
Moderate Grazer Density Stabilizes Forage Availability More Than Patch Burning in Low-Stature Grassland

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Location & Design
2.1.1. Fire Regime
2.1.2. Grazing Regime
2.2. Sampling Procedure
2.3. Data Analysis
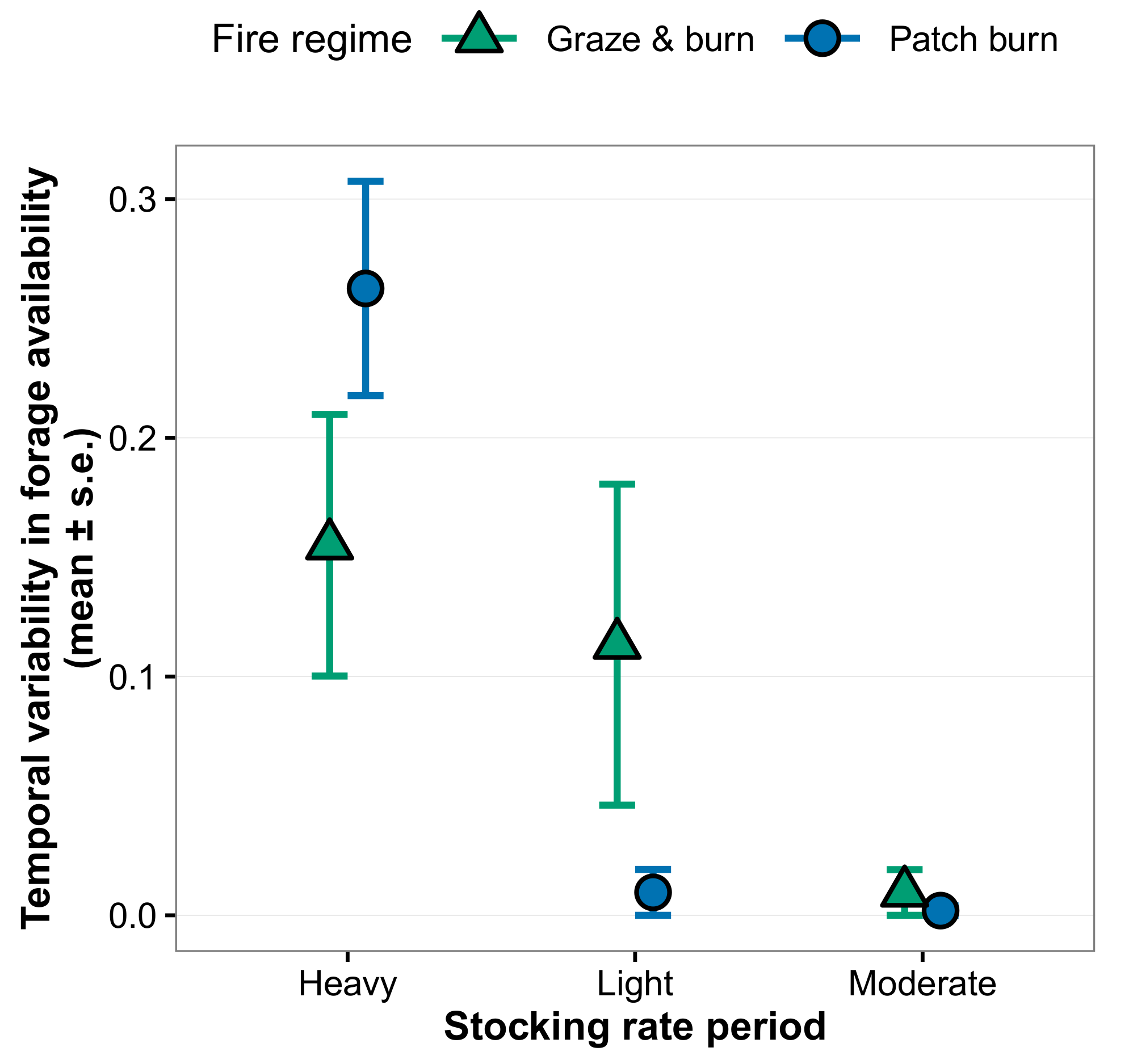
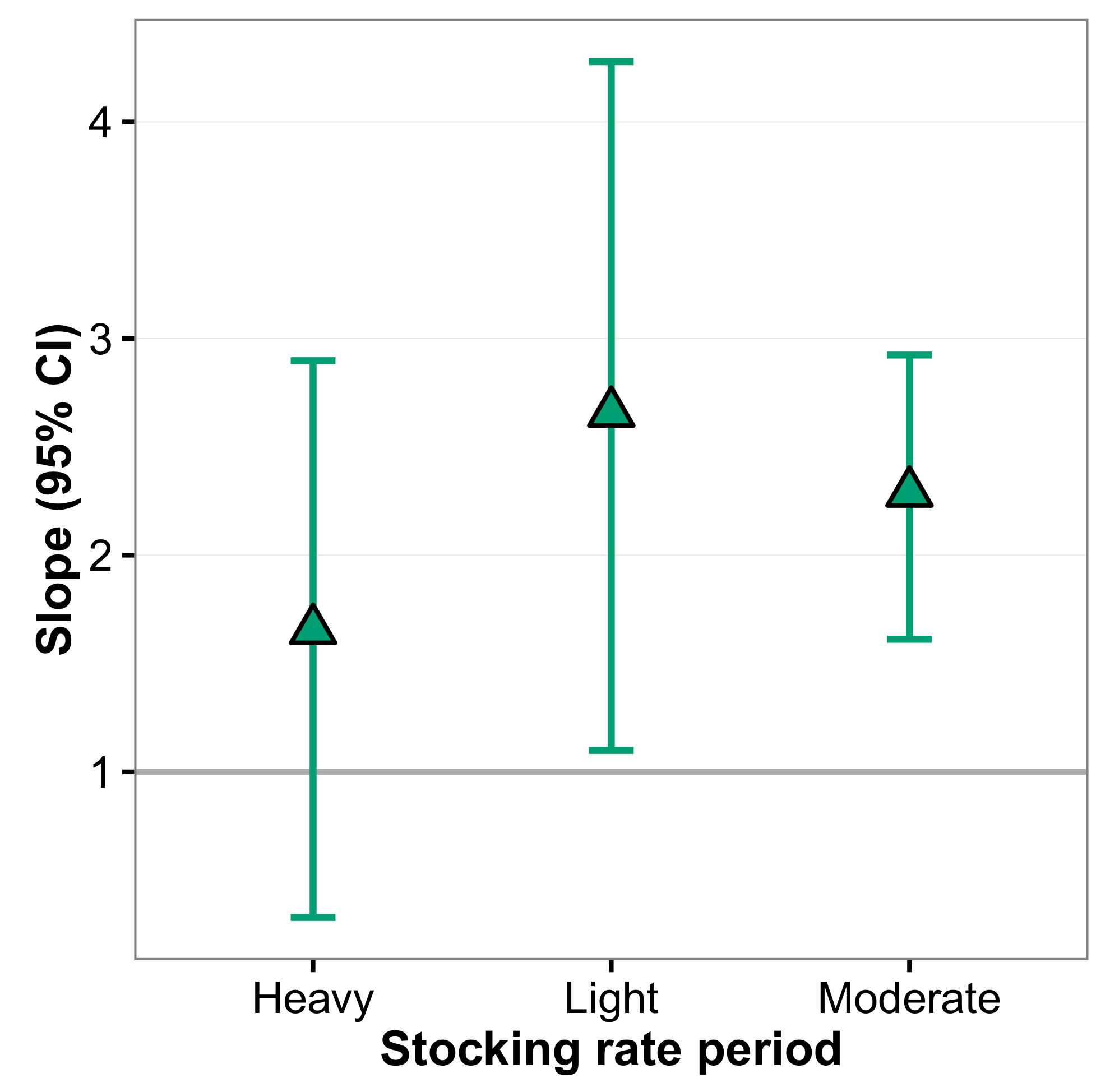
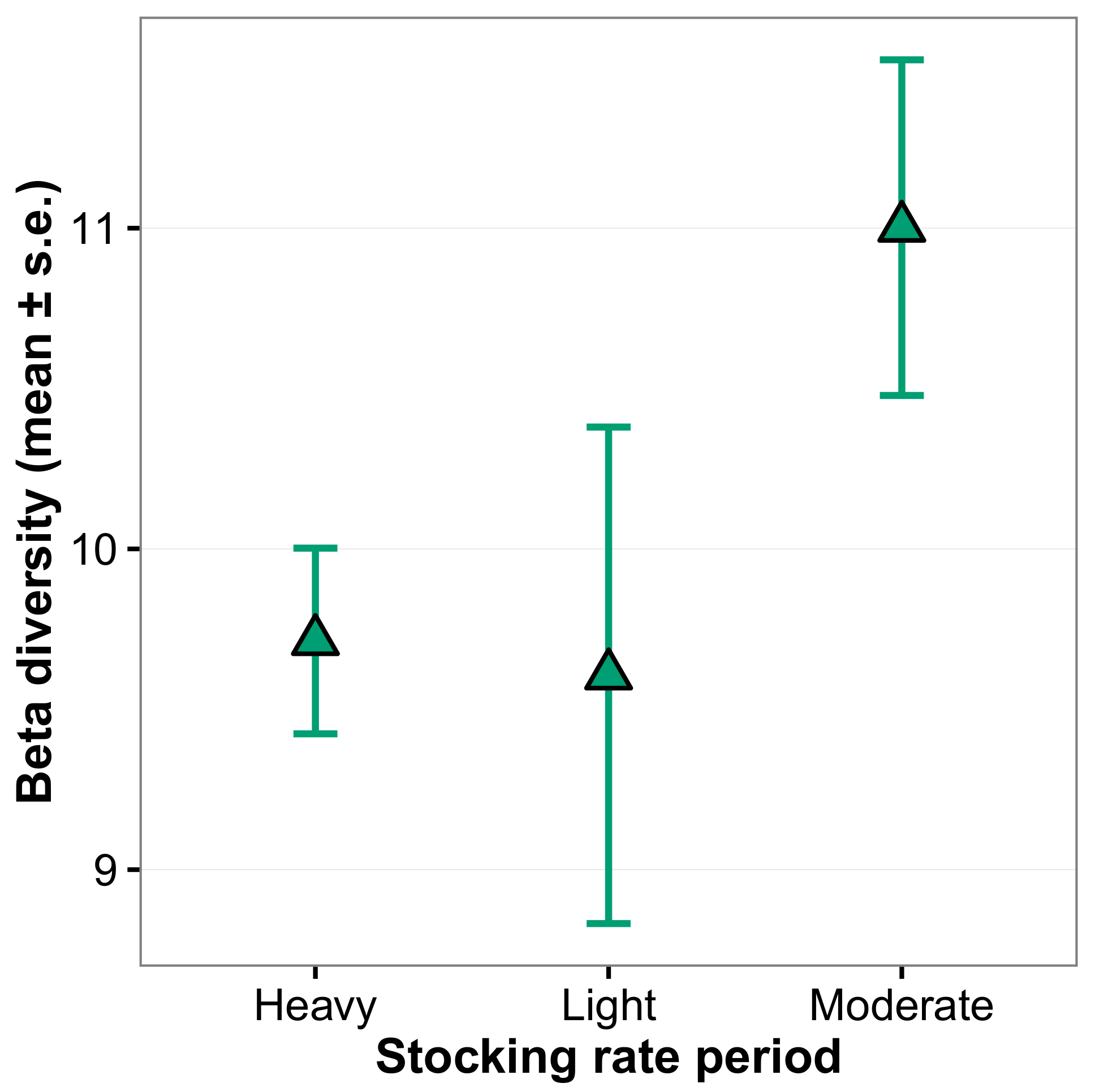
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Additional Statistical Results

{kind=link}
{kind=link}
{kind=link}
| Model | K | AICc | AICc | |
|---|---|---|---|---|
| Grazer density | 5 | −38.61 | 0.00 | 0.78 |
| Grazer density + Fire regime | 6 | −35.00 | 3.61 | 0.12 |
| Grazer density × Fire regime | 8 | 34.10 | 4.47 | 0.08 |
| Null | 3 | 27.00 | 11.56 | 0.002 |
| Fire regime | 4 | 24.10 | 14.47 | 0.001 |
References
- Fuhlendorf, S.D.; Engle, D.M.; Kerby, J.; Hamilton, R. Pyric Herbivory: Rewilding Landscapes through the Recoupling of Fire and Grazing. Conserv. Biol. 2009, 23, 588–598. [Google Scholar] [CrossRef]
- Transeau, E.N. The Prairie Peninsula. Ecology 1935, 16, 423–437. [Google Scholar] [CrossRef]
- Fuhlendorf, S.D.; Engle, D.M. Application of the Fire–Grazing Interaction to Restore a Shifting Mosaic on Tallgrass Prairie. J. Appl. Ecol. 2004, 41, 604–614. [Google Scholar] [CrossRef]
- Archibald, S.; Bond, W.J.; Stock, W.D.; Fairbanks, D.H.K. Shaping the Landscape: Fire-Grazer Interactions in an African Savanna. Ecol. Appl. 2005, 15, 96–109. [Google Scholar] [CrossRef]
- McGranahan, D.; Kirkman, K. Multifunctional Rangeland in Southern Africa: Managing for Production, Conservation, and Resilience with Fire and Grazing. Land 2013, 2, 176. [Google Scholar] [CrossRef]
- Fuhlendorf, S.D.; Harrell, W.C.; Engle, D.M.; Hamilton, R.G.; Davis, C.A.; Leslie, D.M., Jr. Should Heterogeneity Be the Basis for Conservation? Grassland Bird Response to Fire and Grazing. Ecol. Appl. 2006, 16, 1706–1716. [Google Scholar] [CrossRef] [Green Version]
- McGranahan, D.A.; Engle, D.M.; Fuhlendorf, S.D.; Winter, S.J.; Miller, J.R.; Debinski, D.M. Spatial Heterogeneity across Five Rangelands Managed with Pyric-Herbivory. J. Appl. Ecol. 2012, 49, 903–910. [Google Scholar] [CrossRef]
- Courtwright, J. “When We First Come Here It All Looked Like Prairie Land Almost”: Prairie Fire and Plains Settlement. West. Hist. Q. 2007, 38, 157–179. [Google Scholar] [CrossRef]
- Fuhlendorf, S.D.; Fynn, R.W.S.; McGranahan, D.A.; Twidwell, D. Heterogeneity as the Basis for Rangeland Management. In Rangeland Systems: Processes, Management and Challenges; Briske, D.D., Ed.; Springer Series on Environmental Management; Springer International Publishing: Cham, Switzerland, 2017; pp. 169–196. [Google Scholar]
- Twidwell, D.; Rogers, W.E.; Fuhlendorf, S.D.; Wonkka, C.L.; Engle, D.M.; Weir, J.R.; Kreuter, U.P.; Taylor, C.A. The Rising Great Plains Fire Campaign: Citizens’ Response to Woody Plant Encroachment. Front. Ecol. Environ. 2013, 11, e64–e71. [Google Scholar] [CrossRef]
- McGranahan, D.; Engle, D.; Fuhlendorf, S.; Miller, J.; Debinski, D. Multivariate Analysis of Rangeland Vegetation and Soil Organic Carbon Describes Degradation, Informs Restoration and Conservation. Land 2013, 2, 328. [Google Scholar] [CrossRef] [Green Version]
- McGranahan, D.A.; Engle, D.M.; Fuhlendorf, S.D.; Winter, S.L.; Miller, J.R.; Debinski, D.M. Inconsistent Outcomes of Heterogeneity-Based Management Underscore Importance of Matching Evaluation to Conservation Objectives. Environ. Sci. Policy 2013, 31, 53–60. [Google Scholar] [CrossRef] [Green Version]
- McGranahan, D.A.; Engle, D.M.; Fuhlendorf, S.D.; Miller, J.R.; Debinski, D.M. An Invasive Cool-Season Grass Complicates Prescribed Fire Management in a Native Warm-Season Grassland. Nat. Areas J. 2012, 32, 208–214. [Google Scholar]
- McGranahan, D.A.; Engle, D.M.; Miller, J.R.; Debinski, D.M. An Invasive Grass Increases Live Fuel Proportion and Reduces Fire Spread in a Simulated Grassland. Ecosystems 2013, 16, 158–169. [Google Scholar] [CrossRef]
- Duchardt, C.J.; Miller, J.R.; Debinski, D.M.; Engle, D.M. Adapting the Fire-Grazing Interaction to Small Pastures in a Fragmented Landscape for Grassland Bird Conservation. Rangel. Ecol. Manag. 2016, 69, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Foster, C.N.; Sato, C.F.; Lindenmayer, D.B.; Barton, P.S. Integrating Theory into Disturbance Interaction Experiments to Better Inform Ecosystem Management. Glob. Chang. Biol. 2016, 22, 1325–1335. [Google Scholar] [CrossRef]
- Raynor, E.J.; Coon, J.J.; Swartz, T.M.; Morton, L.W.; Schacht, W.H.; Miller, J.R. Shifting Cattle Producer Beliefs on Stocking and Invasive Forage: Implications for Grassland Conservation. Rangel. Ecol. Manag. 2019, 72, 888–898. [Google Scholar] [CrossRef]
- McGranahan, D.A.; Hovick, T.J.; Elmore, R.D.; Engle, D.M.; Fuhlendorf, S.D.; Winter, S.L.; Miller, J.R.; Debinski, D.M. Temporal Variability in Aboveground Plant Biomass Decreases as Spatial Variability Increases. Ecology 2016, 97, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J.; Ellingsen, K.E.; McArdle, B.H. Multivariate Dispersion as a Measure of Beta Diversity. Ecol. Lett. 2006, 9, 683–693. [Google Scholar] [CrossRef]
- McGranahan, D.A.; Hovick, T.J.; Elmore, R.D.; Engle, D.M.; Fuhlendorf, S.D. Moderate Patchiness Optimizes Heterogeneity, Stability, and Beta Diversity in Mesic Grassland. Ecol. Evol. 2018, 8, 5008–5015. [Google Scholar] [CrossRef] [Green Version]
- Allred, B.W.; Scasta, J.D.; Hovick, T.J.; Fuhlendorf, S.D.; Hamilton, R.G. Spatial Heterogeneity Stabilizes Livestock Productivity in a Changing Climate. Agric. Ecosyst. Environ. 2014, 193, 37–41. [Google Scholar] [CrossRef]
- McGranahan, D.A.; Henderson, C.B.; Hill, J.S.; Raicovich, G.M.; Wilson, W.N.; Smith, C.K. Patch Burning Improves Forage Quality and Creates Grassbank in Old-Field Pasture: Results of a Demonstration Trial. Southeast. Nat. 2014, 13, 200–207. [Google Scholar] [CrossRef]
- Spiess, J.W.; McGranahan, D.A.; Geaumont, B.; Sedivec, K.; Lakey, M.; Berti, M.; Hovick, T.J.; Limb, R.F. Patch-Burning Buffers Forage Resources and Livestock Performance to Mitigate Drought in the Northern Great Plains. Rangel. Ecol. Manag. 2020, 73, 473–481. [Google Scholar] [CrossRef]
- Phillips, W.A.; Coleman, S.W. Productivity and Economic Return of Three Warm Season Grass Stocker Systems for the Southern Great Plains. J. Prod. Agric. 1995, 8, 334–339. [Google Scholar] [CrossRef]
- McGranahan, D.A.; Engle, D.M.; Wilsey, B.J.; Fuhlendorf, S.D.; Miller, J.R.; Debinski, D.M. Grazing and an invasive grass confound spatial pattern of exotic and native grassland plant species richness. Basic Appl. Ecol. 2012, 13, 654–662. [Google Scholar] [CrossRef]
- Raynor, E.J.; Griffith, C.D.; Twidwell, D.; Schacht, W.H.; Wonkka, C.L.; Roberts, C.P.; Bielski, C.L.; Debinski, D.M.; Miller, J.R. The Emergence of Heterogeneity in Invasive-Dominated Grassland: A Matter of the Scale of Detection. Landsc. Ecol. 2018, 33, 2103–2119. [Google Scholar] [CrossRef]
- Scasta, J.D.; Duchardt, C.; Engle, D.M.; Miller, J.R.; Debinski, D.M.; Harr, R.N. Constraints to Restoring Fire and Grazing Ecological Processes to Optimize Grassland Vegetation Structural Diversity. Ecol. Eng. 2016, 95, 865–875. [Google Scholar] [CrossRef] [Green Version]
- Hempson, G.P.; Archibald, S.; Bond, W.J.; Ellis, R.P.; Grant, C.C.; Kruger, F.J.; Kruger, L.M.; Moxley, C.; Owen-Smith, N.; Peel, M.J.S.; et al. Ecology of Grazing Lawns in Africa. Biol. Rev. 2015, 90, 979–994. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.J.; Canziani, O.F.; Leary, N.A.; Dokken, D.J.; White, K.S. Climate Change 2001: Impacts, Adaptation, and Vulnerability: Contribution of Working Group II to the Third Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2001; Volume 2. [Google Scholar]
- Derner, J.D.; Hunt, L.; Filho, K.E.; Ritten, J.; Capper, J.; Han, G. Livestock Production Systems. In Rangeland Systems: Processes, Management and Challenges; Briske, D.D., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 347–372. [Google Scholar] [CrossRef] [Green Version]
- IEM. Iowa Environmental Mesonet, Iowa State University Department of Agronomy: Mt Ayr, IA. 2014. Available online: https://mesonet.agron.iastate.edu/climodat/index.phtml?network=IACLIMATE&station=IA5769&report=17 (accessed on 24 February 2021).
- USDA-SCS. Soil Survey of Ringgold County, Iowa; Technical Report; United States Department of Agriculture: Washington, DC, USA, 1992. [Google Scholar]
- McGranahan, D.A. Degradation and Restoration in Remnant Tallgrass Prairie: Grazing History, Soil Carbon, and Invasive Species Affect Community Composition and Response to the Fire-Grazing Interaction. Master’s Thesis, Iowa State University, Ames, IA, USA, 2008. [Google Scholar]
- Harrell, W.C.; Fuhlendorf, S.D. Evaluation of habitat structural measures in a shrubland community. Rangel. Ecol. Manag. 2002, 55, 488–493. [Google Scholar] [CrossRef]
- Vermeire, L.T.; Gillen, R.L. Estimating Herbage Standing Crop with Visual Obstruction in Tallgrass Prairie. J. Range Manag. 2001, 54, 57–60. [Google Scholar] [CrossRef]
- Daubenmire, R. A Canopy-Coverage Method of Vegetational Analysis. Northwest Sci. 1959, 33, 43–64. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Winter, S.L.; Fuhlendorf, S.D.; Goad, C.L.; Davis, C.A.; Hickman, K.R.; Leslie, D.M., Jr. Restoration of the Fire–Grazing Interaction in Artemisia filifolia Shrubland. J. Appl. Ecol. 2012, 49, 242–250. [Google Scholar] [CrossRef]
- Wang, S.; Loreau, M. Ecosystem Stability in Space: α, β and γ Variability. Ecol. Lett. 2014, 17, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Taylor, L.R. Aggregation, variance and the mean. Nature 1961, 189, 732–735. [Google Scholar] [CrossRef]
- Doak, D.F.; Bigger, D.; Harding, E.K.; Marvier, M.A.; O’Malley, R.E.; Thomson, D. The Statistical Inevitability of Stability-Diversity Relationships in Community Ecology. Am. Nat. 1998, 151, 264–276. [Google Scholar] [CrossRef]
- Tilman, D.; Clarence, L.L.; Charles, E.B. Diversity-Stability Relationships: Statistical Inevitability or Ecological Consequence? Am. Nat. 1998, 151, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘Vegan’. Community Ecol. Packag. Version 2013, 2, 1–295. [Google Scholar]
- Maresh Nelson, S.B.; Coon, J.J.; Schacht, W.H.; Miller, J.R. Cattle Select against the Invasive Grass Tall Fescue in Heterogeneous Pastures Managed with Prescribed Fire. Grass Forage Sci. 2019, 74, 486–495. [Google Scholar] [CrossRef]
- Cid, M.S.; Brizuela, M.A. Heterogeneity in Tall Fescue Pastures Created and Sustained by Cattle Grazing. J. Range Manag. 1998, 51, 644–649. [Google Scholar] [CrossRef] [Green Version]
- Petrie, M.D.; Peters, D.P.C.; Yao, J.; Blair, J.M.; Burruss, N.D.; Collins, S.L.; Derner, J.D.; Gherardi, L.A.; Hendrickson, J.R.; Sala, O.E.; et al. Regional Grassland Productivity Responses to Precipitation during Multiyear Above- and below-Average Rainfall Periods. Glob. Chang. Biol. 2018, 24, 1935–1951. [Google Scholar] [CrossRef]
- Milchunas, D.G.; Lauenroth, W.K. Quantitative Effects of Grazing on Vegetation and Soils over a Global Range of Environments. Ecol. Monogr. 1993, 63, 327–366. [Google Scholar] [CrossRef]



| Study Phase | Time Period | Stocking Rate (AUM ) |
|---|---|---|
| Heavy stocking | 2007–2009 | 3.2 ± 0.2 |
| Light stocking | 2010–2011 | 1.5 ± 0.1 |
| Moderate stocking | 2012–2013 | 2.4 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raynor, E.J.; McGranahan, D.A.; Miller, J.R.; Debinski, D.M.; Schacht, W.H.; Engle, D.M. Moderate Grazer Density Stabilizes Forage Availability More Than Patch Burning in Low-Stature Grassland. Land 2021, 10, 395. https://doi.org/10.3390/land10040395
Raynor EJ, McGranahan DA, Miller JR, Debinski DM, Schacht WH, Engle DM. Moderate Grazer Density Stabilizes Forage Availability More Than Patch Burning in Low-Stature Grassland. Land. 2021; 10(4):395. https://doi.org/10.3390/land10040395
Chicago/Turabian StyleRaynor, Edward J., Devan Allen McGranahan, James R. Miller, Diane M. Debinski, Walter H. Schacht, and David M. Engle. 2021. "Moderate Grazer Density Stabilizes Forage Availability More Than Patch Burning in Low-Stature Grassland" Land 10, no. 4: 395. https://doi.org/10.3390/land10040395
APA StyleRaynor, E. J., McGranahan, D. A., Miller, J. R., Debinski, D. M., Schacht, W. H., & Engle, D. M. (2021). Moderate Grazer Density Stabilizes Forage Availability More Than Patch Burning in Low-Stature Grassland. Land, 10(4), 395. https://doi.org/10.3390/land10040395