
The Response of NDVI to Drought at Different Temporal Scales in the Yellow River Basin from 2003 to 2020

Abstract
1. Introduction
2. Materials and Methods
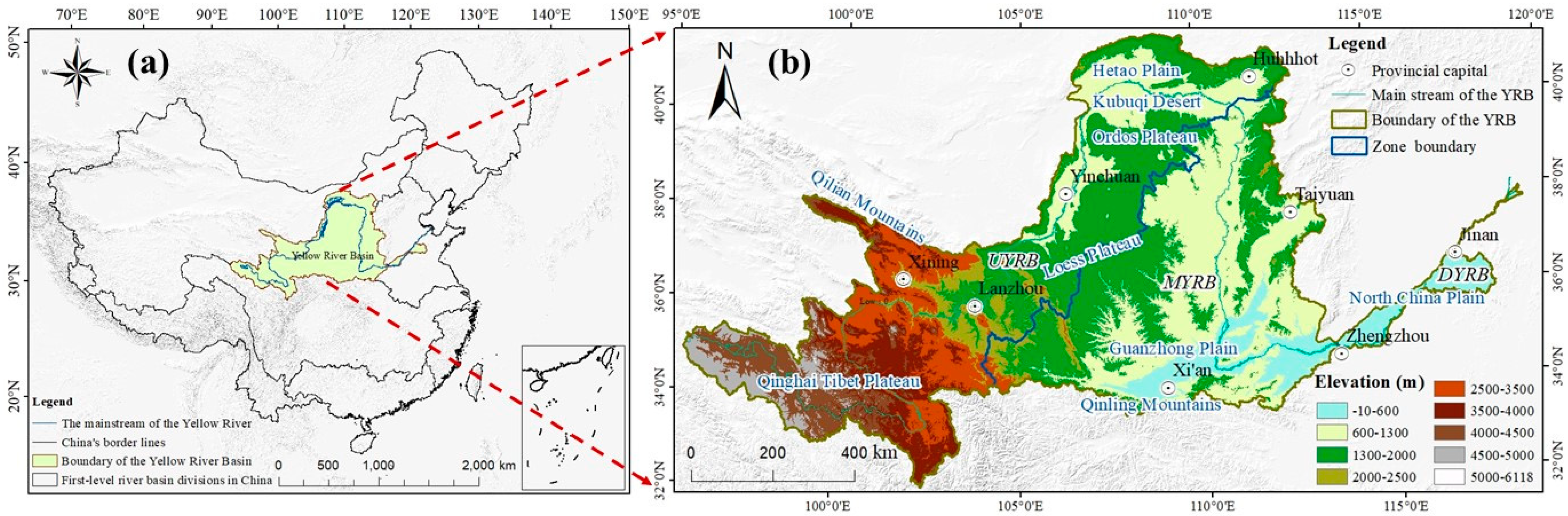
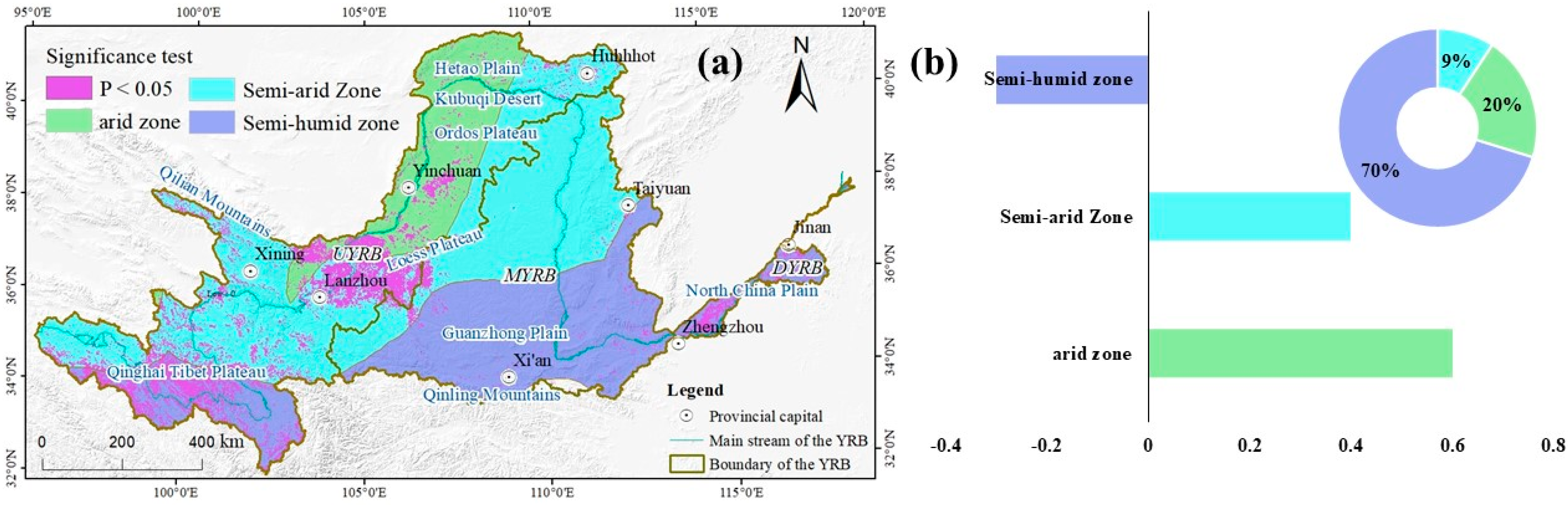
2.1. Study Area
2.2. Method and Materials
2.2.1. Data Source
2.2.2. Linear Trend Method
2.2.3. Correlation Analysis
3. Results
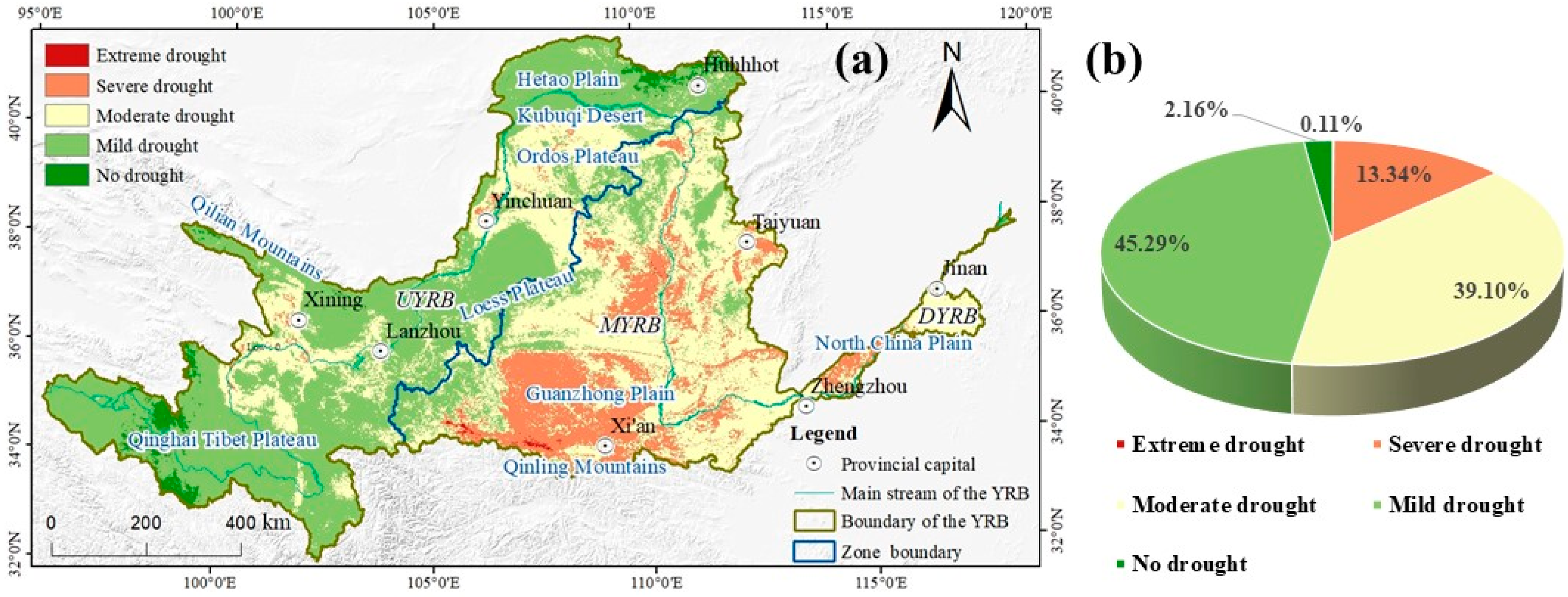
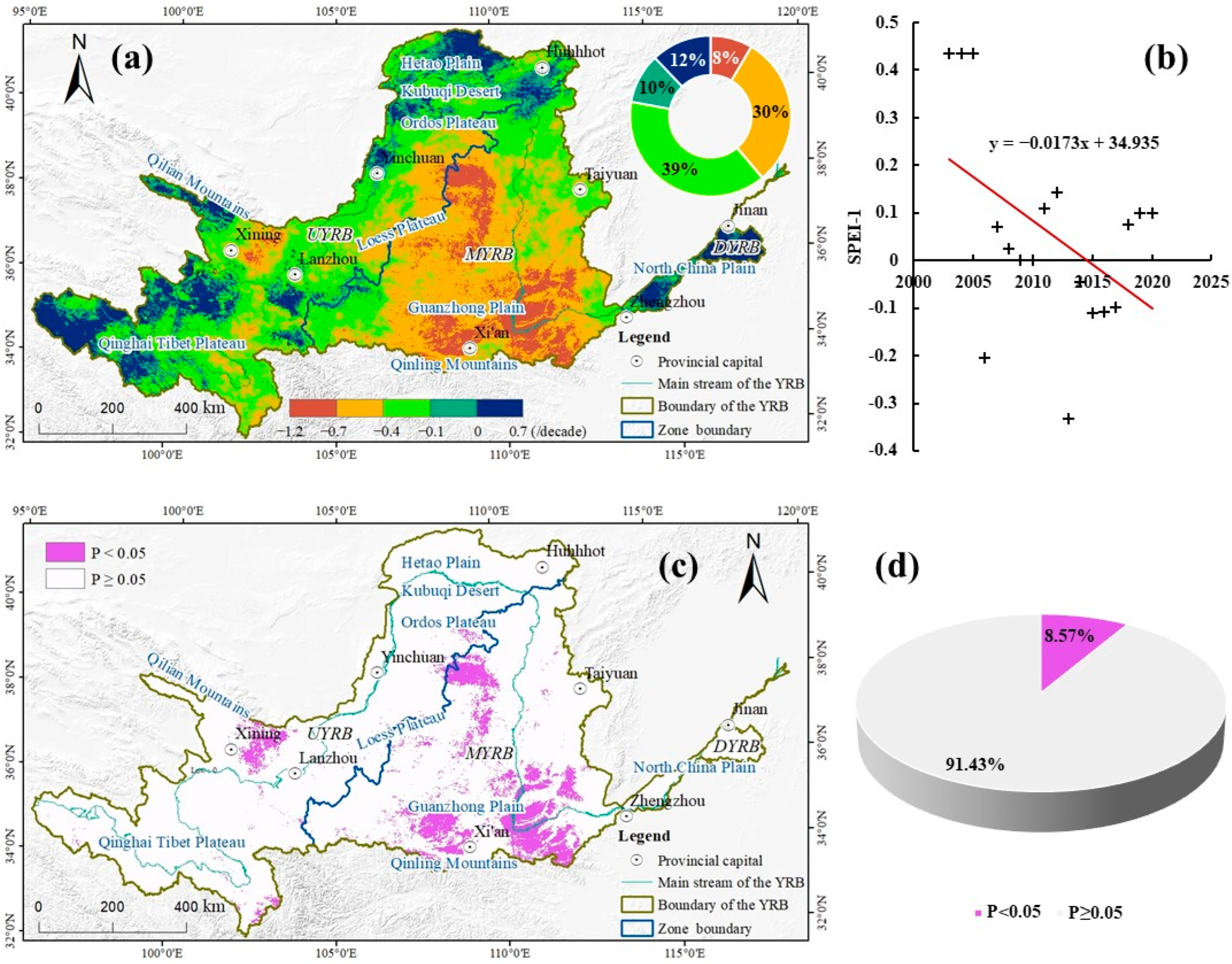
3.1. Spatio-Temporal Distribution Characteristics of SPEI
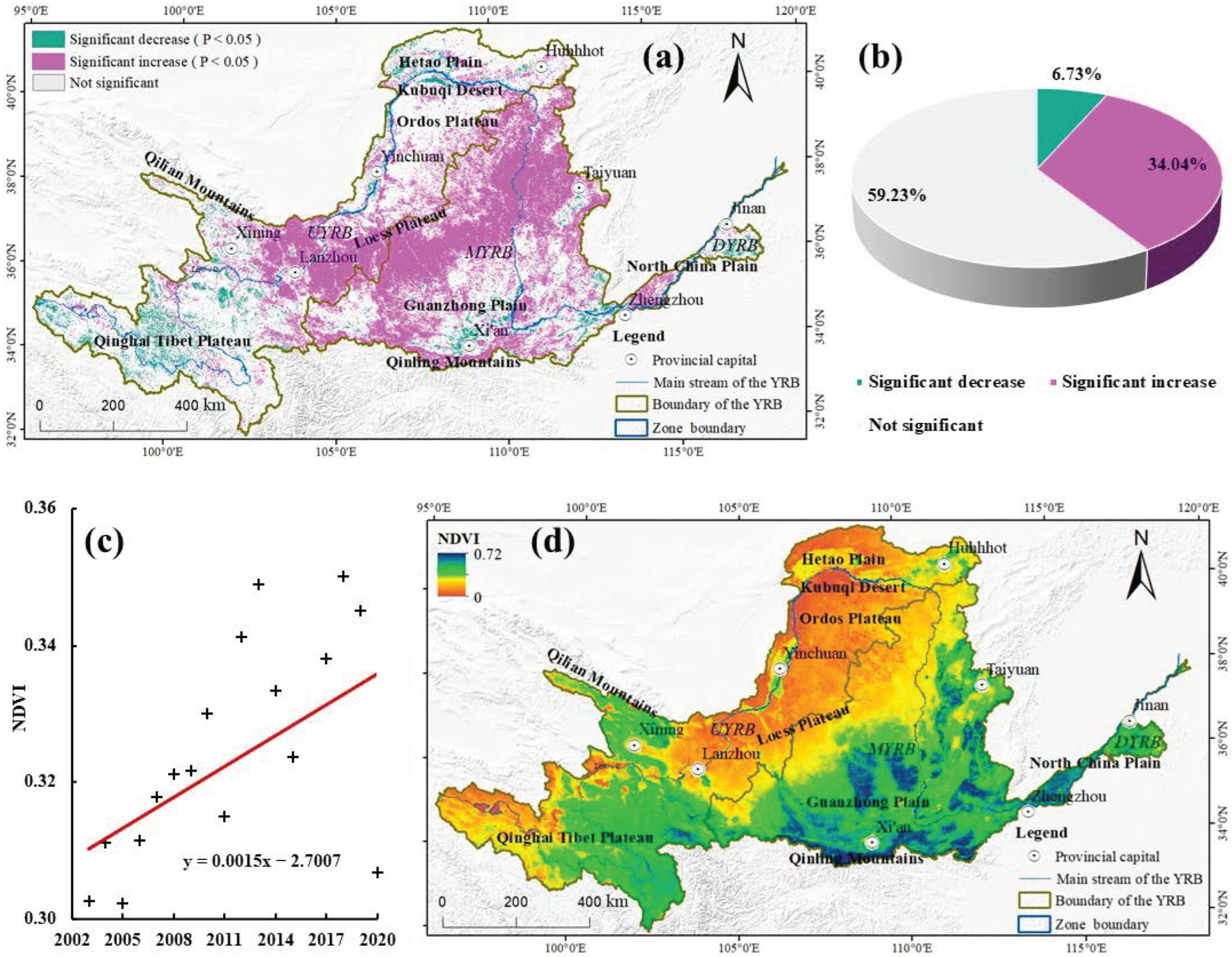
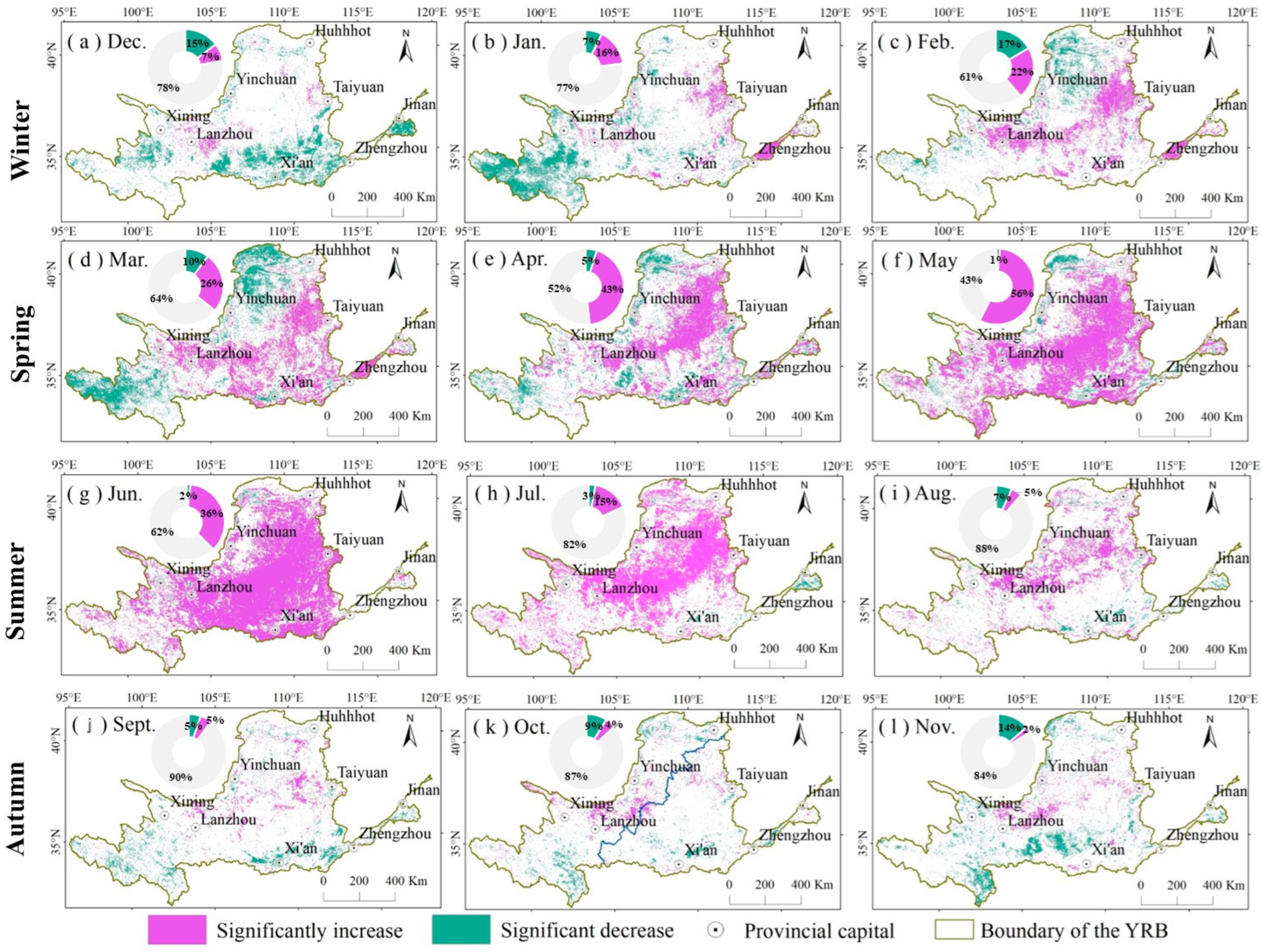
3.2. Spatial and Temporal Distribution Characteristics of NDVI
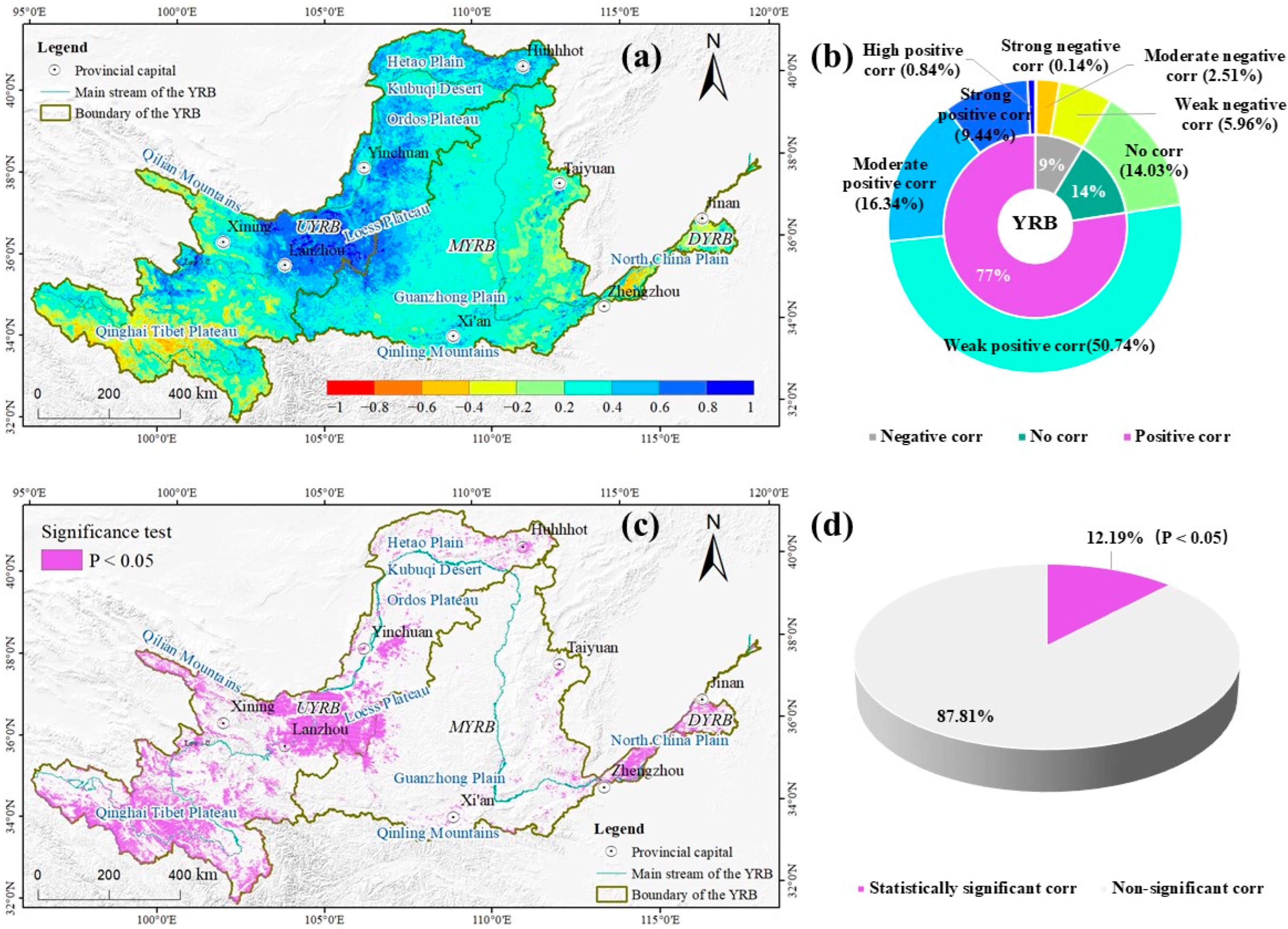
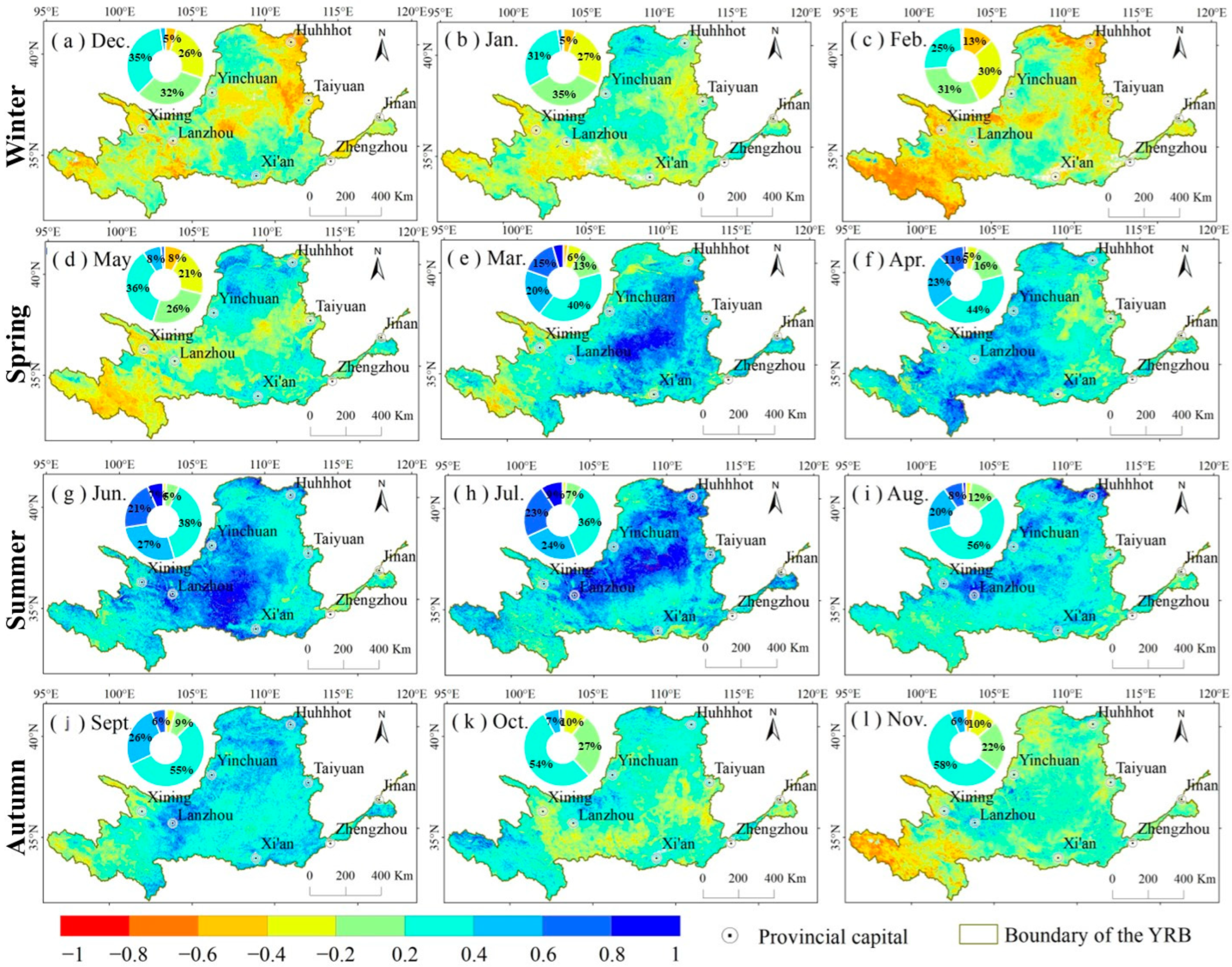
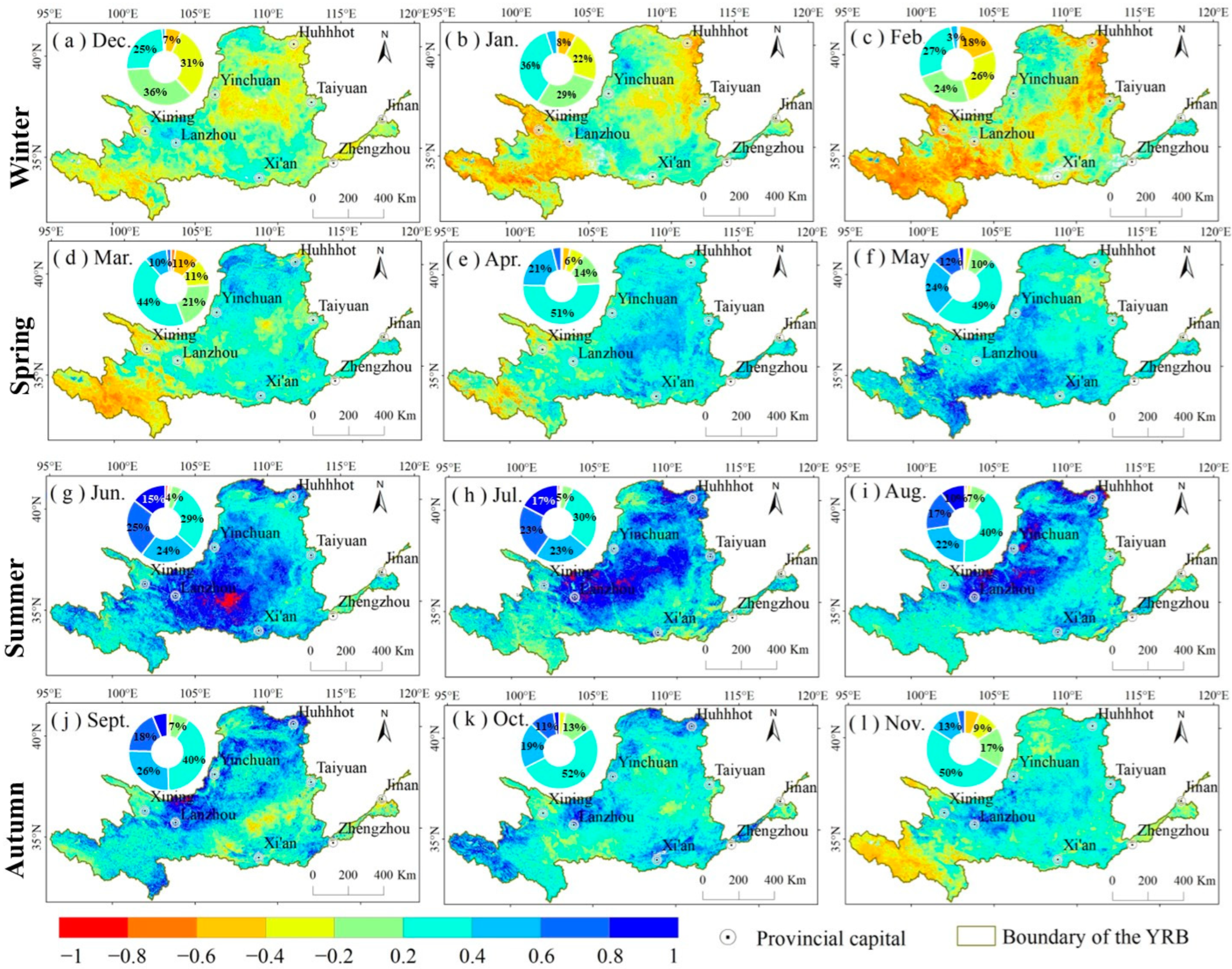
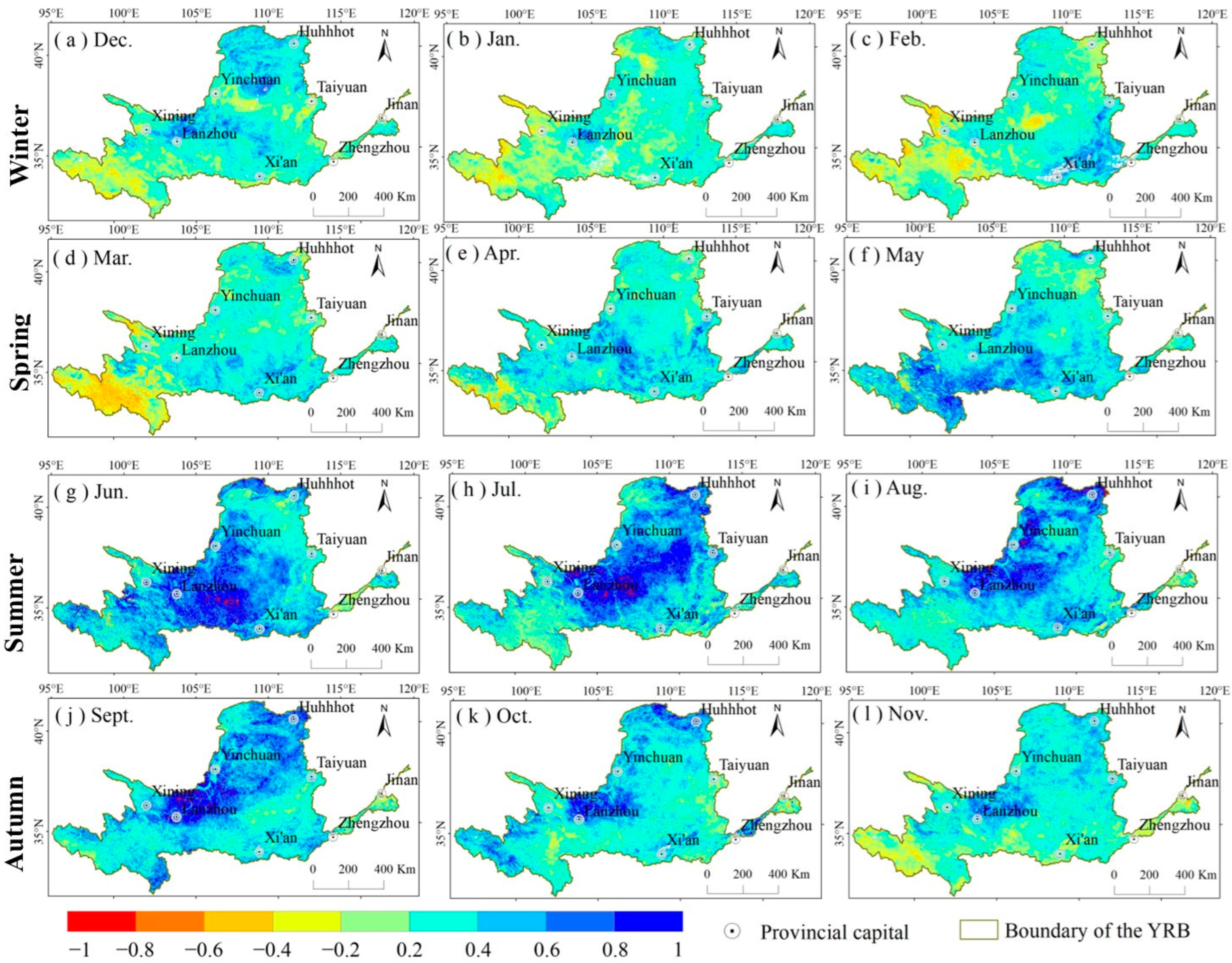
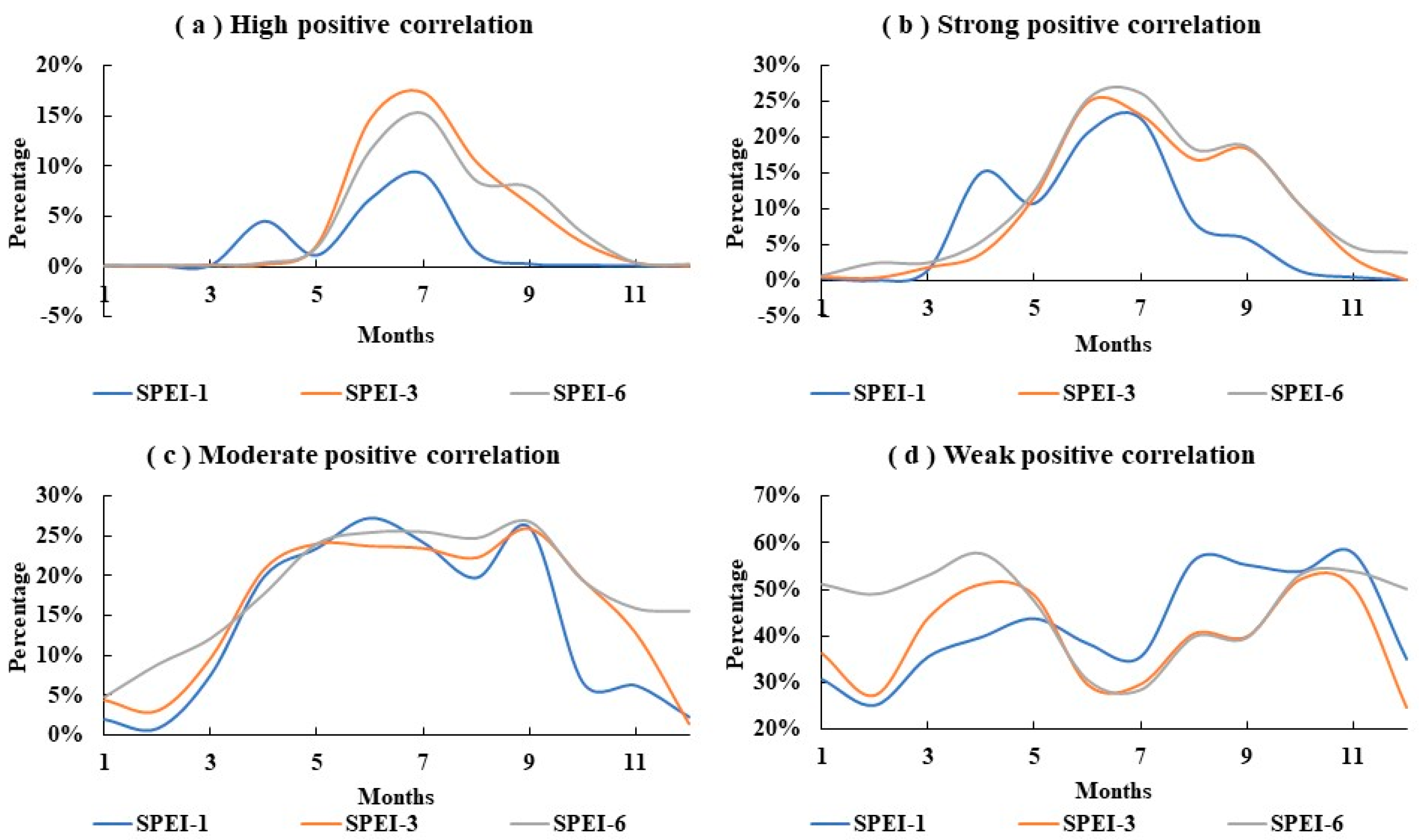
3.3. The Response of NDVI to SPEI
4. Discussion
4.1. The Impact Mechanism of SPEI on NDVI
4.2. Response of NDVI to SPEI
4.3. Uncertainty and Vegetation Management Recommendations
5. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- Novick, K.A.; Ficklin, D.L.; Stoy, P.C.; Williams, C.A.; Bohrer, G.; Oishi, A.C.; Papuga, S.A.; Blanken, P.D.; Noormets, A.; Sulman, B.N.; et al. The increasing importance of atmospheric demand for ecosystem water and carbon fluxes. Nat. Clim. Chang. 2016, 6, 1023–1027. [Google Scholar] [CrossRef]
- Huang, K.; Xia, J.; Wang, Y.; Ahlström, A.; Chen, J.; Cook, R.B.; Cui, E.; Fang, Y.; Fisher, J.B.; Huntzinger, D.N.; et al. Enhanced peak growth of global vegetation and its key mechanisms. Nat. Ecol. Evol. 2018, 2, 1897–1905. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Dickinson, R.E. A review of global terrestrial evapotranspiration: Observation, modeling, climatology, and climatic variability. Rev. Geophys. 2012, 50. [Google Scholar] [CrossRef]
- Piao, S.; Zhang, X.; Chen, A.; Liu, Q.; Lian, X.; Wang, X.; Peng, S.; Wu, X. The impacts of climate extremes on the terrestrial carbon cycle: A review. Sci. China Earth Sci. 2019, 62, 1551–1563. [Google Scholar] [CrossRef]
- Jiao, W.; Wang, L.; Smith, W.K.; Chang, Q.; Wang, H.; D’Odorico, P. Observed increasing water constraint on vegetation growth over the last three decades. Nat. Commun. 2021, 12, 3777. [Google Scholar] [CrossRef]
- Peterson, T.J.; Saft, M.; Peel, M.C.; John, A. Watersheds may not recover from drought. Science 2021, 372, 745–749. [Google Scholar] [CrossRef]
- Shu, Z.; Jin, J.; Zhang, J.; Wang, G.; Lian, Y.; Liu, Y.; Bao, Z.; Guan, T.; He, R.; Liu, C.; et al. 1.5 °C and 2.0 °C of global warming intensifies the hydrological extremes in China. J. Hydrol. 2024, 635, 131229. [Google Scholar] [CrossRef]
- Furtak, K.; Wolińska, A. The impact of extreme weather events as a consequence of climate change on the soil moisture and on the quality of the soil environment and agriculture—A review. Catena 2023, 231, 107378. [Google Scholar] [CrossRef]
- Cao, X.; Zheng, Y.; Lei, Q.; Li, W.; Song, S.; Wang, C.; Liu, Y.; Khan, K. Increasing actual evapotranspiration on the Loess Plateau of China: An insight from anthropologic activities and climate change. Ecol. Indic. 2023, 157, 111235. [Google Scholar] [CrossRef]
- Liu, Y.; Lin, Z.; Wang, Z.; Chen, X.; Han, P.; Wang, B.; Wang, Z.; Wen, Z.; Shi, H.; Zhang, Z.; et al. Discriminating the impacts of vegetation greening and climate change on the changes in evapotranspiration and transpiration fraction over the Yellow River Basin. Sci. Total Environ. 2023, 904, 166926. [Google Scholar] [CrossRef]
- Schumacher, D.L.; Keune, J.; Dirmeyer, P.; Miralles, D.G. Drought self-propagation in drylands due to land–atmosphere feedbacks. Nat. Geosci. 2022, 15, 262–268. [Google Scholar] [CrossRef]
- Yuan, W.; Zheng, Y.; Piao, S.; Ciais, P.; Lombardozzi, D.; Wang, Y.; Ryu, Y.; Chen, G.; Dong, W.; Hu, Z.; et al. Increased atmospheric vapor pressure deficit reduces global vegetation growth. Sci. Adv. 2019, 5, eaax1396. [Google Scholar] [CrossRef]
- Lucht, W.; Prentice, I.C.; Myneni, R.B.; Sitch, S.; Friedlingstein, P.; Cramer, W.; Bousquet, P.; Buermann, W.; Smith, B. Climatic Control of the High-Latitude Vegetation Greening Trend and Pinatubo Effect. Science 2002, 296, 1687–1689. [Google Scholar] [CrossRef]
- Zhu, Z.; Piao, S.; Myneni, R.B.; Huang, M.; Zeng, Z.; Canadell, J.G.; Ciais, P.; Sitch, S.; Friedlingstein, P.; Arneth, A.; et al. Greening of the Earth and its drivers. Nat. Clim. Chang. 2016, 6, 791–795. [Google Scholar] [CrossRef]
- Wang, Z.; Li, J.; Hou, J.; Zhao, K.; Wu, R.; Sun, B.; Lu, J.; Liu, Y.; Cui, C.; Liu, J. Enhanced evapotranspiration induced by vegetation restoration may pose water resource risks under climate change in the Yellow River Basin. Ecol. Indic. 2024, 162, 112060. [Google Scholar] [CrossRef]
- Li, X.; Xu, X.; Tian, W.; Tian, J.; He, C. Contribution of climate change and vegetation restoration to interannual variability of evapotranspiration in the agro–pastoral ecotone in northern China. Ecol. Indic. 2023, 154, 110485. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Rennenberg, H. Molecular and physiological responses of trees to waterlogging stress. Plant Cell Environ. 2014, 37, 2245–2259. [Google Scholar] [CrossRef]
- Lan, X.; Xie, Y.; Liu, Z.; Yang, T.; Huang, L.; Chen, X.; Chen, X.; Lin, K.; Cheng, L. Vegetation greening accelerated hydrological drought in two-thirds of river basins over China. J. Hydrol. 2024, 637, 131436. [Google Scholar] [CrossRef]
- Zhao, M.; Running, S.W. Drought-Induced Reduction in Global Terrestrial Net Primary Production from 2000 Through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef]
- Xu, W.; Yuan, W.; Wu, D.; Zhang, Y.; Shen, R.; Xia, X.; Ciais, P.; Liu, J. Impacts of record-breaking compound heatwave and drought events in 2022 China on vegetation growth. Agric. For. Meteorol. 2024, 344, 109799. [Google Scholar] [CrossRef]
- Xu, H.-j.; Wang, X.-p.; Zhao, C.-y.; Yang, X.-m. Diverse responses of vegetation growth to meteorological drought across climate zones and land biomes in northern China from 1981 to 2014. Agric. For. Meteorol. 2018, 262, 1–13. [Google Scholar] [CrossRef]
- Ye, J.; Gao, Z.; Wu, X.; Lu, Z.; Li, C.; Wang, X.; Chen, L.; Cui, G.; Yu, M.; Yan, G.; et al. Impact of increased temperature on spring wheat yield in northern China. Food Energy Secur. 2021, 10, 368–378. [Google Scholar] [CrossRef]
- Li, Y.; Huang, Y.; Li, Y.; Zhang, H.; Fan, J.; Deng, Q.; Wang, X. Spatiotemporal heterogeneity in meteorological and hydrological drought patterns and propagations influenced by climatic variability, LULC change, and human regulations. Sci. Rep. 2024, 14, 5965. [Google Scholar] [CrossRef]
- Wu, J.; Miao, C.; Zheng, H.; Duan, Q.; Lei, X.; Li, H. Meteorological and Hydrological Drought on the Loess Plateau, China: Evolutionary Characteristics, Impact, and Propagation. J. Geophys. Res. Atmos. 2018, 123, 11–569. [Google Scholar] [CrossRef]
- Apurv, T.; Sivapalan, M.; Cai, X. Understanding the Role of Climate Characteristics in Drought Propagation. Water Resour. Res. 2017, 53, 9304–9329. [Google Scholar] [CrossRef]
- Van Loon, A.F.; Tijdeman, E.; Wanders, N.; Van Lanen, H.A.J.; Teuling, A.J.; Uijlenhoet, R. How climate seasonality modifies drought duration and deficit. J. Geophys. Res. Atmos. 2014, 119, 4640–4656. [Google Scholar] [CrossRef]
- Zhan, C.; Liang, C.; Zhao, L.; Jiang, S.; Niu, K.; Zhang, Y. Drought-related cumulative and time-lag effects on vegetation dynamics across the Yellow River Basin, China. Ecol. Indic. 2022, 143, 109409. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M. Response of vegetation to drought time-scales across global land biomes. Proc. Natl. Acad. Sci. USA 2013, 110, 52–57. [Google Scholar] [CrossRef]
- Wu, D.; Vargas, G.G.; Powers, J.S.; McDowell, N.G.; Becknell, J.M.; Pérez-Aviles, D.; Medvigy, D.; Liu, Y.; Katul, G.G.; Calvo-Alvarado, J.C.; et al. Reduced ecosystem resilience quantifies fine-scale heterogeneity in tropical forest mortality responses to drought. Glob. Chang. Biol. 2021, 28, 2081–2094. [Google Scholar] [CrossRef]
- Jiang, X.; Fang, X.; Zhu, Q.; Jin, J.; Ren, L.; Jiang, S.; Yan, Y.; Yuan, S.; Liao, M. Time-series satellite images reveal abrupt changes in vegetation dynamics and possible determinants in the Yellow River Basin. Agric. For. Meteorol. 2024, 355, 110124. [Google Scholar] [CrossRef]
- Niu, H.; Xiu, Z.; Xiao, D. Impact of land-use change on ecological vulnerability in the Yellow River Basin based on a complex network model. Ecol. Indic. 2024, 166, 112212. [Google Scholar] [CrossRef]
- Shen, J.; Zhao, M.; Tan, Z.; Zhu, L.; Guo, Y.; Li, Y.; Wu, C. Ecosystem service trade-offs and synergies relationships and their driving factor analysis based on the Bayesian belief Network: A case study of the Yellow River Basin. Ecol. Indic. 2024, 163, 112070. [Google Scholar] [CrossRef]
- Ci, F.; Wang, Z.; Hu, Q. Spatial pattern characteristics and optimization policies of low-carbon innovation levels in the urban agglomerations in the Yellow River Basin. J. Clean. Prod. 2024, 439, 140856. [Google Scholar] [CrossRef]
- Wu, J.; Chen, X. Spatiotemporal trends of dryness/wetness duration and severity: The respective contribution of precipitation and temperature. Atmos. Res. 2019, 216, 176–185. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Liang, R.G. Spatial-Temporal Variation Characteristics and Influencing Factors of Drought in Karst Region of Southwest China Based on Remote Sensing. Master’s Thesis, Guizhou Normal University, Guiyang, China, 2022. [Google Scholar]
- Xue, H.; Li, Y.; Dong, G. Analysis of Spatial-temporal Variation Characteristics of Meteorological Drought in the Hexi Corridor Based on SPEI Index. Chin. J. Agrometeorol. 2022, 43, 932–934. [Google Scholar] [CrossRef]
- Zhang, X.; Duan, Y.; Duan, J.; Jian, D.; Ma, Z. A daily drought index based on evapotranspiration and its application in regional drought analyses. Sci. China Earth Sci. 2021, 65, 317–336. [Google Scholar] [CrossRef]
- Li, Y.; Qin, Y. The Response of Net Primary Production to Climate Change: A Case Study in the 400 mm Annual Precipitation Fluctuation Zone in China. Int. J. Environ. Res. Public Health 2019, 16, 1497. [Google Scholar] [CrossRef]
- Ding, Y.; Xu, J.; Wang, X.; Cai, H.; Zhou, Z.; Sun, Y.; Shi, H. Propagation of meteorological to hydrological drought for different climate regions in China. J. Environ. Manag. 2021, 283, 111980. [Google Scholar] [CrossRef]
- Ding, Y.; Xu, J.; Wang, X.; Peng, X.; Cai, H. Spatial and temporal effects of drought on Chinese vegetation under different coverage levels. Sci. Total Environ. 2020, 716, 137166. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, X.; Hao, Z.; Singh, V.P.; Hao, F. Characterization of agricultural drought propagation over China based on bivariate probabilistic quantification. J. Hydrol. 2021, 598, 126194. [Google Scholar] [CrossRef]
- Jiang, T.; Su, X.; Singh, V.P.; Zhang, G. Spatio-temporal pattern of ecological droughts and their impacts on health of vegetation in Northwestern China. J. Environ. Manag. 2022, 305, 114356. [Google Scholar] [CrossRef]
- Ge, W.; Han, J.; Zhang, D.; Wang, F. Divergent impacts of droughts on vegetation phenology and productivity in the Yungui Plateau, southwest China. Ecol. Indic. 2021, 127, 107743. [Google Scholar] [CrossRef]
- Wang, C.-P.; Huang, M.-T.; Zhai, P.-M. Change in drought conditions and its impacts on vegetation growth over the Tibetan Plateau. Adv. Clim. Chang. Res. 2021, 12, 333–341. [Google Scholar] [CrossRef]
- Chen, J.; Shao, Z.; Huang, X.; Zhuang, Q.; Dang, C.; Cai, B.; Zheng, X.; Ding, Q. Assessing the impact of drought-land cover change on global vegetation greenness and productivity. Sci. Total Environ. 2022, 852, 158499. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Y.; Ju, W.; Porcar-Castell, A.; Ye, S.; Zhang, Z.; Brümmer, C.; Urbaniak, M.; Mammarella, I.; Juszczak, R.; et al. Warmer spring alleviated the impacts of 2018 European summer heatwave and drought on vegetation photosynthesis. Agric. For. Meteorol. 2020, 295, 108195. [Google Scholar] [CrossRef]
- Frank, D.; Reichstein, M.; Bahn, M.; Thonicke, K.; Frank, D.; Mahecha, M.D.; Smith, P.; van der Velde, M.; Vicca, S.; Babst, F.; et al. Effects of climate extremes on the terrestrial carbon cycle: Concepts, processes and potential future impacts. Glob. Chang. Biol. 2015, 21, 2861–2880. [Google Scholar] [CrossRef]
- Sippel, S.; Reichstein, M.; Ma, X.; Mahecha, M.D.; Lange, H.; Flach, M.; Frank, D. Drought, Heat, and the Carbon Cycle: A Review. Curr. Clim. Chang. Rep. 2018, 4, 266–286. [Google Scholar] [CrossRef]
- Li, D.; Li, X.; Li, Z.; Fu, Y.; Zhang, J.; Zhao, Y.; Wang, Y.; Liang, E.; Rossi, S. Drought limits vegetation carbon sequestration by affecting photosynthetic capacity of semi-arid ecosystems on the Loess Plateau. Sci. Total Environ. 2024, 912, 168778. [Google Scholar] [CrossRef]
- Cao, S.; He, Y.; Zhang, L.; Chen, Y.; Yang, W.; Yao, S.; Sun, Q. Spatiotemporal characteristics of drought and its impact on vegetation in the vegetation region of Northwest China. Ecol. Indic. 2021, 133, 108420. [Google Scholar] [CrossRef]
- Song, W.; Song, R.; Zhao, Y.; Zhao, Y. Research on the characteristics of drought stress state based on plant stem water content. Sustain. Energy Technol. Assess. 2023, 56, 103080. [Google Scholar] [CrossRef]
- Li, Q.; Xue, Y. Simulated impacts of land cover change on summer climate in the Tibetan Plateau. Environ. Res. Lett. 2010, 5, 015102. [Google Scholar] [CrossRef]
- Jiang, W.; Wang, L.; Zhang, M.; Yao, R.; Chen, X.; Gui, X.; Sun, J.; Cao, Q. Analysis of drought events and their impacts on vegetation productivity based on the integrated surface drought index in the Hanjiang River Basin, China. Atmos. Res. 2021, 254, 105536. [Google Scholar] [CrossRef]
- Jha, S.; Srivastava, R. Impact of drought on vegetation carbon storage in arid and semi-arid regions. Remote Sens. Appl. Soc. Environ. 2018, 11, 22–29. [Google Scholar] [CrossRef]
- Zheng, L.; Lu, J.; Chen, X. Drought offsets the vegetation greenness-induced gross primary productivity from 1982 to 2018 in China. J. Hydrol. 2024, 632, 130881. [Google Scholar] [CrossRef]
- Gouveia, C.M.; Trigo, R.M.; Beguería, S.; Vicente-Serrano, S.M. Drought impacts on vegetation activity in the Mediterranean region: An assessment using remote sensing data and multi-scale drought indicators. Glob. Planet. Chang. 2017, 151, 15–27. [Google Scholar] [CrossRef]
- Scheiter, S.; Kumar, D.; Pfeiffer, M.; Langan, L. Modeling drought mortality and resilience of savannas and forests in tropical Asia. Ecol. Model. 2024, 494, 110783. [Google Scholar] [CrossRef]
- Holtmann, A.; Huth, A.; Bohn, F.; Fischer, R. Assessing the impact of multi-year droughts on German forests in the context of increased tree mortality. Ecol. Model. 2024, 492, 110696. [Google Scholar] [CrossRef]
- Socha, J.; Hawryło, P.; Tymińska-Czabańska, L.; Reineking, B.; Lindner, M.; Netzel, P.; Grabska-Szwagrzyk, E.; Vallejos, R.; Reyer, C.P.O. Higher site productivity and stand age enhance forest susceptibility to drought-induced mortality. Agric. For. Meteorol. 2023, 341, 109680. [Google Scholar] [CrossRef]
- Shen, C.; Ma, R. Estimating suitable hydrothermal conditions for vegetation growth for land use cover across China based on maximum-probability-density monthly NDVI. Remote Sens. Appl. Soc. Environ. 2023, 30, 100958. [Google Scholar] [CrossRef]
- Weng, Z.; Niu, J.; Guan, H.; Kang, S. Three-dimensional linkage between meteorological drought and vegetation drought across China. Sci. Total Environ. 2023, 859, 160300. [Google Scholar] [CrossRef]
- Jiang, L.; Guli·Jiapaer; Bao, A.; Guo, H.; Ndayisaba, F. Vegetation dynamics and responses to climate change and human activities in Central Asia. Sci. Total Environ. 2017, 599–600, 967–980. [Google Scholar] [CrossRef]
- Yuan, M.; Zhao, L.; Lin, A.; Wang, L.; Li, Q.; She, D.; Qu, S. Impacts of preseason drought on vegetation spring phenology across the Northeast China Transect. Sci. Total Environ. 2020, 738, 140297. [Google Scholar] [CrossRef]
- Rahmati, M.; Graf, A.; Poppe Terán, C.; Amelung, W.; Dorigo, W.; Franssen, H.-J.H.; Montzka, C.; Or, D.; Sprenger, M.; Vanderborght, J.; et al. Continuous increase in evaporative demand shortened the growing season of European ecosystems in the last decade. Commun. Earth Environ. 2023, 4, 236. [Google Scholar] [CrossRef]
- Li, Z.; Bai, X.; Tan, Q.; Zhao, C.; Li, Y.; Luo, G.; Chen, F.; Li, C.; Ran, C.; Zhang, S.; et al. Dryness stress weakens the sustainability of global vegetation cooling. Sci. Total Environ. 2024, 909, 168474. [Google Scholar] [CrossRef]
- Wu, C.; Zhong, L.; Yeh, P.J.F.; Gong, Z.; Lv, W.; Chen, B.; Zhou, J.; Li, J.; Wang, S. An evaluation framework for quantifying vegetation loss and recovery in response to meteorological drought based on SPEI and NDVI. Sci. Total Environ. 2024, 906, 167632. [Google Scholar] [CrossRef]
- Akula, N.N.; Abdelhakim, L.; Knazovický, M.; Ottosen, C.-O.; Rosenqvist, E. Plant responses to co-occurring heat and water deficit stress: A comparative study of tolerance mechanisms in old and modern wheat genotypes. Plant Physiol. Biochem. 2024, 210, 108595. [Google Scholar] [CrossRef]
- Zhao, H.; Huang, Y.; Wang, X.; Li, X.; Lei, T. The performance of SPEI integrated remote sensing data for monitoring agricultural drought in the North China Plain. Field Crop. Res. 2023, 302, 109041. [Google Scholar] [CrossRef]
- Liu, S.; Xue, L.; Xiao, Y.; Yang, M.; Liu, Y.; Han, Q.; Ma, J. Dynamic process of ecosystem water use efficiency and response to drought in the Yellow River Basin, China. Sci. Total Environ. 2024, 934, 173339. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Dai, Q.-Y.; Lu, Y.-G.; Zhao, C.; Huang, W.-T.; Xu, M.; Feng, Y.-X. Identification of ecologically sensitive zones affected by climate change and anthropogenic activities in Southwest China through a NDVI-based spatial-temporal model. Ecol. Indic. 2024, 158, 111482. [Google Scholar] [CrossRef]
- Zhang, Q.; Kong, D.; Singh, V.P.; Shi, P. Response of vegetation to different time-scales drought across China: Spatiotemporal patterns, causes and implications. Glob. Planet. Chang. 2017, 152, 1–11. [Google Scholar] [CrossRef]
- Xiao, L.; Zhou, J.; Wu, X.; Anas Khan, M.; Zhao, S.; Wu, X. The dominant influence of terrain and geology on vegetation mortality in response to drought: Exploring resilience and resistance. Catena 2024, 243, 108156. [Google Scholar] [CrossRef]
- Zhang, F.; Geng, M.; Wu, Q.; Liang, Y. Study on the spatial-temporal variation in evapotranspiration in China from 1948 to 2018. Sci. Rep. 2020, 10, 17139. [Google Scholar] [CrossRef]
- Hollunder, R.K.; Mariotte, P.; Carrijo, T.T.; Holmgren, M.; Luber, J.; Stein-Soares, B.; Guidoni-Martins, K.G.; Ferreira-Santos, K.; Scarano, F.R.; Garbin, M.L. Topography and vegetation structure mediate drought impacts on the understory of the South American Atlantic Forest. Sci. Total Environ. 2021, 766, 144234. [Google Scholar] [CrossRef]
- Konapala, G.; Mishra, A.K.; Wada, Y.; Mann, M.E. Climate change will affect global water availability through compounding changes in seasonal precipitation and evaporation. Nat. Commun. 2020, 11, 3044. [Google Scholar] [CrossRef]
- Padrón, R.S.; Gudmundsson, L.; Decharme, B.; Ducharne, A.; Lawrence, D.M.; Mao, J.; Peano, D.; Krinner, G.; Kim, H.; Seneviratne, S.I. Observed changes in dry-season water availability attributed to human-induced climate change. Nat. Geosci. 2020, 13, 477–481. [Google Scholar] [CrossRef]
- Liu, M.; Huang, J.; Sun, A.Y.; Wang, K.; Chen, H. What roles can water-stressed vegetation play in agricultural droughts? Sci. Total Environ. 2022, 803, 149810. [Google Scholar] [CrossRef]
- Buermann, W.; Forkel, M.; O’Sullivan, M.; Sitch, S.; Friedlingstein, P.; Haverd, V.; Jain, A.K.; Kato, E.; Kautz, M.; Lienert, S.; et al. Widespread seasonal compensation effects of spring warming on northern plant productivity. Nature 2018, 562, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, S.; Wang, H.; Huang, Q.; Li, P.; Zheng, X.; Wang, Z.; Jiang, S.; Leng, G.; Li, J.; et al. Warming and greening exacerbate the propagation risk from meteorological to soil moisture drought. J. Hydrol. 2023, 622, 129716. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, X.; Yang, L.; Chen, N.; Nam, W.-H.; Niyogi, D. Urbanization-induced drought modification: Example over the Yangtze River Basin, China. Urban Clim. 2022, 44, 101231. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drought Grade | SPEI Value |
|---|---|
| Extreme drought | SPEI ≤ −2.00 |
| Severe drought | −2 < SPE I ≤ −1.5 |
| Moderate drought | −1.5 < SPEI ≤ −1 |
| Mild drought | −1 < SPEI ≤ −0.5 |
| Normal | SPEI ≥ −0.5 |
| Level of CC | CC |
|---|---|
| High positive correlation | 0.8 ≤ CC < 1.0 |
| Strong positive correlation | 0.6 ≤ CC < 0.8 |
| Moderate positive correlation | 0.4 ≤ CC < 0.6 |
| Weak positive correlation | 0.2 ≤ CC < 0.4 |
| No correlation | −0.2 < CC < 0.2 |
| Weak negative correlation | −0.4 < CC ≤ −0.2 |
| Moderate negative correlation | −0.6 < CC ≤ −0.4 |
| Strong negative correlation | −0.8 < CC ≤ −0.6 |
| High negative correlation | −1 < CC ≤ −0.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W. The Response of NDVI to Drought at Different Temporal Scales in the Yellow River Basin from 2003 to 2020. Water 2024, 16, 2416. https://doi.org/10.3390/w16172416
Liu W. The Response of NDVI to Drought at Different Temporal Scales in the Yellow River Basin from 2003 to 2020. Water. 2024; 16(17):2416. https://doi.org/10.3390/w16172416
Chicago/Turabian StyleLiu, Wen. 2024. "The Response of NDVI to Drought at Different Temporal Scales in the Yellow River Basin from 2003 to 2020" Water 16, no. 17: 2416. https://doi.org/10.3390/w16172416
APA StyleLiu, W. (2024). The Response of NDVI to Drought at Different Temporal Scales in the Yellow River Basin from 2003 to 2020. Water, 16(17), 2416. https://doi.org/10.3390/w16172416