
A Quantitative Survey of Effect of Semi-Natural Habitat Composition and Configuration on Landscape Heterogeneity in Arable Land System



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
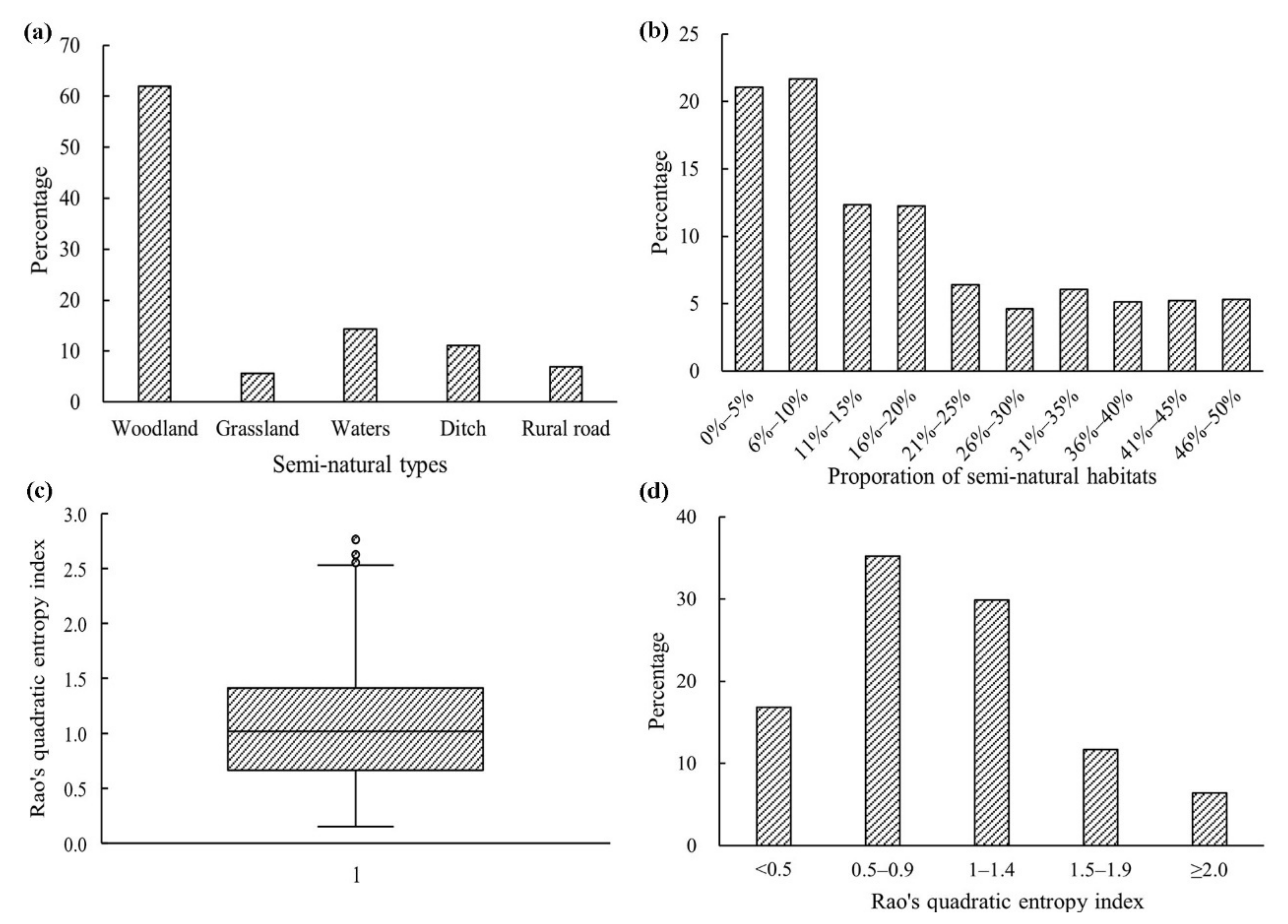
2.2. Semi-Natural Habitat Composition
2.3. Semi-Natural Habitat Configuration
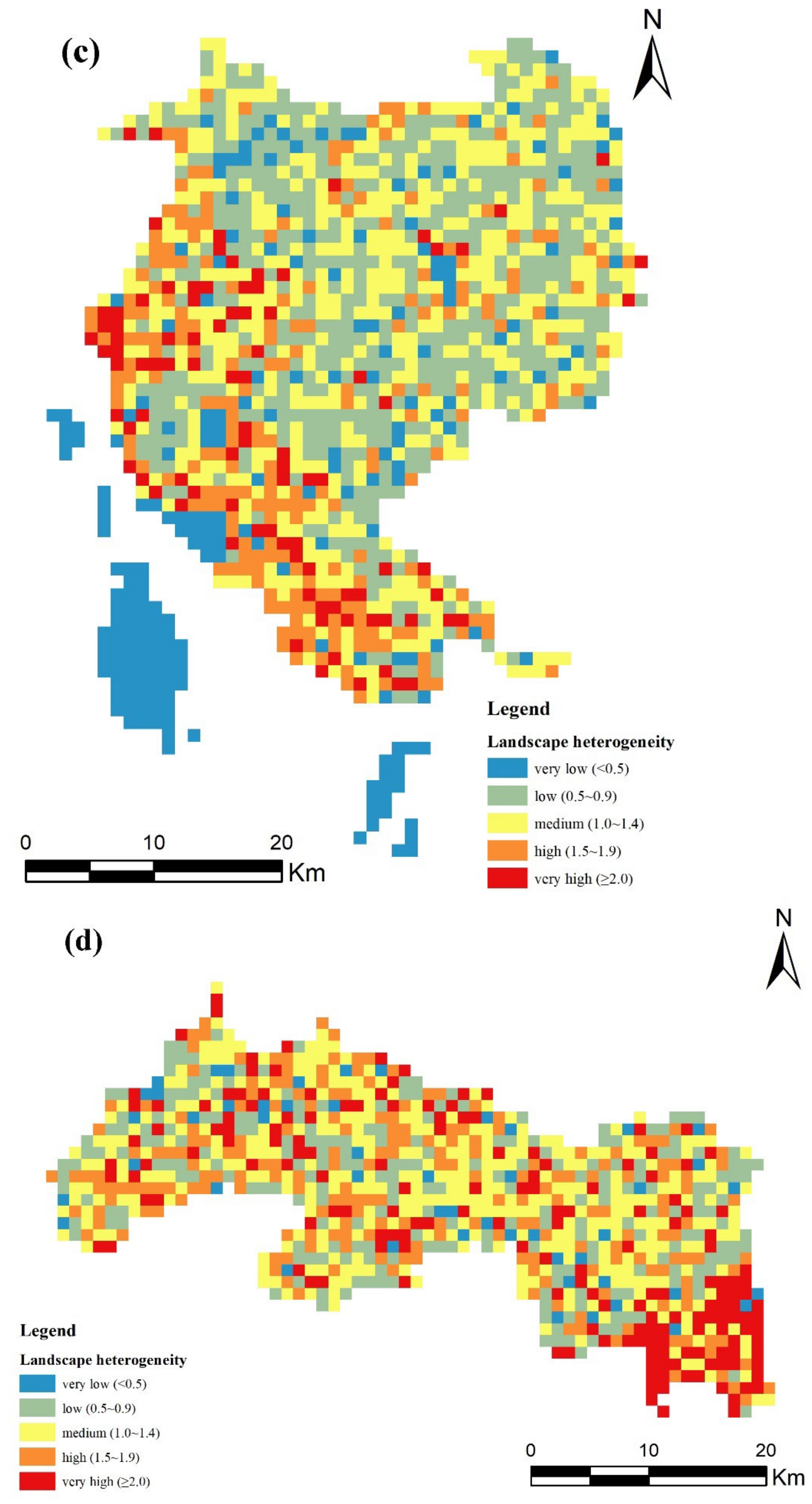
2.4. Landscape Heterogeneity
2.5. Statistical Analysis
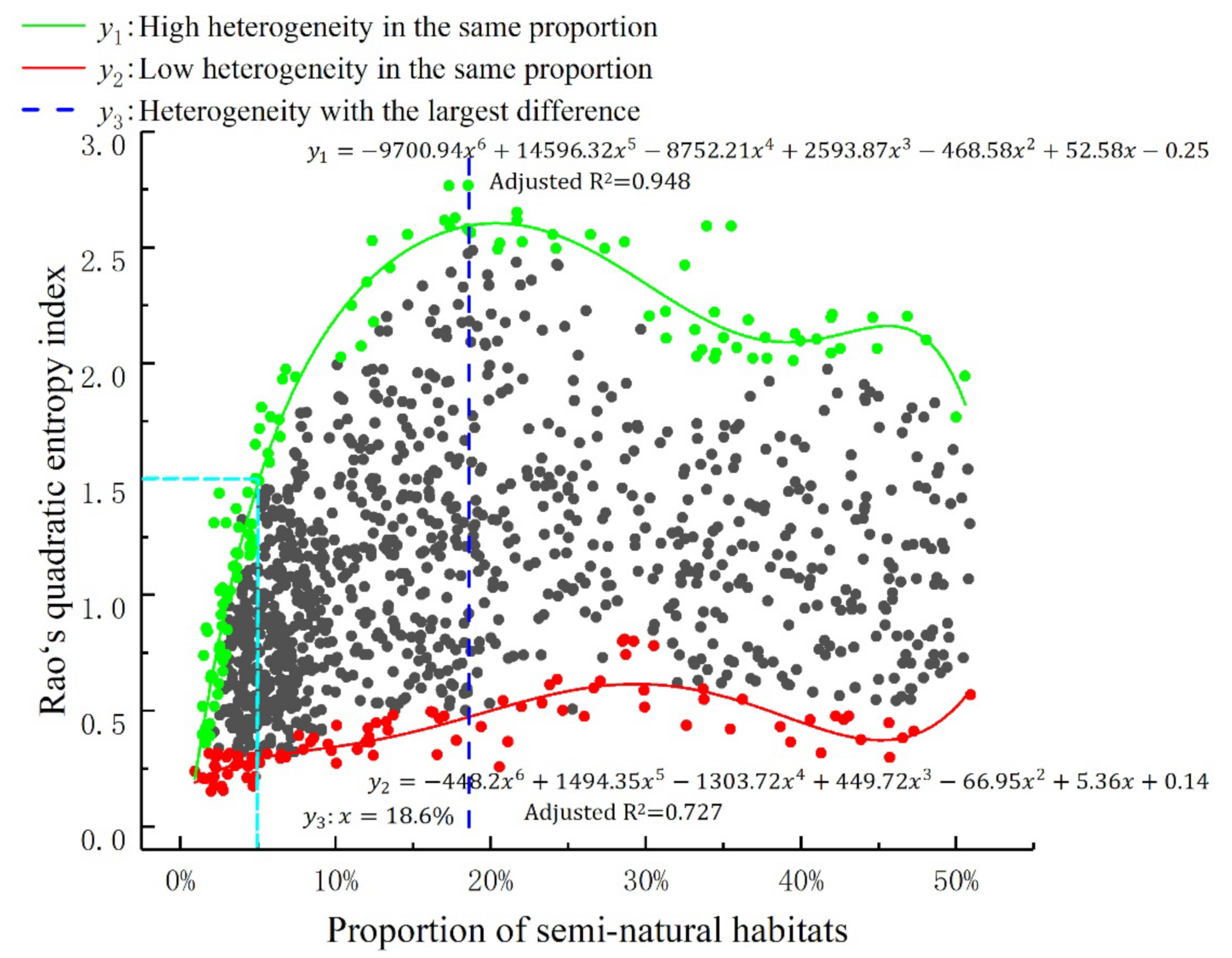
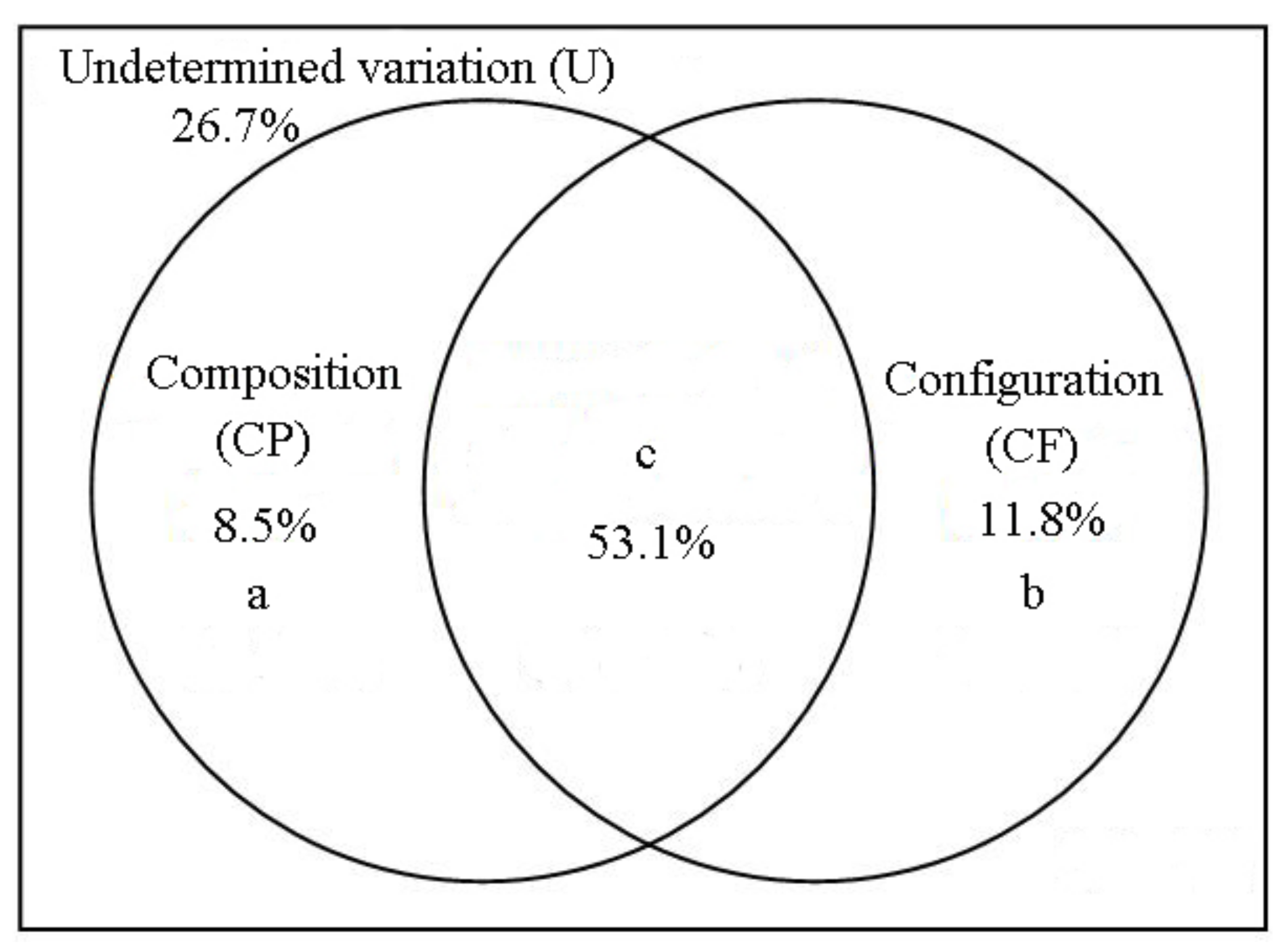
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Campbell, B.M.; Beare, D.J.; Bennett, E.M.; Hall-Spencer, J.M.; Ingram, J.S.; Jaramillo, F.; Ortiz, R.; Ramankutty, N.; Sayer, J.A.; Shindell, D. Agriculture production as a major driver of the Earth system exceeding planetary boundaries. Ecol. Soc. 2017, 22, 8. [Google Scholar] [CrossRef]
- Tscharntke, T.; Tylianakis, J.M.; Rand, T.A.; Didham, R.K.; Fahrig, L.; Batáry, P.; Bengtsson, J.; Clough, Y.; Crist, T.O.; Dormann, C.F.; et al. Landscape moderation of biodiversity patterns and processes—Eight hypotheses. Biol. Rev. 2012, 87, 661–685. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Raven, P.H.; Wagner, D.L. Agricultural intensification and climate change are rapidly decreasing insect biodiversity. Proc. Natl. Acad. Sci. USA 2021, 118, e2002548117. [Google Scholar] [CrossRef]
- Alsterberg, C.; Roger, F.; Sundbäck, K.; Juhanson, J.; Hulth, S.; Hallin, S.; Gamfeldt, L.; Naturvetenskapliga, F.; Faculty, O.S.; Göteborgs, U.; et al. Habitat diversity and ecosystem multifunctionality-The importance of direct and indirect effects. Sci. Adv. 2017, 3, e1601475. [Google Scholar] [CrossRef] [Green Version]
- Tamburini, G.; Bommarco, R.; Wanger, T.C.; Kremen, C.; van der Heijden, M.G.A.; Liebman, M.; Hallin, S.; Sveriges, L. Agricultural diversification promotes multiple ecosystem services without compromising yield. Sci. Adv. 2020, 6, eaba1715. [Google Scholar] [CrossRef]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- García-Feced, C.; Weissteiner, C.J.; Baraldi, A.; Paracchini, M.L.; Maes, J.; Zulian, G.; Kempen, M.; Elbersen, B.; Pérez-Soba, M. Semi-natural vegetation in agricultural land: European map and links to ecosystem service supply. Agron. Sustain. Dev. 2015, 35, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Zulian, G.; Maes, J.; Paracchini, M. Linking Land Cover Data and Crop Yields for Mapping and Assessment of Pollination Services in Europe. Land 2013, 2, 472–492. [Google Scholar] [CrossRef] [Green Version]
- Rega, C.; Bartual, A.M.; Bocci, G.; Sutter, L.; Albrecht, M.; Moonen, A.; Jeanneret, P.; van der Werf, W.; Pfister, S.C.; Holland, J.M.; et al. A pan-European model of landscape potential to support natural pest control services. Ecol. Indic. 2018, 90, 653–664. [Google Scholar] [CrossRef]
- Wezel, A.; Casagrande, M.; Celette, F.; Vian, J.; Ferrer, A.; Peigné, J. Agroecological practices for sustainable agriculture. A review. Agron. Sustain. Dev. 2014, 34, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Batáry, P.; Dicks, L.V.; Kleijn, D.; Sutherland, W.J. The role of agri-environment schemes in conservation and environmental management. Conserv. Biol. 2015, 29, 1006–1016. [Google Scholar] [CrossRef] [Green Version]
- Merckx, T.; Pereira, H.M. Reshaping agri-environmental subsidies: From marginal farming to large-scale rewilding. Basic Appl. Ecol. 2015, 16, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Commission Implementing Regulation (EU). 2021/2289 of 21 December 2021 laying down rules for the application of Regulation (EU) 2021/2115 of the European Parliament and of the Council on the presentation of the content of the CAP Strategic Plans and on the electronic system for the secure exchange of information. Off. J. Eur. Union 2021. Available online: http://data.europa.eu/eli/reg_impl/2021/2289/oj (accessed on 22 May 2022).
- Czúcz, B.; Baruth, B.; Terres, J.M.; Gallego, J.; Hagyo, A.; Angileri, V.; Nocita, M.; Soba, M.P.; Koeble, R.; Paracchini, M.L. Classification and Quantification of Landscape Features in Agricultural Land across the EU; European Commission: Brussels, Belgium, 2022. [Google Scholar]
- Tscharntke, T.; Batáry, P.; Dormann, C.F. Set-aside management: How do succession, sowing patterns and landscape context affect biodiversity? Agric. Ecosyst. Environ. 2011, 143, 37–44. [Google Scholar] [CrossRef]
- Holzkämper, A.; Lausch, A.; Seppelt, R. Optimizing landscape configuration to enhance habitat suitability for species with contrasting habitat requirements. Ecol. Model. 2006, 198, 277–292. [Google Scholar] [CrossRef]
- Harlio, A.; Kuussaari, M.; Heikkinen, R.K.; Arponen, A. Incorporating landscape heterogeneity into multi-objective spatial planning improves biodiversity conservation of semi-natural grasslands. J. Nat. Conserv. 2019, 49, 37–44. [Google Scholar] [CrossRef]
- Viana, C.M.; Freire, D.; Abrantes, P.; Rocha, J.; Pereira, P. Agricultural land systems importance for supporting food security and sustainable development goals: A systematic review. Sci. Total Environ. 2022, 806, 150718. [Google Scholar] [CrossRef]
- Mitchell, M.S.; Powell, R.A. A mechanistic home range model for optimal use of spatially distributed resources. Ecol. Model. 2004, 177, 209–232. [Google Scholar] [CrossRef]
- Fahrig, L.; Nuttle, W.K. Population Ecology in Spatially Heterogeneous Environments. In Ecosystem Function in Heterogeneous Landscapes; Lovett, G.M., Turner, M.G., Jones, C.G., Weathers, K.C., Eds.; Springer: New York, NY, USA, 2005. [Google Scholar]
- Pasher, J.; Mitchell, S.W.; King, D.J.; Fahrig, L.; Smith, A.C.; Lindsay, K.E. Optimizing landscape selection for estimating relative effects of landscape variables on ecological responses. Landsc. Ecol. 2013, 28, 371–383. [Google Scholar] [CrossRef]
- Zheng, X.; Zhu, J. A new climatic classification of afforestation in Three-North regions of China with multi-source remote sensing data. Theor. Appl. Climatol. 2017, 127, 465–480. [Google Scholar] [CrossRef]
- O’Neill, R.V.; Hunsaker, C.T.; Timmins, S.P.; Jackson, B.L.; Jones, K.B.; Riitters, K.H.; Wickham, J.D. Scale problems in reporting landscape pattern at the regional scale. Landsc. Ecol. 1996, 11, 169–180. [Google Scholar] [CrossRef]
- Martin, E.A.; Dainese, M.; Clough, Y.; Báldi, A.; Bommarco, R.; Gagic, V.; Garratt, M.P.D.; Holzschuh, A.; Kleijn, D.; Kovács Hostyánszki, A.; et al. The interplay of landscape composition and configuration: New pathways to manage functional biodiversity and agroecosystem services across Europe. Ecol. Lett. 2019, 22, 1083–1094. [Google Scholar] [CrossRef] [Green Version]
- Uuemaa, E.; Antrop, M.; Roosaare, J.; Marja, R.; Mander, Ü. Landscape Metrics and Indices: An Overview of Their Use in Landscape Research. Living Rev. Landsc. Res. 2009, 3, 1–28. [Google Scholar] [CrossRef]
- Hesselbarth, M.H.K.; Sciaini, M.; With, K.A.; Wiegand, K.; Nowosad, J. landscapemetrics: An open-source R tool to calculate landscape metrics. Ecography 2021, 42, 1648–1657. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Varela, E.; Roces-Díaz, J.V.; Álvarez-Álvarez, P. Detection of landscape heterogeneity at multiple scales: Use of the Quadratic Entropy Index. Landsc. Urban Plan 2016, 153, 149–159. [Google Scholar] [CrossRef]
- Doxa, A.; Prastacos, P. Using Rao’s quadratic entropy to define environmental heterogeneity priority areas in the European Mediterranean biome. Biol. Conserv. 2020, 241, 108366. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5–6. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 22 May 2022).
- Amaral, T.S.D.; Santos, J.S.D.; Rosa, F.F.; Pessôa, M.B.; Chaves, L.J.; Ribeiro, M.C.; Collevatti, R.G. Agricultural Landscape Heterogeneity Matter: Responses of Neutral Genetic Diversity and Adaptive Traits in a Neotropical Savanna Tree. Front. Genet. 2021, 11, 606222. [Google Scholar] [CrossRef]
- Sirami, C.; Gross, N.; Baillod, A.B.; Bertrand, C.; Carrié, R.; Hass, A.; Henckel, L.; Miguet, P.; Vuillot, C.; Alignier, A.; et al. Increasing crop heterogeneity enhances multitrophic diversity across agricultural regions. Proc. Natl. Acad. Sci. USA 2019, 116, 16442–16447. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Regulation (EU) No. 1307/2013 of the European Parliament and of the Council of 17 December 2013. Off. J. Eur. Union 2013 608–670. Available online: http://data.europa.eu/eli/reg/2013/1307/oj (accessed on 22 May 2022).
- European Commission. The Post-2020 Common Agricultural Policy: Environmental Benefits and Simplification; European Network for Rural Development: Eisenstadt, Austria, 2019; CID: 20.500.12592/svct60. [Google Scholar]
- Rotchés-Ribalta, R.; Ruas, S.; Ahmed, K.D.; Gormally, M.; Moran, J.; Stout, J.; White, B.; HUallacháin, D.Ó. Assessment of semi-natural habitats and landscape features on Irish farmland: New insights to inform EU Common Agricultural Policy implementation. Ambio 2021, 50, 346–359. [Google Scholar] [CrossRef]
- Li, L.; Hu, R.; Huang, J.; Bürgi, M.; Zhu, Z.; Zhong, J.; Lü, Z. A farmland biodiversity strategy is needed for China. Nat. Ecol. Evol. 2020, 4, 772–774. [Google Scholar] [CrossRef]
- Guo, X.; Bian, Z.; Wang, S.; Wang, Q.; Zhang, Y.; Zhou, J.; Lin, L. Prediction of the spatial distribution of soil arthropods using a random forest model: A case study in Changtu County, Northeast China. Agric. Ecosyst. Environ. 2020, 292, 106818. [Google Scholar] [CrossRef]
- Zheng, H.; Cao, S. Threats to China’s Biodiversity by Contradictions Policy. Ambio 2015, 44, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wu, Y.; Manevski, K.; Fu, M.; Yin, X.; Chen, F. A Framework for the Heterogeneity and Ecosystem Services of Farmland Landscapes: An Integrative Review. Sustainability 2021, 13, 12463. [Google Scholar] [CrossRef]
- Holland, J.M.; Bianchi, F.J.; Entling, M.H.; Moonen, A.; Smith, B.M.; Jeanneret, P. Structure, function and management of semi-natural habitats for conservation biological control: A review of European studies. Pest Manag. Sci. 2016, 72, 1638–1651. [Google Scholar] [CrossRef]
- Begg, G.S.; Cook, S.M.; Dye, R.; Ferrante, M.; Franck, P.; Lavigne, C.; Lövei, G.L.; Mansion-Vaquie, A.; Pell, J.K.; Petit, S.; et al. A functional overview of conservation biological control. Crop Prot. 2017, 97, 145–158. [Google Scholar] [CrossRef]
- Cunningham, M.A.; Johnson, D.H. Proximate and landscape factors influence grassland bird distributions. Ecol. Appl. 2006, 16, 1062–1075. [Google Scholar] [CrossRef]
- Montgomery, I.; Caruso, T.; Reid, N. Hedgerows as Ecosystems: Service Delivery, Management, and Restoration. Annu. Rev. Ecol. Evol. Syst. 2020, 51, 81–102. [Google Scholar] [CrossRef]
- Holden, J.; Grayson, R.P.; Berdeni, D.; Bird, S.; Chapman, P.J.; Edmondson, J.L.; Firbank, L.G.; Helgason, T.; Hodson, M.E.; Hunt, S.F.P.; et al. The role of hedgerows in soil functioning within agricultural landscapes. Agric. Ecosyst. Environ. 2019, 273, 1–12. [Google Scholar] [CrossRef]
- Søndergaard, M.; Jeppesen, E.; Jensen, J.P. Pond or lake: Does it make any difference? Arch. Hydrobiol. 2005, 162, 143–165. [Google Scholar] [CrossRef]
- Herzon, I.; Helenius, J. Agricultural drainage ditches, their biological importance and functioning. Biol. Conserv. 2008, 141, 1171–1183. [Google Scholar] [CrossRef]
- Ahiablame, L.; Chaubey, I.; Smith, D. Nutrient Content at the Sediment-Water Interface of Tile-Fed Agricultural Drainage Ditches. Water 2010, 2, 411–428. [Google Scholar] [CrossRef] [Green Version]
- Fahrig, L.; Baudry, J.; Brotons, L.; Burel, F.G.; Crist, T.O.; Fuller, R.J.; Sirami, C.; Siriwardena, G.M.; Martin, J. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Lett. 2011, 14, 101–112. [Google Scholar] [CrossRef]
- Jackson, L.; van Noordwijk, M.; Bengtsson, J.; Foster, W.; Lipper, L.; Pulleman, M.; Said, M.; Snaddon, J.; Vodouhe, R. Biodiversity and agricultural sustainagility: From assessment to adaptive management. Curr. Opin. Environ. Sustain. 2010, 2, 80–87. [Google Scholar] [CrossRef]
- Ansell, D.; Freudenberger, D.; Munro, N.; Gibbons, P. The cost-effectiveness of agri-environment schemes for biodiversity conservation: A quantitative review. Agric. Ecosyst. Environ. 2016, 225, 184–191. [Google Scholar] [CrossRef]
- Li, X.; Zhang, J.; Huang, J.; Lin, W.; Wu, S.; Ma, M. To Preserve Green Buffer under Polarization and Diffusion Effects of a Fast-Developing Megalopolis. Land 2022, 11, 724. [Google Scholar] [CrossRef]
- Rudnick, D.; Ryan, S.J.; Beier, P.; Cushman, S.A.; Dieffenbach, F.; Epps, C.; Gerber, L.R.; Hartter, J.N.; Jenness, J.S.; Kintsch, J.; et al. The role of landscape connectivity in planning and implementing conservation and restoration priorities. Issues Ecol. 2012, 16, 1–20. [Google Scholar]
- Brückmann, S.V.; Krauss, J.; Steffan-Dewenter, I. Butterfly and plant specialists suffer from reduced connectivity in fragmented landscapes. J. Appl. Ecol. 2010, 47, 799–809. [Google Scholar] [CrossRef]
- Perović, D.; Gámez-Virués, S.; Börschig, C.; Klein, A.; Krauss, J.; Steckel, J.; Rothenwöhrer, C.; Erasmi, S.; Tscharntke, T.; Westphal, C. Configurational landscape heterogeneity shapes functional community composition of grassland butterflies. J. Appl. Ecol. 2015, 52, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Fraterrigo, J.M.; Pearson, S.M.; Turner, M.G. Joint effects of habitat configuration and temporal stochasticity on population dynamics. Landsc. Ecol. 2009, 24, 863–877. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Van der Werf, W. The Effect of the Area and Configuration of Hibernation Sites on the Control of Aphids by Coccinella septempunctata (Coleoptera: Coccinellidae) in Agricultural Landscapes: A Simulation Study. Environ. Entomol. 2003, 32, 1290–1304. [Google Scholar] [CrossRef] [Green Version]
- Thies, C.; Tscharntke, T. Landscape structure and biological control in agroecosystems. Science 1999, 285, 893–895. [Google Scholar] [CrossRef] [PubMed]
- Steingröver, E.G.; Geertsema, W.; van Wingerden, W.K.R.E. Designing agricultural landscapes for natural pest control: A transdisciplinary approach in the Hoeksche Waard (The Netherlands). Landsc. Ecol. 2010, 25, 825–838. [Google Scholar] [CrossRef]
- Samways, M.J.; Barton, P.S.; Birkhofer, K.; Chichorro, F.; Deacon, C.; Fartmann, T.; Fukushima, C.S.; Gaigher, R.; Habel, J.C.; Hallmann, C.A.; et al. Solutions for humanity on how to conserve insects. Biol. Conserv. 2020, 242, 108427. [Google Scholar] [CrossRef]
- Aguilera, G.; Roslin, T.; Miller, K.; Tamburini, G.; Birkhofer, K.; Caballero Lopez, B.; Lindström, S.A.M.; Öckinger, E.; Rundlöf, M.; Rusch, A.; et al. Crop diversity benefits carabid and pollinator communities in landscapes with semi-natural habitats. J. Appl. Ecol. 2020, 57, 2170–2179. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Name | Formula | Range |
|---|---|---|---|
| AI | Aggregation index | 100 | 0 ≤ AI ≤ 100 |
| COHESION | Patch cohesion index | 100 | 0 < COHESION < 100 |
| DIVISION | Landscape division index | 0 < DIVISION < 1 | |
| ENN_MN | Mean Euclidean nearest neighbor index | ENN_MN > 0 | |
| LSI | Landscape shape index | LSI ≥ 1 | |
| SHAPE_MN | Mean shape index | SHAPE_MN ≥ 1 | |
| SPLIT | Splitting index | 1 ≤ SPLIT ≤ number of cells squared |
| Rank | Semi-Natural Habitat Types | R | p |
|---|---|---|---|
| 1 | Woodland, Waters, Ditch | 0.3478 | <0.01 |
| 2 | Woodland, Waters, Grassland, Ditch | 0.3368 | <0.01 |
| 3 | Woodland, Ditch | 0.3327 | <0.01 |
| 4 | Woodland, Ditch, Grassland | 0.3283 | <0.01 |
| 5 | Woodland, Waters, Ditch, Rural roads | 0.3103 | <0.01 |
| 6 | Woodland, Grassland, Waters, Rural roads, Ditch | 0.309 | <0.01 |
| 7 | Woodland, Waters | 0.2963 | <0.01 |
| 8 | Woodland, Grassland, Ditch, Rural roads | 0.2878 | <0.01 |
| 9 | Woodland, Waters, Grassland | 0.2872 | <0.01 |
| 10 | Woodland, Ditch, Rural roads | 0.2846 | <0.01 |
| Rank | Semi-Natural Habitat Configuration | R | p |
|---|---|---|---|
| 1 | AI + COHESION | 0.5205 | <0.01 |
| 2 | AI + COHESION + SPLIT | 0.5144 | <0.01 |
| 3 | AI | 0.5018 | <0.01 |
| 4 | AI + LSI + COHESION | 0.4905 | <0.01 |
| 5 | AI + SPLIT | 0.4889 | <0.01 |
| 6 | AI + LSI + COHESION + SPLIT | 0.4845 | <0.01 |
| 7 | AI + SHAPEMN + COHESION | 0.4694 | <0.01 |
| 8 | AI + LSI | 0.4671 | <0.01 |
| 9 | AI + SHAPEMN + COHESION + SPLIT | 0.4646 | <0.01 |
| 10 | AI + ENNMN + COHESION | 0.4627 | <0.01 |
| Name | F | p |
|---|---|---|
| Semi-natural habitat composition + semi-natural habitat configuration | 81.4 | 0.002 |
| Semi-natural habitat composition | 110 | 0.002 |
| Semi-natural habitat configuration | 95.5 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Guan, M.; Bian, Z.; Wang, Q. A Quantitative Survey of Effect of Semi-Natural Habitat Composition and Configuration on Landscape Heterogeneity in Arable Land System. Land 2022, 11, 1018. https://doi.org/10.3390/land11071018
Guo X, Guan M, Bian Z, Wang Q. A Quantitative Survey of Effect of Semi-Natural Habitat Composition and Configuration on Landscape Heterogeneity in Arable Land System. Land. 2022; 11(7):1018. https://doi.org/10.3390/land11071018
Chicago/Turabian StyleGuo, Xiaoyu, Minghao Guan, Zhenxing Bian, and Qiubing Wang. 2022. "A Quantitative Survey of Effect of Semi-Natural Habitat Composition and Configuration on Landscape Heterogeneity in Arable Land System" Land 11, no. 7: 1018. https://doi.org/10.3390/land11071018
APA StyleGuo, X., Guan, M., Bian, Z., & Wang, Q. (2022). A Quantitative Survey of Effect of Semi-Natural Habitat Composition and Configuration on Landscape Heterogeneity in Arable Land System. Land, 11(7), 1018. https://doi.org/10.3390/land11071018