
Influence of Increased Freshwater Inflow on Nitrogen and Phosphorus Budgets in a Dynamic Subtropical Estuary, Barataria Basin, Louisiana



Abstract
:1. Introduction
2. Materials and Methods
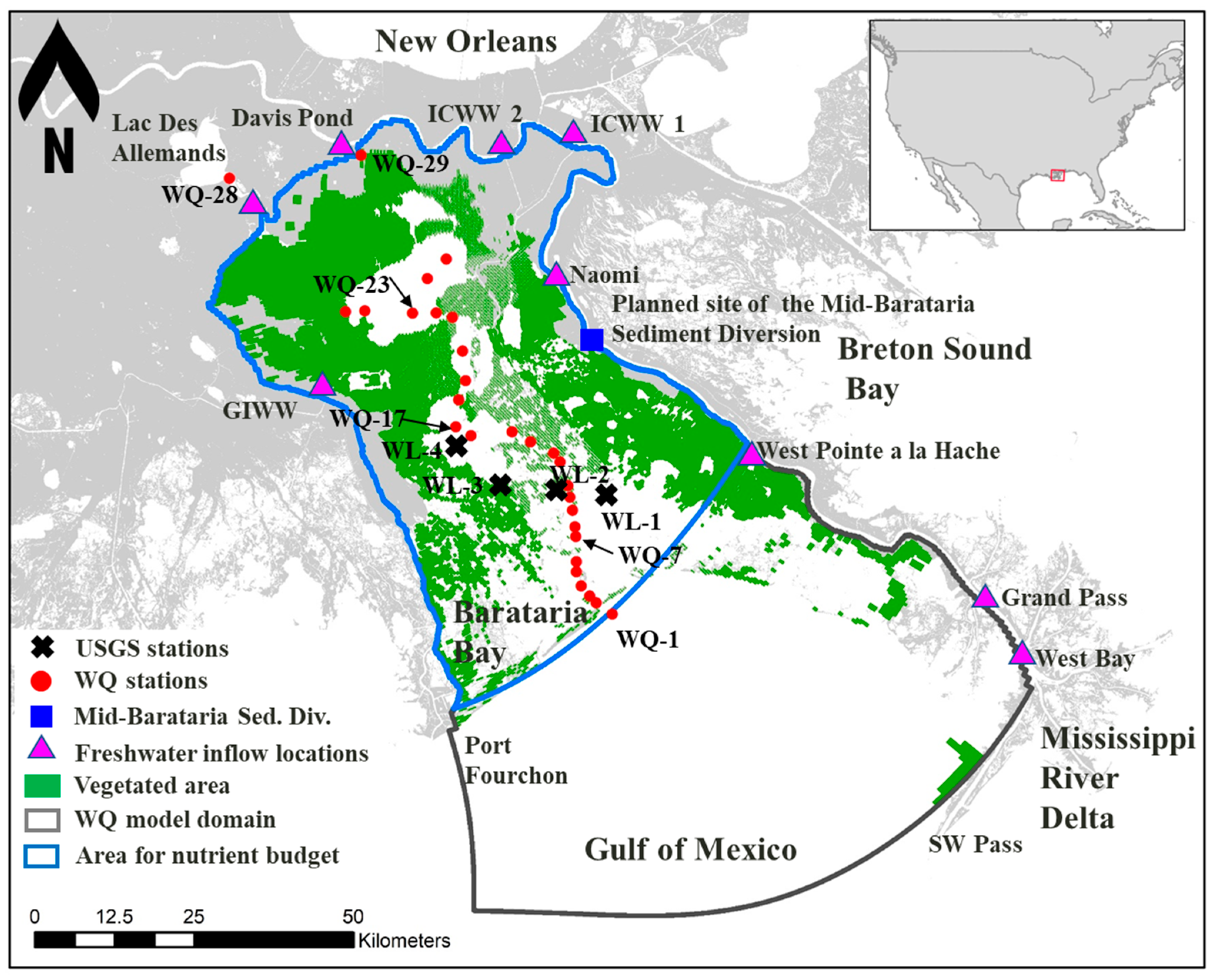
2.1. Study Area
2.2. Model
2.3. Current Freshwater Inflows
2.4. Water Quality Data
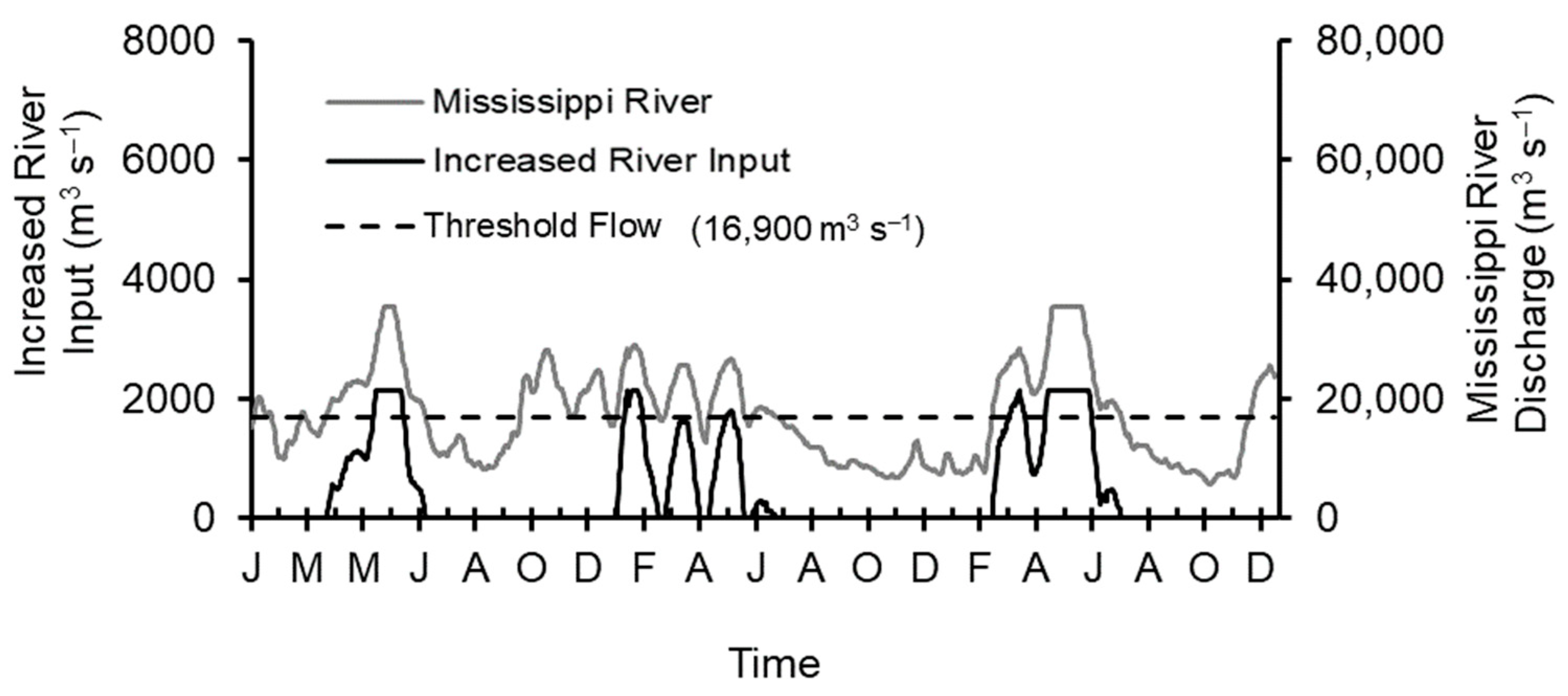
2.5. Increased River Input Scenario
2.6. Nutrient Budget Analysis
3. Results
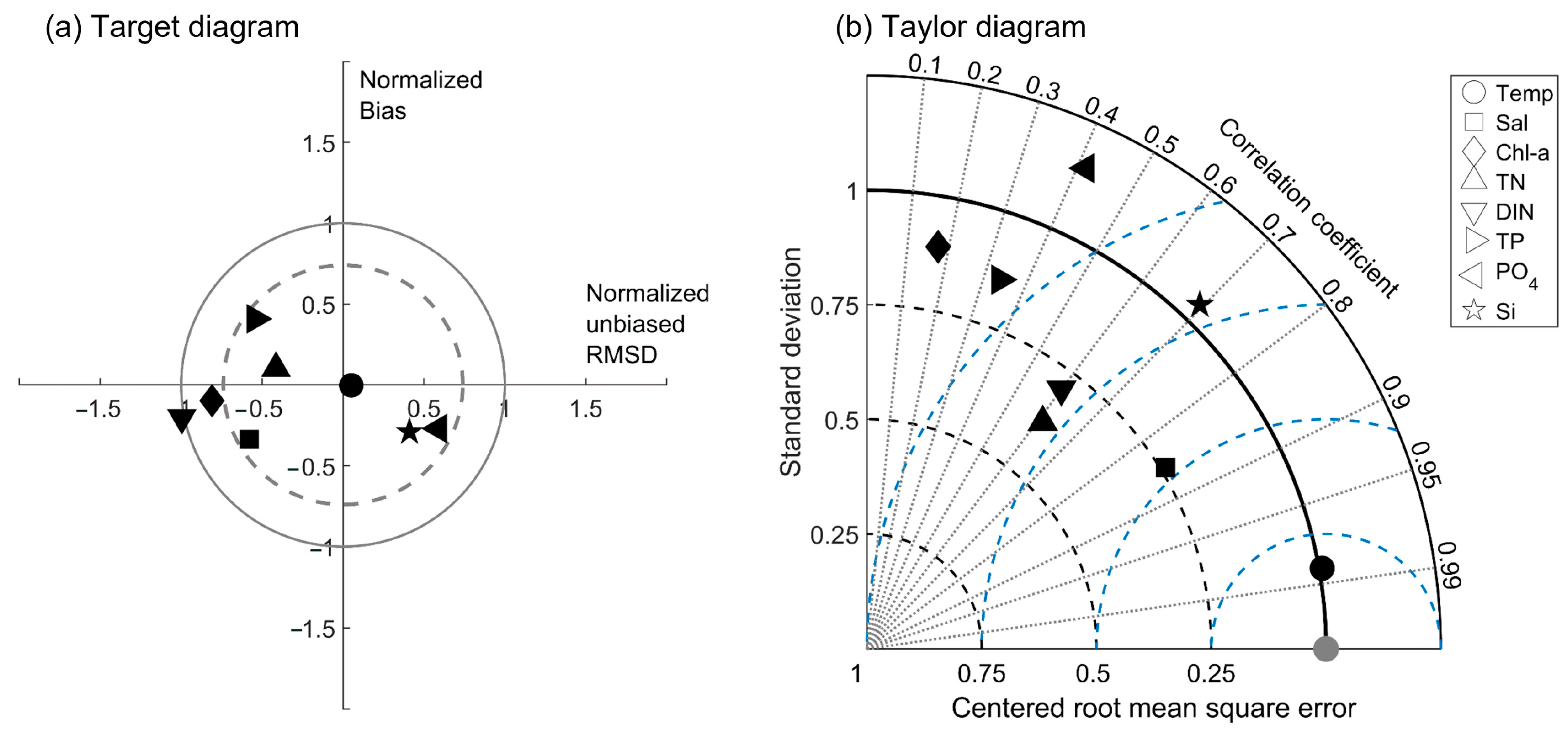
3.1. Model Performance
3.2. TN and TP Budgets with Current River Input
3.2.1. TN Budget
3.2.2. TP Budget
3.3. Interannual Water, TN, and TP Fluxes Variation with Current River Input
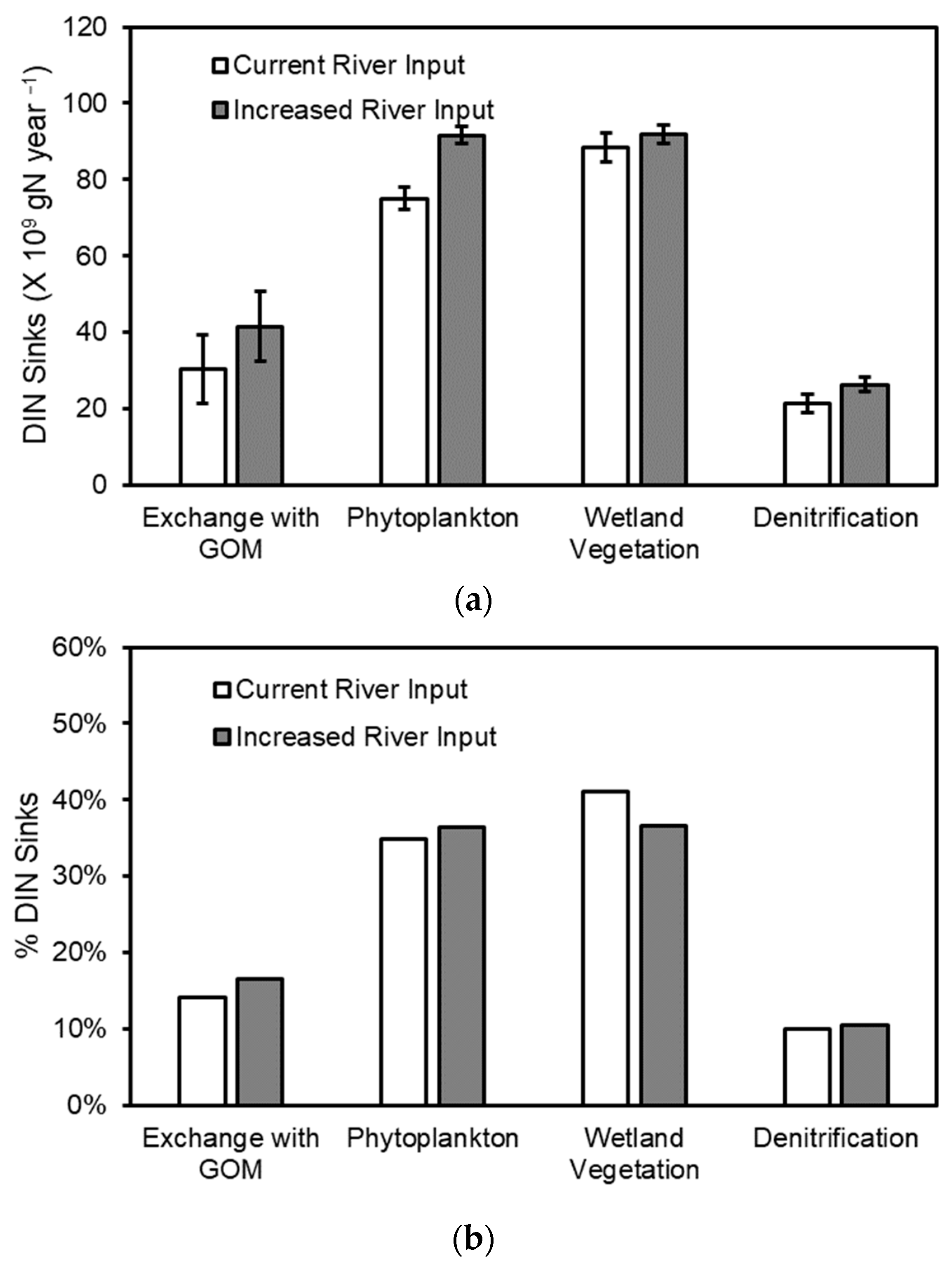
3.4. Effect of Increased River Input on Nutrient Pools and Fluxes within the Basin
4. Discussion
4.1. TN and TP Budgets with Current River Input
4.2. Interannual Water, TN, and TP Fluxes with Current River Input
4.3. Impact of Increased River Input on TN and TP Budgets
4.4. Comparing to Other Ecosystems
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Constituents |
|---|---|
| Phytoplankton | Freshwater diatoms (FDIATOMS), Freshwater flagellates (FFLGELA), Green algae (GREENS), Microcystis spp. (MICROSYSTIS), Anabaena spp. (ANABAENA), Marine diatoms (MDIATOM), Marine flagellates (MFLAGELA), dinoflagellates (DINOFLAG) |
| Emergent vegetation | Typha spp. (TYDO), Phragmites spp. (PHAU7), Spartina alterniflora (SPAL), Spartina patens (SPPA), Sagittaria lancifolia (SALA), Sagittaria latifolia (SALA2), Zizaniopsis miliacea (ZIMI) |
| Particulate organic matter | Carbon POC1-4 *, and Dissolved Organic Carbon (DOC); Nitrogen PON1-4 *, and DON; Phosphorus POP1-4 *, DOP; Sulfur POS1-4 *, DOS |
| Ammonium, nitrate | NH4, NO3 |
| Dissolved phosphate, adsorbed phosphate, vivianite-P, apatite-P | PO4, AAP, VIVP, APATP |
| Dissolved silicate, opal silicate | Si, OPAL |
| Dissolved oxygen | OXY |
| Sulfate, dissolved sulfide, particulate sulfide | SO4, SUD, SUP |
| Inorganic sediment types | IM1-3 |
| Salinity | Salinity |
| Coefficients | F. Diatoms | F. Flagellates | Greens | Micro-cystis | Anabaena | M. Diatoms | M. Flagellates | Dino- Flagellates | |
|---|---|---|---|---|---|---|---|---|---|
| Pmax at 0 °C | Orig. | 0.45 | 0.35 | 0.07 | 0.047 | 0.19 | 0.07 | 0.09 | 0.13 |
| Rev. | 0.09 | 0.02 | 0.14 | 0.06 | 0.05 | 0.06 | 0.10 | 0.09 | |
| Pmax temp. funct. | Orig. | Exp. | Linear | Linear | Linear | Exp. | Linear | Linear | Linear |
| Rev. | Linear | Linear | Linear | Linear | Linear | Linear | Linear | Linear | |
| Pmax temp. coeff. | Orig. | 1.06 | 0.0 | 3.0 | 5.0 | 1.09 | −4.5 | −1 | 4.8 |
| Rev. | 10.0 | −8.0 | 15.0 | 10.0 | 10.0 | −10.0 | 10.0 | 5.5 | |
| Resp at 0 °C | Orig. | 0.031 | 0.031 | 0.012 | 0.012 | 0.06 | 0.06 | 0.06 | 0.06 |
| Rev. | 0.038 | 0.04 | 0.034 | 0.030 | 0.042 | 0.038 | 0.038 | 0.04 | |
| Resp temp. coeff. | Orig. | 1.072 | 1.072 | 1.072 | 1.072 | 1.072 | 1.066 | 1.066 | 1.066 |
| Rev. | 1.060 | 1.070 | 1.070 | 1.070 | 1.080 | 1.090 | 1.070 | 1.066 | |
| Mort at 0 °C/35 ppt | Orig. | 0.42 | 0.42 | 0.42 | 0.54 | 0.54 | 0.07 | 0.07 | 0.075 |
| Rev. | 0.20 | 0.20 | 0.20 | 0.30 | 0.20 | 0.04 | 0.04 | 0.04 | |
| Mort at 0 ppt | Orig. | 0.035 | 0.035 | 0.035 | 0.08 | 0.08 | 0.42 | 0.42 | 0.42 |
| Rev. | 0.055 | 0.080 | 0.045 | 0.06 | 0.06 | 0.40 | 0.40 | 0.40 | |
| Mort temp. coeff. | Orig. | 1.080 | 1.080 | 1.080 | 1.080 | 1.080 | 1.072 | 1.072 | 1.072 |
| Rev. | 1.080 | 1.070 | 1.080 | 1.080 | 1.075 | 1.080 | 1.080 | 1.080 | |
| Mort stress shape B1 | Orig. | 0.002 | 0.002 | 0.002 | 0.001 | 0.001 | 0.002 | 0.002 | 0.003 |
| Rev. | 0.0008 | 0.0008 | 0.0006 | 0.0006 | 0.0006 | 0.001 | 0.0015 | 0.0015 | |
| Mort stress shape B2 | Orig. | 8000 | 8000 | 11,500 | 6000 | 6000 | 6000 | 6000 | 6000 |
| Rev. | 7000 | 8000 | 10,000 | 10,000 | 11,000 | 4000 | 5000 | 6000 | |
| Process | Coefficients | Water Column | ||||
|---|---|---|---|---|---|---|
| Original | Revised | |||||
| Vegetated | Un-Vegetated | Vegetated | Un-Vegetated | |||
| Organic matter | b_poc2doc | Fraction POC2 converted to DOC (-) | 0.10 | 0.10 | 0.05 | 0.05 |
| b_poc3doc | Fraction POC3 converted to DOC (-) | 0.10 | 0.10 | 0.03 | 0.03 | |
| Settling, other than phytoplankton | V0SedPOC | Settling velocity for POC 1 to 4 (m/day) | 1.0 | 0.5 | 1.0 | 0.15 |
| V0SedIM1 | Settling velocity for silt (m/day) | 0.15 | 0.025 | 0.3 | 0.15 | |
| V0SedIM2 | Settling velocity for clay (m/day) | 0.05 | 0.005 | 0.1 | 0.05 | |
| V0SedIM3 | Settling velocity for sand (m/day) | 3.0 | 3.0 | 2.0 | 1.0 | |
Appendix B




| Water Quality Variable | Mean Obs. | Mean Model | Bias (B) | Correlation Coefficient (R) | Root Mean Square Error (RMSE) |
|---|---|---|---|---|---|
| Salinity | 6.95 | 4.61 | −2.34 | 0.85 | 4.71 |
| Temperature (°C) | 22.7 | 22.7 | −0.05 | 0.98 | 1.16 |
| Chl-a (ug L−1) | 16.3 | 14.7 | −1.60 | 0.17 | 13.3 |
| TN (mg L−1) | 0.60 | 0.66 | 0.06 | 0.61 | 0.25 |
| DIN (mg L−1) | 0.22 | 0.17 | −0.04 | 0.60 | 0.22 |
| TP (mg L−1) | 0.05 | 0.07 | 0.02 | 0.34 | 0.03 |
| PO4 (mg L−1) | 0.04 | 0.03 | −0.01 | 0.41 | 0.03 |
| Si (mg L−1) | 1.84 | 1.32 | −0.53 | 0.69 | 0.92 |
References
- Couvillion, B.R.; Beck, H.; Schoolmaster, D.; Fischer, M. Land Area Change in Coastal Louisiana from 1932 to 2016. U.S. Geological Survey Scientific Investigations Map 3381; U.S. Geological Survey: Reston, VA, USA, 2017.
- Barras, J.A.; Beville, S.; Britsch, D.; Hartley, S.; Hawes, S.R.; Johnston, J.; Kemp, P.; Kinler, Q.; Martucci, A.; Porthouse, J.; et al. Historical and Projected Coastal Louisiana Land Changes: 1978–2050; U.S. Geological Survey Open File Report 03–334; U.S. Geological Survey: Reston, VA, USA, 2003.
- Tornqvist, T.E.; Wallace, D.J.; Storms, J.E.A.; Wallinga, J.; van Dam, R.L.; Blaauw, M.; Derksen, M.S.; Klerks, C.J.W.; Meijneken, C.; Snijders, E.M.A. Mississippi Delta subsidence primarily caused by compaction of Holocene strata. Nat. Geosci. 2008, 1, 173–176. [Google Scholar] [CrossRef]
- Blum, M.D.; Roberts, H.H. Drowning of the Mississippi Delta due to insufficient sediment supply and global sea-level rise. Nat. Geosci. 2009, 2, 488–491. [Google Scholar] [CrossRef]
- Turner, R.E. Intertidal vegetation and commercial yields of penaeid shrimp. Trans. Am. Fish. Soc. 1977, 106, 411–416. [Google Scholar] [CrossRef]
- Day, R.H.; Williams, T.M.; Swarzenski, C.M. Hydrology of tidal freshwater forested wetlands of the southeastern United States. In Ecology of Tidal Freshwater Forested Wetlands of the Southeastern United States; Springer: Berlin/Heidelberg, Germany, 2007; pp. 29–63. [Google Scholar]
- Coastal Protection and Restoration Authority. Coastal Protection and Restoration Authority: Strategic Plan Fiscal Year 2017–2018 through Fiscal Year 2021–2022; Strategic fiscal plan; Coastal Protection and Restoration Authority: Baton Rouge, LA, USA, 2017.
- Twilley, R.R.; Rivera-Monroy, V. Sediment and Nutrient Tradeoffs in Restoring Mississippi River Delta: Restoration vs Eutrophication. J. Contemp. Water Res. Educ. 2009, 141, 39–44. [Google Scholar] [CrossRef]
- Bargu, S.; Justic, D.; White, J.R.; Lane, R.; Day, J.; Paerl, H.; Raynie, R. Mississippi River diversions and phytoplankton dynamics in deltaic Gulf of Mexico estuaries: A review. Estuarine Coast. Shelf Sci. 2019, 221, 39–52. [Google Scholar] [CrossRef]
- Turner, R.E.; Rabalais, N.N. Changes in Mississippi River Water Quality This Century. Bioscience 1991, 41, 140–147. [Google Scholar] [CrossRef]
- Nixon, S.W.; Ammerman, J.W.; Atkinson, L.P.; Berounsky, V.M.; Billen, G.; Boicourt, W.C.; Boynton, W.R.; Church, T.M.; DiToro, D.M.; Elmgren, R.; et al. The fate of nitrogen and phosphorus at the land-sea margin of the North Atlantic Ocean. Biogeochemistry 1996, 35, 141–180. [Google Scholar] [CrossRef]
- Rabalais, N.N.; Turner, R.E.; Scavia, D. Beyond Science into Policy: Gulf of Mexico Hypoxia and the Mississippi River. Bioscience 2002, 52, 129–142. [Google Scholar] [CrossRef]
- Tao, B.; Tian, H.; Ren, W.; Yang, J.; Yang, Q.; He, R.; Cai, W.; Lohrenz, S. Increasing Mississippi river discharge throughout the 21st century influenced by changes in climate, land use, and atmospheric CO2. Geophys. Res. Lett. 2014, 41, 4978–4986. [Google Scholar] [CrossRef]
- Sinha, E.; Michalak, A.M.; Balaji, V. Eutrophication will increase during the 21st century as a result of precipitation changes. Science 2017, 357, 405. [Google Scholar] [CrossRef]
- Baustian, M.M.; Clark, F.R.; Jerabek, A.S.; Wang, Y.; Bienn, H.C.; White, E.D. Modeling current and future freshwater inflow needs of a subtropical estuary to manage and maintain forested wetland ecological conditions. Ecol. Indic. 2018, 85, 791–807. [Google Scholar] [CrossRef]
- Kingsford, R.T. Ecological impacts of dams, water diversions and river management on floodplain wetlands in Australia. Austral. Ecol. 2000, 25, 19. [Google Scholar] [CrossRef]
- Kingsford, R.T. Conservation management of rivers and wetlands under climate change—A synthesis. Mar. Freshw. Res. 2011, 62, 217–222. [Google Scholar] [CrossRef]
- D’Elia, C.F.; Boynton, W.R.; Sanders, J.G. A watershed perspective on nutrient enrichment, science, and policy in the Patuxent River, Maryland: 1960–2000. Estuaries 2003, 26, 171–185. [Google Scholar] [CrossRef]
- Boynton, W.; Garber, J.H.; Summers, R.; Kemp, W.M. Inputs, Transformations, and Transport of Nitrogen and Phosphorus in Chesapeake Bay and Selected Tributaries. Estuaries 1995, 18, 285–314. [Google Scholar] [CrossRef]
- Nixon, S.W. Prehistoric Nutrient Inputs and Productivity in Narragansett Bay. Estuaries 1997, 20, 253. [Google Scholar] [CrossRef]
- Williams, M.R.; Fisher, T.R.; Boynton, W.R.; Cerco, C.F.; Kemp, M.W.; Eshleman, K.N.; Kim, S.; Hood, R.; Fiscus, D.A.; Radcliffe, G.R. An integrated modelling system for management of the Patuxent River estuary and basin, Maryland, USA. Int. J. Remote Sens. 2006, 27, 3705–3726. [Google Scholar] [CrossRef]
- Laurent, A.; Fennel, K. Simulated reduction of hypoxia in the northern Gulf of Mexico due to phosphorus limitation. Elementa: Sci. Anthr. 2013, 2, 22. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Q.; Hu, K.; La Peyre, M.K. A Modeling Study of the Impacts of Mississippi River Diversion and Sea-Level Rise on Water Quality of a Deltaic Estuary. Estuaries Coasts 2017, 40, 1028–1054. [Google Scholar] [CrossRef]
- Laurent, A.; Fennel, K.; Ko, D.S.; Lehrter, J. Climate Change Projected to Exacerbate Impacts of Coastal Eutrophication in the Northern Gulf of Mexico. J. Geophys. Res. Oceans 2018, 123, 3408–3426. [Google Scholar] [CrossRef]
- Smits, J.G.C.; Van Beek, J.K.L. ECO: A Generic Eutrophication Model Including Comprehensive Sediment-Water Interaction. Edited by Vishal Shah. PLoS ONE 2013, 8, e68104. [Google Scholar] [CrossRef] [PubMed]
- Swenson, E.M.; Cable, J.E.; Fry, B.; Justic, D.; Das, A.; Snedden, G.; Swarzenski, C. Estuarine flushing times influenced by freshwater diversions. Coast. Hydrol. Process. 2006, 33, 403–412. [Google Scholar]
- Das, A.; Justić, D.; Swenson, E. Modeling estuarine-shelf exchanges in a deltaic estuary: Implications for coastal carbon budgets and hypoxia. Ecol. Model. 2010, 221, 978–985. [Google Scholar] [CrossRef]
- Das, A.; Justic, D.; Swenson, E.; Turner, R.E.; Inoue, M.; Park, D. Coastal land loss and hypoxia: The ‘outwelling’ hypothesis revisited. Environ. Res. Lett. 2011, 6, 025001. [Google Scholar] [CrossRef]
- Fitzgerald, D.M.; Kulp, M.; Penland, S.; Flocks, J.; Kindinger, J. Morphologic and stratigraphic evolution of muddy ebb-tidal deltas along a subsiding coast: Barataria Bay, Mississippi River delta. Sedimentology 2004, 51, 1157–1178. [Google Scholar] [CrossRef]
- Penland, S.; Ramsey, K.E.; McBride, R.A.; Mestayer, J.T.; Westphal, K.A. Relative Sea Level Rise and Delta-Plain Development in the Terrebonne Parish Region; Coastal Geology Technical Report No. 4; Louisiana Geological Survey: Baton Rouge, LA, USA, 1988. [Google Scholar]
- Turner, R.E.; Swenson, E.M.; Milan, C.S.; Lee, J.M. Spatial variations in Chlorophyll a, C, N, and P in a Louisiana estuary from 1994 to 2016. Hydrobiologia 2019, 834, 131–144. [Google Scholar] [CrossRef]
- Turner, R.E. Water quality data from the Barataria Basin, 1994–2016. In Distributed by: Gulf of Mexico Research Initiative Information and Data Cooperative (GRIIDC); Harte Research Institute, Texas A&M University–Corpus Christi: Corpus Christi, TX, USA, 2017. [Google Scholar] [CrossRef]
- Gagliano, S.; Culley, P.; Earle, D., Jr.; King, P.; Latiolais, C.; Light, P.; Rowland, A.; Shlemon, R.; van Beek, J.L. Environmental Atlas and Mulituse Management Plan for South-Central Louisiana; Department of the Army, New Orleans District Corps of Engineers, Office of Sea Grant, National Oceanic and Atmospheric Administration: Washington, DC, USA, 1973.
- Wiseman, W.J., Jr.; Swenson, E.M.; Power, J.; Swenson, E.M.; Power, J. Salinity Trends in Louisiana Estuaries. Estuaries 1990, 13, 265–271. [Google Scholar] [CrossRef]
- Habib, E.; Larson, B.F.; Nuttle, W.K.; Rivera-Monroy, V.H.; Nelson, B.R.; Meselhe, E.A.; Twilley, R.R. Effect of rainfall spatial variability and sampling on salinity prediction in an estuarine system. J. Hydrol. 2008, 350, 56–67. [Google Scholar] [CrossRef]
- Wiseman, W.J., Jr.; Swenson, E.M. Modeling the effects of produced water discharges on estuarine salinity. In Environmental Impacts of Produced Water Discharges in Coastal Louisiana; Boesch, D.F., Rabalais, N.N., Eds.; Louisiana Universities Marine Consortium: Chauvin, Louisiana, 1989. [Google Scholar]
- Orlando, S.P.J.; Rozas, L.P.; Ward, G.H.; Klien, C.J. Salinity Characteristics of Gulf of Mexico Estuaries; National Oceanic and Atmospheric Administration, Office of Ocean Resources Conservation and Assessment: Silver Spring, MD, USA, 1993.
- McKee, K.L.; Mendelssohn, I.A.; Materne, M.D. Acute salt marsh dieback in the Mississippi River Deltaic Plain: A drought-induced phenomenon? Glob. Ecol. Biogeogr. 2004, 13, 65–73. [Google Scholar] [CrossRef]
- Deltares. Delft3D-FLOW User Manual; Deltares: Delft, The Netherlands, 2014. [Google Scholar]
- Hu, K.; Chen, Q.; Wang, H.; Hartig, E.K.; Orton, P.M. Numerical modeling of salt marsh morphological change induced by Hurricane Sandy. Coast. Eng. 2018, 132, 63–81. [Google Scholar] [CrossRef]
- Tehranirad, B.; Herdman, L.; Nederhoff, K.; Erikson, L.; Cifelli, R.; Pratt, G.; Leon, M.; Barnard, P. Effect of Fluvial Discharges and Remote Non-Tidal Residuals on Compound Flood Forecasting in San Francisco Bay. Water 2020, 12, 2481. [Google Scholar] [CrossRef]
- Smits, J. Delft3D-ECO: Model Documentation; Deltares: Delft, The Netherlands, 2013. [Google Scholar]
- Deltares. D-Water Quality: Versatile Water Quality Modelling in 1D, 2D or 3D Systems including Physical, (Bio)Chemical and Biological Processes; User Manual Version 4.99.34158; Deltares: Delft, The Netherlands, 2014. [Google Scholar]
- Baustian, M.M.; Meselhe, E.; Jung, H.; Sadid, K.; Duke-Sylvester, S.M.; Visser, J.M.; Allison, M.A.; Moss, L.C.; Ramatchandirane, C.; van Maren, D.S.; et al. Development of an Integrated Biophysical Model to represent morphological and ecological processes in a changing deltaic and coastal ecosystem. Environ. Model. Softw. 2018, 109, 402–419. [Google Scholar] [CrossRef]
- Visser, J.M.; Duke-Sylvester, S.; Broussard, W.; Carter, J. Vegetation Model Technical Report (Appendix D-4); Techinal Report; CPRA: Baton Rouge, LA, USA, 2012. [Google Scholar]
- Brown, S.; Couvillion, B.R.; Dong, Z.; Meselhe, E.; Visser, J.M.; Wang, Y.; White, E.D. 2017 Coastal Master Plan: Attachment C3-23: ICM Calibration, Validation, and Performance Assessment; Version Final. 2017 Coastal Master Plan; Coastal Protection and Restoration Authority: Baton Rouge, LA, USA, 2017.
- Allison, M.A.; Demas, C.R.; Ebersole, B.A.; Kleiss, B.A.; Little, C.D.; Meselhe, E.A.; Powell, N.J.; Pratt, T.C.; Vosburg, B.M. A water and sediment budget for the lower Mississippi–Atchafalaya River in flood years 2008–2010: Implications for sediment discharge to the oceans and coastal restoration in Louisiana. J. Hydrol. 2012, 432–433, 84–97. [Google Scholar] [CrossRef]
- U.S. Geological Survey. USGS Water Data for the Nation: U.S. Geological Survey National Water Information System Database; U.S. Geological Survey: Reston, VA, USA, 2016. [CrossRef]
- Brown, S. 2017 Coastal Master Plan: Attachment C3-26: Hydrology and Water Quality Boundary Conditions; Version Final. Louisiana’s Comprehensive Master Plan for a Sustainable Coast; Coastal Master Plan: Baton Rouge, LA, USA, 2017.
- McCorquodale, J.A.; Roblin, R.J.; Georgiou, I.Y.; Haralampides, K.A. Salinity, Nutrient, and Sediment Dynamics in the Pontchartrain Estuary. J. Coast. Res. 2009, 10054, 71–87. [Google Scholar] [CrossRef]
- National Atmospheric Deposition Program. NADP/NTN Monitoring Location LA30. NADP (National Atmospheric Deposition Program). 2002. Available online: http://nadp.sws.uiuc.edu/ (accessed on 4 May 2015).
- Meselhe, E.A.; Sadid, K.; Xing, F.; Jung, H.; Baustian, M.M.; Smits, J.G.C.; Maren, B.D.S.; Duke-Sylvester, S.M.; Visser, J.M. Mississippi River Hydrodynamics and Delta Management Study (MRHDMS): Assessment and Analysis of Alternatives; Final Report Task Order 36; The Water Institute of the Gulf. Funded by the Coastal Protection and Restoration Authority: Baton Rouge, LA, USA, 2016. [Google Scholar]
- Taylor, K.E. Summarizing multiple aspects of model performance in a single diagram. J. Geophys. Res. Atmos. 2001, 106, 7183–7192. [Google Scholar] [CrossRef]
- Jolliff, J.K.; Kindle, J.C.; Shulman, I.; Penta, B.; Friedrichs, M.A.; Helber, R.; Arnone, R.A. Summary diagrams for coupled hydrodynamic-ecosystem model skill assessment. J. Mar. Syst. 2009, 76, 64–82. [Google Scholar] [CrossRef]
- Los, F.; Blaas, M. Complexity, accuracy and practical applicability of different biogeochemical model versions. J. Mar. Syst. 2010, 81, 44–74. [Google Scholar] [CrossRef]
- Vaccare, J.; Meselhe, E.; White, J.R. The denitrification potential of eroding wetlands in Barataria Bay, LA, USA: Implications for river reconnection. Sci. Total. Environ. 2019, 686, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Dettmann, E.H. Effect of Water Residence Time on Annual Export and Denitrification of Nitrogen in Estuaries: A Model Analysis. Estuaries 2001, 24, 481. [Google Scholar] [CrossRef]
- Peierls, B.L.; Hall, N.S.; Paerl, H.W. Non-monotonic Responses of Phytoplankton Biomass Accumulation to Hydrologic Variability: A Comparison of Two Coastal Plain North Carolina Estuaries. Estuaries Coasts 2012, 35, 1376–1392. [Google Scholar] [CrossRef]
- Valdes-Weaver, L.M.; Piehler, M.F.; Pinckney, J.L.; Howe, K.E.; Rossignol, K.; Paerl, H.W. Long-term temporal and spatial trends in phytoplankton biomass and class-level taxonomic composition in the hydrologically variable Neuse-Pamlico estuarine continuum, North Carolina, U.S.A. Limnol. Oceanogr. 2006, 51, 1410–1420. [Google Scholar] [CrossRef]
- Boynton, W.R.; Murray, L.; Hagy, J.D.; Stokes, C.J.; Kemp, W.M. A comparative analysis of eutrophication patterns in a temperate coastal lagoon. Estuaries 1996, 19, 408–421. [Google Scholar] [CrossRef]
- Smith, S.V. Responses of Kaneohe Bay, Hawaii, to relaxation of sewage stress. In Estuaries and Nutrients; Neilson, B.J., Cronin, L.E., Eds.; Humana: Clifton, NJ, USA, 1981; pp. 391–410. [Google Scholar]
- Yurkovskis, A.; Wulff, F.; Rahm, L.; Andruzaitis, A.; Rodrigues-Medina, M. A nutrient budget of Gulf of Riga; Baltic Sea. Estuar. Coast. Shelf Sci. 1993, 37, 113–127. [Google Scholar] [CrossRef]
- Nixon, S.W.; Oviatt, C.A.; Frithsen, J.; Sullivan, B. Nutrients and the productivity of estuarine and coastal marine systems. J. Limnol. Soc. South Afr. 1986, 121, 43–71. [Google Scholar]
- Engqvist, A. Long-term nutrient balances in the eutrophication of the Himmerfjarden estuary. Estuar. Coast. Shelf Sci. 1996, 42, 483–507. [Google Scholar] [CrossRef]
- Valiela, I.; Costa, J.E. Eutrophication of Buttermilk Bay, a Cape Cod coastal embayment: Concentrations of nutrients and watershed nutrient budgets. Environ. Manag. 1988, 124, 539–553. [Google Scholar] [CrossRef]
- Eyre, B.D.; McKee, L.J. Carbon, nitrogen, and phosphorus budgets for a shallow subtropical coastal embayment (Moreton Bay, Australia). Limnol. Oceanogr. 2002, 474, 1043–1055. [Google Scholar] [CrossRef]
- Degobbis, D.; Gilmartin, M. Nitrogen, phosphorus, and biogenic silicon budgets for the northern Adriatic Sea. Oceanol. Acta 1990, 13, 31–45. [Google Scholar]
- National Oceanographic and Atmospheric Administration/Environmental Protection Agency (NOAA/EPA). Strategic Assessment of Near Coastal Waters, Susceptibility of East Coast Estuaries to Nutrient Discharges: Passamaquoddy Bay to Chesapeake Bay; Strategic Assessment Branch, NOS/NOAA: Rockville, MD, USA, 1989.
- Nixon, S.W.; Granger, S.L.; Nowicki, B.L. An assessment of the annual mass balance of carbon, nitrogen and phosphorus in Narragansett Bay. Biogeochemistry 1995, 31, 15–61. [Google Scholar] [CrossRef]
- Hager, S.W.; Schemel, L.E. Sources of nitrogen and phosphorus to Northern San Francisco Bay. Estuaries 1992, 15, 40–52. [Google Scholar] [CrossRef]
- Mortazavi, B.; Iverson, R.L.; Huang, W.; Lewis, F.G.; Caffrey, J. Nitrogen budget of Apalachicola Bay, a bar-built estuary in the northeastern Gulf of Mexico. Mar. Ecol. Process Ser. 2000, 195, 1–14. [Google Scholar] [CrossRef]
- Stammerjohn, S.E.; Smith, E.; Boynton, W.R.; Kemp, W.M. Potential Impacts from Marinas and Boats in Baltimore Harbor; Chesapeake Research Consortium Publication Number 139: Chesapeake Research Consortium: Solomons, MD, USA, 1991. [Google Scholar]
- Boynton, W.R.; Burger, N.H.; Stankelis, R.M.; Rohland, F.M.; Hagy, J.D., III; Frank, J.M.; Matteson, L.L.; Weir, M.M. An Environmental Evaluation of Back River with Selected Data from Patapsco River; Ref. No. [UMCES]CBL 98-112b; Chesapeake Biological Laboratory: Solomons, MD, USA, 1998; 90p. [Google Scholar]
- Day, J.W.; Conner, W.H.; DeLaune, R.D.; Hopkinson, C.S.; Hunter, R.G.; Shaffer, G.P.; Kandalepas, D.; Keim, R.F.; Kemp, G.P.; Lane, R.R.; et al. A Review of 50 Years of Study of Hydrology, Wetland Dynamics, Aquatic Metabolism, Water Quality and Trophic Status, and Nutrient Biogeochemistry in the Barataria Basin, Mississippi Delta—System Functioning, Human Impacts and Restoration Approaches. Water 2021, 13, 642. [Google Scholar] [CrossRef]




| Freshwater Inflows (×109 m3 year−1) | Years | ||
|---|---|---|---|
| 2009 | 2010 | 2011 | |
| Davis Pond Freshwater Diversion | 3.54 | 0.36 | 0.09 |
| Intracoastal Waterway (ICWW) 1 and 2 | 0.10 | 0.10 | 0.10 |
| Lac des Allemands | 1.41 | 1.41 | 1.41 |
| Naomi Siphon | 0.11 | 0.36 | 0.09 |
| Gulf Intracoastal Waterway (GIWW) | 1.84 | 1.84 | 1.84 |
| West Pointe a la Hache Siphon | 0.40 | 0.65 | 0.28 |
| Net precipitation | 0.71 | 0.74 | −0.60 |
| All in Barataria Basin | 8.11 | 5.46 | 3.21 |
| All in Barataria Basin with Increased River Input | 11.4 | 13.2 | 16.9 |
| Grand Pass | 54.4 | 49.9 | 51.5 |
| West Bay | 34.2 | 31.0 | 32.2 |
| Water Column Pool | Gains (×109 g N year−1) | Losses (×109 g N year−1) | Net (×109 g N year−1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Current River Input | Increased River Input | Current River Input | Increased River Input | Current River Input | Increased River Input | |||||||
| TN | DIN | TN | DIN | TN | DIN | TN | DIN | TN | DIN | TN | DIN | |
| Water inflows | 8.8 | 5.7 | 35.9 | 25.5 | 0.0 | 0.0 | 0.0 | 0.0 | 8.8 | 5.7 | 35.9 | 25.5 |
| Exchange with GOM | 96.6 | 37.8 | 99.7 | 39.5 | 97.1 | 30.5 | 120.7 | 41.6 | −0.5 | 7.3 | −20.9 | −2.1 |
| Sediment/Soils | 101.6 | 51.8 | 118.6 | 66.2 | 144.3 | 22.8 | 169.3 | 36.5 | −42.8 | 29.0 | −50.7 | 29.7 |
| Atmosphere | 0.6 | 0.6 | 0.6 | 0.6 | 0.0 | 0.0 | 0.0 | 0.0 | 0.6 | 0.6 | 0.6 | 0.6 |
| Internal process | 15.0 | 15.0 | 17.5 | 17.5 | 0.0 | 0.0 | 0.0 | 0.0 | 15.0 | 15.0 | 17.5 | 17.5 |
| Phytoplankton | 132.4 | 17.0 | 159.9 | 20.2 | 132.3 | 75.0 | 159.8 | 91.7 | 0.1 | −58.0 | 0.1 | −71.5 |
| Wetland vegetation | 33.7 | 0.0 | 35.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 33.7 | 0.0 | 35.1 | 0.0 |
| Denitrification | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | −0.0 | −0.0 | −0.0 | −0.0 |
| Sediment/Soils Pool | Gains (×109 g N year−1) | Losses (×109 g N year−1) | Net (×109 g N year−1) | |||||||||
| Current River Input | Increased River Input | Current River Input | Increased River Input | Current River Input | Increased River Input | |||||||
| TN | DIN | TN | DIN | TN | DIN | TN | DIN | TN | DIN | TN | DIN | |
| Water column | 144.3 | 22.8 | 169.3 | 36.5 | 101.6 | 51.8 | 118.6 | 66.2 | 42.8 | −29.0 | 50.7 | −29.7 |
| Atmosphere | 2.5 | 2.5 | 2.5 | 2.5 | 0.0 | 0.0 | 0.0 | 0.0 | 2.5 | 2.5 | 2.5 | 2.5 |
| Wetland vegetation | 54.6 | 0.0 | 56.8 | 0.0 | 88.3 | 88.3 | 91.9 | 91.9 | −33.7 | −88.3 | −35.1 | −91.9 |
| Internal process | 133.7 | 133.7 | 143.3 | 143.3 | 0.0 | 0.0 | 0.0 | 0.0 | 133.7 | 133.7 | 143.3 | 143.3 |
| Denitrification | 0.0 | 0.0 | 0.0 | 0.0 | 21.4 | 21.4 | 26.4 | 26.4 | −21.4 | −21.4 | −26.4 | −26.4 |
| Seepage/Burial | 9.9 | 2.5 | 8.3 | 2.2 | 0 | 0 | 0 | 0 | 9.9 | 2.5 | 8.3 | 2.2 |
| Water Column Pool | Gains (×109 g P year−1) | Losses (×109 g P year−1) | Net (×109 g P year−1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Current River Input | Increased River Input | Current River Input | Increased River Input | Current River Input | Increased River Input | |||||||
| TP | PO4 | TP | PO4 | TP | PO4 | TP | PO4 | TP | PO4 | TP | PO4 | |
| Water inflows | 0.9 | 0.4 | 2.9 | 1.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.9 | 0.4 | 2.9 | 1.3 |
| Exchange with GOM | 9.4 | 3.8 | 9.6 | 3.7 | 9.7 | 3.4 | 11.3 | 3.7 | −0.3 | 0.4 | −1.7 | 0.0 |
| Sediment/Soils | 12.3 | 8.5 | 13.0 | 8.8 | 14.1 | 4.7 | 15.5 | 4.9 | −1.9 | 3.8 | −2.5 | 3.9 |
| Atmosphere | 0.1 | 0.1 | 0.1 | 0.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.1 | 0.1 | 0.1 | 0.1 |
| Phytoplankton | 13.1 | 1.7 | 14.5 | 1.8 | 13.1 | 7.5 | 14.1 | 8.4 | 0.0 | −5.8 | 0.4 | −6.6 |
| Internal process | 0.0 | 1.2 | 0.0 | 1.3 | 0.0 | 0.1 | 0.0 | 0.1 | 0.0 | 1.1 | 0.0 | 1.2 |
| Wetland vegetation | 1.2 | 0.0 | 1.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 1.2 | 0.0 | 1.2 | 0.0 |
| Sediment/Soils Pool | Gains (×109 g P year−1) | Losses (×109 g P year−1) | Net (×109 g P year−1) | |||||||||
| Current River Input | Increased River Input | Current River Input | Increased River Input | Current River Input | Increased River Input | |||||||
| TP | PO4 | TP | PO4 | TP | PO4 | TP | PO4 | TP | PO4 | TP | PO4 | |
| Water | 14.1 | 4.7 | 15.5 | 4.9 | 12.3 | 8.5 | 13.0 | 8.8 | 1.9 | −3.8 | 2.5 | −3.9 |
| Atmosphere | 0.2 | 0.2 | 0.2 | 0.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.2 | 0.2 | 0.2 | 0.2 |
| Internal process | 0.0 | 11.2 | 0.0 | 11.6 | 0.0 | 4.4 | 0.0 | 4.6 | 0.0 | 6.8 | 0.0 | 7.0 |
| Wetland vegetation | 2.0 | 0.0 | 2.1 | 0.0 | 3.2 | 3.2 | 3.3 | 3.3 | −1.2 | −3.2 | −1.2 | −3.3 |
| Seepage/Burial | 0.0 | 0.0 | 0.0 | 0.0 | 0.8 | 0.0 | 1.5 | 0.0 | −0.8 | 0.0 | −1.5 | 0.0 |
| Water Volume (×109 m3) | Residence Time (Days) | TN (DIN) ×109 g N year−1 | TP (PO4) ×109 g P year−1 | ||||
|---|---|---|---|---|---|---|---|
| Water Inflows | Exchange with GOM | Water Inflows | Exchange with GOM | ||||
| Current River Input | 2009 | 2.13 | 87.7 | 12.8 (8.5) | −2.8 (6.2) | 1.3 (0.55) | −0.5 (0.3) |
| 2010 | 2.08 | 117.8 | 7.9 (5.5) | −0.5 (7.5) | 0.8 (0.37) | −0.3 (0.4) | |
| 2011 | 2.07 | 142.2 | 5.9 (3.3) | 1.8 (8.2) | 0.5 (0.29) | 0.0 (0.6) | |
| Increased River Input | 2009 | 2.15 | 37.1 | 35.5 (26.1) | −20.2 (−1.6) | 3.1 (1.3) | −1.8 (0.0) |
| 2010 | 2.45 | 44.8 | 33.8 (25.3) | −19.0 (−3.0) | 2.8 (1.4) | −1.8 (−0.3) | |
| 2011 | 2.09 | 35.9 | 38.5 (25.1) | −23.6 (−1.7) | 2.8 (1.2) | −1.6 (0.3) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, H.; Nuttle, W.; Baustian, M.M.; Carruthers, T. Influence of Increased Freshwater Inflow on Nitrogen and Phosphorus Budgets in a Dynamic Subtropical Estuary, Barataria Basin, Louisiana. Water 2023, 15, 1974. https://doi.org/10.3390/w15111974
Jung H, Nuttle W, Baustian MM, Carruthers T. Influence of Increased Freshwater Inflow on Nitrogen and Phosphorus Budgets in a Dynamic Subtropical Estuary, Barataria Basin, Louisiana. Water. 2023; 15(11):1974. https://doi.org/10.3390/w15111974
Chicago/Turabian StyleJung, Hoonshin, William Nuttle, Melissa M. Baustian, and Tim Carruthers. 2023. "Influence of Increased Freshwater Inflow on Nitrogen and Phosphorus Budgets in a Dynamic Subtropical Estuary, Barataria Basin, Louisiana" Water 15, no. 11: 1974. https://doi.org/10.3390/w15111974
APA StyleJung, H., Nuttle, W., Baustian, M. M., & Carruthers, T. (2023). Influence of Increased Freshwater Inflow on Nitrogen and Phosphorus Budgets in a Dynamic Subtropical Estuary, Barataria Basin, Louisiana. Water, 15(11), 1974. https://doi.org/10.3390/w15111974