
Extreme Weather Events and Climate Variability Provide a Lens to How Shallow Lakes May Respond to Climate Change

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Study Lakes
2.2. Sampling and Laboratory Methods
2.2.1. Lake Okeechobee
2.2.2. Lake George
2.2.3. Lake Taihu
2.3. Data Analysis
2.3.1. Lake Okeechobee
2.3.2. Lake George
2.3.3. Lake Taihu
3. Results
3.1. Lake Okeechobee
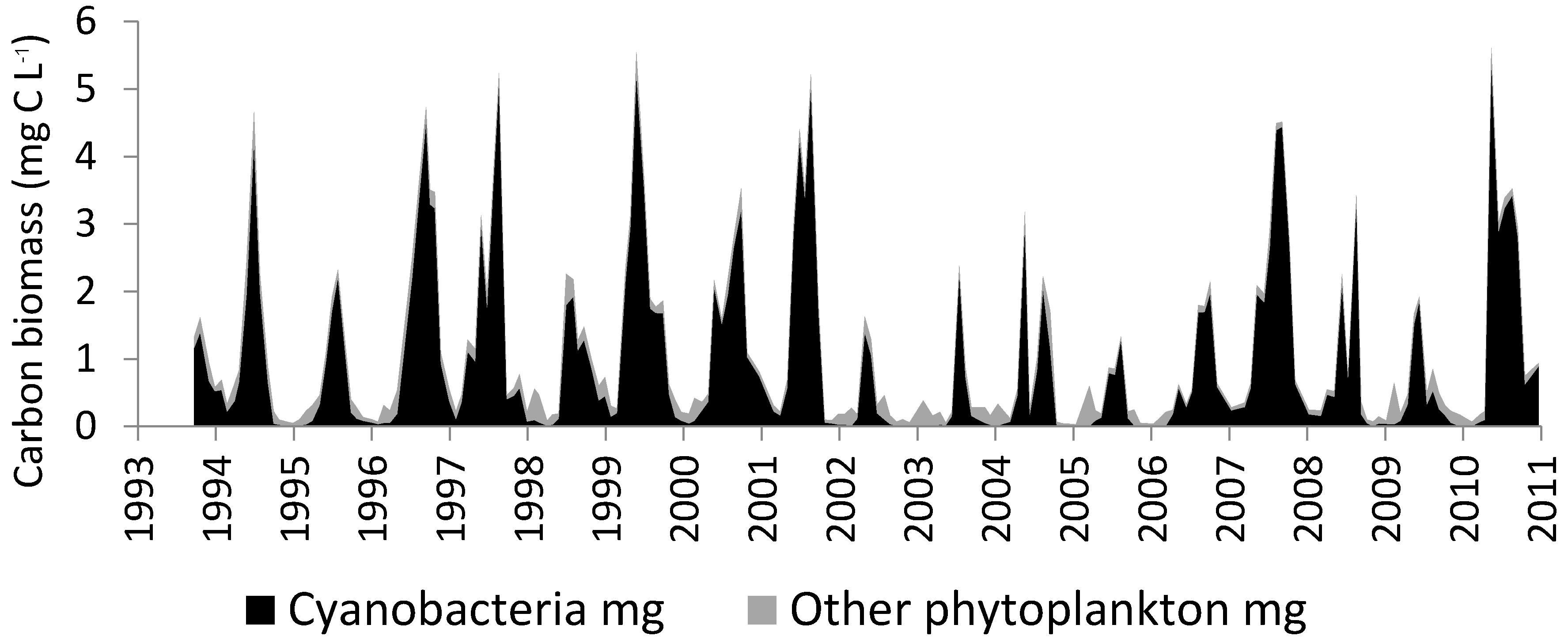
3.2. Lake George
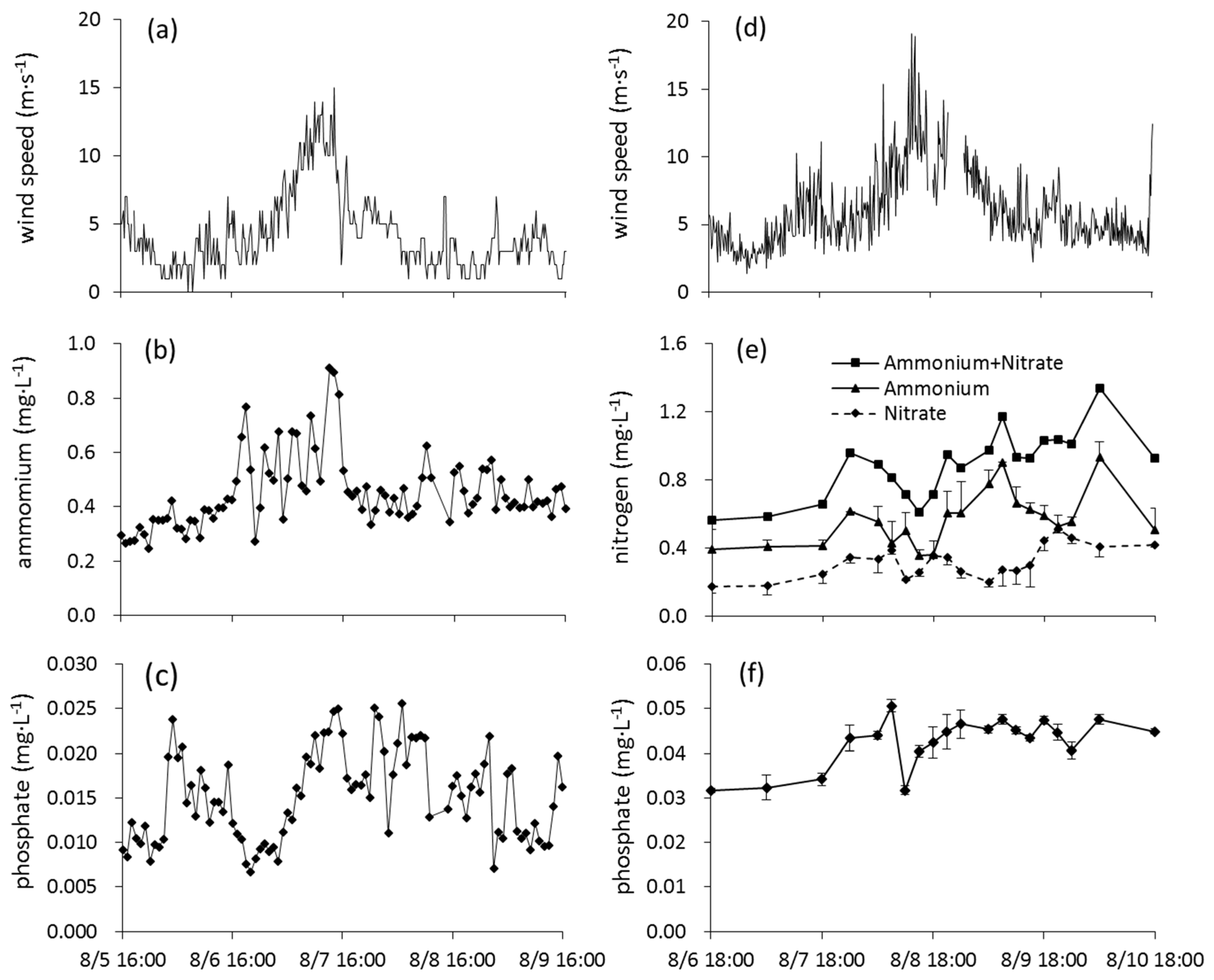
3.3. Lake Taihu
4. Discussion
- Is the lake located in a region that experiences tropical cyclones? If yes, this increases the risk of highly disruptive impacts from cyclones that likely will be exacerbated in a future with cyclones of greater intensity due to global warming. The data presented here indicate that all of the lakes are resilient to the effects of tropical cyclones and recover on time scales of months. But what if in the future those storms are more intense or they follow one-another in close sequence? That could lead to a situation where there is insufficient time for the system to recover before being hit by another storm (Figure 11).
- Is the lake located in a region where oceanic cycles and their teleconnections result in year-to-year or decadal variation in rainfall, which applies to both Lakes Okeechobee and George? If yes, and those ocean cycles are changed as a result of climate change [23], it could markedly alter the dynamics of the lake ecosystems and their long-term response to management actions.
- Does the lake have a high dynamic ratio, which indicates a large surface area relative to mean depth, which characterize all three lakes in this study? If yes, then for a given amount of wind action on the water surface, a greater amount of energy will shear the sediments, making these lakes more prone to disruption by wind events.
- Do the sediments contain a high proportion of inorganic solids? If yes, these lakes may recover more quickly because those inorganic solids will block light, limiting phytoplankton development while also not supporting bacterial growth. This appears to be what happened in Lake Okeechobee, especially after the impact of a third hurricane.
- Are concentrations of TP and TN high in the sediments? If yes, then there is a source of nutrients that can be carried into the water column to stimulate algal and bacterial production. This occurred in Lake Taihu, where dissolved nutrients were released from the sediments into the overlying water during the strong wind period, followed by a bloom period. It also occurred in Lake Okeechobee, and it likely is what stimulated the long-term dominance of cyanobacteria blooms in Lake Apopka, Florida [48].
- Are cyanobacteria, including bloom-forming species, dominant, as observed in all three lakes in this study? If yes, then it is more likely, considering the factors listed above, that extreme events or changes in water depth and global temperature might stimulate CyanoHABs. Here, we have considered the effects of global climate change on the management of shallow lakes only from the perspective of wind and flushing events. There also is evidence that, in a warmer future, cyanobacteria will be favored over other algae in lakes based on optimal growth rates at elevated temperatures and stronger vertical stratification [56] and that lakes with high TP and TN concentrations that undergo warming could experience a non-linear and unpredictable rise in bloom frequency [6]. When combined with the above-mentioned factors, it may render some lakes beyond the reach of any currently available rehabilitation methods, and it supports the view that nutrient reduction is critical now, rather than later when these various effects of global climate change amplify nutrient effects [57,58].
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F. (Ed.) Cambridge University Press: Cambridge, UK, 2014.
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystem structure. Nature 2011, 413, 591–596. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W. Nuisance phytoplankton blooms in coastal, estuarine, and inland waters. Limnol. Oceanogr. 1988, 33, 823–847. [Google Scholar] [CrossRef]
- Zhu, M.; Paerl, H.W.; Zhu, G.; Wu, T.; Li, W.; Shi, K.; Zhao, L.; Zhang, Y.; Qin, B.; Caruso, A.M. The role of tropical cyclones in stimulating Cyanobacteria (Microcystis spp.) blooms in hypertrophic Lake Taihu, China. Harmful Algae 2014, 39, 310–321. [Google Scholar] [CrossRef]
- Jackson, L.J.; Lauridsen, T.L.; Søndergaard, M.; Jeppesen, E. A comparison of shallow Danish and Canadian lakes and implications for climate change. Freshw. Biol. 2007, 52, 1782–1792. [Google Scholar] [CrossRef]
- Kosten, S.; Huszar, V.L.M.; Bécares, E.; Costa, L.S.; Van Donk, E.; Hansson, L.A.; Jeppesen, E.; Kruk, C.; Lacerot, G.; Mazzeo, N.; et al. Warmer climate boosts cyanobacterial dominance in shallow lakes. Glob. Chang. Biol. 2012, 18, 118–126. [Google Scholar] [CrossRef]
- Moss, B.; Stephen, D.; Balayla, D.M.; Bécares, E.; Collings, S.E.; Frenández-Aláez, C.; Frenández-Aláez, M.; Ferriol, C.; García, P.; Gomá, J.; et al. Continental-scale patterns of nutrient and fish effects on shallow lakes; synthesis of a pan-European mesocosm experiment. Freshw. Biol. 2004, 49, 1633–1649. [Google Scholar] [CrossRef]
- Jeppesen, E.; Meerhoff, M.; Davidson, T.A.; Trolle, D.; Søndergaard, M.; Lauridsen, T.L.; Beklioğlu, M.; Brucet, S.; Bolta, P.; González-Bergonzoni, I.; et al. Climate change impacts on lakes: An integrated ecological perspective based on a multi-faceted approach, with special focus on shallow lakes. J. Limnol. 2014, 73, 84–107. [Google Scholar] [CrossRef]
- Shurin, J.B.; Clasen, J.L.; Hamish, S.G.; Pavel, K.; Thompson, P.L. Warming shifts top-down and bottom-up control of pond food web structure and function. Phil. Trans. R. Soc. B 2012, 367, 3008–3017. [Google Scholar] [CrossRef] [PubMed]
- Emanuel, K. Increasing destructiveness of tropical cyclones over the past 30 years. Nature 2005, 436, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.R.; Ji, Z.G. Calibration and verification of a spectral wind-wave model for Lake Okeechobee. Ocean Eng. 2001, 28, 571–584. [Google Scholar] [CrossRef]
- Havens, K.E.; Beaver, J.R.; Casamatta, D.; East, T.; James, R.T.; McCormick, P.; Phlips, E.J.; Rodusky, A.J. Hurricane effects on the planktonic food web of a large subtropical lake. J. Plankton Res. 2011, 33, 1081–1094. [Google Scholar] [CrossRef]
- Havens, K.E.; Jin, K.R.; Rodusky, A.J.; Sharfstein, B.; Brady, M.A.; East, T.L.; Iricanin, N.; James, R.T.; Harwell, M.C.; Steinman, A.D. Hurricane effects on a shallow lake ecosystem and its response to a controlled manipulation of water level. Sci. World 2001, 1, 44–70. [Google Scholar] [CrossRef] [PubMed]
- Beaver, J.R.; Casamatta, D.A.; East, T.L.; Havens, K.E.; Rodusky, A.J.; James, R.T.; Tausz, C.E.; Buccier, K.M. Extreme weather events influence the phytoplankton community structure in a large lowland subtropical lake (Lake Okeechobee, Florida, USA). Hydrobiologia 2013, 709, 213–226. [Google Scholar] [CrossRef]
- Paerl, H.W.; Christian, R.R.; Bales, J.D.; Peierls, B.L.; Hall, N.S.; Joyner, A.R.; Riggs, S.R. Assessing the response of the Pamlico Sound, North Carolina, USA to human and climatic disturbances: Management implications. In Critical Habitats of Environmental Change; Kennish, M., Paerl, H., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 17–42. [Google Scholar]
- Hall, N.S.; Paerl, H.W.; Peierls, B.L.; Whipple, A.C.; Rossignol, K.L. Effects of climatic variability on phytoplankton biomass and community structure in the eutrophic, microtidal, New River Estuary, North Carolina, USA. Est. Coast. Shelf Sci. 2013, 117, 70–82. [Google Scholar] [CrossRef]
- Paerl, H.W.; Hall, N.S.; Peierls, B.L.; Rossignol, K.L.; Joyner, A.R. Hydrologic variability and its control of phytoplankton community structure and function in two shallow, coastal, lagoonal ecosystems: The Neuse and New River Estuaries, North Carolina, USA. Estuaries Coasts 2014, 37, 31–45. [Google Scholar] [CrossRef]
- Cόzar, A.; Bruno, M.; Bergamino, N.; Ubeda, B.; Bracchini, L.; Dattilo, A.M.; Loiselle, S.A. Basin-scale control on the phytoplankton biomass in Lake Victoria, Africa. PLoS ONE 2012, 7, e29962. [Google Scholar] [CrossRef] [PubMed]
- Van Cleave, K.; Lenters, J.D.; Wang, J.; Verhamme, E.M. A regime shift in Lake Superior ice cover, evaporation, and water temperature following the warm El Niño winter of 1997–1998. Limnol. Oceanogr. 2014, 59, 1889–1898. [Google Scholar] [CrossRef]
- Dokulil, M.T.; Jagsch, A.; George, G.D.; Anneville, O.; Jankowski, T.; Wahl, B.; Blenckner, T.; Teubner, K. Twenty years of spatially coherent deepwater warming in lakes across Europe related to the North Atlantic Oscillation. Limnol. Oceanogr. 2006, 51, 2787–2793. [Google Scholar] [CrossRef]
- Paerl, H.W.; Richards, R.C.; Leonard, R.L.; Goldman, C.R. Seasonal nitrate cycling as evidence for complete vertical mixing in Lake Tahoe, California-Nevada. Limnol. Oceanogr. 1975, 20, 1–8. [Google Scholar] [CrossRef]
- Abtew, W.; Trimble, P. El Niño-Southern Oscillation link to south Florida hydrology and water management applications. Water Resour. Manag. 2010, 24, 4255–4271. [Google Scholar] [CrossRef]
- Cai, W.; Santoso, A.; Wang, G.; Yeh, S.W.; An, S.; Cobb, K.M.; Collins, M.; Guilyardi, E.; Lengaigne, M. ENSO and greenhouse warming. Nat. Clim. Chang. 2015, 5, 849–859. [Google Scholar] [CrossRef]
- Havens, K.E.; Fulton, R.S.; Beaver, J.R.; Samples, E.E.; Colee, J. Effects of climate variability on cladoceran zooplankton and cyanobacteria in a shallow subtropical lake. J. Plankton Res. 2016. [Google Scholar] [CrossRef]
- Phlips, E.J.; Aldridge, F.J.; Hansen, P.; Zimba, P.V.; Ihnat, J.; Conroy, M.; Ritter, R.R. Spatial and temporal variability of trophic state parameters in a shallow subtropical lake (Lake Okeechobee, Florida, USA). Arch. Hydrobiol. 1993, 128, 437–458. [Google Scholar]
- Aldridge, F.J.; Phlips, E.J.; Schelske, C.L. The use of nutrient enrichment bioassays to test for spatial and temporal distribution of limiting factors affecting phytoplankton dynamics in Lake Okeechobee, Florida. Arch. Hydrobiol. Adv. Limnol. 1995, 45, 177–190. [Google Scholar]
- Phlips, E.J.; Hendrickson, J.; Quinlan, E.; Cichra, M. Meteorological influences on algal bloom potential in a nutrient-rich blackwater river. Freshw. Biol. 2007, 52, 2141–2155. [Google Scholar] [CrossRef]
- Phlips, E.J.; Cichra, M.; Aldridge, F.J.; Jembeck, J.; Hendrickson, J.; Brody, R. Light availability and variations in phytoplankton standing crops in a nutrient-rich blackwater river. Limnol. Oceanogr. 2000, 45, 916–929. [Google Scholar] [CrossRef]
- Hendrickson, J.; Lowe, E.F.; Dobberfuhl, D.; Sucsy, P.; Campbell, D. Characteristics of Accelerated Eutrophication in the Lower St. Johns River Estuary and Recommended Targets to Achieve Water Quality Goals for the Fulfillment of TMDL and PLRG Objectives; St. Johns River Water Management District: Palatka, FL, USA, 2003. [Google Scholar]
- Methods for Chemical Analysis of Water and Waste Water; United States Environmental Protection Agency: Washington, DC, USA, 1979.
- South Florida Water Management District Field Sampling Manual; SFWMD: West Palm Beach, FL, USA, 2008.
- Lund, J.G.W.; Kipling, C.; LeCren, E.D. The inverted microscope method of estimating algal numbers and the statistical basis of estimates by counting. Hydrobiologia 1958, 11, 143–170. [Google Scholar] [CrossRef]
- Rocha, O.; Duncan, A. The relationship between cell carbon and cell-volume in fresh-water algal species used in zooplanktonic studies. J. Plankton Res. 1985, 7, 279–294. [Google Scholar] [CrossRef]
- Auer, B.; Elzer, U.; Arndt, H. Comparison of pelagic food webs in lakes along a trophic gradient and with seasonal aspects: Influence of resource and predation. J. Plankton Res. 2004, 26, 697–709. [Google Scholar] [CrossRef]
- Havens, K.E.; Beaver, J.R.; East, T.L. Plankton biomass partitioning in a eutrophic subtropical lake: Comparison with results from temperate lake ecosystems. J. Plankton Res. 2007, 18, 1605–1625. [Google Scholar] [CrossRef]
- Edler, L. Recommendations on Methods for Marine Biological Studies in the Baltic Sea: Phytoplankton and Chlorophyll; The Baltic Marine Biologists: Stockholm, Sweden, 1979. [Google Scholar]
- Lundgren, A. Experimental lake fertilization in the Kuokkel Area, northern Sweden: Changes in sestonic carbon and the role of phytoplankton. Int. Ver. Theor. Ang. Limnol. Verh. 1978, 20, 863–868. [Google Scholar]
- Ahlgren, G. Comparison of methods for estimation of phytoplankton carbon. Arch. Hydrobiol. 1983, 98, 489–508. [Google Scholar]
- Stefan, H.; Skoglund, T.; Megard, R.O. Wind control of algal growth in eutrophic lakes. J. Environ. Eng. Div. ASCE 1976, 102, 1201–1203. [Google Scholar]
- Wetzel, R.G. Limnology; Academic Press: New York, NY, USA, 2001. [Google Scholar]
- Oswald, W.J.; Gataas, H.B. Phytosynthesis in sewage treatment. Trans. Am. Soc. Civ. Eng. 1957, 122, 73–97. [Google Scholar]
- Hu, C.M. A novel ocean color index to detect floating algae in the global oceans. Remote Sens. Environ. 2009, 113, 2118–2129. [Google Scholar] [CrossRef]
- Webster, P.J.; Holland, G.J.; Curry, J.A.; Chang, H.R. Changes in tropical cyclone number, duration, and intensity in a warming environment. Science 2005, 309, 1844–1846. [Google Scholar] [CrossRef] [PubMed]
- Elsner, J.B.; Kossin, J.P.; Jagger, T.H. The increasing intensity of the strongest tropical cyclones. Nature 2008, 455, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.; Emanuel, K. Grey swan tropical cyclones. Nat. Clim. Chang. 2016, 6, 106–111. [Google Scholar] [CrossRef]
- Moss, B.; Madgwick, J.; Phillips, G. A Guide to the Restoration of Nutrient-Enriched Shallow Lakes; W.W. Hawes, Inc.: London, UK, 1996. [Google Scholar]
- Scheffer, M. Alternative stable stages in eutrophic shallow freshwater systems: A minimal model. Hydrobiol. Bull. 1989, 23, 73–83. [Google Scholar] [CrossRef]
- Bachmann, R.W.; Hoyer, M.V.; Vinzon, S.B.; Canfield, D.E. The origin of the fluid mud layer in Lake Apopka, Florida. Limnol. Oceanogr. 2005, 50, 629–635. [Google Scholar] [CrossRef]
- James, R.T.; Chimney, M.J.; Sharfstein, B. Hurricane effects on a shallow lake ecosystem, Lake Okeechobee, Florida (USA). Arch. Hydrobiol. 2005, 163, 225–239. [Google Scholar] [CrossRef]
- Abtew, W.; Iricanin, N. Hurricane effects on south Florida water management system: A case study of Hurricane Wilma of October 2005. J. Spat. Hydrol. 2008, 8, 2–20. [Google Scholar]
- Gaiser, E.E.; Deyrup, N.D.; Bachmann, R.W.; Battoe, L.E.; Swain, H.M. Effects of climate variability on transparency and thermal structure in subtropical, monomictic Lake Annie, Florida. Arch. Hydrobiol. 2009, 175, 217–230. [Google Scholar] [CrossRef]
- Srifa, A.; Phlips, E.J.; Cichra, M.F.; Hendrickson, J.C. Phytoplankton dynamics in a subtropical lake dominated by cyanobacteria: Cyanobacteria like it hot and sometimes dry. Aquat. Ecol. 2016. [Google Scholar] [CrossRef]
- Piehler, M.F.; Dyble, J.; Moisander, P.H.; Chapman, A.D.; Hendrickson, J.; Paerl, H.W. Interactions between nitrogen dynamics and the phytoplankton community in Lake George, Florida, USA. Lake Reserv. Manag. 2009, 25, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Doron, M. Aquatic Nitrogen Fixation Patterns, Rates and Controls in a Shallow, Subtropical Lake. Master’ Thesis, University of Florida, Gainesville, FL, USA, 2010. [Google Scholar]
- Dong, L. Factors Controlling Phytoplankton Community and Dynamics during Cyanobacterial Bloom. Master’s Thesis, University of Florida, Gainesville, FL, USA, 2010. [Google Scholar]
- Paerl, H.W.; Huisman, J. Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Moss, B.; Kosten, S.; Meerhoff, M.; Battarbee, R.W.; Jeppesen, E.; Mazzeo, N.; Havens, K.; Lacerot, G.; Liu, Z.; De Meester, L.; et al. Allied attack: Climate change and eutrophication. Inland Waters 2011, 1, 101–105. [Google Scholar] [CrossRef]
- Havens, K.E.; Paerl, H.W. Climate change at a crossroad for control of harmful algal blooms. Environ. Sci. Technol. 2015, 49, 12605–12606. [Google Scholar] [CrossRef] [PubMed]












© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Havens, K.; Paerl, H.; Phlips, E.; Zhu, M.; Beaver, J.; Srifa, A. Extreme Weather Events and Climate Variability Provide a Lens to How Shallow Lakes May Respond to Climate Change. Water 2016, 8, 229. https://doi.org/10.3390/w8060229
Havens K, Paerl H, Phlips E, Zhu M, Beaver J, Srifa A. Extreme Weather Events and Climate Variability Provide a Lens to How Shallow Lakes May Respond to Climate Change. Water. 2016; 8(6):229. https://doi.org/10.3390/w8060229
Chicago/Turabian StyleHavens, Karl, Hans Paerl, Edward Phlips, Mengyuan Zhu, John Beaver, and Akeapot Srifa. 2016. "Extreme Weather Events and Climate Variability Provide a Lens to How Shallow Lakes May Respond to Climate Change" Water 8, no. 6: 229. https://doi.org/10.3390/w8060229
APA StyleHavens, K., Paerl, H., Phlips, E., Zhu, M., Beaver, J., & Srifa, A. (2016). Extreme Weather Events and Climate Variability Provide a Lens to How Shallow Lakes May Respond to Climate Change. Water, 8(6), 229. https://doi.org/10.3390/w8060229