1. Introduction
Soil compaction is a form of degradation affecting future global food security and continues to be a problem for farmers in many parts of the world [
1]. Over the past few decades, management practices and field activities as well as wheel traffic from various heavy farm machinery have led to an increase in soil compaction, prompting global concerns on soil quality, hydrological properties, and crop production [
2,
3,
4,
5,
6]. It can significantly reduce total soil porosity, change pore size distribution, affect hydrological processes, and impact crop production for food security [
1,
2,
3,
5,
6,
7,
8].
Several approaches have been utilized to alleviate soil compaction in agricultural lands including mechanical, natural, and biological. Mechanical methods such as deep tillage or subsoiling are good practical solutions in alleviating compaction problems in subsurface soil layers; however, these processes have detrimental effects on soil quality and organic matter decomposition as well as involving considerable expenditures of time, money and fuel [
2,
9,
10,
11].
Natural repeated freezing and thawing cycles and their dynamic forces have been proven effective tools for lessening soil compaction and improving soil physical quality in the top layers over winter periods in cold regions [
3,
12].
Biological methods such as deep-rooted cover crops can be another potential solution to ameliorate the negative effects of soil compaction, particularly in no-tillage farming production [
11,
13,
14]. Cover crops (e.g., radish, safflower, turnip, etc.) with vigorous taproots can reduce soil compaction by penetrating and loosening the compacted layer. Eventually, these roots decompose over time and form root channels and large voids that enable air, water, nutrient and roots of subsequent crops to move more deeply through the soil profile, thus enhancing soil macropores and physical quality [
15,
16].
Some researches refer to this biological process as a “bio-drilling” or “biological drilling” or “bio-tillage” which is the root-induced formation of voids “bio-pores” that present deep paths for air, water, nutrient movement, and root growth, and penetration of subsequent crops [
15,
16,
17,
18].
Ehlers et al. [
17] evaluated soil penetration resistance and root growth of oats in tilled and untilled soils. They concluded that root channels or “bio-pores” created by roots of preceding crops acted as passages for subsequent crops in the soil profile in no-tillage systems.
Williams and Weil [
15] observed that soybean roots grew through the compacted plow pan soil using channels and “bio-pores” created by decomposition of cover crop roots. Similarly, Chen and Weil [
16] evaluated the effect of three cover crop roots in compacted soils. They concluded that forage radish and rapeseed had at least twice as many roots as rye in a highly compacted soil and determined that tap-rooted crops may
penetrate the compacted soil layer far better than fibrous-rooted crops.
Recently, Rosolem and Pivetta [
11] investigated both mechanical and biological methods to alleviate soil compaction in tropical clay soils. In their study, they evaluated root growth and activity of soybean and maize grown in rotation with cover crops. They found that including ruzi grass and castor cover crops in the rotation increased root growth, activity and “bio-pores” in the soil profile as well as increased soybean yield.
We hypothesized that cover crops and cool-season oilseed crops with different root systems (i.e., vigorous taproots, fibrous roots) would have a potential to alleviate soil compaction in agricultural lands. Globally, soil compaction can affect crop productivity and millions of hectares are affected by soil compaction that cost farmers millions of dollars every year. Crop roots can improve compacted soils and eliminate the need for expensive tillage operations, which tend to provide only a short-term benefit while reversing benefits of no-till production on soil quality. Further, information about using crop roots as an approach to biologically managing and loosening compacted soils in the MonDak region (eastern Montana and western North Dakota, USA) of the northern Great Plains is lacking. Therefore, the objective of this study was to evaluate the effect of previous crop roots on soil compaction in 2 yr durum wheat rotations with camelina, carinata and a cover crop mix planted in place of fallow in a no-tillage farming system.
2. Materials and Methods
2.1. Field Experiment
A long-term field study was carried out from 2014 to 2020 at the USDA-ARS dryland site located approximately 11 km north of Culbertson (48°33′ N, 104°50′ W, altitude 589 m) in the northern Great Plains, USA. The soil at the research site was classified as Dooley sandy loam (fine-loamy, mixed, superactive, frigid Typic Argiustolls) (
https://soilseries.sc.egov.usda.gov (accessed on 16 February 2021)), derived from glacial till parent material, with a slope of 0 to 2%. The amounts of sand, silt, and clay in Ap horizon (0–15 cm) were approximately 62%, 22%, and 16%, respectively, and in Bt (15–30 cm) horizon were approximately 60%, 21%, and 19%, respectively. The presence of argillic horizon in this layer can greatly restrict water flow, root development and has higher density values and less macropores. The soil sampled prior to initiation of this experiment contained approximately 1.7% organic matter and has 7.2 pH at the 0–20 cm depth [
19]. Monthly precipitation amounts during 2014–2020 growing seasons at the study site are given in
Table 1. The previous cropping system was continuous spring or spring wheat–summer fallow at this site for over 30 years. Annual tillage to a depth of 15 to 20 cm was typical prior to 2005 to control weeds and prepare the seedbed for planting. Thereafter, no-tillage was practiced and weeds were controlled with herbicides. The moldboard plow has not been used at this field for over 25 years.
Crops in a 2 yr rotation with durum wheat (Triticum durum D.) were camelina (Camelina sativa L.), carinata (Brassica carinata A.), both belonging to the Brassicace family, and a cover crop mix (10 species). The cover crop mix was composed of 12% lentil (Lens culinaris), 19% forage pea (Pisum sativum), 6% radish (Raphanus sativus), 3% turnip (Brassica rapa subsp), 3% winter canola (Brassica napus), 6% flax (Linum usitatissimum), 13% forage sorghum (Sorghum vulgare Pers), 6% German millet (Setaria italic), 19% cowpea (Vigna unguiculata), and 13% buckwheat (Fagopyrum esculentum). We planted a diversified cover crop mix to maximize ecosystem services with inclusion of functional groups including pollinator species, warm and cool season plants, broadleafs and grasses, legumes, and crucifer species. A highly variable climate in the region typically favors different species productivity among years, hence the desire to plant a highly diversified cover crop mix.
Planting for all crops in the study took place in late April to early May using a custom built no-till research drill with double-shoot Barton single-disk openers on 20.3 cm row spacing. Durum “Grenora” was planted at 67 kg ha−1. Camelina “Suneson”, carinata “A110”, and the cover crop mix were planted at 9.0, 7.3, and 26.9 kg ha−1. Crop seeds were harvested with a Kincaid 8XP research combine (manufactured in Haven, KS, USA).
Camelina seed was typically harvested late July to early August; durum harvest was early to mid-August. Carinata seed was harvested late August to mid-September. The cover crop mix was swathed for forage at pea bloom, typically occurring in mid-July. Following forage harvest, the cover crop was allowed to regrow unharvested and was terminated each year with a killing frost that typically occurred late September to mid-October.
Fertilizer was banded at planting 5 cm below and 5 cm to the side of seed. Urea (45-0-0) was applied to durum, camelina, and carinata based on yield goals of 2350, 1800, and 1800 kg ha
−1, resulting in N application rates of 129, 90, and 117 kg ha
−1, respectively, according to fertilizer guidelines for Montana crops [
20]. Urea rates were adjusted by subtracting N from monoammonium phosphate and ammonium sulfate fertilizer and from residual soil nitrate that was determined from soil samples (0–60 cm depth) taken the previous fall. All plots received 56 and 45 kg ha
−1 monoammonium phosphate (11-52-0) and potassium chloride (0-0-60) and camelina and carinata plots received 112 kg ha
−1 ammonium sulfate (20-0-0-24) banded at planting.
Weed control consisted of preplant and post-harvest applications of glyphosate herbicide at 3.36 kg a.i. ha–1. Durum also received an in-crop application of 0.68 kg a.i. ha–1 bromoxynil and 0.09 kg a.i ha–1 fenoxaprop herbicides. Camelina and carinata received 9.0 kg a.i. ha–1 ethylfluralin in late fall preceding spring planting.
Research plots in the rotation were arranged in randomized complete block design with three replications. Each crop phase of the rotation was present in every year for a total of 18 plots from three different rotations. Each plot measured 6 m wide × 15 m long.
2.2. Field Measurements
Soil penetration resistance (PR) was measured using a hand-held digital stainless steel cone-tipped penetrometer (12.8 mm diameter, 30° cone) (Field Scout, SC 900 Soil Compaction Meter; Spectrum Technologies, Inc., Plainfield, IL, USA). Measurements were made at three locations on a diagonal transect across each plot, one at the middle of the plot and the other two at each end of the plot. Penetration resistance readings (MPa) were recorded to the 30 cm depth at intervals of 2.5 cm [
3,
19].
At the time of PR measurements, soil moisture contents were determined for each plot using a digital TDR at three measurements per plot adjacent to PR positions (Field Scout, TDR 300 Soil Moisture Meter; Spectrum Technologies, Inc., Plainfield, IL, USA.
Soil PR and volumetric moisture content were monitored shortly after planting and after harvest in each plot every year throughout the course of the study.
Measurements were taken on 13 May, 11 September in 2014; 30 April, 11 September in 2015; 2 May, 26 September in 2016; 5 May, 4 October in 2017; 3 May, 2 October in 2018; 8 May, 8 October in 2019; and 21 April in 2020.
2.3. Statistical Analysis
The mixed model of SAS was used for data analysis [
21] using repeated measures over time as randomized block model with previous crop as a fixed effect, and year and replication as random effects.
The variables PR and volumetric moisture content measured by the TDR were tested for linearity and a significant linear correlation was found between these two soil parameters (r = −0.254, p = 0.009). Therefore, analysis of covariance was used to analyze soil PR for the compacted layer of 8–20 cm depth using moisture content as a covariable parameter. Mean separation utilized least significant differences at p ≤ 0.05.
3. Results and Discussion
Statistical results from analysis of covariance indicated that soil PR varied significantly by year (p < 0.0001) and not significantly by previous crop (p = 0.0554), with a significant interaction for previous crop × year (p = 0.0014).
Results indicated that the effect of previous crop on soil moisture content was not significant in all six years (
Table 2).
The effects of previous crop on soil PR and volumetric moisture content in the compacted layer of 8–20 cm for the 2015, 2016, 2017, 2018, 2019, and 2020 growing seasons are presented in
Table 3. The effect of previous crop on soil PR was not significant in all years except in 2015 and 2017. Variations in the PR among crops in 2015 and 2017 may be related to the soil variability within the field (
Table 3).
In 2014, soil PR averaged across all 18 plots was 2.244 MPa where durum or spring wheat was a previous crop for over 30 years. Results from 2014 showed all plots had a compacted layer that exceeded the critical compaction level of 2 MPa between the 8–20 cm depth [
22], presumably due to a history of tillage that likely caused a plow pan layer, presence of argillic horizon, continuous monocropping of spring wheat, and wheel traffic caused by other farming activities and management practices (
Figure 1).
Average soil penetration resistance across camelina, carinata, cover crop mix, and durum plots for the 8–20 cm layer after one, two, and three cycles of the 2 yr rotation were 2.370, 1.678, and 1.521 MPa in 2016, 2018, and 2020, respectively.
After one cycle of the 2 yr rotation (2014–2016), soil PR measured in spring 2016 indicated that the compacted layer continued to exist at the same depth of 8–20 cm. After two cycles (2014–2018), soil PR measurements in spring 2018 indicated that the compacted layer was decreased below the critical level of 2 MPa by 25.2% regardless of the rooting characteristics of the rotation crops. After three cycles (2014–2020), soil PR measurements in spring 2020 indicated that the compacted layer was reduced once again by 32.2% (
Figure 1).
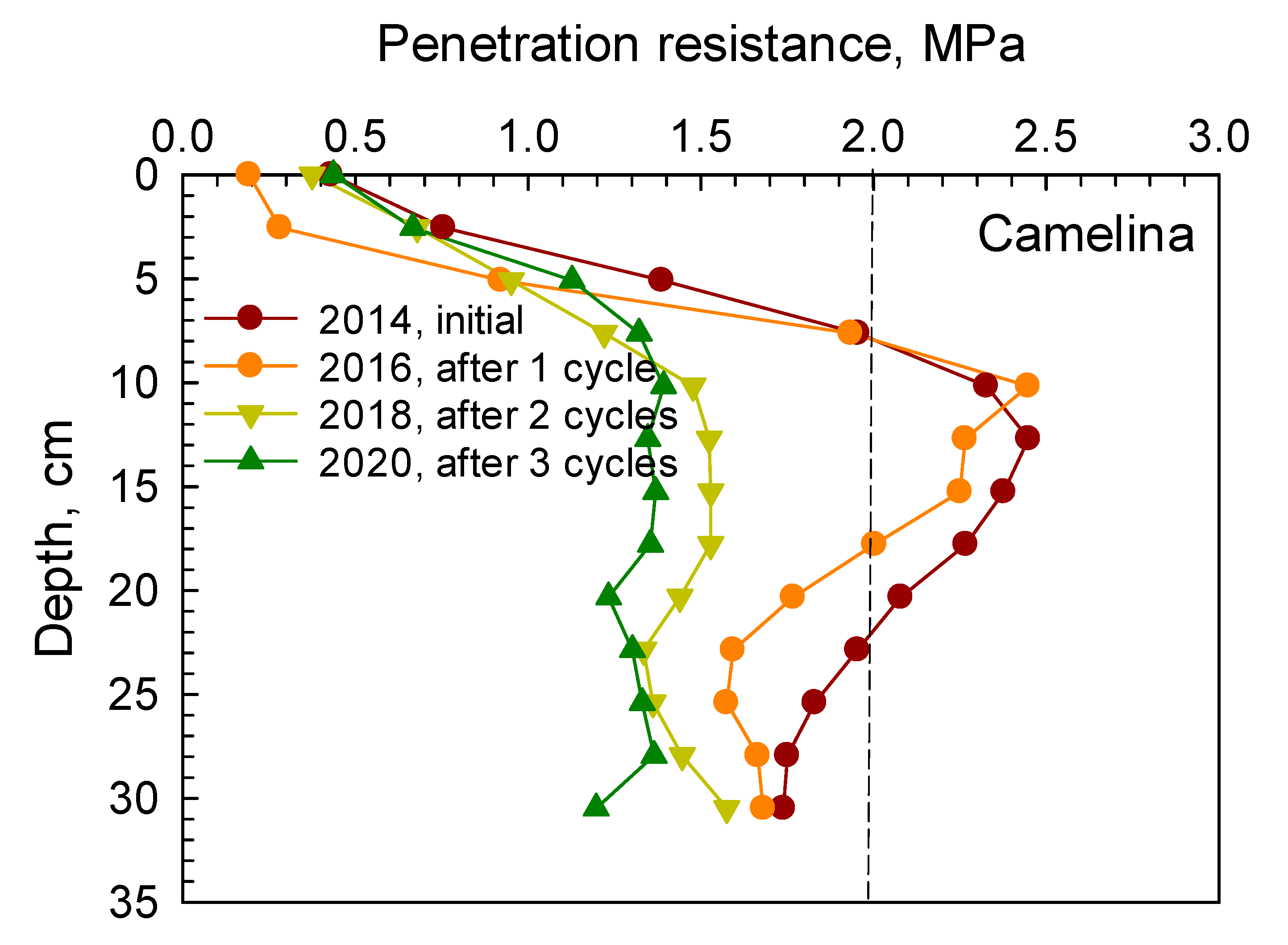
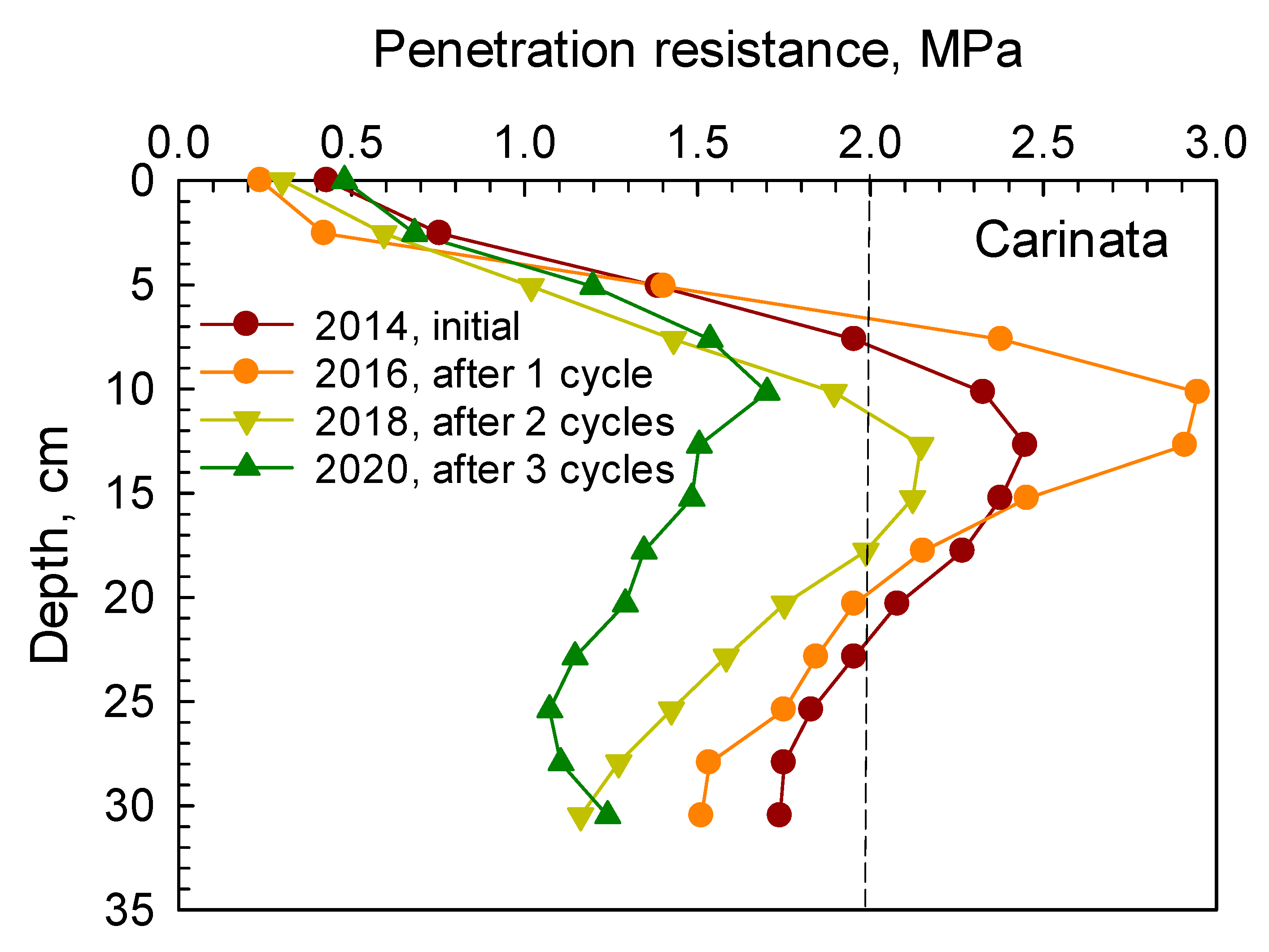
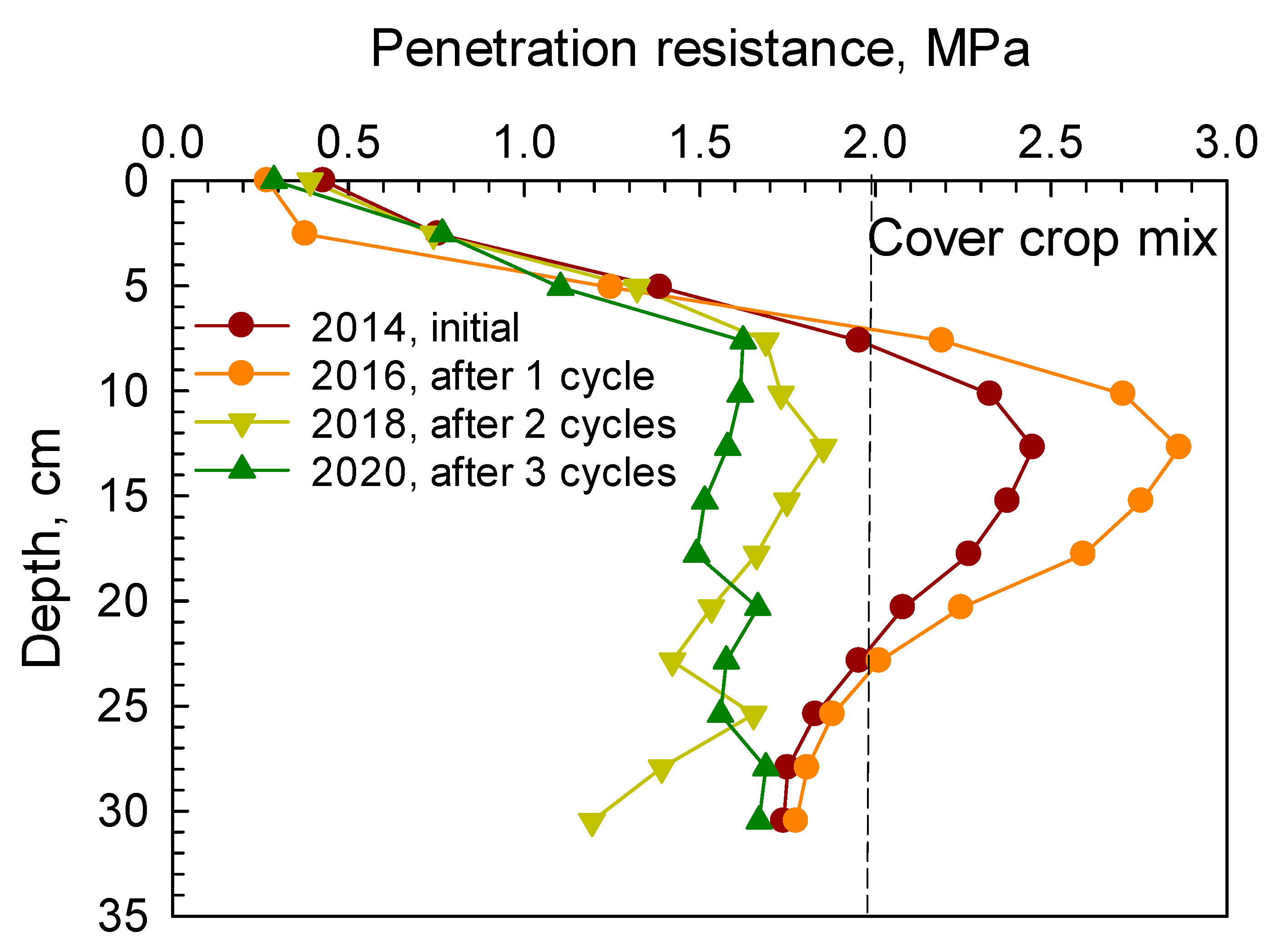
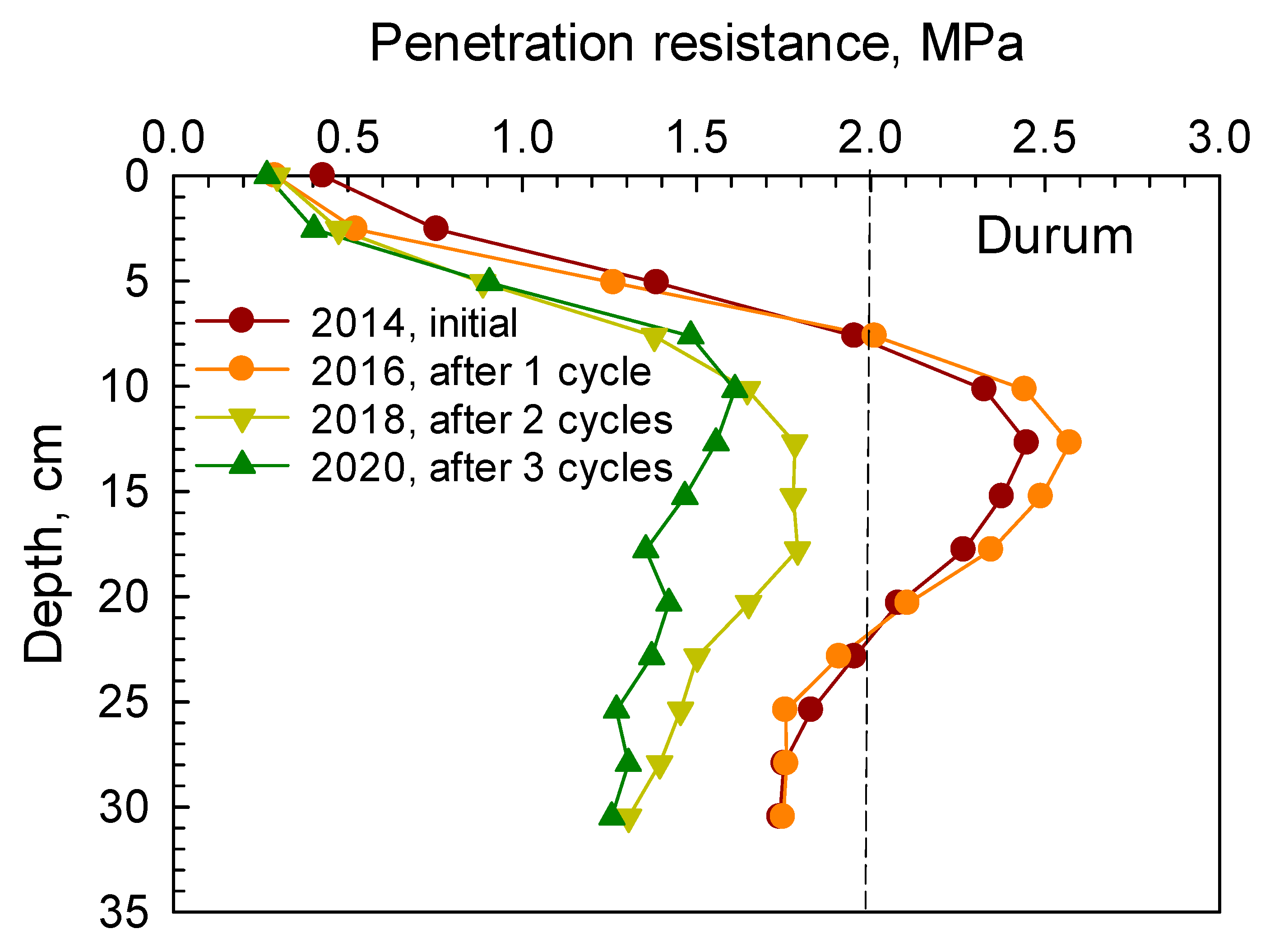
Similarly, soil PR from 0 to 30 cm depth under camelina, carinata, cover crop mix, and durum are presented in
Figure 2,
Figure 3,
Figure 4 and
Figure 5, respectively. Soil penetration resistance for the compacted layer under these crops exhibited similar patterns as described in
Figure 1.
After one cycle of the 2 yr rotation (2014–2016), soil PR measured in spring 2016 under camelina, carinata, cover crop mix, and durum indicated that the compacted layer was present at the 8–20 cm depth. After two and three cycles, soil PR in the compacted layer was decreased below the threshold of 2 MPa under each crop. Reductions in soil PR in the compacted layer after two and three cycles under camelina, carinata, cover crop mix, and durum are given in
Table 4. These reductions ranged between 19.3–34% and 29.5–35% after two and three cycles, respectively.
The root systems of cover crop mix and durum tended to have greater effects in loosening the compacted soil by creating more root channels and voids than both camelina and carinata after three cycles of the rotation (
Table 4). The cover crop mix treatment had various crops with a vigorous taproot system such as radish, forage sorghum, and turnip that penetrated through the compacted soil layer, decomposed over time, and created voids and passages through the dense layer.
Concluding results indicated that root systems of cover crops used in this study required approximately four years to be decomposed and produced root channels and pore spaces in the compacted soil profile. These findings showed that roots of camelina, carinata, cover crop mix, and durum could have a potential to decrease soil penetration resistance in the compacted layer by penetrating the compacted layer, eventually decompose, and form biological voids and channels, which enable air, water, nutrient and root growth for following crops to move easily through the compacted layer.
Figure 6 shows roots for various crops (sorghum, radish, turnip, etc.) in the cover crop mix treatment growing and penetrating through the compacted soil layer in summer of 2017. These roots will decompose over time and form large voids and root channels in the compacted layer.
The decomposition rate of cover crop roots and plant residues primarily depends on many factors including soil aeration, moisture content, temperature, texture, acidity, tillage, crop species, nitrogen availability, plant characteristics as well as microbial diversity and activities in the soil [
23].
The results of the study confirm previous research that showed that tap-rooted crops are more effective than fibrous-rooted crops in ameliorating compacted soils [
16,
18,
24]. Nevertheless, further investigations on different textured soils with various tap-rooted crops (i.e., rye grass, radish, safflower, turnip, Lucerne or alfalfa, rape, sugarbeet, chicory, etc.) as well as cover crops with fibrous-rooted crops are required for conclusive recommendations on crop rotations to avoid/recover from soil compaction in a no-tillage system.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}