Drought-Induced Salinity Intrusion Affects Nitrogen Removal in a Deltaic Ecosystem (Po River Delta, Northern Italy)

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
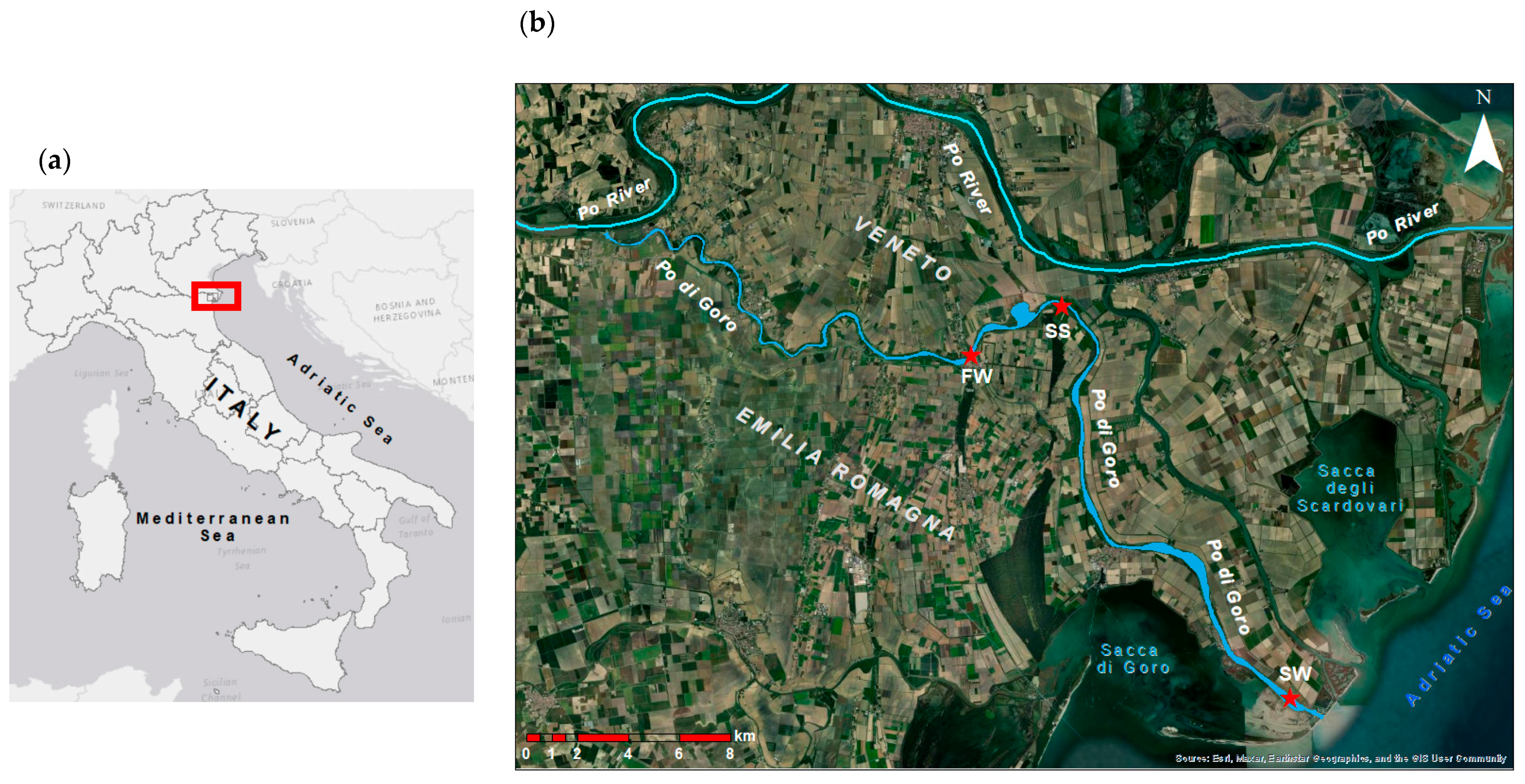
2.1. Area of Study
2.2. Sediment and Water Sampling
2.3. Benthic Flux Measurements and Sediment Characterization
2.4. Measurement of Denitrification and DNRA Rates
2.5. Statistical Analysis
3. Results and Discussion
3.1. The Summer Drought of 2022
3.2. Benthic N Fluxes along the Salinity Gradient
3.3. Denitrification and DNRA Rates
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dugdale, R.C.; Wilkerson, F.P.; Hogue, V.E.; Marchi, A. The Role of Ammonium and Nitrate in Spring Bloom Development in San Francisco Bay. Estuar. Coast. Shelf Sci. 2007, 73, 17–29. [Google Scholar] [CrossRef]
- Howarth, R.W. Coastal Nitrogen Pollution: A Review of Sources and Trends Globally and Regionally. Harmful Algae 2008, 8, 14–20. [Google Scholar] [CrossRef]
- Kroeze, C.; Bouwman, L.; Seitzinger, S. Modeling Global Nutrient Export from Watersheds. Curr. Opin. Environ. Sustain. 2012, 4, 195–202. [Google Scholar] [CrossRef]
- Cornwell, J.C.; Kemp, W.M.; Kana, T.M. Denitrification in coastal ecosystems: Methods, environmental controls, and ecosystem level controls, a review. Aquat. Ecol. 1999, 33, 41–54. [Google Scholar] [CrossRef]
- Santoro, A.E. Microbial Nitrogen Cycling at the Saltwater–Freshwater Interface. Hydrogeol J. 2010, 18, 187–202. [Google Scholar] [CrossRef]
- Zhou, Z.; Ge, L.; Huang, Y.; Liu, Y.; Wang, S. Coupled Relationships among Anammox, Denitrification, and Dissimilatory Nitrate Reduction to Ammonium along Salinity Gradients in a Chinese Estuarine Wetland. J. Environ. Sci. 2021, 106, 39–46. [Google Scholar] [CrossRef]
- Gervasio, M.P.; Soana, E.; Vincenzi, F.; Castaldelli, G. An Underestimated Contribution of Deltaic Denitrification in Reducing Nitrate Export to the Coastal Zone (Po River–Adriatic Sea, Northern Italy). Water 2022, 14, 501. [Google Scholar] [CrossRef]
- Tiedje, J.M.; Sexstone, A.J.; Myrold, D.D.; Robinson, J.A. Denitrification: Ecological Niches, Competition and Survival. Antonie Van Leeuwenhoek 1983, 48, 569–583. [Google Scholar] [CrossRef]
- Piña-Ochoa, E.; Álvarez-Cobelas, M. Denitrification in Aquatic Environments: A Cross-System Analysis. Biogeochemistry 2006, 81, 111–130. [Google Scholar] [CrossRef]
- Burgin, A.J.; Hamilton, S.K. Have We Overemphasized the Role of Denitrification in Aquatic Ecosystems? A Review of Nitrate Removal Pathways. Front. Ecol. Environ. 2007, 5, 89–96. [Google Scholar] [CrossRef]
- Heil, J.; Vereecken, H.; Brüggemann, N. A Review of Chemical Reactions of Nitrification Intermediates and Their Role in Nitrogen Cycling and Nitrogen Trace Gas Formation in Soil: Chemical Reactions of Nitrification Intermediates in Soil. Eur. J. Soil Sci. 2016, 67, 23–39. [Google Scholar] [CrossRef]
- Scott, J.T.; McCarthy, M.J.; Gardner, W.S.; Doyle, R.D. Denitrification, Dissimilatory Nitrate Reduction to Ammonium, and Nitrogen Fixation along a Nitrate Concentration Gradient in a Created Freshwater Wetland. Biogeochemistry 2008, 87, 99–111. [Google Scholar] [CrossRef]
- Xia, R.; Zhang, Y.; Critto, A.; Wu, J.; Fan, J.; Zheng, Z.; Zhang, Y. The Potential Impacts of Climate Change Factors on Freshwater Eutrophication: Implications for Research and Countermeasures of Water Management in China. Sustainability 2016, 8, 229. [Google Scholar] [CrossRef]
- Pörtner, H.-O.; Roberts, D.C.; Tignor, M.; Poloczanska, E.S.; Mintenbeck, K.; Alegría, A.; Craig, M.; Langsdorf, S.; Löschke, S.; Möller, V. (Eds.) IPCC, 2022: Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022; p. 3056. [Google Scholar]
- Gao, X.; Giorgi, F. Increased Aridity in the Mediterranean Region under Greenhouse Gas Forcing Estimated from High Resolution Simulations with a Regional Climate Model. Glob. Planet. Change 2008, 62, 195–209. [Google Scholar] [CrossRef]
- Carvalho, D.; Pereira, S.; Silva, R.; Rocha, A. Aridity and Desertification in the Mediterranean under EURO-CORDEX Future Climate Change Scenarios. Clim. Change 2022, 174, 28. [Google Scholar] [CrossRef]
- Howarth, R.W.; Swaney, D.P.; Butler, T.J.; Marino, R. Rapid Communication: Climatic Control on Eutrophication of the Hudson River Estuary. Ecosystems 2000, 3, 210–215. [Google Scholar] [CrossRef]
- Feyen, L.; Dankers, R. Impact of Global Warming on Streamflow Drought in Europe. J. Geophys. Res. 2009, 114, D17116. [Google Scholar] [CrossRef]
- Statham, P.J. Nutrients in Estuaries—An Overview and the Potential Impacts of Climate Change. Sci. Total Environ. 2012, 434, 213–227. [Google Scholar] [CrossRef]
- Osborne, R.I.; Bernot, M.J.; Findlay, S.E.G. Changes in Nitrogen Cycling Processes Along a Salinity Gradient in Tidal Wetlands of the Hudson River, New York, USA. Wetlands 2015, 35, 323–334. [Google Scholar] [CrossRef]
- Xie, R.; Rao, P.; Pang, Y.; Shi, C.; Li, J.; Shen, D. Salt Intrusion Alters Nitrogen Cycling in Tidal Reaches as Determined in Field and Laboratory Investigations. Sci. Total Environ. 2020, 729, 138803. [Google Scholar] [CrossRef]
- Giblin, A.E.; Weston, N.B.; Banta, G.T.; Tucker, J.; Hopkinson, C.S. The Effects of Salinity on Nitrogen Losses from an Oligohaline Estuarine Sediment. Estuaries Coasts 2010, 33, 1054–1068. [Google Scholar] [CrossRef]
- Marchant, H.K.; Lavik, G.; Holtappels, M.; Kuypers, M.M.M. The Fate of Nitrate in Intertidal Permeable Sediments. PLoS ONE 2014, 9, e104517. [Google Scholar] [CrossRef]
- Herbert, E.R.; Boon, P.; Burgin, A.J.; Neubauer, S.C.; Franklin, R.B.; Ardón, M.; Hopfensperger, K.N.; Lamers, L.P.M.; Gell, P. A Global Perspective on Wetland Salinization: Ecological Consequences of a Growing Threat to Freshwater Wetlands. Ecosphere 2015, 6, art206. [Google Scholar] [CrossRef]
- Gervasio, M.P.; Soana, E.; Granata, T.; Colombo, D.; Castaldelli, G. An Unexpected Negative Feedback between Climate Change and Eutrophication: Higher Temperatures Increase Denitrification and Buffer Nitrogen Loads in the Po River (Northern Italy). Environ. Res. Lett. 2022, 17, 084031. [Google Scholar] [CrossRef]
- Bellafiore, D.; Ferrarin, C.; Maicu, F.; Manfè, G.; Lorenzetti, G.; Umgiesser, G.; Zaggia, L.; Levinson, A.V. Saltwater Intrusion in a Mediterranean Delta Under a Changing Climate. J. Geophys. Res. Ocean. 2021, 126, e2020JC016437. [Google Scholar] [CrossRef]
- Viaroli, P.; Giordani, G.; Bartoli, M.; Naldi, M.; Azzoni, R.; Nizzoli, D.; Ferrari, I.; Comenges, J.M.Z.; Bencivelli, S.; Castaldelli, G.; et al. The Sacca Di Goro Lagoon and an Arm of the Po River. In Estuaries; Wangersky, P.J., Ed.; The Handbook of Environmental Chemistry; Springer: Berlin/Heidelberg, Germany, 2005; Volume 5H, pp. 197–232. ISBN 978-3-540-00270-3. [Google Scholar]
- Bonaldo, D.; Bellafiore, D.; Ferrarin, C.; Ferretti, R.; Ricchi, A.; Sangelantoni, L.; Vitelletti, M.L. The Summer 2022 Drought: A Taste of Future Climate for the Po Valley (Italy)? Reg. Environ. Change 2023, 23, 1. [Google Scholar] [CrossRef]
- Dalsgaard, T.; Nielsen, L.P.; Brotas, V.; Viaroli, P.; Underwood, G.; Nedwell, D.; Sundback, K.; Rysgaard, S.; Miles, A.; Bartoli, M.; et al. Protocol Handbook for NICE-Nitrogen Cycling in Estuaries: A Project under the EU Research Programme: Marine Science and Technology (MAST III); Ministry of Environment and Energy National Environmental Research Institute: Denmark; Department of Lake and Estuarine Ecology: Silkeborg, Denmark, 2000; pp. 1–62. [Google Scholar]
- Armstrong, F.A.J.; Stearns, C.R.; Strickland, J.D.H. The Measurement of Upwelling and Subsequent Biological Process by Means of the Technicon Autoanalyzer® and Associated Equipment. In Deep Sea Research and Oceanographic Abstracts, 1967th ed.; Elsevier: Amsterdam, The Netherlands; Volume 14.
- Golterman, H.L.; Clymo, R.S.; Ohnstand, M.A.M. Methods for Physical and Chemical Analysis of Fresh Waters, I.B.P.; Blackwell: Oxford, MS, USA, 1978; Volume 8. [Google Scholar]
- Bower, C.E.; Holm-Hansen, T. A Salicylate–Hypochlorite Method for Determining Ammonia in Seawater. Can. J. Fish. Aquat. Sci. 1980, 37, 794–798. [Google Scholar] [CrossRef]
- Nielsen, L.P. Denitrification in Sediment Determined from Nitrogen Isotope Pairing. FEMS Microbiol. Lett. 1992, 86, 357–362. [Google Scholar] [CrossRef]
- Kana, T.M.; Darkangelo, C.; Hunt, M.D.; Oldham, J.B.; Bennett, G.E.; Cornwell, J.C. Membrane Inlet Mass Spectrometer for Rapid High-Precision Determination of N2, O2, and Ar in Environmental Water Samples. Anal. Chem. 1994, 66, 4166–4170. [Google Scholar] [CrossRef]
- Eyre, B.; Ferguson, A. Comparison of Carbon Production and Decomposition, Benthic Nutrient Fluxes and Denitrification in Seagrass, Phytoplankton, Benthic Microalgae- and Macroalgae-Dominated Warm-Temperate Australian Lagoons. Mar. Ecol. Prog. Ser. 2002, 229, 43–59. [Google Scholar] [CrossRef]
- Magri, M.; Benelli, S.; Bonaglia, S.; Zilius, M.; Castaldelli, G.; Bartoli, M. The Effects of Hydrological Extremes on Denitrification, Dissimilatory Nitrate Reduction to Ammonium (DNRA) and Mineralization in a Coastal Lagoon. Sci. Total Environ. 2020, 740, 140169. [Google Scholar] [CrossRef]
- Warembourg, F.R. Nitrogen Fixation in Soil and Plant Systems; R Knowles and T H Black-burn; Academic Press Inc.: New York, NY, USA, 1993. [Google Scholar]
- Risgaard-Petersen, N.; Rysgaard, S. Nitrate Reduction in Sediments and Waterlogged Soil Measured by 15N Techniques. In Methods in Applied Soil Microbiology; Academic Press Inc.: London, UK, 1995. [Google Scholar]
- Toreti, A.; Masante, D.; Acosta Navarro, J.; Bavera, D.; Cammalleri, C.; De Felice, M.; de Jager, A.; Di Ciollo, C.; Hrast Es-senfelder, A.; Maetens, W.; et al. Drought in Europe July 2022: GDO Analytical Report; Publications Office of the European Un-Ion: Luxembourg, 2022. [Google Scholar]
- Marchina, C.; Natali, C.; Fazzini, M.; Fusetti, M.; Tassinari, R.; Bianchini, G. Extremely Dry and Warm Conditions in Northern Italy during the Year 2015: Effects on the Po River Water. Rend. Fis. Acc. Lincei 2017, 28, 281–290. [Google Scholar] [CrossRef]
- Chatzidaki, E.; Vecchi, A. Estate 2022: Presente e Futuro Si Toccano; Ecoscienza—ARPAE: Bologna, Italy, 2022. [Google Scholar]
- Montanari, A. Hydrology of the Po River: Looking for Changing Patterns in River Discharge. Hydrol. Earth Syst. Sci. 2012, 16, 3739–3747. [Google Scholar] [CrossRef]
- Autorità di Bacino Distrettuale del Fiume Po. Osservatorio Permanente Sugli Utilizzi Idrici Del Distretto Idrografico Del Fiume Po; Autorità di Bacino Distrettuale del Fiume Po: Parma, Italy, 2022. [Google Scholar]
- Maicu, F.; De Pascalis, F.; Ferrarin, C.; Umgiesser, G. Hydrodynamics of the Po River-Delta-Sea System. J. Geophys. Res. Ocean. 2018, 123, 6349–6372. [Google Scholar] [CrossRef]
- Sholkovitz, E.R. Flocculation of Dissolved Organic and Inorganic Matter during the Mixing of River Water and Seawater. Geochim. Et Cosmochim. Acta 1976, 40, 831–845. [Google Scholar] [CrossRef]
- Asmala, E.; Bowers, D.G.; Autio, R.; Kaartokallio, H.; Thomas, D.N. Qualitative Changes of Riverine Dissolved Organic Matter at Low Salinities Due to Flocculation: Riverine DOM Flocculation. J. Geophys. Res. Biogeosci. 2014, 119, 1919–1933. [Google Scholar] [CrossRef]
- Neubauer, S.C.; Piehler, M.F.; Smyth, A.R.; Franklin, R.B. Saltwater Intrusion Modifies Microbial Community Structure and Decreases Denitrification in Tidal Freshwater Marshes. Ecosystems 2019, 22, 912–928. [Google Scholar] [CrossRef]
- Ardón, M.; Morse, J.L.; Colman, B.P.; Bernhardt, E.S. Drought-Induced Saltwater Incursion Leads to Increased Wetland Nitrogen Export. Glob. Chang. Biol. 2013, 19, 2976–2985. [Google Scholar] [CrossRef]
- Æsøy, A.; Ødegaard, H.; Bentzen, G. The Effect of Sulphide and Organic Matter on the Nitrification Activity in a Biofilm Process. Water Sci. Technol. 1998, 37, 115–122. [Google Scholar] [CrossRef]
- Joye, S.B.; Hollibaugh, J.T. Influence of Sulfide Inhibition of Nitrification on Nitrogen Regeneration in Sediments. Science 1995, 270, 623–625. [Google Scholar] [CrossRef]
- Lisa, J.A.; Song, B.; Tobias, C.R.; Hines, D.E. Genetic and Biogeochemical Investigation of Sedimentary Nitrogen Cycling Communities Responding to Tidal and Seasonal Dynamics in Cape Fear River Estuary. Estuar. Coast. Shelf Sci. 2015, 167, A313–A323. [Google Scholar] [CrossRef]
- Kraft, B.; Tegetmeyer, H.E.; Sharma, R.; Klotz, M.G.; Ferdelman, T.G.; Hettich, R.L.; Geelhoed, J.S.; Strous, M. The Environmental Controls That Govern the End Product of Bacterial Nitrate Respiration. Science 2014, 345, 676–679. [Google Scholar] [CrossRef] [PubMed]
- King, D.; Nedwell, D.B. The Influence of Nitrate Concentration upon the End-Products of Nitrate Dissimilation by Bacteria in Anaerobic Salt Marsh Sediment. FEMS Microbiol. Lett. 1985, 31, 23–28. [Google Scholar] [CrossRef]
- An, S.; Gardner, W. Dissimilatory Nitrate Reduction to Ammonium (DNRA) as a Nitrogen Link, versus Denitrification as a Sink in a Shallow Estuary (Laguna Madre/Baffin Bay, Texas). Mar. Ecol. Prog. Ser. 2002, 237, 41–50. [Google Scholar] [CrossRef]
- Magri, M.; Benelli, S.; Castaldelli, G.; Bartoli, M. The Seasonal Response of in Situ Denitrification and DNRA Rates to Increasing Nitrate Availability. Estuar. Coast. Shelf Sci. 2022, 271, 107856. [Google Scholar] [CrossRef]
- Koop-Jakobsen, K.; Giblin, A.E. The Effect of Increased Nitrate Loading on Nitrate Reduction via Denitrification and DNRA in Salt Marsh Sediments. Limnol. Oceanogr. 2010, 55, 789–802. [Google Scholar] [CrossRef]
- Nizzoli, D.; Carraro, E.; Nigro, V.; Viaroli, P. Effect of Organic Enrichment and Thermal Regime on Denitrification and Dissimilatory Nitrate Reduction to Ammonium (DNRA) in Hypolimnetic Sediments of Two Lowland Lakes. Water Res. 2010, 44, 2715–2724. [Google Scholar] [CrossRef]
- Bonaglia, S.; Klawonn, I.; De Brabandere, L.; Deutsch, B.; Thamdrup, B.; Brüchert, V. Denitrification and DNRA at the Baltic Sea Oxic-Anoxic Interface: Substrate Spectrum and Kinetics: Denitrification and DNRA at the Oxic-Anoxic Interface. Limnol. Oceanogr. 2016, 61, 1900–1915. [Google Scholar] [CrossRef]
- Broman, E.; Zilius, M.; Samuiloviene, A.; Vybernaite-Lubiene, I.; Politi, T.; Klawonn, I.; Voss, M.; Nascimento, F.J.A.; Bonaglia, S. Active DNRA and Denitrification in Oxic Hypereutrophic Waters. Water Res. 2021, 194, 116954. [Google Scholar] [CrossRef]
- Murphy, A.E.; Bulseco, A.N.; Ackerman, R.; Vineis, J.H.; Bowen, J.L. Sulphide Addition Favours Respiratory Ammonification (DNRA) over Complete Denitrification and Alters the Active Microbial Community in Salt Marsh Sediments. Environ. Microbiol 2020, 22, 2124–2139. [Google Scholar] [CrossRef]
- Po River Basin District Authority. Monthly Bullettin of the Permanent Observatory on Water Uses in the Po River District (August 2022); Autorità di Bacino Distrettuale del Fiume Po: Parma, Italy, 2022. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Distance from the Outlet (km) | O2 (mg L−1) | Salinity (mg L−1) | NO3− (µM) | NH4+ (µM) | Ntot (µM) |
|---|---|---|---|---|---|---|
| FW | 25 | 4.7 | 1.5 | 39 | 14 | 107 |
| SS | 22 | 2.9 | 5.4 | 40 | 23 | 118 |
| SW | 1.5 | 5.8 | 30.1 | 11 | 3 | 68 |
| FW | SS | SW | |
|---|---|---|---|
| Typology | Sandy | Muddy–Sandy | Clay |
| Porosity | 0.47 ± 0.12 | 0.65 ± 0.02 | 0.62 ± 0.05 |
| Density (g mL−1) | 1.80 ± 0.11 | 1.47 ± 0.04 | 1.56 ± 0.05 |
| OM (%) | 1.00 | 3.9 | 1.90 |
| Parameter | p | F | Tukey’s Test |
|---|---|---|---|
| NOx− flux | <0.05 | 5.6 | FW vs. SW |
| NH4+ flux | NS | 0.9 | - |
| Dtot | <0.001 | 30.9 | FW vs. SW SS vs. SW |
| Dw | <0.001 | 47.1 | FW vs. SS SS vs. SW FW vs. SW |
| Dn | <0.01 | 15.1 | FW vs. SW SS vs. SW |
| DNRAtot | <0.05 | 4.9 | SS vs. SW |
| DNRAw | NS | 4.1 | - |
| DNRAn | NS | 2.5 | - |
| Location | T (°C) | Salinity (‰) | NO3− (μM) | OM (%) | DNRAtot (μmol N m−2 h−1) | Dtot (μmol N m−2 h−1) | Reference |
|---|---|---|---|---|---|---|---|
| Pit Lake Ca’ Stanga—Hypolimnetic Sediments (Italy) | 6.5 | - | 202 | 1 * | 7 ± 4 | 163 ± 12 | [57] |
| Pit Lake Verde—Hypolimnetic Sediments (Italy) | 12.5 | 102 | 2 * | 4 ± 1 | 31 ± 5 | ||
| Goro Lagoon—Site Giralda (Italy) | 23 | 5 | 32 | 2 * | 48 | 217 | [36] |
| 21.7 | 7 | 79 | 7 | 65 ± 11 | 152 ± 36 | [55] | |
| 105 | 45 ± 11 | 290 ± 94 | |||||
| 236 | 62 ± 8 | 414 ± 123 | |||||
| 437 | 59 ± 11 | 377 ± 65 | |||||
| 874 | 74 ± 8 | 472 ± 50 | |||||
| 1722 | 108 ± 23 | 631 ± 51 | |||||
| Goro Lagoon—Site Gorino (Italy) | 25 | 28 | 23 | 0.24 * | 35 | 72 | [36] |
| 22.5 | 16 | 61 | 9 | 176 ± 14 | 290 ± 58 | [55] | |
| 113 | 127 ± 17 | 399 ± 94 | |||||
| 226 | 241 ± 102 | 690 ± 109 | |||||
| 462 | 150 ± 11 | 713 ± 36 | |||||
| 871 | 334 ± 65 | 1077 ± 225 | |||||
| 1725 | 235 ± 42 | 1304 ± 196 | |||||
| Baltic Sea—Site A | 6 | 10 | 1.3 | - | 0.15 | 40 | [58] |
| Baltic Sea—Site B | 12 | 0.5 | 0.67 | 22 | |||
| Baltic Sea—Site C | 10 | 0.9 | 0.13 | ||||
| Curonian Lagoon—Baltic Sea (Sediments) | 22 | 0.3 | 1 | 13 * | 2 ± 1 | 0.95 ± 0.06 | [59] |
| Plum Island Sound Estuary—Coastal Sediments (Massachusetts, USA) | 23 | 22 | - | 13 ± 2 | 26 ± 0.6 | [60] | |
| Plum Island Sound Estuary—Tidal Creek (Massachusetts, USA) | 24 | 25 | 7 | - | 20 ± 5 | 27 ± 2 | [56] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gervasio, M.P.; Soana, E.; Vincenzi, F.; Magri, M.; Castaldelli, G. Drought-Induced Salinity Intrusion Affects Nitrogen Removal in a Deltaic Ecosystem (Po River Delta, Northern Italy). Water 2023, 15, 2405. https://doi.org/10.3390/w15132405
Gervasio MP, Soana E, Vincenzi F, Magri M, Castaldelli G. Drought-Induced Salinity Intrusion Affects Nitrogen Removal in a Deltaic Ecosystem (Po River Delta, Northern Italy). Water. 2023; 15(13):2405. https://doi.org/10.3390/w15132405
Chicago/Turabian StyleGervasio, Maria Pia, Elisa Soana, Fabio Vincenzi, Monia Magri, and Giuseppe Castaldelli. 2023. "Drought-Induced Salinity Intrusion Affects Nitrogen Removal in a Deltaic Ecosystem (Po River Delta, Northern Italy)" Water 15, no. 13: 2405. https://doi.org/10.3390/w15132405
APA StyleGervasio, M. P., Soana, E., Vincenzi, F., Magri, M., & Castaldelli, G. (2023). Drought-Induced Salinity Intrusion Affects Nitrogen Removal in a Deltaic Ecosystem (Po River Delta, Northern Italy). Water, 15(13), 2405. https://doi.org/10.3390/w15132405