
Microtopography Affects the Diversity and Stability of Vegetation Communities by Regulating Soil Moisture

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Areas and Experimental Design
2.2. Vegetation Survey
2.3. Soil Data
2.4. Data Analysis
2.4.1. Vegetation Parameters
- (1)
- Vegetation diversity
- (2)
- Vegetation stability
- Cumulative percentage of species:
- Cumulative relative frequency:
- Smooth curve fitting equations:
2.4.2. Soil Moisture Content
3. Results
3.1. Characterization of Soil Water Content Under Different Microtopographic Conditions
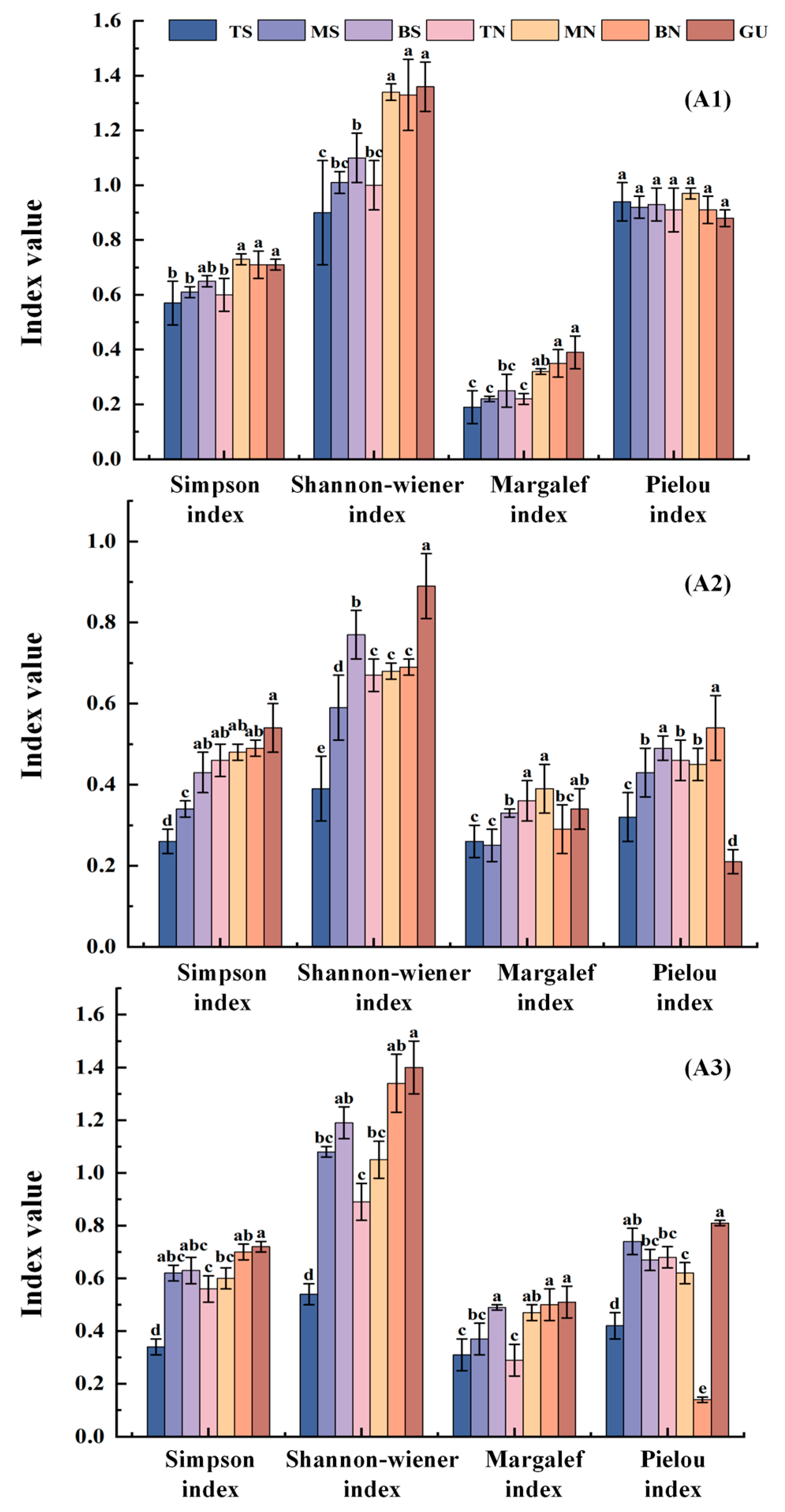
3.2. Characteristics of Changes in Vegetation Community Diversity Under Different Microtopographic Conditions
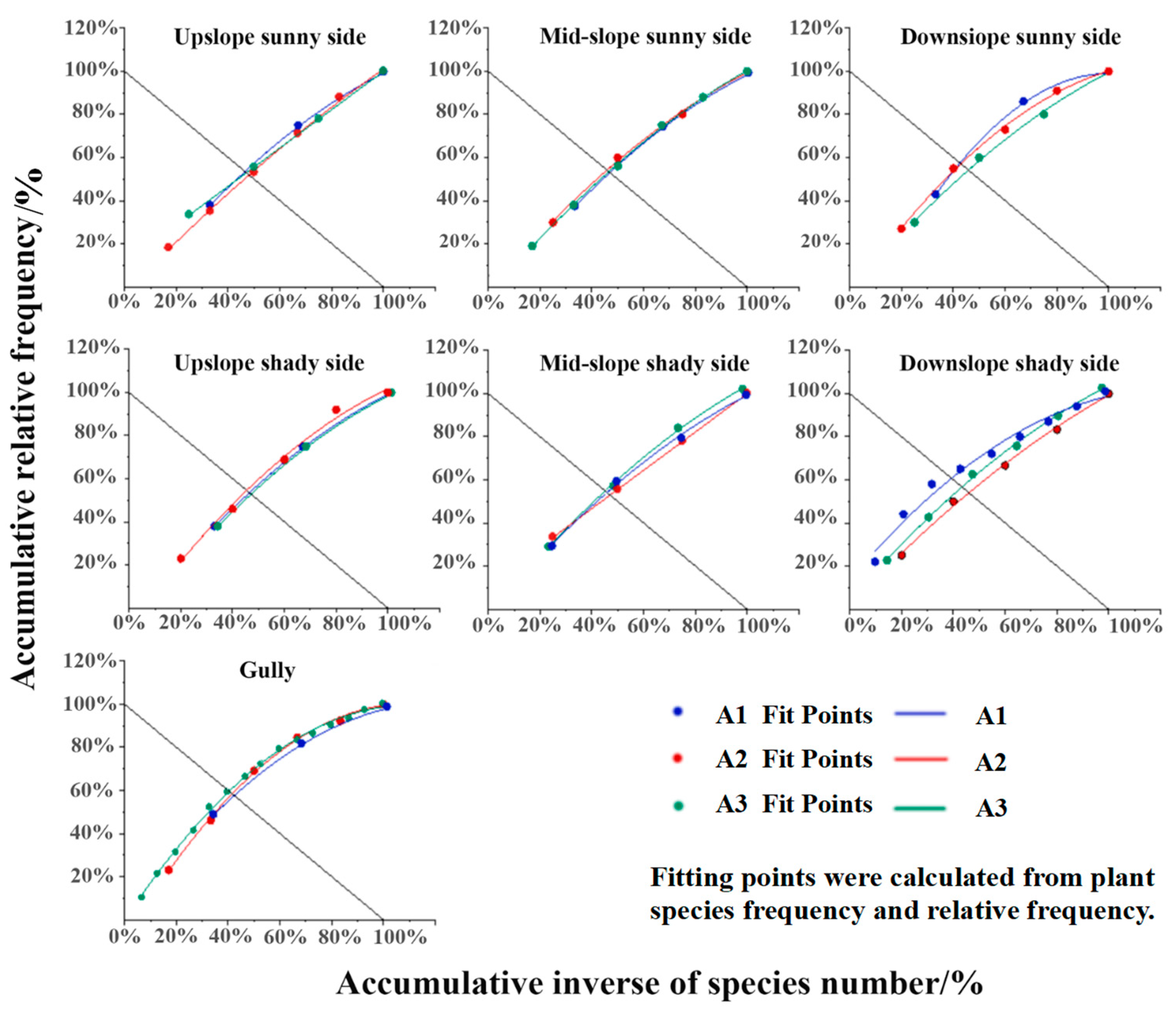
3.3. Characteristics of Vegetation Community Stability Changes Under Different Microtopographic Conditions
3.4. Correlation Analysis Between Soil Moisture and Vegetation Community Diversity and Stability
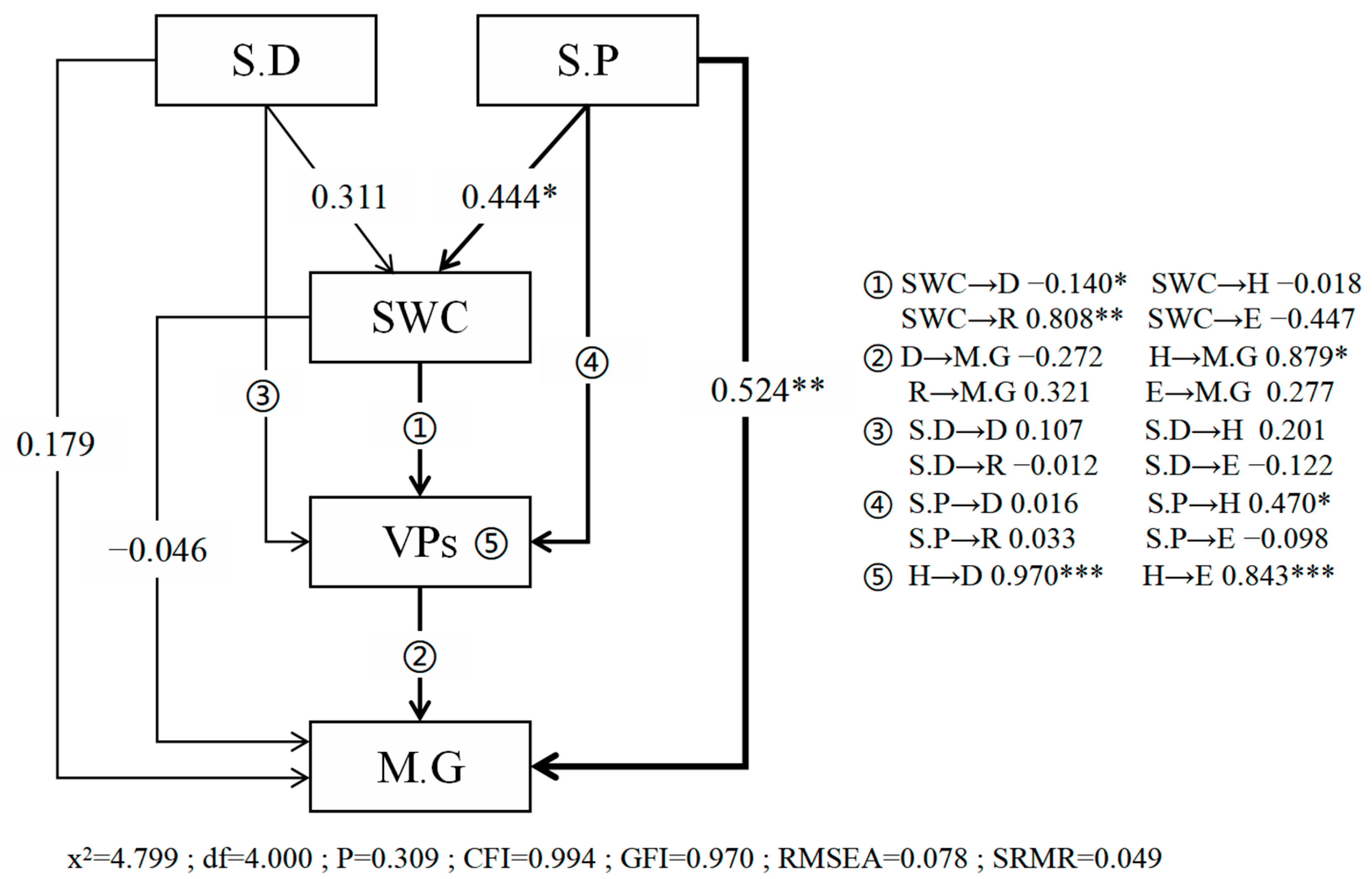
3.5. Microtopography-Soil Moisture-Vegetation Community Relationship Analysis
4. Discussion
4.1. The Impact of Microtopography on Soil Moisture
4.2. The Impact of Microtopography on Vegetation Community Diversity and Stability
4.3. The Relationship Between Microtopography, Soil Moisture, and Vegetation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- He, J.; Shi, X.; Fu, Y. Identifying vegetation restoration effectiveness and driving factors on different micro-topographic types of hilly Loess Plateau: From the perspective of ecological resilience. J. Environ. Manag. 2021, 289, 112562. [Google Scholar] [CrossRef]
- Ju, J.; Masek, J.G. The vegetation greenness trend in Canada and US Alaska from 1984–2012 Landsat data. Remote Sens. Environ. 2016, 176, 1–16. [Google Scholar] [CrossRef]
- Huang, L. Advances and perspectives on soil water research in China’s Loess Plateau. Earth-Sci. Rev. 2019, 199, 102962. [Google Scholar] [CrossRef]
- Alarcón, R.; Hernández-Plaza, E.; Navarrete, L.; Sánchez, M.J.; Escudero, A.; Hernanz, J.L.; Sánchez-Giron, V.; Sánchez, A. Effects of no-tillage and non-inversion tillage on weed community diversity and crop yield over nine years in a Mediterranean cereal-legume cropland. Soil Tillage Res. 2018, 179, 54–62. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of plant diversity to ecosystems: Immediate, filterand founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Liao, C.; Liu, B.; Xu, Y.; Li, Y.; Li, H. Effect of topography and protecting barriers on revegetation of sandy land, Southern Tibetan Plateau. Sci. Rep. 2019, 9, 6501. [Google Scholar] [CrossRef]
- Sun, C.; Chai, Z.; Liu, G.; Xue, S. Changes in species diversity patterns and spatial heterogeneity during the secondary succession of grassland vegetation on the Loess Plateau, China. Front. Plant Sci. 2017, 8, 1465. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.J.; Royer, P.D.; Cobb, N.S.; Breshears, D.D.; Ford, P.L. Precipitation thresholds and drought-induced tree die-off: Insights from patterns of Pinus edulis mortality along an environmental stress gradient. New Phytol. 2013, 200, 413–421. [Google Scholar] [CrossRef]
- Kong, W.; Yao, Y.; Zhao, Z.; Qin, X.; Zhu, H.; Wei, X.; Su, M.; Wang, Z.; Bao, K.; Su, M. Effects of vegetation and slope aspect on soil nitrogen mineralization during the growing season in sloping lands of the Loess Plateau. Catena 2019, 172, 753–763. [Google Scholar] [CrossRef]
- Fang, J.-Y.; Shen, Z.-H.; Cui, H.-T. Ecological characteristics of mountains and re-search issues of mountain ecology. Biodivers Sci. 2004, 12, 10–19. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Carscadden, K.; Mirotchnick, N. Beyond species: Functional diversity and the maintenance of ecological processes and services. J. Appl. Ecol. 2011, 48, 1079–1087. [Google Scholar]
- Sun, W.; Shao, Q.; Liu, J.; Zhai, J. Assessing the effects of land use and topography on soil erosion on the Loess Plateau in China. Catena 2014, 121, 151–163. [Google Scholar] [CrossRef]
- Gao, X.; Zhao, X.; Wu, P.; Brocca, L.; Zhang, B. Effects of large gullies on catchment-scale soil moisture spatial behaviors: A case study on the Loess Plateau of China. Geoderma 2016, 261, 1–10. [Google Scholar] [CrossRef]
- Qiao, H.; Li, Q.; He, G.; Ji, T.; Yang, D.Y.; Wen, T.; Guan, W.H.; Liu, Z.G.; Yang, J.Y.; Liu, X.N. Biodiversity and community stability of mountain meadow vegetation in response to slope orientation. J. Grassl. Sci. 2022, 30, 2173–2181. [Google Scholar] [CrossRef]
- Lei, S.; Liao, L.; Wang, J.; Zhang, L.; Ye, Z.; Liu, G.; Zhang, C. Relationship between plant diversity and Godron community stability in alpine grassland and its environmental drivers. J. Grassl. Sci. 2023, 32, 1–12. [Google Scholar]
- Zhao, W.; Zhang, Y.; Zhu, Q. Effects of microtopography on tree growth characteristic in loess slope of northern Shaanxi. J. Basic Sci. Eng. 2014, 22, 69. [Google Scholar]
- Jiang, C.; Zhang, H.; Wang, X.; Feng, Y.; Labzovskii, L. Challenging the land degradation in China’s Loess Plateau: Benefits, limitations, sustainability, and adaptive strategies of soil and water conservation. Ecol. Eng. 2019, 127, 135–150. [Google Scholar] [CrossRef]
- Yu, Y.; Zhao, W.; Martinez-Murillo, J.F.; Pereira, P. Loess Plateau: From degradation to restoration. Sci. Total Environ. 2020, 738, 140206. [Google Scholar]
- Yang, L.; Wei, W.; Chen, L.; Jia, F.; Mo, B. Spatial variations of shallow and deep soil moisture in the semi-arid Loess Plateau, China. Hydrol. Earth Syst. Sci. 2012, 16, 3199–3217. [Google Scholar]
- Liu, Y.; Du, J.; Xu, X.; Kardol, P.; Hu, D. Microtopography-induced ecohydrological effects alter plant community structure. Geoderma 2020, 362, 114119. [Google Scholar]
- Yao, S.; Li, H. Agricultural productivity changes induced by the sloping land conversion program: An analysis of Wuqi county in the Loess Plateau region. Environ. Manag. 2010, 45, 541–550. [Google Scholar]
- Lin, Y.; Yao, S. Impact of the Sloping Land Conversion Program on rural household income: An integrated estimation. Land Use Policy 2014, 40, 56–63. [Google Scholar]
- Yu, B.; Liu, G.; Liu, Q.; Wang, X.; Feng, J.; Huang, C. Soil moisture variations at different topographic domains and land use types in the semi-arid Loess Plateau, China. Catena 2018, 165, 125–132. [Google Scholar]
- Cui, M.; Zhou, R.; Zheng, C.; Wei, J.; Wan, J.; Zhao, Y.; Wen, Z. Responses of Aboveground Biomass and Soil Moisture and Nutrients to Microtopographic Changes in Grassland in the Loess Hilly Region. Res. Soil Water Conserv. 2024, 31, 18–26. [Google Scholar]
- Huffman, R.L.; Fangmeier, D.D.; Elliot, W.J.; Workman, S.R.; Schwab, G.O. Soil and Water Conservation Engineering; American Society of Agricultural and Biological Engineers: St. Joseph, MI, USA, 2013. [Google Scholar]
- Chen, Y.; Jiao, J.; Yan, X.; Li, J.; Vanmaercke, M.; Wang, N. Response of gully morphology and density to the spatial and rainy-season monthly variation of rainfall at the regional scale of the Chinese Loess Plateau. Catena 2024, 236, 107773. [Google Scholar]
- Chase, J.M.; Knight, T.M. Scale-dependent effect sizes of ecological drivers on biodiversity: Why standardised sampling is not enough. Ecol. Lett. 2013, 16, 17–26. [Google Scholar]
- Mallarino, A.P.; Wittry, D.J. Efficacy of grid and zone soil sampling approaches for site-specific assessment of phosphorus, potassium, pH, and organic matter. Precis. Agric. 2004, 5, 131–144. [Google Scholar]
- Shannon, C.E. The mathematical theory of communication. MD Comput. 1997, 14, 306–317. [Google Scholar]
- Margalef, R. Information Theory in Ecology; Real Academia de Ciencias y Artes de Barcelona: Barcelona, Spain, 1973. [Google Scholar]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Theor. Biol. 1966, 13, 131–144. [Google Scholar]
- Li, X.; Yang, W.; Sun, T.; Su, L. Framework of multidimensional macrobenthos biodiversity to evaluate ecological restoration in wetlands. Environ. Res. Lett. 2019, 14, 054003. [Google Scholar] [CrossRef]
- Li, S.; Sun, T.; Yang, W.; Cui, B.; Yin, X. A biodiversity evaluation framework for restoration of aquatic macrophyte communities in shallow lakes driven by hydrological process management. Hydrol. Process. 2021, 35, e13983. [Google Scholar] [CrossRef]
- Godron, M. Some aspecets of heterogeneity in grasslands of Cantal. Statis. Ecol. 1972, 3, 397–415. [Google Scholar]
- Zheng, Y. Comparison of methods for studying stability of forest community. Sci. Silvae Sin. 2000, 36, 28–32. [Google Scholar] [CrossRef]
- Li, L.; Jiang, E.; Yin, H.; Wu, K.; Dong, G. Ultrashort-term responses of riparian vegetation restoration to adjacent cycles of ecological water conveyance scheduling in a hyperarid endorheic river basin. J. Environ. Manag. 2022, 320, 115803. [Google Scholar] [CrossRef]
- Jin, S.S.; Zhang, Y.Y.; Zhou, M.L.; Dong, X.M.; Chang, C.H.; Wang, T.; Yan, D.F. Interspecific association and community stability of tree species in natural secondary forests at different altitude gradients in the southern Taihang Mountains. Forests 2022, 13, 373. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, W.; Wang, L.; Liu, Y.; Liu, Y.; Feng, Q. Relationship between soil water content and soil particle size on typical slopes of the Loess Plateau during a drought year. Sci. Total Environ. 2019, 648, 943–954. [Google Scholar] [CrossRef]
- Miyasaka, T.; Okuro, T.; Miyamori, E.; Zhao, X.; Takeuchi, K. Effects of different restoration measures and sand dune topography on short-and long-term vegetation restoration in northeast China. J. Arid Environ. 2014, 111, 1–6. [Google Scholar] [CrossRef]
- Gao, R.; Zhao, Y.G.; Liu, X.F.; Li, W.Y.; Du, Y.J.; Zhang, X.X. Effects of stand age and slope position of Caragana korshinskii plantations on soil aggregate stability in the loess hilly region. Acta Ecol. Sin. 2020, 40, 2964–2974. [Google Scholar] [CrossRef]
- Luo, J.; Zheng, Z.; Li, T.; He, S.; Zhang, X.; Huang, H.; Wang, Y. Quantifying the contributions of soil surface microtopography and sediment concentration to rill erosion. Sci. Total Environ. 2021, 752, 141886. [Google Scholar] [CrossRef]
- Tian, X.; Gao, K.; Zhang, L.J.; Yu, Y.Q.; Han, G.D. Effect of slope position on spatial distribution of soil water and vegetation in sandy land. J. Soil Water Conserv. 2015, 35, 12–16. [Google Scholar] [CrossRef]
- Zhang, L.; Yu, Y.; Gao, K. Response of plant functional groups and their diversity to microtopographic changes in sandy areas. J. Grassl. 2015, 23, 41–46. [Google Scholar] [CrossRef]
- Coverdale T, C.; Davies A, B. Unravelling the relationship between plant diversity and vegetation structural complexity: A review and theoretical framework. J. Journal of Ecology. 2023, 111, 1378–1395. [Google Scholar]
- Jahantigh, M.; Pessarakli, M. Causes and effects of gully erosion on agricultural lands and the environment. Commun. Soil Sci. Plant Anal. 2011, 42, 2250–2255. [Google Scholar]
- Tong, X.; Wang, K.; Brandt, M.; Yue, Y.; Liao, C.; Fensholt, R. Assessing future vegetation trends and restoration prospects in the karst regions of southwest China. Remote Sens. 2016, 8, 357. [Google Scholar] [CrossRef]
- Gao, X.; Wu, P.; Zhao, X.; Shi, Y.; Wang, J.; Zhang, B. Soil moisture variability along transects over a well-developed gully in the Loess Plateau, China. Catena 2011, 87, 357–367. [Google Scholar] [CrossRef]
- Chesson, P.; Gebauer, R.L.E.; Schwinning, S.; Huntly, N.; Wiegand, K.; Ernest, M.S.K.; Sher, A.; Novoplansky, A.; Weltzin, J.F. Resource pulses, species interactions, and diversity maintenance in arid and semi-arid environments. Oecologia 2004, 141, 236–253. [Google Scholar]
- Wang, J.; Zhao, W.; Zhang, X.; Liu, Y.; Wang, S.; Liu, Y. Effects of reforestation on plant species diversity on the Loess Plateau of China: A case study in Danangou catchment. Sci. Total Environ. 2019, 651, 979–989. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Deng, L.; Yan, W.M.; Shangguan, Z.P. Interaction of soil water storage dynamics and long-term natural vegetation succession on the Loess Plateau, China. Catena 2016, 137, 52–60. [Google Scholar] [CrossRef]
- Yu, B.; Liu, G.; Liu, Q.; Feng, J.; Wang, X.; Han, G.; Huang, C. Effects of micro-topography and vegetation type on soil moisture in a large gully on the Loess Plateau of China. Hydrol. Res. 2018, 49, 1255–1270. [Google Scholar]
- Li, C.; Yang, X.; Chen, Y.; Zhou, X.; Li, S.; Cui, X.; Tang, G. Assessing the influence of geo-environmental factors on discontinuous gully erosion at regional scales: A case study of spoon gullies on the loess plateau of China. J. Environ. Manag. 2025, 373, 123940. [Google Scholar] [CrossRef] [PubMed]
- Fan, F.; Gu, X.; Luo, J.; Zhang, B.; Liu, H.; Yang, H.; Wang, L. Identification of gully erosion activity and its influencing factors: A case study of the Sunshui River Basin. PLoS ONE 2024, 19, e0309672. [Google Scholar] [CrossRef] [PubMed]
- de Mazancourt, C.; Isbell, F.; Larocque, A.; Berendse, F.; De Luca, E.; Grace, J.B.; Haegeman, B.; Polley, H.W.; Roscher, C.; Schmid, B.; et al. Predicting ecosystem stability from community composition and biodiversity. Ecol. Lett. 2013, 16, 617–625. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Zhang, C.; Meng, F.-R.; Bourque, C.P.-A.; Huang, Z.; Li, X. Impacts of re-vegetation on soil water dynamics in a semiarid region of Northwest China. Sci. Total Environ. 2024, 911, 168496. [Google Scholar] [CrossRef]
- Brandt, M.; Hiernaux, P.; Rasmussen, K.; Mbow, C.; Kergoat, L.; Tagesson, T.; Ibrahim, Y.Z.; Wélé, A.; Tucker, C.J.; Fensholt, R. Assessing woody vegetation trends in Sahelian drylands using MODIS based seasonal metrics. Remote Sens. Environ. 2016, 183, 215–225. [Google Scholar] [CrossRef]
- Tian, Q.; Zhang, X.; Wang, M.; Wang, H.J.; Yi, H.J.; He, J.; Xu, X.M.; Lyu, D.; Sun, W.N. Relationship between plant compositional diversity and community stability with soil aggregates under vegetation succession. China Environ. Sci. 2022, 42, 5885–5895. [Google Scholar] [CrossRef]
- Jiao, F.; Wen, Z.M.; An, S.S. Changes in soil properties across a chronosequence of vegetation restoration on the Loess Plateau of China. Catena 2011, 86, 110–116. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area Number | Sample Plot | Slope Micro-Topography | Longitude and Latitude | Elevation/m | Quadrat Vegetation Type | Dominant Species * |
|---|---|---|---|---|---|---|
| A1 | 1 | TS | 107°84′37″ E, 37°38′63″ N | 1797 | grass | Linum perenne L. Stipa capillata L. |
| 2 | MS | 107°84′39″ E, 37°38′58″ N | 1771 | grass | Linum perenne L. Stipa capillata L. Artemisia frigida Willd. | |
| 3 | BS | 107°84′39″ E, 37°38′58″ N | 1743 | grass | Linum perenne L. Stipa capillata L. Stipa bungeana Trin. | |
| 4 | TN | 107°84′53″ E, 37°38′71″ N | 1788 | grass | Linum perenne L. Stipa capillata L. | |
| 5 | MN | 107°85′89″ E, 37°38′90″ N | 1691 | grass | Stipa capillata L. Artemisia frigida Willd. Potentilla acaulis L. | |
| 6 | BN | 107°85′86″ E, 37°38′95″ N | 1658 | grass | Caragana Korshinskii Kom. Stipa capillata L. Cymbaria mongolica Maxim. | |
| 7 | GU | 107°85′72″ E, 37°38′38″ N | 1648 | grass | Artemisia frigida Willd. Stipa capillata L. Lespedeza tomentosa (Thunb.) Siebold ex Maxim. | |
| A2 | 8 | TS | 108°14′12″ E, 37°12′76″ N | 1549 | arbor-grass | Prunus sibirica L. (arbor) Stipa capillata L. (grass) |
| 9 | MS | 108°14′02″ E, 37°12′75″ N | 1503 | arbor-grass | Prunus sibirica L. (arbor) Stipa capillata L. (grass) Artemisia stechmanniana Besser.(grass) | |
| 10 | BS | 108°19′41″ E, 37°47′05″ N | 1472 | arbor-grass | Prunus sibirica L. (arbor) Pyrus betulifolia Bunge.(arbor) Stipa capillata L. (grass) | |
| 11 | TN | 108°14′06″ E, 37°12′56″ N | 1528 | arbor-shrub-grass | Prunus sibirica L. (arbor) Hippophae rhamnoides L. (shrub) Stipa capillata L. (grass) | |
| 12 | MN | 108°14′07″ E, 37°12′58″ N | 1504 | arbor-shrub-grass | Prunus sibirica L. (arbor) Hippophae rhamnoides L. (shrub) Artemisia stechmanniana Besser.(grass) Stipa capillata L. (grass) | |
| 13 | BN | 108°14′08″ E, 37°12′54″ N | 1476 | arbor-shrub-grass | Populus simonii Carrière Prunus sibirica L. (arbor) Caragana korshinskii Kom.(shrub) Stipa capillata L. (grass) | |
| 14 | GU | 108°14′08″ E, 37°12′55″ N | 1465 | arbor-shrub-grass | Populus simonii.(arbor) Carrière Prunus sibirica L. (arbor) Caragana korshinskii Kom.(shrub) Stipa capillata L. (grass) | |
| A3 | 15 | TS | 109°66′57″ E, 36°11′43″ N | 1246 | arbor-grass | Pinus tabuliformis Carrière.(arbor) Acer tataricum L. (arbor) Allium ramosum L. (grass) |
| 16 | MS | 109°66′60″ E, 36°11′41″ N | 1228 | arbor-grass | Pinus tabuliformis Carrière.(arbor) Prunus sibirica L. (arbor) Allium ramosum L. (grass) Phtheirospermum japonicum (Thunb.) Kanitz.(grass) | |
| 17 | BS | 109°66′61″ E, 36°11′39″ N | 1212 | arbor-grass | Acer tataricum L. (arbor) Allium ramosum L. (grass) Rubia cordifolia L. (grass) | |
| 18 | TN | 109°65′89″ E, 36°11′27″ N | 1234 | arbor-grass | Pinus tabuliformis Carrière.(arbor) Quercus wutaishansea Mary.(arbor) Allium ramosum L. (grass) | |
| 19 | MN | 109°65′86″ E, 36°11′22″ N | 1215 | arbor-shrub-grass | Pinus tabuliformis Carrière.(arbor) Quercus wutaishansea Mary.(arbor) Allium ramosum L. (grass) Rubus parvifolius L. (grass) | |
| 20 | BN | 109°65′85″ E, 36°11′26″ N | 1199 | arbor-shrub-grass | Pinus tabuliformis Carrière.(arbor) Acer tataricum L. (arbor) Allium ramosum L. (grass) Chrysanthemum chanetii H. Lév.(grass) | |
| 21 | GU | 109°65′85″ E, 36°11′25″ N | 1193 | arbor-shrub-grass | Tamarix chinensis Lour.(arbor) Rubus parvifolius L. (grass) Amethystea caerulea L. (grass) |
| Vegetation Parameters | Formula |
|---|---|
| Shannon–Wiener index | H = −) |
| Margalef index | R = (S − 1)/lnN |
| Simpson index | D = 1 − |
| Pielou index | E = H/lnS |
| Area Number | Microtopography | Fitting Curve | R2 | Coordinates of the Intersection Point with the Line y = 100 − x | Euclidean Distance from the Stable Point of the Community (20, 80) |
|---|---|---|---|---|---|
| TS | y = −0.09x2 + 1.57x − 0.49 | 0.9991 | (46.89,53.11) | 38.18 | |
| MS | y = −0.09x2 + 1.57x − 0.49 | 0.9991 | (46.49,53.51) | 36.77 | |
| BS | y = −0.26x2 + 2.49x − 1.24 | 0.9989 | (43.60,56.40) | 33.94 | |
| A1 | TN | y = −0.09x2 + 1.57x − 0.49 | 0.9991 | (46.49,53.51) | 36.77 |
| MN | y = −0.02x2 + 1.42x − 0.40 | 0.9981 | (45.45,54.55) | 35.36 | |
| BN | y = 0.11x2 + 1.43x − 0.57 | 0.9833 | (40.47,59.53) | 28.28 | |
| GU | y = 0.03x2 + 1.63x − 0.67 | 0.9985 | (42.56,57.44) | 32.53 | |
| TS | y = −0.03x2 + 1.24x − 0.20 | 0.9979 | (48.54,51.45) | 41.01 | |
| MS | y = −0.02x2 + 1.42x − 0.40 | 0.9944 | (45.45,54.55) | 35.36 | |
| BS | y = −0.04x2 + 1.72x − 0.68 | 0.9969 | (40.65,59.35) | 29.70 | |
| A2 | TN | y = −0.09x2 + 1.64x − 0.54 | 0.9874 | (43.52,56.48) | 33.94 |
| MN | y = 0.11x2 + 0.89x − 0.004 | 0.9998 | (47.06,52.94) | 38.18 | |
| BN | y = 0.02x2 + 1.27x − 0.30 | 0.9953 | (46.02,53.98) | 36.77 | |
| GU | y = −0.09x2 + 1.99x − 0.91 | 0.9970 | (41.94,58.06) | 31.11 | |
| TS | y = −0.03x2 + 1.24x − 0.20 | 0.9987 | (48.54,51.46) | 41.01 | |
| MS | y = −0.02x2 + 1.42x − 0.40 | 0.9981 | (45.83,54.17) | 36.77 | |
| BS | y = −0.02x2 + 1.41x − 0.39 | 0.9986 | (45.45,54.55) | 35.36 | |
| A3 | TN | y = −0.09x2 + 1.57x − 0.49 | 0.9991 | (46.89,53.11) | 38.18 |
| MN | y = −0.09x2 + 1.64x − 0.54 | 0.9874 | (44.44,55.55) | 33.94 | |
| BN | y = −0.1x2 + 1.86x − 0.75 | 0.9977 | (43.49,56.51) | 32.53 | |
| GU | y = −0.01x2 + 1.82x − 0.82 | 0.9983 | (40.50,59.50) | 29.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, L.; Liu, Y.; Liu, J.; Kang, H.; Liu, Z.; Tuo, F.; Gan, S.; Ren, Y.; Yi, C.; Hu, G. Microtopography Affects the Diversity and Stability of Vegetation Communities by Regulating Soil Moisture. Water 2025, 17, 1012. https://doi.org/10.3390/w17071012
Han L, Liu Y, Liu J, Kang H, Liu Z, Tuo F, Gan S, Ren Y, Yi C, Hu G. Microtopography Affects the Diversity and Stability of Vegetation Communities by Regulating Soil Moisture. Water. 2025; 17(7):1012. https://doi.org/10.3390/w17071012
Chicago/Turabian StyleHan, Lei, Yang Liu, Jie Liu, Hongliang Kang, Zhao Liu, Fengwei Tuo, Shaoan Gan, Yuxuan Ren, Changhua Yi, and Guiming Hu. 2025. "Microtopography Affects the Diversity and Stability of Vegetation Communities by Regulating Soil Moisture" Water 17, no. 7: 1012. https://doi.org/10.3390/w17071012
APA StyleHan, L., Liu, Y., Liu, J., Kang, H., Liu, Z., Tuo, F., Gan, S., Ren, Y., Yi, C., & Hu, G. (2025). Microtopography Affects the Diversity and Stability of Vegetation Communities by Regulating Soil Moisture. Water, 17(7), 1012. https://doi.org/10.3390/w17071012