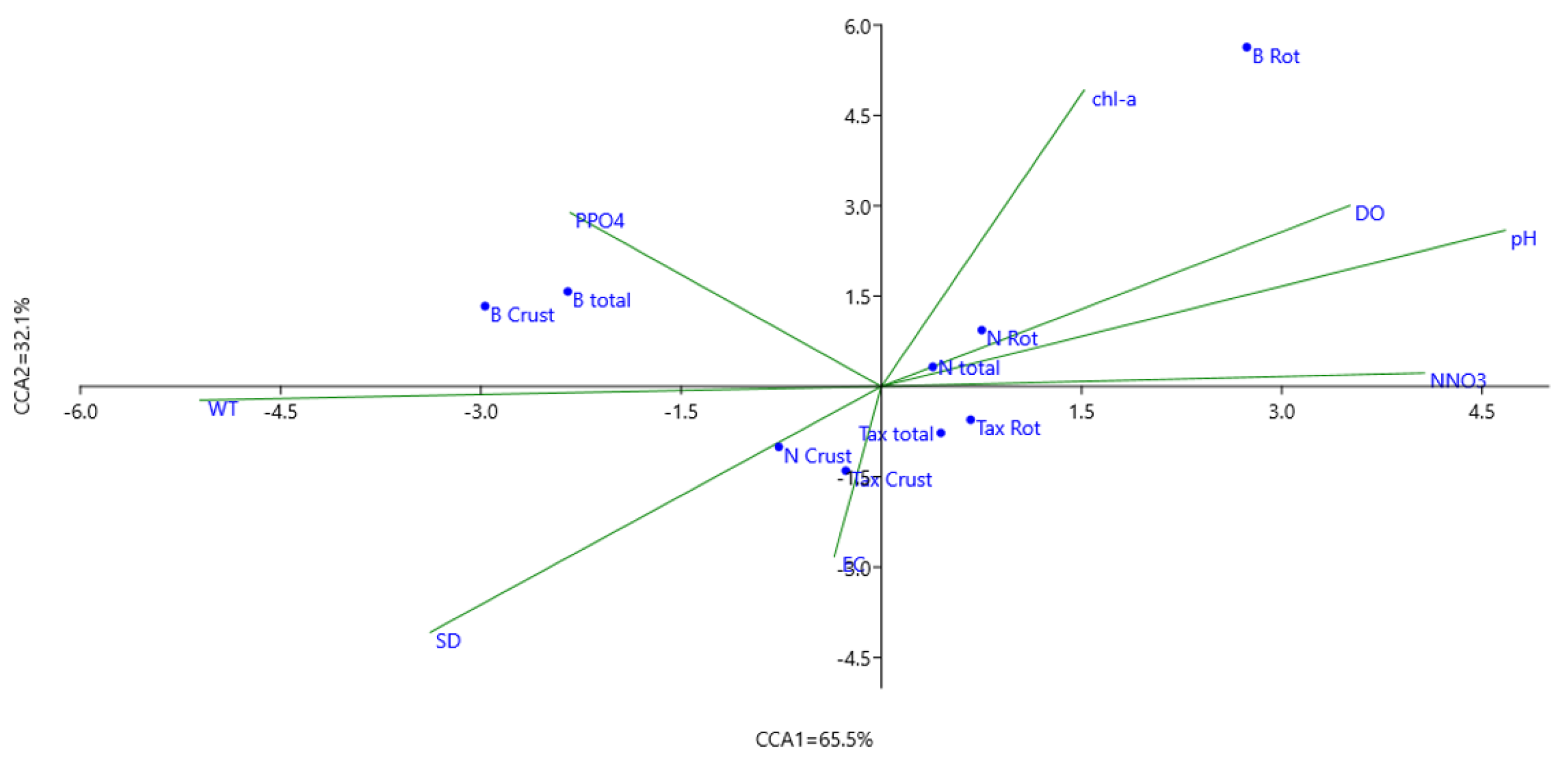
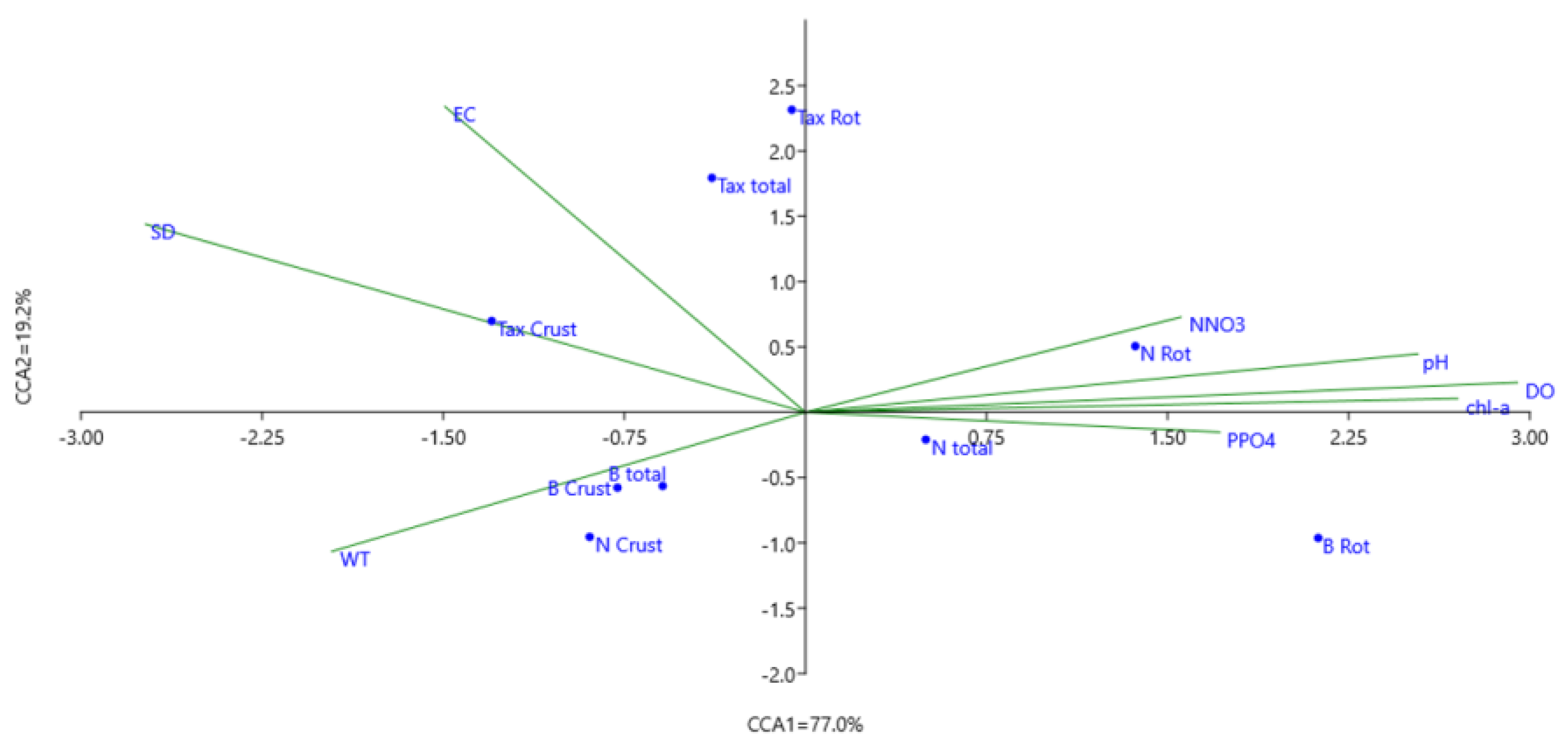
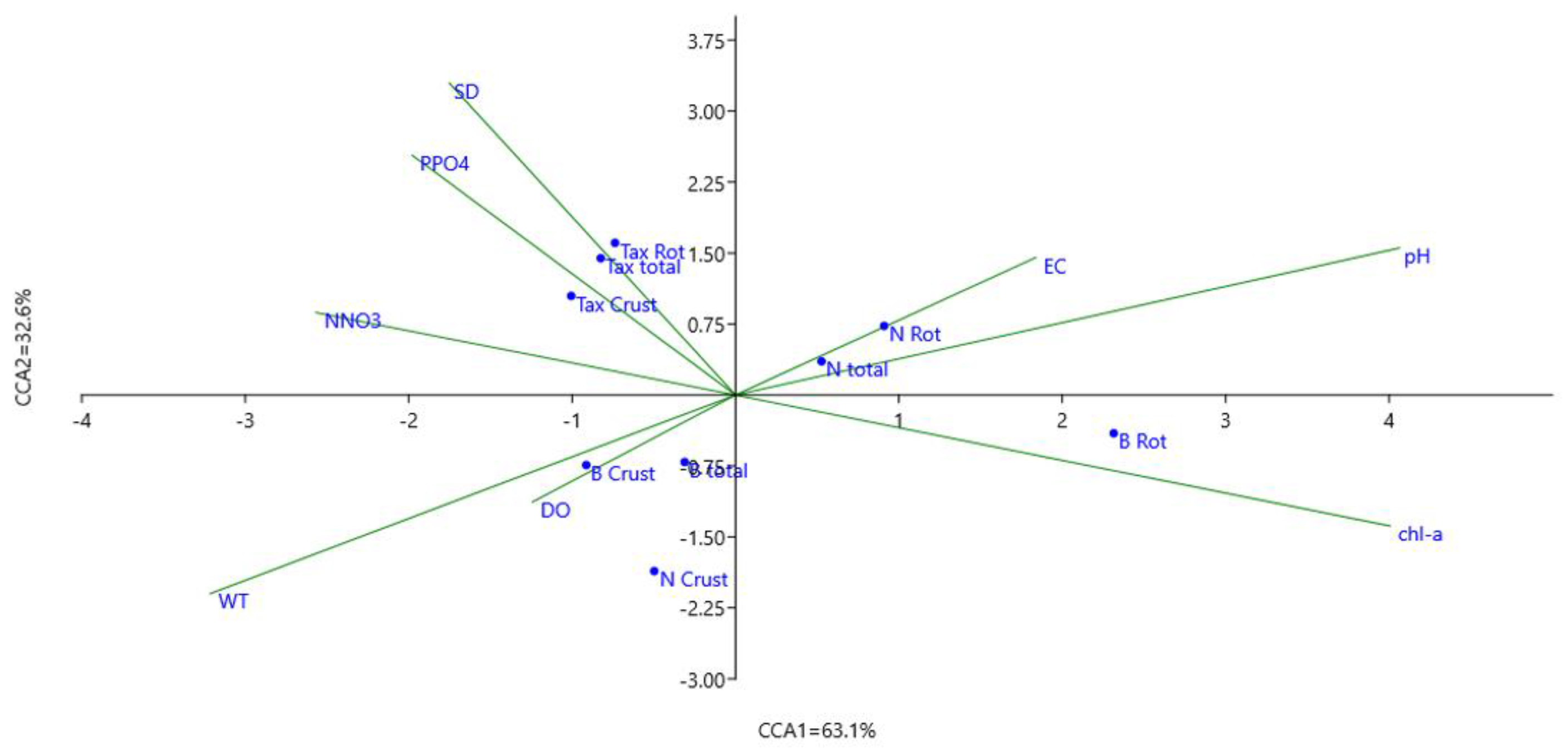
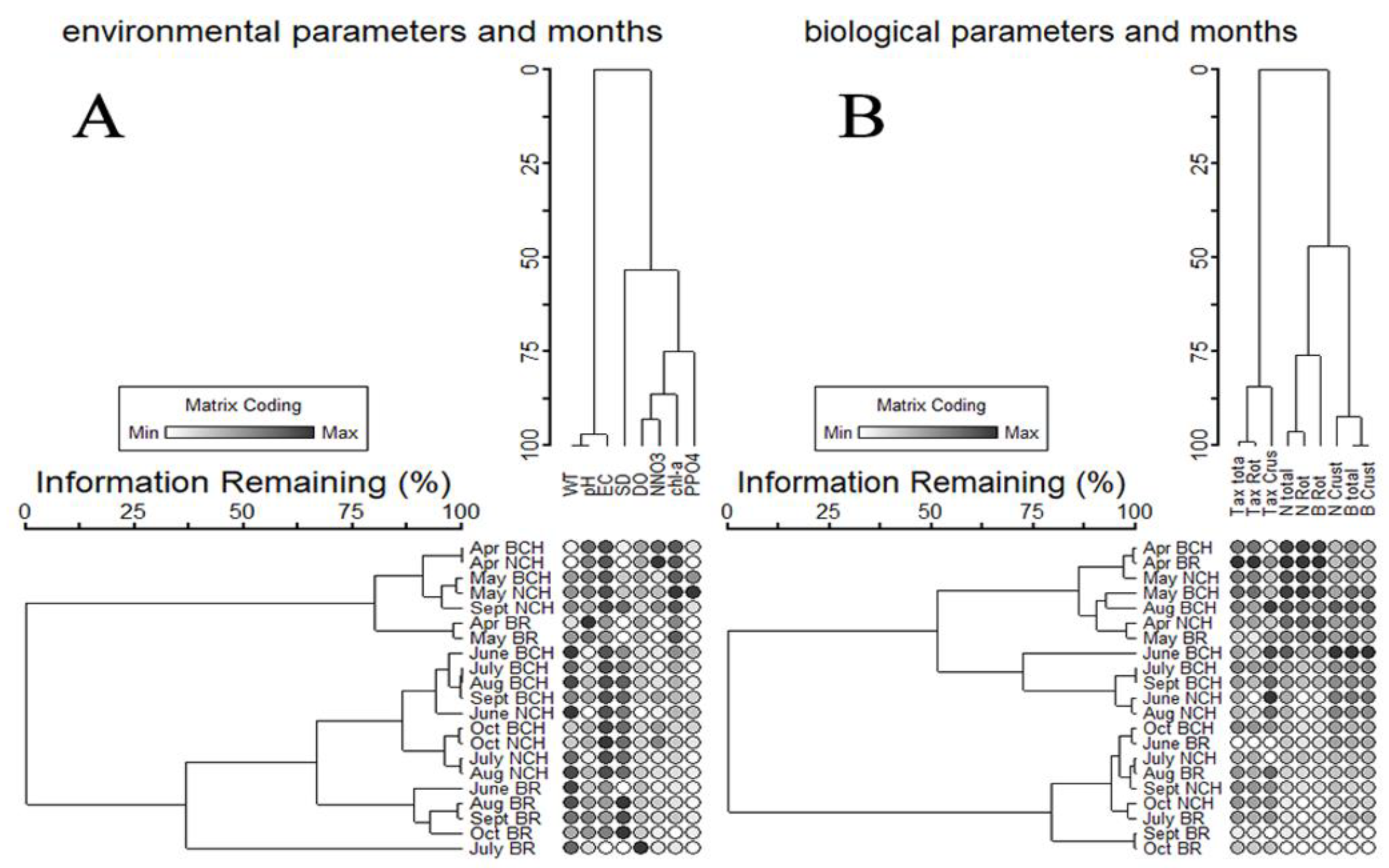
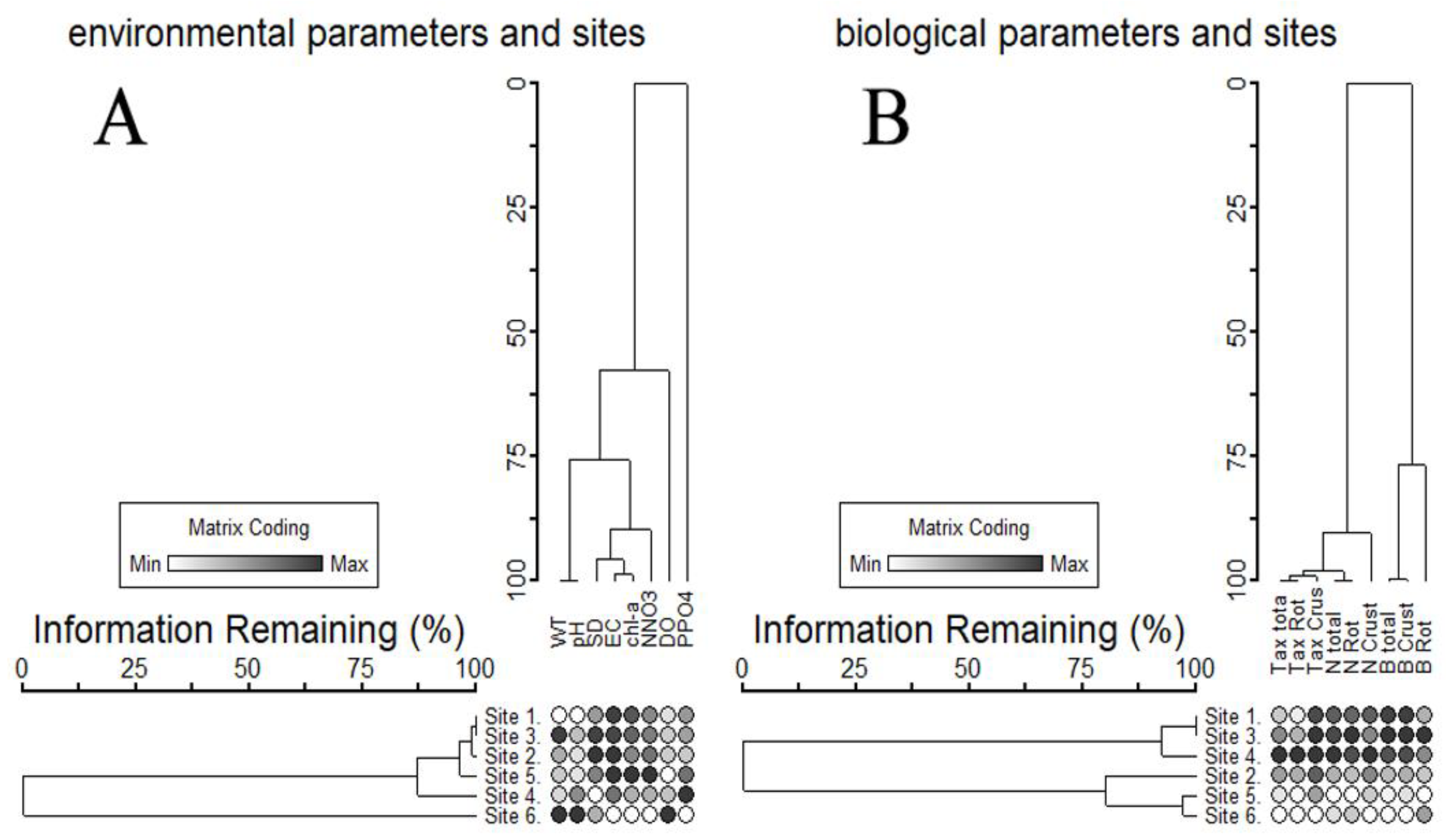
3.2. Influence of Environmental Factors on Zooplankton Communities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gorączko, M. About the need and possibilities to build a modern hydrological monitoring system within the Bydgoszcz Water Junction. Logistyka 2015, 3, 1534–1544. (In Polish) [Google Scholar]
- Florescu, L.; Parpală, L.; Dumitrache, A.; Moldoveanu, M. Spatial and Temporal Distribution of the Zooplankton Biomass in Sfântu Gheorghe Branch (The Danube Delta, Romania) in Relation to Environmental Factors. Trav. Muséum Natl. d’Histoire Nat. Grigore Antipa 2013, 56, 109–124. [Google Scholar] [CrossRef] [Green Version]
- Pringle, C.M. Hydrologic connectivity and the management of biological reserves: A global perspective. Ecol. Appl. 2001, 11, 981–998. [Google Scholar] [CrossRef]
- Pringle, C. What is hydrologic connectivity and why is it ecologically important? Hydrol. Process. 2003, 17, 2685–2689. [Google Scholar] [CrossRef]
- Vasylieva, O.M.; Novitskyi, R.O.; Hubanova, N.L.; Horchanok, A.V.; Sapronova, V.O. Dynamics of quality indicators of water status in the principal channel “Dnipro–Donbas” resulting of seasonal pumping. Agrology 2019, 2, 106–111. [Google Scholar] [CrossRef]
- Kim, J.; Mandrak, N.E. Assessing the potential movement of invasive fishes through the Welland Canal. J. Great Lakes Res. 2016, 42, 1102–1108. [Google Scholar] [CrossRef] [Green Version]
- Czerniawski, R.; Slugocki, L. Analysis of zooplankton assemblages from man-made ditches in relation to current velocity. Oceanol. Hydrobiol. Stud. 2017, 46, 199–211. [Google Scholar] [CrossRef]
- Sługocki, Ł.; Czerniawski, R.; Kowalska-Góralska, M.; Teixeira, C.A. Hydro-modifications matter: Influence of vale transformation on microinvertebrate communities (Rotifera, Cladocera, and Copepoda) of upland rivers. Ecol. Indic. 2021, 122, 107259. [Google Scholar] [CrossRef]
- Zhao, K.; Wang, L.; Riseng, C.; Wehrly, K.; Pan, Y.; Song, K.; Wang, Q. Factors determining zooplankton assemblage difference among a man-made lake, connecting canals, and the water-origin river. Ecol. Indic. 2018, 84, 488–496. [Google Scholar] [CrossRef]
- Napiórkowski, P.; Napiórkowska, T. The diversity and longitudinal changes of zooplankton in the lower course of a large, regulated European river (the lower Vistula River, Poland). Biologia 2013, 68, 1163–1171. [Google Scholar] [CrossRef] [Green Version]
- Sharma, C.; Tiwari, R.P. Studies on Zooplanktons of fresh water reservoir at Lony Dam Theonther Rewa (M.P.). Int. J. Pharm. Life Sci. 2011, 2, 492–495. [Google Scholar]
- Arya, A.; Singh, N. Assessment of water quality and zooplankton population of canal water in western U.P. Progress. Agric. 2017, 17, 66–71. [Google Scholar] [CrossRef]
- Majagi, S.; Naik, J.; Chitra, J. Seasonal Investigation on the Zooplankton Diversity and Distribution in Relation to Water Quality at Chikklingdalli Dam, Karnataka. Int. J. Res. Anal. Rev. 2019, 1, 754–767. [Google Scholar]
- Pace, M.L.; Findlay, S.E.; Lints, D. Zooplankton in advective environments: The Hudson River community and a comparative analysis. Can. J. Fish. Aquat. Sci. 1992, 49, 1060–1069. [Google Scholar] [CrossRef]
- Chang, K.H.; Doi, H.; Imai, H.; Gunji, F.; Nakano, S.I. Longitudinal changes in zooplankton distribution below a reservoir outfall with reference to river planktivory. Limnology 2008, 9, 125–133. [Google Scholar] [CrossRef]
- Czerniawski, R.; Pilecka-Rapacz, M. Summer zooplankton in small rivers in relation to selected conditions. Cent. Eur. J. Biol. 2011, 6, 659–674. [Google Scholar] [CrossRef] [Green Version]
- Ostroumov, S.A. Some aspects of water filtering activity of filter-feeders. Hydrobiologia 2005, 542, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Weber, M.J.; Brown, M.L. Effects of common carp on aquatic ecosystems 80 years after “carp as a dominant”: Ecological Insights for Fisheries Management. Rev. Fish. Sci. 2009, 17, 524–537. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Wardle, D.A. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Gomes, L.F.; Pereira, H.R.; Gomes, A.C.A.M.; Vieira, M.C.; Martins, P.R.; Roitman, I.; Vieira, L.C.G. Zooplankton functional-approach studies in continental aquatic environments: A systematic review. Aquat. Ecol. 2019, 53, 191–203. [Google Scholar] [CrossRef]
- Burdis, R.M.; Hoxmeier, R.J.H. Seasonal zooplankton dynamics in main channel and backwater habitats of the Upper Mississippi River. Hydrobiologia 2011, 667, 69–87. [Google Scholar] [CrossRef]
- Shivashankar, P.; Venkataramana, G.V. Zooplankton diversity of Chikkadevarayana canal in relation to physico-chemical characteristics. J. Environ. Biol. 2013, 34, 819–824. [Google Scholar]
- Jażdżewski, K.; Konopacka, A. Invasive Ponto-Caspian species in waters of the Vistula and Oder basins and of the southern Baltic Sea. In Invasive Aquatic Species of Europe; Leppökoski, E., Gollasch, S., Olenin, S., Eds.; Kluwer Academic Publisher: Dordrecht, The Netherlands, 2002; pp. 384–398. [Google Scholar] [CrossRef]
- Bazzuri, M.E.; Gabellone, N.A.; Solari, L.C. Zooplankton-population dynamics in the Salado-River basin (Buenos Aires, Argentina) in relation to hydraulic works and resulting wetland function. Aquat. Sci. 2020, 82, 48. [Google Scholar] [CrossRef]
- Abbas, M.F. Ecological study of zooplankton in the Shatt Al-Basrah canal, Basrah-Iraq. Mesopot. J. Mar. Sci. 2015, 30, 67–80. [Google Scholar]
- Napiórkowski, P.; Napiórkowska, T. The structure and spatial heterogeneity of zooplankton in small lake river Wel (Poland). Limnol. Rev. 2014, 14, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Ruwini Yasadari Perera, L.G. A preliminary study on the variation of zooplankton diversity, abundance and density in a selected branch of Diyawanna Oya Canal System. OUSL J. 2015, 9, 61–81. [Google Scholar] [CrossRef] [Green Version]
- MacIsaac, H.J.; Gilbert, J.J. Competition between Rotifers and Cladocerans of Different Body Sizes. Oecologia 1989, 81, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Romanovsky, Y.E.; Feniova, I.Y. Competition among Cladocera: Effect of Different Levels of Food Supply. Oikos 1985, 44, 243–252. [Google Scholar] [CrossRef]
- Fussmann, G. The importance of crustacean zooplankton in structuring rotifer and phytoplankton communities; an enclosure study. J. Plankton Res. 1996, 18, 1897–1915. [Google Scholar] [CrossRef] [Green Version]
- Babiński, Z.; Habel, M.; Szumińska, D. Mechanisms and causes of silting of the Bydgoszcz Canal. In Revitalization of the Vistula—Oder Waterway as an Opportunity for the Region’s Economy; Babiński, Z., Ed.; Logo Press: Bydgoszcz, Poland, 2008; pp. 65–78, (Summary in English). [Google Scholar]
- Babiński, Z.; Habel, M.; Szatten, D.; Dąbrowski, J. Influence of hydrological and sedimentological processes on the functioning of inland waterway transport on an example of Brdyujście Regatta Track. Sci. J. Marit. Univ. Szczec. 2014, 37, 16–21. [Google Scholar]
- Harris, R.P.; Wiebe, P.H.; Lenz, J.; Skjoldal, H.R.; Huntley, M. ICES Zooplankton Methodology Manual; Elsevier: Amsterdam, The Netherlands, 2000; 684p. [Google Scholar] [CrossRef]
- Nogrady, T. Rotifera: Biology, Ecology and Systematics. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Dumont, H.J., Ed.; SPB Academic Publishing: The Hague, The Netherlands, 1993; 142p. [Google Scholar]
- Napiórkowski, P.; Bąkowska, M.; Mrozińska, N.; Szymańska, M.; Kolarova, N.; Obolewski, K. The effect of hydrological connectivity on the zooplankton structure in floodplain lakes of a regulated large river (the lower Vistula, Poland). Water 2019, 11, 1924. [Google Scholar] [CrossRef] [Green Version]
- Einsle, U. Copepoda: Cyclopoida: Genera Cyclops, Megacyclops, Acanthocyclops. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World; Dumont, H.J., Ed.; SPB Academic Publishing: Amsterdam, The Netherlands, 1996; p. 83. [Google Scholar]
- Radwan, S.; Bielańska-Grajner, I.; Ejsmont-Karabin, J. Rotifers (Rotifera, Monogononta). In Freshwater Fauna of Poland; University of Lodz Press: Łódź, Poland, 2004; p. 579. [Google Scholar]
- Błędzki, L.A.; Rybak, J.I. Freshwater Crustacean Zooplankton of Europe. Cladocera and Copepoda (Calanoida, Cyclopoida) Key to species identification; Springer International Publishing: Cham, Switzerland, 2016. [Google Scholar] [CrossRef]
- Smirnov, N.N. Cladocera: The Chydorinae and Sayciinae (Chydoridae) of the world. In Guides to the Identification of the Microinvertebrates of the Continental Waters of the World 11; Dumont, H.J., Ed.; SPB Academic Publishing: Amsterdam, The Netherlands, 1996; p. 197. [Google Scholar]
- Ter Braak, C.J.; Verdonschot, P.F. Canonical correspondence analysis and related multivariate methods in aquatic ecology. Aquat. Sci. 1995, 57, 255–289. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistic software package for education and data analysis. Paleontol. Electron. 2001, 4, 178. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD: Multivariate Analysis of Ecological Data; Version 6 for Windows [User’s Guide]; MjM Software Design: Gleneden Beach, OR, USA, 2011. [Google Scholar]
- Demetraki-Paleolog, A. Planktonic rotifers diversity in selected rivers of the Vistula, Wieprz and San drainage-basins. Teka Kom. Ochr. Kszt. Środ. Przyr. OL PAN 2004, 1, 44–50. [Google Scholar]
- Marneffe, Y.; Descy, J.P.; Thomé, J.P. The zooplankton of the lower river Meuse, Belgium: Seasonal changes and impact of industrial and municipal discharges. Hydrobiologia 1996, 319, 1–13. [Google Scholar] [CrossRef]
- Chará-Serna, A.M.; Casper, A. How do large river zooplankton communities respond to abiotic and biotic drivers over time? A complex and spatial dependent example. Freshw. Biol. 2020, 66, 391–405. [Google Scholar] [CrossRef]
- Scheffer, M.; Jeppesen, E. Alternative stable state. In Ecology of Shallow Lakes; Scheffer, M., Ed.; Chapman and Hall: London, UK, 1998; 357p. [Google Scholar] [CrossRef]
- Arora, J.; Mehra, N.K. Species diversity of planktonic and epiphytic rotifers in the backwaters of the Delhi segment of the Yamuna River, with remarks on new records from India. Zool. Stud. Taipei 2003, 42, 239–247. [Google Scholar]
- Dumont, H.J. Workshop on taxonomy and biogeography. In Rotatoria. Developments in Hydrobiology; Dumont, H.J., Green, J., Eds.; Springer: Dordrecht, The Netherlands, 1980; Volume 1, pp. 205–206. [Google Scholar] [CrossRef]
- Dumont, H.J. Biogeography of rotifers. In Biology of Rotifers; Pejler, B., Starkweather, R., Nogrady, T., Eds.; Springer: Dordrecht, The Netherlands, 1983; pp. 19–30. [Google Scholar] [CrossRef]
- Armengol, X.; Esparcia, A.; Miracle, M.R. Rotifer vertical distribution in a strongly stratified lake: A multivariate analysis. Hydrobiologia 1998, 387–388, 161–170. [Google Scholar] [CrossRef]
- Deveci, A.; Dügel, M.; Külköylüoğlu, O. Zooplankton of Lake Sünnet (Bolu, Turkey) and determination of some environmental variables. Rev. Hydrobiol. 2011, 4, 115–130. [Google Scholar]
- Lair, N. Abiotic vs. biotic factors: Lessons drawn from rotifers in the Middle Loire, a meandering river monitored from 1995 to 2002, during low flow periods. In Rotifera X. Developments in Hydrobiology; Herzig, A., Gulati, R.D., Jersabek, C.D., May, L., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 181. [Google Scholar] [CrossRef]
- Branco, C.W.C.; Silveira, R.D.M.L.; Marinho, M.M. Flood pulse acting on a zooplankton community in a tropical river (upper Paraguay river, northern Pantanal, Brazil). Fundam. Appl. Limnol. 2018, 192, 23–42. [Google Scholar] [CrossRef]
- Jiménez-Contreras, J.; Nandini, S.; Sarma, S.S.S. Diversity of Rotifera (Monogononta) and egg ratio of selected taxa in the canals of Xochimilco (Mexico City). Wetlands 2018, 38, 1033–1044. [Google Scholar] [CrossRef]
- Kim, H.W.; Joo, G.J.; Walz, N. Zooplankton Dynamics in the Hyper-Eutrophic Nakdong River System (Korea) Regulated by an Estuary Dam and Side Channels. Int. Rev. Hydrobiol. 2001, 86, 127–143. [Google Scholar] [CrossRef]
- Kuczyńska-Kippen, N.M.; Nagengast, B.; Celewicz-Gołdyn, S.; Klimko, M. Zooplankton community structure within various macrophyte stands of a small water body in relation to seasonal changes in water level. Oceanol. Hydrobiol. Stud. 2009, 38, 125–133. [Google Scholar] [CrossRef]
- Karpowicz, M.; Ejsmont-Karabin, J.; Strzałek, M. Biodiversity of zooplankton (Rotifera and crustacea) in water soldier (Stratiotes aloides) habitats. Biologia 2016, 71, 563–573. [Google Scholar] [CrossRef]
- Kentzer, A.; Dembowska, E.; Giziński, A.; Napiórkowski, P. Influence of the Włocławek Reservoir on hydrochemistry and plankton of a large, lowland river (the Lower Vistula River, Poland). Ecol. Eng. 2010, 36, 1747–1753. [Google Scholar] [CrossRef]
- Hillbricht-Ilkowska, A. Shallow lakes in lowland river systems: Role in transport and transformations of nutrients and in biological diversity. In Shallow Lakes’ 98. Hydrobiologia 1999, 408–409, 349–358. [Google Scholar] [CrossRef]
- Carlson, R.E. A trophic state index for lakes. Limnol. Oceanogr. 1977, 22, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Czerniawski, R.; Kowalska-Góralska, M. Spatial changes in zooplankton communities in a strong human-mediated river ecosystem. PeerJ 2018, 6, e5087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cumming, G.S. The impact of low-head dams on fish species richness in Wisconsin, USA. Ecol. Appl. 2004, 14, 1495–1506. [Google Scholar] [CrossRef]
- Shurin, J.B.; Havel, J.E. Hydrologic connections and overland dispersal in an exotic freshwater crustacean. Biol. Invasions 2002, 4, 431–439. [Google Scholar] [CrossRef]
- Wu, N.; Tang, T.; Fu, X.; Jiang, W.; Li, F.; Zhou, S.; Cai, Q. Impacts of cascade run-of-river dams on benthic diatoms in the Xiangxi River, China. Aquat. Sci. 2010, 72, 117–125. [Google Scholar] [CrossRef]
- Zhou, S.; Tang, T.; Wu, N.; Fu, X.; Cai, Q. Impacts of a Small Dam on Riverine Zooplankton. Int. Rev. Hydrobiol. 2008, 93, 297–311. [Google Scholar] [CrossRef]
- Kuczyńska-Kippen, N.M.; Nagengast, B. The influence of the spatial structure of hydromacrophytes and differentiating habitat on the structure of rotifer and cladoceran communities. Hydrobiologia 2006, 559, 203–212. [Google Scholar] [CrossRef]
- Vadadi-Fülöp, C.; Hufnagel, L.; Jablonszky, G.; Zsuga, K. Crustacean plankton abundance in the Danube River and in its side arms in Hungary. Biologia 2009, 64, 1184–1195. [Google Scholar] [CrossRef] [Green Version]
- Viroux, L. Seasonal and longitudinal aspects of microcrustacean (Cladocera, Copepoda) dynamics in a lowland river. J. Plankton Res. 2002, 24, 281–292. [Google Scholar] [CrossRef]
- Ejsmont-Karabin, J.; Węgleńska, T. Changes in the zooplankton structure in the transitory river-lake-river zone. The River Krutynia system, Mazurian Lake District. Zesz. Nauk. Pol. Akad. Nauk. Kom. Nauk. PAN Człowiek i Środowisko 1996, 13, 263–289. (In Polish) [Google Scholar]
- Walks, D.J.; Cyr, H. Movement of plankton through lake-stream systems. Freshw. Biol. 2004, 49, 745–759. [Google Scholar] [CrossRef]
- Dembowska, E.A. The Use of Phytoplankton in the Assessment of Water Quality in the Lower Section of Poland’s Largest River. Water 2021, 13, 3471. [Google Scholar] [CrossRef]
- Shayestehfar, A.; Soleimani, M.; Mousavi, S.N.; Shirazi, F. Ecological study of rotifers from Kor river, Fars. Iran. J. Environ. Biol. 2008, 29, 715–720. [Google Scholar] [PubMed]
- Orcutt, J.D.; Pace, M.L. Seasonal dynamics of rotifer and crustacean zooplankton populations in a eutrophic, monomictic lake with a note on rotifer sampling techniques. Hydrobiologia 1984, 119, 73–80. [Google Scholar] [CrossRef]
- Vadadi-Fülöp, C.; Hufnagel, L. Climate change and plankton phenology in freshwater: Current trends and future commitments. J. Limnol. 2014, 73, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Hansson, L.A.; Nicolle, A.; Brodersen, J.; Romare, P.; Nilsson, P.A.; Brönmark, C.; Skov, C. Consequences of fish predation, migration, and juvenile ontogeny on zooplankton spring dynamics. Limnol. Oceanogr. 2007, 52, 696–706. [Google Scholar] [CrossRef]
- Edmondson, W.T. Reproductive Rate of Planktonic Rotifers as Related to Food and Temperature in Nature. Ecol. Monogr. 1965, 35, 61–111. [Google Scholar] [CrossRef]
- Jafari, N.; Nabavi, M.S.; Akhavan, M. Ecological investigation of zooplankton abundance in the river Haraz, northeast Iran: Impact of environmental variables. Arch. Biol. Sci. 2011, 63, 785–798. [Google Scholar] [CrossRef]
- Ning, N.S.; Nielsen, D.L.; Hillman, T.J.; Suter, P.J. The influence of planktivorous fish on zooplankton communities in riverine slackwaters. Freshw. Biol. 2010, 55, 360–374. [Google Scholar] [CrossRef]
- Baranyi, C.; Hein, T.; Holarek, C.; Keckeis, S.; Schiemer, F. Zooplankton biomass and community structure in a Danube River floodplain system: Effects of hydrology. Freshw. Biol. 2002, 47, 473–482. [Google Scholar] [CrossRef]
- Balkić, A.G.; Ternjej, I.; Špoljar, M. Hydrology driven changes in the rotifer trophic structure and implications for food web interactions. Ecohydrology 2018, 11, e1917. [Google Scholar] [CrossRef]
- Chaparro, G.; Fontanarrosa, M.S.; O’Farrell, I. Colonization and succession of zooplankton after a drought: Influence of hydrology and free-floating plant dynamics in a floodplain lake. Wetlands 2015, 36, 85–100. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bydgoszcz Canal | Noteć Canal | Brda River | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Site 1 | Site 2 | Site 3 | Site 4 | Mean-Range | Site 5 Mean | Range | Site 6 Mean | Range | |
| WT (°C) | 16.4 | 16.7 | 17.0 | 16.5 | 16.7 (7.6–23.0) | 16.6 | (7.6–25.8) | 17.0 | (9.2–23.4) |
| SD (m) | 1.05 | 1.29 | 1.26 | 0.84 | 1.11 (0.45–2.10) | 1.11 | (0.45–1.50) | 2.20 | (2.00–2.50) |
| EC (µS cm−1) | 623 | 674 | 586 | 391 | 569 (152–1115) | 672 | (525–1120) | 122 | (101–202) |
| DO (mg/L) | 8.3 | 9.5 | 9.4 | 10.0 | 9.3 (2.7–16.3) | 6.8 | (1.9–12.1) | 8.8 | (6.6–14.2) |
| pH | 7.4 | 7.5 | 7.6 | 7.7 | 7.6 (6.7–8.9) | 7.5 | (6.5–8.3) | 8.0 | (7.0–9.5) |
| chl-a (µg/L) | 13.46 | 10.44 | 12.42 | 8.39 | 11.18 (0.30 31.31) | 15.78 | (0.92–51.48) | 5.42 | (0–23.81) |
| NNO3− (mg/L) | 0.55 | 0.59 | 0.57 | 0.47 | 0.54 (0.15–1.41) | 0.73 | (0.28–1.67) | 0.31 | (0.21–0.49) |
| PPO42− (mg/L) | 0.16 | 0.10 | 0.15 | 0.28 | 0.14 (0.02–0.63) | 0.21 | (0.03–0.89) | 0.04 | (0.01–0.07) |
| Bydgoszcz Canal | Noteć Canal | Brda River | ||||
|---|---|---|---|---|---|---|
| Site 1 | Site 2 | Site 3 | Site 4 | Site 5 | Site 6 | |
| Rotifers | 39 | 45 | 40 | 45 | 32 | 38 |
| Crustaceans | 12 | 12 | 15 | 13 | 11 | 11 |
| Total | 51 | 57 | 55 | 58 | 43 | 49 |
| Dominant species and percent of domination | Keratella cochlearis * 62% | Anuraeopsis fissa * 33% | Keratella cochlearis * 61% | Keratella cochlearis * 29% | Keratella cochlearis * 60% | Keratella cochlearis * 26% |
| Keratella quadrata * 6% | Keratella cochlearis * 6% | Keratella quadrata * 7% | Keratella quadrata * 18% | Keratella quadrata * 14% | Keratella quadrata * 23% | |
| Polyarthra remata * 8% | Polyarthra dolichoptera * 21% | Polyarthra dolichoptera * 20% | Polyarthra dolichoptera * 23% | Polyarthra dolichoptera * 6% | Polyarthra dolichoptera * 35% | |
| Bosmina longirostris ** 17% | Bosmina longirostris ** 34% | Bosmina longirostris ** 7% | Bosmina longirostris ** 17% | Bosmina longirostris ** 8% | Polyarthra remata * 12% | |
| nauplius ** 7% | nauplius ** 6% | nauplius ** 5% | nauplius ** 13% | nauplius ** 12% | nauplius ** 4% | |
| Bydgoszcz Canal | Noteć Canal | Brda River | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Site 1 | Site 2 | Site 3 | Site 4 | Mean-Range | Site 5 Mean | Range | Site 6 Mean | Range | |
| Tax total | 16 | 17 | 17 | 21 | 18 (11–18) | 15 | (13–18) | 14 | (8–29) |
| Tax Rot | 11 | 13 | 13 | 16 | 13 (5–20) | 11 | (6–15) | 11 | (6–25) |
| Tax Crust | 5 | 4 | 4 | 5 | 4.5 (1–8) | 4 | (2–7) | 3 | (2–5) |
| N total | 574 | 332 | 662 | 744 | 578 (28–2478) | 196 | (25–640) | 246 | (16–1354) |
| N Rot | 429 | 247 | 577 | 496 | 437 (8–3430) | 161 | (15–620) | 229 | (14–1338) |
| N Crust | 150 | 88 | 86 | 249 | 143 (4–1420) | 35 | (10–78) | 17 | (2–38) |
| B total | 5.204 | 1.511 | 5.773 | 4.541 | 4.258 (0.113–26.285) | 0.634 | (0.108–1.661) | 0.494 | (0.026–1.349) |
| B Rot | 0.142 | 0.120 | 0.273 | 0.183 | 0.180 (0.004–0.971) | 0.066 | (0.003–0.243) | 0.161 | (0.027–0.882) |
| B Crust | 5.062 | 1.391 | 5.500 | 4.358 | 4.078 (0.046–26.259) | 0.568 | (0.099–1.656) | 0.333 | (0.023–0.383) |
| H’ index | 1.91 | 1.90 | 1.93 | 1.92 | 1.92 | 1.94 | 1.85 | ||
| J’ index | 0.57 | 0.50 | 0.49 | 0.71 | 0.57 | 0.48 | 0.32 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolarova, N.; Napiórkowski, P. How Do Specific Environmental Conditions in Canals Affect the Structure and Variability of the Zooplankton Community? Water 2022, 14, 979. https://doi.org/10.3390/w14060979
Kolarova N, Napiórkowski P. How Do Specific Environmental Conditions in Canals Affect the Structure and Variability of the Zooplankton Community? Water. 2022; 14(6):979. https://doi.org/10.3390/w14060979
Chicago/Turabian StyleKolarova, Nikola, and Paweł Napiórkowski. 2022. "How Do Specific Environmental Conditions in Canals Affect the Structure and Variability of the Zooplankton Community?" Water 14, no. 6: 979. https://doi.org/10.3390/w14060979