
The Bacterial Degradation of Lignin—A Review

 ,
,  and
and
Abstract
:1. Introduction
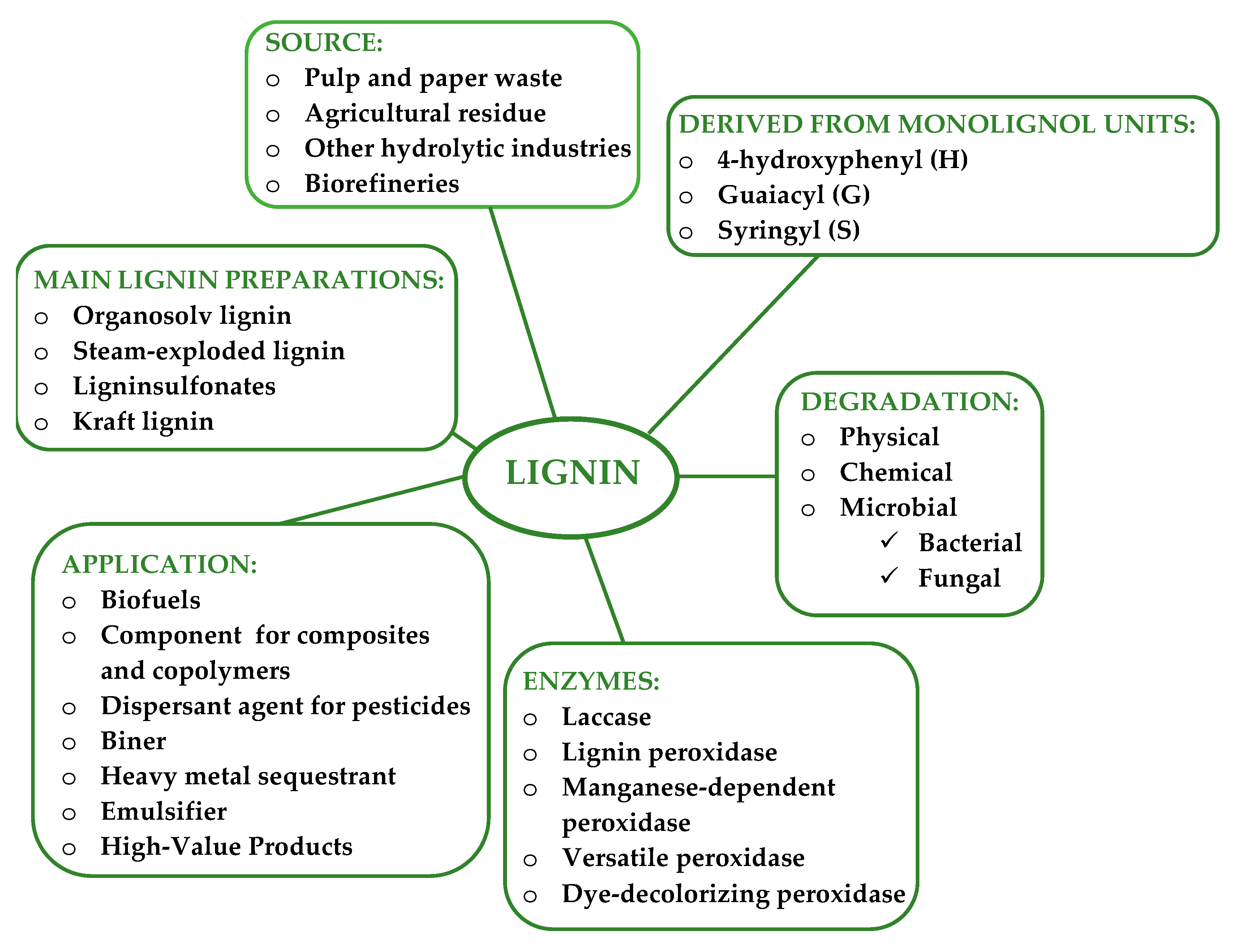
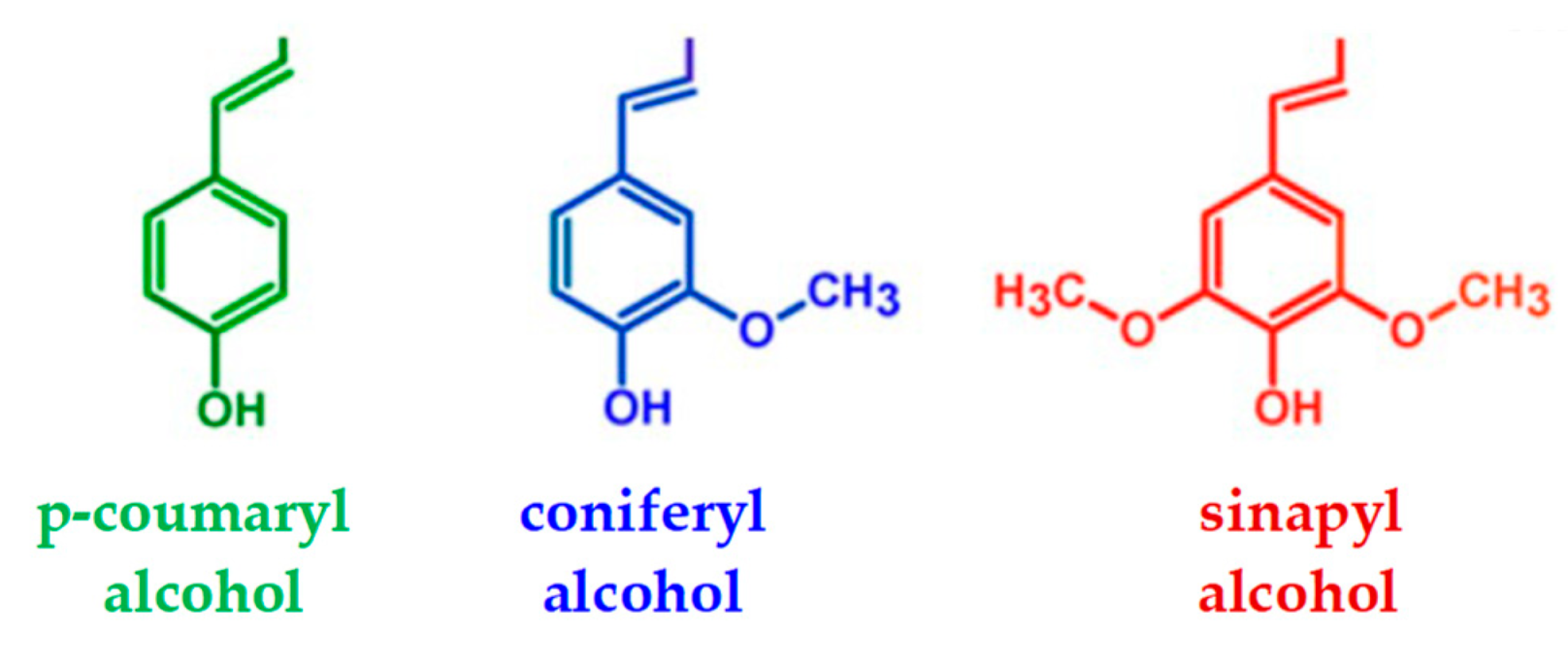
2. Lignin Degradation
3. Biological Lignin Degradation
4. Lignin Degradation by Bacteria

{kind=link}
{kind=link}
{kind=link}
| Bacteria | Reference |
|---|---|
| Bacillus Bacillus ligniniphilus L1 Bacillus sp. Bacillus sp. ITRC-S8 Bacillus magnetrium Bacillus altitudinis SL7 Bacillus flexus RMWW II Bacillus pumilus Bacillus atrophaeus Bacillus cereus Bacillus megaterium Bacillus sp. (AY952465) Bacillus endophyticus Bacillus subtilis | [11] [12] [48,49] [37] [50] [51] [52] [53] [53] [54] [55] [56] [48] [48,57] |
| Arthrobacter sp. C2 | [27] |
| Planococcus sp. TRC1 | [58] |
| Pseudomonas Pseudomonas sp. Q18 Pseudomonas putida NX-1 Pseudomonas putida KT2440 Pseudomonas putida A514 Pseudomonas fluorescens Pseudomonas aeruginosa Pseudomonas plecoglossicida | [11] [59] [60] [61] [62] [40] [63] [50] |
| Brevibacillus thermoruber | [8] |
| Streptomyces Streptomyces sp. Streptomyces badius ATCC 39117 Streptomyces viridosporus T7A Streptomyces coelicolor A3(2) Streptomyces cinnamoneus | [11] [6] [64] [65] [66] [39] |
| Microbacterium sp. | [6] |
| Rhodococcus Rhodococcus opacus PD630 Rhodococcus jostii RHA1 Rhodococcus opacus DSM 1069 Rhodococcus erythropolis | [11] [67,68] [16] [68] [69] |
| Amycolatopsis sp. 75iv2 | [70,71] |
| Enterobacter Enterobacter lignolyticus SCF1 Enterobacter soil sp. nov. Enterobacter aerogenes | [11] [47,72] [73] [74] |
| Paenibacillus sp. (AY952466) Paenibacillus glucanolyticus Paenibacillus sp. strain LD1 | [56] [75] [76] |
| Caldicellulosiruptor bescii | [77] |
| Thermobifida fusca | [78] |
| Clostridium thermocellum | [79] |
| Brucella | [40] |
| Ochrobactrum | [40] |
| Sphingobium | [40] |
| Sphingomonas | [40] |
| Sphingobacterium | [45] |
| Escherichia coli | [40] |
| Aneurinibacillus aneurinilyticus (AY856831) | [56] |
| Azotobacter | [55] |
| Citrobacter sp. Citrobacter freundii | [80] [81] |
| Klebsiella pneumonia | [57,80] |
| Pantoea sp. | [82] |
| Pseudochrobactrum glaciale | [82] |
| Serratia marcescens Serratia liquefaciens | [55,80] [83] |
5. Reports of Lignin Degradation by Bacteria
5.1. Lignin Degradation by Bacillus ligniniphilus L1, Halotolerant Bacterium
5.2. Lignin Degradation by Arthrobacter sp. C2, Psychrotrophic Bacterium
5.3. Lignin Degradation by Pseudomonas sp. Q18
5.4. Lignin Degradation by Brevibacillus thermoruber
5.5. Lignin Degradation by Bacteria Streptomyces sp. and Microbacterium sp., White-Rot Fungi Coriolus versicolor and Trametes gallica, and Purified Laccase Enzyme
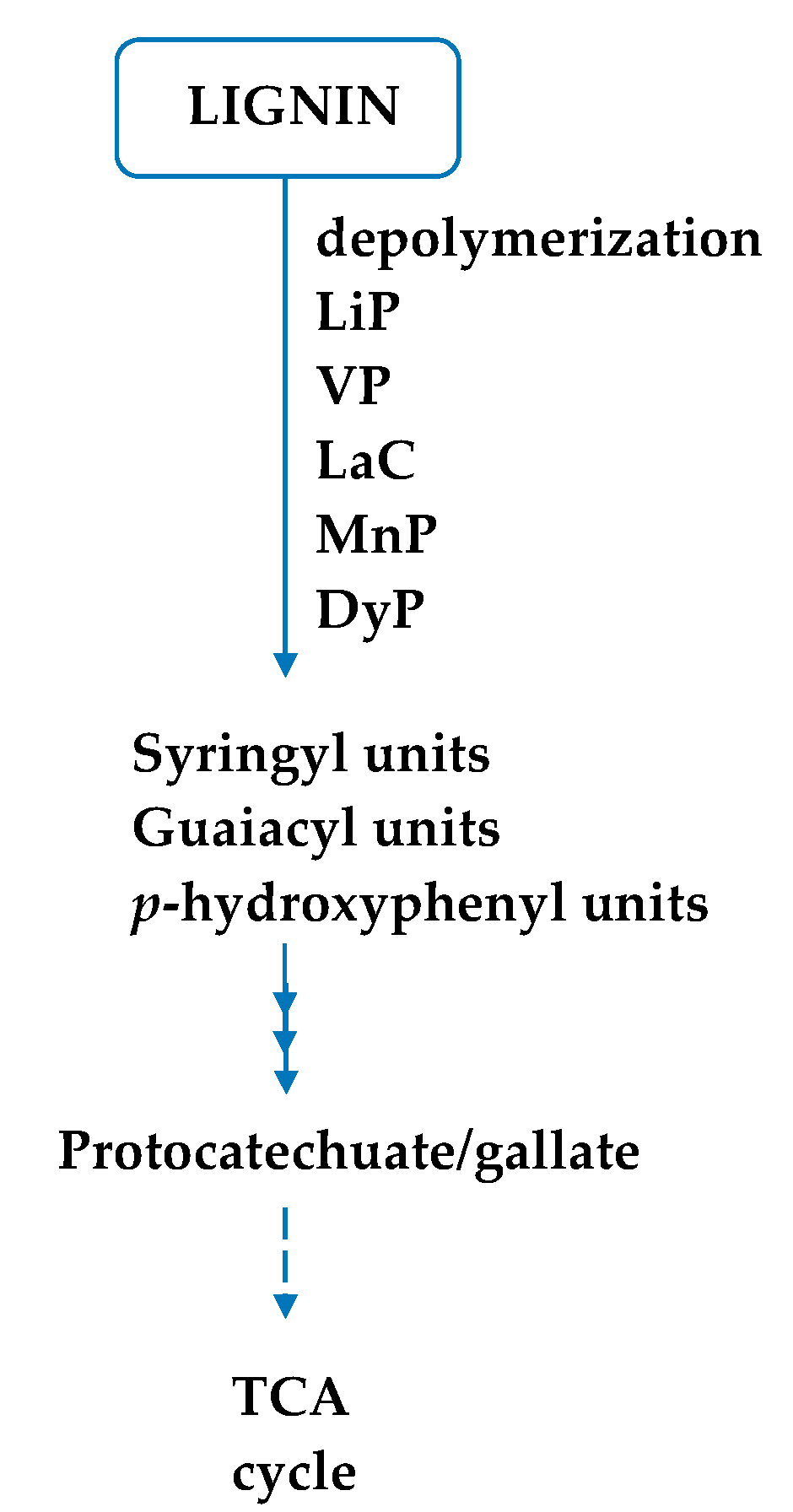
6. Ligninolytic Enzymes
7. Degradation of Lignin from Pulp and Paper Manufacturing Industry
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef] [PubMed]
- Vanholme, R.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin engineering. Curr. Opin. Plant Biol. 2008, 11, 278–285. [Google Scholar] [CrossRef]
- Calvo-Flores, F.G.; Dobado, J.A. Lignin as renewable raw material. ChemSusChem 2010, 3, 1227–1235. [Google Scholar] [CrossRef]
- Christopher, L.P.; Yao, B.; Ji, Y. Lignin biodegradation with laccase-mediator systems. Front. Energy Res. 2014, 2, 12. [Google Scholar] [CrossRef]
- Fisher, A.B.; Fong, S.S. Lignin biodegradation and industrial implications. AIMS Bioeng. 2014, 1, 92–112. [Google Scholar] [CrossRef]
- Asina, F.; Brzonova, I.; Voeller, K.; Kozliak, E.; Kubátová, A.; Yao, B.; Ji, Y. Biodegradation of lignin by fungi, bacteria and laccases. Bioresour. Technol. 2016, 220, 414–424. [Google Scholar] [CrossRef] [Green Version]
- Brebu, M.; Vasile, C. Thermal degradation of lignin—A review. Cellul. Chem. Technol. 2010, 44, 353–363. [Google Scholar]
- Niu, J.; Li, X.; Qi, X.; Ren, Y. Pathway analysis of the biodegradation of lignin by Brevibacillus thermoruber. Bioresour. Technol. 2021, 341, 125875. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.K.; Gupta, N.; Kumar, P.; Dashti, M.G.; Tirth, V.; Khan, S.H.; Yadav, K.K.; Islam, S.; Choudhary, N.; Algahtani, A.; et al. Recent Advances in Synthesis and Degradation of Lignin and Lignin Nanoparticles and Their Emerging Applications in Nanotechnology. Materials 2022, 15, 953. [Google Scholar] [CrossRef] [PubMed]
- Mandal, D.D.; Singh, G.; Majumdar, S.; Chanda, P. Challenges in developing strategies for the valorization of lignin—A major pollutant of the paper mill industry. Environ. Sci. Pollut. Res. 2023, 30, 11119–11140. [Google Scholar] [CrossRef]
- Lee, S.; Kang, M.; Bae, J.H.; Sohn, J.H.; Sung, B.H. Bacterial valorization of lignin: Strains, enzymes, conversion pathways, biosensors, and perspectives. Front. Bioeng. Biotechnol. 2019, 7, 209. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Zhang, P.; Xie, C.; Zhang, W.; Sun, J.; Qian, W.J.; Yang, B. Biodegradation of alkaline lignin by Bacillus ligniniphilus L1. Biotechnol. Biofuels 2017, 10, 44. [Google Scholar] [CrossRef] [Green Version]
- Sigoillot, J.C.; Berrin, J.G.; Bey, M.; Lesage-Meessen, L.; Levasseur, A.; Lomascolo, A.; Record, E.; Uzan-Boukhris, E. Fungal strategies for lignin degradation. In Lignins: Biosynthesis, Biodegradation and Bioengineering; Jouanin, L., Lapierre, C., Eds.; Academic Press: Cambridge, MA, USA, 2012; Volume 61, pp. 263–308. [Google Scholar] [CrossRef]
- Brown, M.E.; Chang, M.C.Y. Exploring bacterial lignin degradation. Curr. Opin. Chem. Biol. 2014, 19, 1–7. [Google Scholar] [CrossRef]
- Bugg, T.D.H.; Ahmad, M.; Hardiman, E.M.; Rahmanpour, R. Pathways for degradation of lignin in bacteria and fungi. Nat. Prod. Rep. 2011, 28, 1883–1896. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Taylor, C.R.; Pink, D.; Burton, K.; Eastwood, D.; Bending, G.D.; Bugg, T.D.H. Development of novel assays for lignin degradation: Comparative analysis of bacterial and fungal lignin degraders. Mol. Biosyst. 2010, 6, 815–821. [Google Scholar] [CrossRef] [PubMed]
- Mandlekar, N.; Cayla, A.; Rault, F.; Giraud, S.; Salaün, F.; Malucelli, G.; Guan, J.-P. An overview on the use of lignin and its derivatives in fire retardant polymer systems. In Lignin-Trends and Applications; Poletto, M., Ed.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.; Abu-Omar, M.M. Chemicals from lignin. In Encyclopedia of Sustainable Technologies; Elsevier: Amsterdam, The Netherlands, 2017; pp. 573–585. ISBN 9780128046777. [Google Scholar]
- Elango, B.; Rajendran, P.; Bornmann, L. A scientometric analysis of international collaboration and growth of literature at the macro level. Malays. J. Libr. Inf. Sci. 2015, 20, 41–50. [Google Scholar]
- Kamali, M.; Khodaparast, Z. Review on recent developments on pulp and paper mill wastewater treatment. Ecotoxicol Env. Safe 2015, 114, 326–342. [Google Scholar] [CrossRef]
- Singhal, A.; Thakur, I.S. Decolourization and detoxification of pulp and paper mill effluent by Emericella nidulans var. nidulans. J. Hazard. Mater. 2009, 171, 619–625. [Google Scholar] [CrossRef]
- Cotana, F.; Cavalaglio, G.; Nicolini, A.; Gelosia, M.; Coccia, V.; Petrozzi, A.; Brinchi, L. Lignin as co-product of second generation bioethanol production from lingo-cellulosic biomass. Energy Procedia 2014, 45, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.-S.; Chang, C.-N.; Chiang, Y.-P.; Sung, H.-F.; Chao, A.C. Photocatalytic degradation of lignin using Pt/TiO2 as the catalyst. Chemosphere 2008, 71, 998–1004. [Google Scholar] [CrossRef]
- Kansal, S.K.; Singh, M.; Sud, D. Studies on TiO2/ZnO photocatalysed degradation of lignin. J. Hazard. Mater. 2008, 153, 412–417. [Google Scholar] [CrossRef]
- Jing, H.; Xue, A.; Wang, Z.; Xia, R.; Wang, L.; Tang, Y.; Wan, P.; Chen, Y. Electrochemical degradation of lignin by ROS. Sustain. Chem. 2020, 1, 345–360. [Google Scholar] [CrossRef]
- Zoghlami, A.; Paës, G. Lignocellulosic biomass: Understanding recalcitrance and predicting hydrolysis. Front. Chem. 2019, 7, 874. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.; Cheng, Y.; Zang, H.; Chen, X.; Wang, Y.; Zhang, Y.; Wang, J.; Shen, X.; Li, C. Biodegradation of lignin and the associated degradation pathway by psychrotrophic Arthrobacter sp. C2 from the cold region of China. Cellulose 2019, 27, 1423–1440. [Google Scholar] [CrossRef]
- Zeng, Y.; Zhao, S.; Yang, S.; Ding, S.Y. Lignin plays a negative role in the biochemical process for producing lignocellulosic biofuels. Curr. Opin. Biotechnol. 2014, 27, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Dueñas, F.J.; Martínez, Á.T. Microbial degradation of lignin: How a bulky recalcitrant polymer is efficiently recycled in nature and how we can take advantage of this. Microb. Biotechnol. 2009, 2, 164–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Hamid, A.M.; Solbiati, J.O.; Cann, I.K.O. Insights into lignin degradation and its potential industrial applications. Adv. Appl. Microbiol. 2013, 82, 1–28. [Google Scholar] [CrossRef]
- Guo, F.F.; Shi, W.J.; Sun, W.; Li, X.Z.; Wang, F.F.; Zhao, J.; Qu, Y. Differences in the adsorption of enzymes onto lignins from diverse types of lignocellulosic biomass and the underlying mechanism. Biotechnol. Biofuels 2014, 7, 38. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, C. Lignocellulosic residues: Biodegradation and bioconversion by fungi. Biotechnol. Adv. 2009, 27, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, O.Y.; Brink, D.P.; Prothmann, J.; Ravi, K.; Sun, M.; García-Hidalgo, J.; Sandahl, M.; Hulteberg, C.P.; Turner, C.; Lidén, G.; et al. Biological valorization of low molecular weight lignin. Biotechnol. Adv. 2016, 34, 1318–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grgas, D.; Štefanac, T.; Barešić, M.; Toromanović, M.; Ibrahimpašić, J.; Vukušić Pavičić, T.; Habuda-Stanić, M.; Herceg, Z.; Landeka Dragičević, T. Co-composting of Sewage Sludge, Green Waste, and Food Waste. J. Sustain. Dev. Energy Water Environ. Syst. 2023, 11, 1100415. [Google Scholar] [CrossRef]
- Tuomela, M.; Vikman, M.; Hatakka, A.; Itävaara, M. Biodegradation of lignin in a compost environment: A review. Bioresour. Technol. 2000, 72, 169–183. [Google Scholar] [CrossRef]
- Knežević, A.; Milovanović, I.; Stajić, M.; Lončar, N.; Brčeski, I.; Vukojević, J.; Cilerdžić, J. Lignin degradation by selected fungal species. Bioresour. Technol. 2013, 138, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Raj, A.; Krishna Reddy, M.M.; Chandra, R. Identification of low molecular weight aromatic compounds by gas chromatography–mass spectrometry (GC–MS) from kraft lignin degradation by three Bacillus sp. Int. Biodeterior. Biodegrad. 2007, 59, 292–296. [Google Scholar] [CrossRef]
- Masai, E.; Katayama, Y.; Fukuda, M. Genetic and biochemical investigations on bacterial catabolic pathways for lignin-derived aromatic compounds. Biosci. Biotechnol. Biochem. 2007, 71, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Janusz, G.; Pawlik, A.; Sulej, J.; Świderska-Burek, U.; Jarosz-Wilkołazka, A.; Paszczyński, A. Lignin degradation: Microorganisms, enzymes involved, genomes analysis and evolution. FEMS Microbiol. Rev. 2017, 41, 941–962. [Google Scholar] [CrossRef] [Green Version]
- Tian, J.H.; Pourcher, A.M.; Bouchez, T.; Gelhaye, E.; Peu, P. Occurrence of lignin degradation genotypes and phenotypes among prokaryotes. Appl. Microbiol. Biotechnol. 2014, 98, 9527–9544. [Google Scholar] [CrossRef] [PubMed]
- Spiker, J.K.; Crawford, D.L.; Thiel, E.C. Oxidation of phenolic and non-phenolic substrates by the lignin peroxidase of Streptomyces viridosporus T7A. Appl. Microbiol. Biotechnol. 1992, 37, 518–523. [Google Scholar] [CrossRef]
- Ahmad, M.; Roberts, J.N.; Hardiman, E.M.; Singh, R.; Eltis, L.D.; Bugg, T.D.H. Identification of DypB from Rhodococcus jostii RHA1 as a lignin peroxidase. Biochemistry 2011, 50, 5096–5107. [Google Scholar] [CrossRef] [PubMed]
- Janusz, G.; Pawlik, A.; Świderska-Burek, U.; Polak, J.; Sulej, J.; Jarosz-Wilkołazka, A.; Paszczyński, A. Laccase Properties, Physiological Functions, and Evolution. Int. J. Mol. Sci. 2020, 21, 966. [Google Scholar] [CrossRef] [Green Version]
- De Gonzalo, G.; Colpa, D.I.; Habib, M.H.M.; Fraaije, M.W. Bacterial enzymes involved in lignin degradation. J. Biotechnol. 2016, 236, 110–119. [Google Scholar] [CrossRef] [Green Version]
- Rashid, G.M.M.; Taylor, C.R.; Liu, Y.; Zhang, X.; Rea, D.; Fülöp, V.; Bugg, T.D.H. Identification of manganese superoxide dismutase from Sphingobacterium sp. T2 as a novel bacterial enzyme for lignin oxidation. ACS Chem. Biol. 2015, 10, 2286–2294. [Google Scholar] [CrossRef] [PubMed]
- Wells, T.; Ragauskas, A.J. Biotechnological opportunities with the β-ketoadipate pathway. Trends Biotechnol 2012, 30, 627–637. [Google Scholar] [CrossRef]
- De Angelis, K.M.; Sharma, D.; Varney, R.; Simmons, B.; Isern, N.G.; Markilllie, L.M.; Nicora, C.; Norbeck, A.D.; Taylor, R.C.; Aldrich, J.T.; et al. Evidence supporting dissimilatory and assimilatory lignin degradation in Enterobacter lignolyticus SCF1. Front. Microbiol. 2013, 4, 280. [Google Scholar] [CrossRef] [Green Version]
- Ojha, A.K.; Tiwari, M. Lignin decolorization and degradation of pulp paper mill effluent by ligninolytic bacteria. Iran. J. Energy Environ. 2016, 7, 282–293. [Google Scholar] [CrossRef]
- Bandounas, L.; Wierckx, N.J.P.; De Winde, J.H.; Ruijssenaars, H.J. Isolation and characterization of novel bacterial strains exhibiting ligninolytic potential. BMC Biotechnol. 2011, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Paliwal, R.; Uniyal, S.; Rai, J.P.N. Evaluating the potential of immobilized bacterial consortium for black liquor biodegradation. Environ. Sci. Pollut. Res. 2015, 22, 6842–6853. [Google Scholar] [CrossRef]
- Khan, S.I.; Zarin, A.; Ahmed, S.; Hasan, F.; Belduz, A.O.; Çanakçi, S.; Khan, S.; Badshah, M.; Farman, M.; Shah, A.A. Degradation of lignin by Bacillus altitudinis SL7 isolated from pulp and paper mill effluent. Water Sci. Technol. 2022, 85, 420–432. [Google Scholar] [CrossRef]
- Kumar, A.; Priyadarshinee, R.; Singha, S.; Sengupta, B.; Roy, A.; Dasgupta, D.; Mandal, T. Biodegradation of alkali lignin by Bacillus flexus RMWW II: Analyzing performance for abatement of rice mill wastewater. Water Sci. Technol. 2019, 80, 1623–1632. [Google Scholar] [CrossRef]
- Huang, X.F.; Santhanam, N.; Badri, D.V.; Hunter, W.J.; Manter, D.K.; Decker, S.R.; Vivanco, J.M.; Reardon, K.F. Isolation and characterization of lignin-degrading bacteria from rainforest soils. Biotechnol. Bioeng. 2013, 110, 1616–1626. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, B.B.; Chandra, R. Biodegradation of phenol in batch culture by pure and mixed strains of Paenibacillus sp. and Bacillus cereus. Pol. J. Microbiol. 2009, 58, 319–325. [Google Scholar] [PubMed]
- Morii, H.; Nakamiya, K.; Kinoshita, S. Isolation of a lignin decolorizing bacterium. J. Ferment. Bioeng. 1995, 80, 296–299. [Google Scholar] [CrossRef]
- Chandra, R.; Raj, A.; Purohit, H.J.; Kapley, A. Characterization and optimization of three potential aerobic bacterial strains for kraft lignin degradation from pulp paper waste. Chemosphere 2007, 67, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Chandra, R. Syntrophic co-culture of Bacillus subtilis and Klebsiella pneumonia for degradation of Kraft lignin discharged from rayon grade pulp industry. J. Environ. Sci. 2015, 33, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, S.; Priyadarshinee, R.; Kumar, A.; Mandal, T.; Dasgupta Mandal, D. Exploring Planococcus sp. TRC1 for carotenoid pigment production and detoxification of paper mill effluent in immobilized fluidized bed reactor. J. Clean. Prod. 2019, 211, 1389–1402. [Google Scholar] [CrossRef]
- Yang, C.; Yue, F.; Cui, Y.; Xu, Y.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Biodegradation of lignin by Pseudomonas sp. Q18 and the characterization of a novel bacterial DyP-type peroxidase. J. Ind. Microbiol. Biotechnol. 2018, 45, 913–927. [Google Scholar] [CrossRef]
- Xu, Z.; Qin, L.; Cai, M.; Hua, W.; Jin, M. Biodegradation of kraft lignin by newly isolated Klebsiella pneumoniae, Pseudomonas putida, and Ochrobactrum tritici strains. Environ. Sci. Pollut. Res. 2018, 25, 14171–14181. [Google Scholar] [CrossRef]
- Salvachúa, D.; Karp, E.M.; Nimlos, C.T.; Vardon, D.R.; Beckham, G.T. Towards lignin consolidated bioprocessing: Simultaneous lignin depolymerization and product generation by bacteria. Green Chem. 2015, 17, 4951–4967. [Google Scholar] [CrossRef]
- Lin, L.; Cheng, Y.; Pu, Y.; Sun, S.; Li, X.; Jin, M.; Pierson, E.A.; Gross, D.C.; Dale, B.E.; Dai, S.Y.; et al. Systems biology-guided biodesign of consolidated lignin conversion. Green Chem. 2016, 18, 5536–5547. [Google Scholar] [CrossRef]
- Tiku, D.K.; Kumar, A.; Chaturvedi, R.; Makhijani, S.D.; Manoharan, A.; Kumar, R. Holistic bioremediation of pulp mill effluents using autochthonous bacteria. Int. Biodeterior. Biodegrad. 2010, 64, 173–183. [Google Scholar] [CrossRef]
- McCarthy, A.J. Lignocellulose-degrading actinomycetes. FEMS Microbiol. Rev. 1987, 3, 145–163. [Google Scholar] [CrossRef]
- Crawford, D.L.; Pometto, A.L.; Crawford, R.L. Lignin degradation by Streptomyces viridosporus: Isolation and characterization of a new polymeric lignin degradation intermediate. Appl. Environ. Microbiol. 1983, 45, 898–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumdar, S.; Lukk, T.; Solbiati, J.O.; Bauer, S.; Nair, S.K.; Cronan, J.E.; Gerlt, J.A. Roles of small laccases from Streptomyces in lignin degradation. Biochemistry 2014, 53, 4047–4058. [Google Scholar] [CrossRef]
- He, Y.; Li, X.; Ben, H.; Xue, X.; Yang, B. Lipid production from dilute alkali corn stover lignin by Rhodococcus strains. ACS Sustain. Chem. Eng. 2017, 5, 2302–2311. [Google Scholar] [CrossRef]
- Kosa, M.; Ragauskas, A.J. Lignin to lipid bioconversion by oleaginous Rhodococci. Green Chem. 2013, 15, 2070–2074. [Google Scholar] [CrossRef]
- Taylor, C.R.; Hardiman, E.M.; Ahmad, M.; Sainsbury, P.D.; Norris, P.R.; Bugg, T.D.H. Isolation of bacterial strains able to metabolize lignin from screening of environmental samples. J. Appl. Microbiol. 2012, 113, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.E.; Walker, M.C.; Nakashige, T.G.; Iavarone, A.T.; Chang, M.C.Y. Discovery and characterization of heme enzymes from unsequenced bacteria: Application to microbial lignin degradation. J. Am. Chem. Soc. 2011, 133, 18006–18009. [Google Scholar] [CrossRef]
- Antai, S.P.; Crawford, D.L. Degradation of softwood, hardwood, and grass lignocelluloses by two Streptomyces strains. Appl. Environ. Microbiol. 1981, 42, 378–380. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, K.M.; D’Haeseleer, P.; Chivian, D.; Fortney, J.L.; Khudyakov, J.; Simmons, B.; Woo, H.; Arkin, A.P.; Davenport, K.W.; Goodwin, L.; et al. Complete genome sequence of “Enterobacter lignolyticus” SCF1. Stand. Genom. Sci. 2011, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Manter, D.K.; Hunter, W.J.; Vivanco, J.M. Enterobacter soli sp. nov.: A lignin-degrading γ-proteobacteria isolated from soil. Curr. Microbiol. 2011, 62, 1044–1049. [Google Scholar] [CrossRef]
- Deschamps, A.M.; Mahoudeau, G.; Lebeault, J.M. Fast degradation of kraft lignin by bacteria. Eur. J. Appl. Microbiol. Biotechnol. 1980, 9, 45–51. [Google Scholar] [CrossRef]
- Mathews, S.L.; Grunden, A.M.; Pawlak, J. Degradation of lignocellulose and lignin by Paenibacillus glucanolyticus. Int. Biodeterior. Biodegrad. 2016, 110, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Raj, A.; Kumar, S.; Haq, I.; Singh, S.K. Bioremediation and toxicity reduction in pulp and paper mill effluent by newly isolated ligninolytic Paenibacillus sp. Ecol. Eng. 2014, 71, 355–362. [Google Scholar] [CrossRef]
- Kataeva, I.; Foston, M.B.; Yang, S.-J.; Pattathil, S.; Biswal, A.K.; Poole, F.L.; Basen, M.; Rhaesa, A.M.; Thomas, T.P.; Azadi, P.; et al. Carbohydrate and lignin are simultaneously solubilized from unpretreated switchgrass by microbial action at high temperature. Energy Environ. Sci. 2013, 6, 2186–2195. [Google Scholar] [CrossRef]
- Deng, Y.; Fong, S.S. Metabolic engineering of Thermobifida fusca for direct aerobic bioconversion of untreated lignocellulosic biomass to 1-propanol. Metab. Eng. 2011, 13, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Akinosho, H.O.; Yoo, C.G.; Dumitrache, A.; Natzke, J.; Muchero, W.; Brown, S.D.; Ragauskas, A.J. Elucidating the structural changes to Populus lignin during consolidated bioprocessing with Clostridium thermocellum. ACS Sustain. Chem. Eng. 2017, 5, 7486–7491. [Google Scholar] [CrossRef]
- Chandra, R.; Abhishek, A.; Sankhwar, M. Bacterial decolorization and detoxification of black liquor from rayon grade pulp manufacturing paper industry and detection of their metabolic products. Bioresour. Technol. 2011, 102, 6429–6436. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R.; Abhishek, A. Bacterial decolorization of black liquor in axenic and mixed condition and characterization of metabolites. Biodegradation 2011, 22, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R.; Singh, R.; Yadav, S. Effect of bacterial inoculum ratio in mixed culture for decolourization and detoxification of pulp paper mill effluent. J. Chem. Technol. Biotechnol. 2012, 87, 436–444. [Google Scholar] [CrossRef]
- Haq, I.; Kumar, S.; Kumari, V.; Singh, S.K.; Raj, A. Evaluation of bioremediation potentiality of ligninolytic Serratia liquefaciens for detoxification of pulp and paper mill effluent. J. Hazard. Mater. 2016, 305, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Shintani, N.; Sugano, Y.; Shoda, M. Decolorization of kraft pulp bleaching effluent by a newly isolated fungus, Geotrichum candidum Dec 1. J. Wood Sci. 2002, 48, 402–408. [Google Scholar] [CrossRef]
- Guo, X.; Xie, C.; Wang, L.; Li, Q.; Wang, Y. Biodegradation of persistent environmental pollutants by Arthrobacter sp. Environ. Sci. Pollut. Res. 2019, 26, 8429–8443. [Google Scholar] [CrossRef]
- Niewerth, H.; Schuldes, J.; Parschat, K.; Kiefer, P.; Vorholt, J.A.; Daniel, R.; Fetzner, S. Complete genome sequence and metabolic potential of the quinaldine-degrading bacterium Arthrobacter sp. Rue61a. BMC Genom. 2012, 13, 534. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Zhou, J.; Gao, Y.; Guan, Y.; Li, J.; Tang, X.; Xu, B.; Ding, J.; Huang, Z. Molecular and biochemical characterizations of a new low-temperature active mannanase. Folia Microbiol. 2015, 60, 483–492. [Google Scholar] [CrossRef]
- Struvay, C.; Feller, G. Optimization to Low Temperature Activity in Psychrophilic Enzymes. Int. J. Mol. Sci. 2012, 13, 11643–11665. [Google Scholar] [CrossRef] [Green Version]
- Winquist, E.; Moilanen, U.; Mettälä, A.; Leisola, M.; Hatakka, A. Production of lignin modifying enzymes on industrial waste material by solid-state cultivation of fungi. Biochem. Eng. J. 2008, 42, 128–132. [Google Scholar] [CrossRef]
- Sharma, P.; Goel, R.; Capalash, N. Bacterial laccases. World J. Microbiol. Biotechnol. 2007, 23, 823–832. [Google Scholar] [CrossRef]
- Margot, J.; Bennati-Granier, C.; Maillard, J.; Blánquez, P.; Barry, D.A.; Holliger, C. Bacterial versus fungal laccase: Potential for micropollutant degradation. AMB Express 2013, 3, 63. [Google Scholar] [CrossRef] [Green Version]
- Adhi, T.P.; Korus, R.A.; Crawford, D.L. Production of major extracellular enzymes during lignocellulose degradation by two Streptomycetes in agitated submerged culture. Appl. Environ. Microbiol. 1989, 55, 1165–1168. [Google Scholar] [CrossRef] [Green Version]
- Camarero, S.; Bockle, B.; Martinez, M.J.; Martinez, A.T. Manganese-mediated lignin degradation by Pleurotus pulmonarius. Appl. Environ. Microbiol. 1996, 62, 1070–1072. [Google Scholar] [CrossRef] [Green Version]
- Koljonen, K.; Österberg, M.; Kleen, M.; Fuhrmann, A.; Stenius, P. Precipitation of lignin and extractives on kraft pulp: Effect on surface chemistry, surface morphology and paper strength. Cellulose 2004, 11, 209–224. [Google Scholar] [CrossRef]
- Lv, Y.; Chen, Y.; Sun, S.; Hu, Y. Interaction among multiple microorganisms and effects of nitrogen and carbon supplementations on lignin degradation. Bioresour. Technol. 2014, 155, 144–151. [Google Scholar] [CrossRef]
- da Silva Coelho-Moreira, J.; Maciel, G.M.; Castoldi, R.; da Silva Mariano, S.; Inácio, F.D.; Bracht, A.; Peralta, R.M. Involvement of lignin-modifying enzymes in the degradation of herbicides. In Herbicides—Advances in Research; Price, A.J., Kelton, J.A., Eds.; InTechOpen: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Hofrichter, M.; Ullrich, R. Heme-thiolate haloperoxidases: Versatile biocatalysts with biotechnological and environmental significance. Appl. Microbiol. Biotechnol. 2006, 71, 276–288. [Google Scholar] [CrossRef]
- Levasseur, A.; Piumi, F.; Coutinho, P.M.; Rancurel, C.; Asther, M.; Delattre, M.; Henrissat, B.; Pontarotti, P.; Asther, M.; Record, E. FOLy: An integrated database for the classification and functional annotation of fungal oxidoreductases potentially involved in the degradation of lignin and related aromatic compounds. Fungal Genet. Biol. 2008, 45, 638–645. [Google Scholar] [CrossRef]
- Messerschmidt, A.; Huber, R. The blue oxidases, ascorbate oxidase, laccase and ceruloplasmin modelling and structural relationships. Eur. J. Biochem. 1990, 187, 341–352. [Google Scholar] [CrossRef]
- Li, K.; Xu, F.; Eriksson, K.E. Comparison of fungal laccases and redox mediators in oxidation of a nonphenolic lignin model compound. Appl. Environ. Microbiol. 1999, 65, 2654–2660. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.W.S. Structure and action mechanism of ligninolytic enzymes. Appl. Biochem. Biotechnol. 2009, 157, 174–209. [Google Scholar] [CrossRef]
- Miki, K.; Renganathan, V.; Gold, M.H. Mechanism of β-aryl ether dimeric lignin model compound oxidation by lignin peroxidase by Phanerochaete chrysosporium. Biochemistry 1986, 25, 4790–4796. [Google Scholar] [CrossRef]
- Khindaria, A.; Yamazaki, I.; Aust, S.D. Stabilization of the veratryl alcohol cation radical by lignin peroxidase. Biochemistry 1996, 35, 6418–6424. [Google Scholar] [CrossRef]
- Kuan, I.C.; Johnson, K.A.; Tien, M. Kinetic analysis of manganese peroxidase. The reaction with manganese complexes. J. Biol. Chem. 1993, 268, 20064–20070. [Google Scholar] [CrossRef]
- Pérez-Boada, M.; Ruiz-Dueñas, F.J.; Pogni, R.; Basosi, R.; Choinowski, T.; Martínez, M.J.; Piontek, K.; Martínez, A.T. Versatile peroxidase oxidation of high redox potential aromatic compounds: Site-directed mutagenesis, spectroscopic and crystallographic investigation of three long-range electron transfer pathways. J. Mol. Biol. 2005, 354, 385–402. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, E.; Mate, D.M.; Gonzalez-Perez, D.; Molina-Espeja, P.; Camarero, S.; Martínez, A.T.; Ballesteros, A.O.; Alcalde, M. Directed evolution of ligninolytic oxidoreductases: From functional expression to stabilization and beyond. In Cascade Biocatalysis; Riva, S., Fessner, W.D., Eds.; Wiley-VCH GmbH & Co. KgaA: Weinheim, Germany, 2014; pp. 1–22. [Google Scholar] [CrossRef]
- Hofrichter, M.; Ullrich, R.; Pecyna, M.J.; Liers, C.; Lundell, T. New and classic families of secreted fungal heme peroxidases. Appl. Microbiol. Biotechnol. 2010, 87, 871–897. [Google Scholar] [CrossRef] [PubMed]
- Zámocký, M.; Hofbauer, S.; Schaffner, I.; Gasselhuber, B.; Nicolussi, A.; Soudi, M.; Pirker, K.F.; Furtmüller, P.G.; Obinger, C. Independent evolution of four heme peroxidase superfamilies. Arch. Biochem. Biophys. 2015, 574, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Falade, A.O.; Nwodo, U.U.; Iweriebor, B.C.; Green, E.; Mabinya, L.V.; Okoh, A.I. Lignin peroxidase functionalities and prospective applications. Microbiologyopen 2017, 6, e00394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, A.; Mendes, S.; Brissos, V.; Martins, L.O. New dye-decolorizing peroxidases from Bacillus subtilis and Pseudomonas putida MET94: Towards biotechnological applications. Appl. Microbiol. Biotechnol. 2014, 98, 2053–2065. [Google Scholar] [CrossRef]
- Ogola, H.J.O.; Kamiike, T.; Hashimoto, N.; Ashida, H.; Ishikawa, T.; Shibata, H.; Sawa, Y. Molecular characterization of a novel peroxidase from the cyanobacterium Anabaena sp. strain PCC 7120. Appl. Environ. Microbiol. 2009, 75, 7509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arapova, O.V.; Chistyakov, A.V.; Tsodikov, M.V.; Moiseev, I.I. Lignin as a Renewable Resource of Hydrocarbon Products and Energy Carriers (A Review). Pet. Chem. 2020, 60, 227–243. [Google Scholar] [CrossRef]
- Colpa, D.I.; Fraaije, M.W.; van Bloois, E. DyP-type peroxidases: A promising and versatile class of enzymes. J. Ind. Microbiol. Biotechnol. 2014, 41, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Rybczyńska-Tkaczyk, K.; Korniłłowicz-Kowalska, T. Biosorption optimization and equilibrium isotherm of industrial dye compounds in novel strains of microscopic fungi. Int. J. Environ. Sci. Technol. 2016, 13, 2837–2846. [Google Scholar] [CrossRef] [Green Version]
- Yeber, M.C.; Silva, T. The ability of a bacterial strain to remove a phenolic structure as an approach to pulp and paper mill wastewater treatment: Optimization by experimental design. Water 2022, 14, 3296. [Google Scholar] [CrossRef]
- Haq, I.; Mazumder, P.; Kalamdhad, A.S. Recent advances in removal of lignin from paper industry wastewater and its industrial applications—A review. Bioresour. Technol. 2020, 312, 123636. [Google Scholar] [CrossRef]
- Costa, S.; Dedola, D.G.; Pellizzari, S.; Blo, R.; Rugiero, I.; Pedrini, P.; Tamburini, E. Lignin biodegradation in pulp-and paper mill wastewater by selected white rot fungi. Water 2017, 9, 935. [Google Scholar] [CrossRef] [Green Version]
- Abhishek, A.; Dwivedi, A.; Tandan, N.; Kumar, U. Comparative bacterial degradation and detoxification of model and Kraft lignin from pulp paper wastewater and its metabolites. Appl. Water Sci. 2017, 7, 757–767. [Google Scholar] [CrossRef] [Green Version]
- Chandra, R.; Singh, R. Decolourization and detoxification of rayon grade pulp paper mill effluent by mixed bacterial culture isolated from pulp paper mill effluent polluted site. Biochem. Eng. J. 2012, 61, 49–58. [Google Scholar] [CrossRef]
- Raghukumar, C.; D’Souza-Ticlo, D.; Verma, A.K. Treatment of colored effluents with lignin-degrading enzymes: An emerging role of marine-derived fungi. Crit. Rev. Microbiol. 2008, 34, 189–206. [Google Scholar] [CrossRef] [PubMed]
- Amr, A.H.; Abd-Elsalam, H.E.; Hafez, E.E. Molecular characterization of two native Egyptian ligninolytic bacterial strains. J. Appl. Sci. Res. 2008, 4, 1291–1296. [Google Scholar]
- Dashtban, M.; Schraft, H.; Syed, T.A.; Qin, W. Fungal biodegradation and enzymatic modification of lignin. Int. J. Biochem. Mol. Biol. 2010, 1, 36–50. [Google Scholar] [PubMed]
- Lokeshwari, N.; Srinikethan, G.; Joshia, S.G.; Shasikala, I.; Srikanth, B.B.; Sushma, L. Isolation of fungi for delignification of Pulp and Paper mill effluent. Int. J. Curr. Eng. Technol. 2013, 1, 124–128. [Google Scholar]
- Gomathi, V.; Ramanathan, A.; Sivaramaiah, N.; Ramanjaneya, V.R.; Jayasimha, D. Decolourization of paper mill effluent by immobilized cells of Phanerochaete chrysosporium. Int. J. Plant Animal Environ. Sci. 2012, 2, 141–146. [Google Scholar]
- Malaviya, P.; Rathore, V.S. Bioremediation of pulp and paper mill effluent by a novel fungal consortium isolated from polluted soil. Bioresour. Technol. 2007, 98, 3647–3651. [Google Scholar] [CrossRef]
- Singhal, A.; Thakur, I.S. Decolourization and detoxification of pulp and paper mill effluent by Cryptococcus sp. Biochem. Eng. J. 2009, 46, 21–27. [Google Scholar] [CrossRef]
- Sumathi, S.; Hung, Y.T. Treatment of pulp and paper mill wastes. In Waste Treatment in the Process Industries; Sumathi, S., Ed.; CRC Press: Boca Raton, FL, USA, 2006; pp. 453–497. [Google Scholar]
- Barapatre, A.; Jha, H. Degradation of alkali lignin by two ascomycetes and free radical scavenging activity of the products. Biocatal Biotransform. 2017, 35, 269–286. [Google Scholar] [CrossRef]
- Haq, I.; Kumar, S.; Raj, A.; Lohani, M.; Satyanarayana, G.N.V. Genotoxicity assessment of pulp and paper mill effluent before and after bacterial degradation using Allium cepa test. Chemosphere 2017, 169, 642–650. [Google Scholar] [CrossRef] [PubMed]
- Hossain, K.; Ismail, N. Bioremediation and detoxification of pulp and paper mill effluent: A review. Res. J. Environ. Toxicol. 2015, 9, 113–134. [Google Scholar] [CrossRef] [Green Version]
- Tyagi, S.; Kumar, V.; Singh, J.; Teotia, P.; Bisht, S.; Sharma, S. Bioremediation of pulp and paper mill effluent by dominant aboriginal microbes and their consortium. Int. J. Environ. Res. 2014, 8, 561–568. [Google Scholar] [CrossRef]
- Majumdar, S.; Paul, I.; Dey, S.; Dutta, S.; Mandal, T.; Dasgupta Mandal, D. Biotransformation of paper mill sludge by Serratia marcescens NITDPER1 for prodigiosin and cellulose nanocrystals: A strategic valorization approach. Biochem. Eng. J. 2020, 164, 107766. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grgas, D.; Rukavina, M.; Bešlo, D.; Štefanac, T.; Crnek, V.; Šikić, T.; Habuda-Stanić, M.; Landeka Dragičević, T. The Bacterial Degradation of Lignin—A Review. Water 2023, 15, 1272. https://doi.org/10.3390/w15071272
Grgas D, Rukavina M, Bešlo D, Štefanac T, Crnek V, Šikić T, Habuda-Stanić M, Landeka Dragičević T. The Bacterial Degradation of Lignin—A Review. Water. 2023; 15(7):1272. https://doi.org/10.3390/w15071272
Chicago/Turabian StyleGrgas, Dijana, Matea Rukavina, Drago Bešlo, Tea Štefanac, Vlado Crnek, Tanja Šikić, Mirna Habuda-Stanić, and Tibela Landeka Dragičević. 2023. "The Bacterial Degradation of Lignin—A Review" Water 15, no. 7: 1272. https://doi.org/10.3390/w15071272
APA StyleGrgas, D., Rukavina, M., Bešlo, D., Štefanac, T., Crnek, V., Šikić, T., Habuda-Stanić, M., & Landeka Dragičević, T. (2023). The Bacterial Degradation of Lignin—A Review. Water, 15(7), 1272. https://doi.org/10.3390/w15071272