
_Kazoglou.png)

Potential of Forage Grasses in Phytoremediation of Lead through Production of Phytoliths in Contaminated Soils

 , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Soil Characterization and Experimental Design
2.2. Measurements and Analytical Determinations
2.3. Statistics
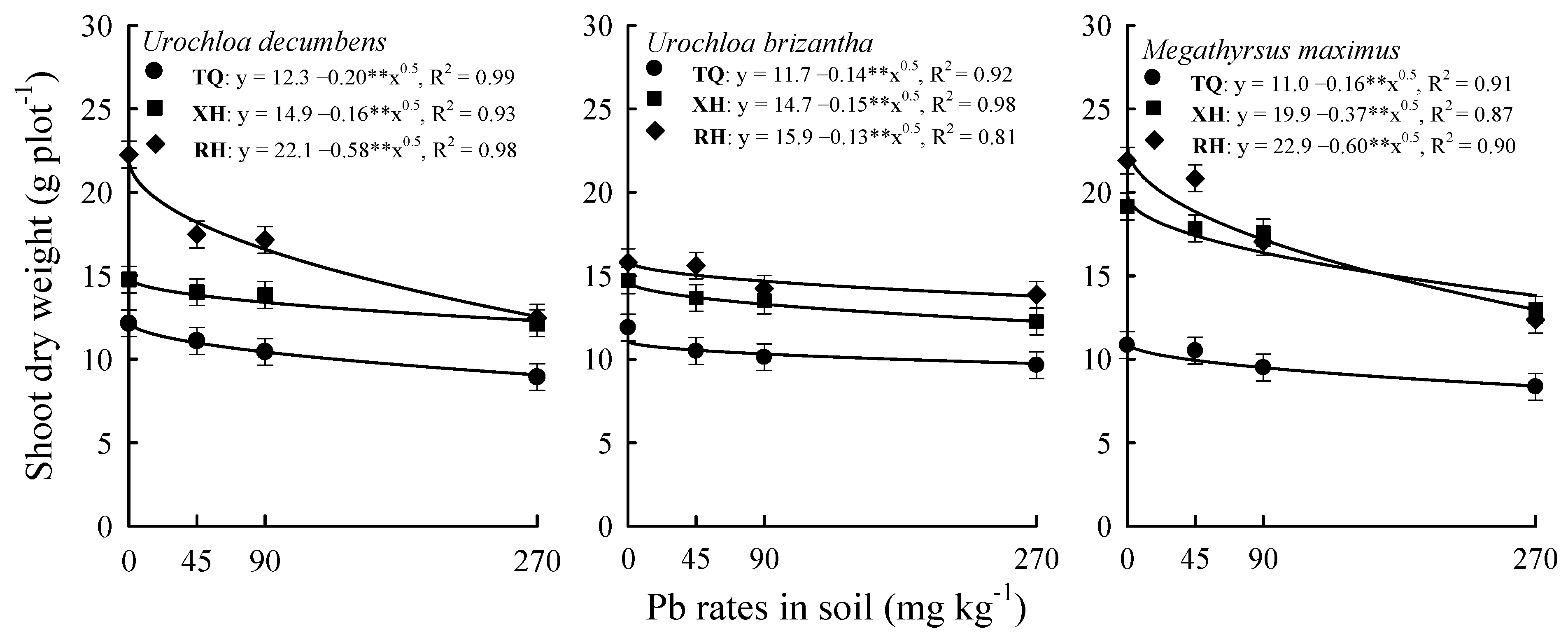
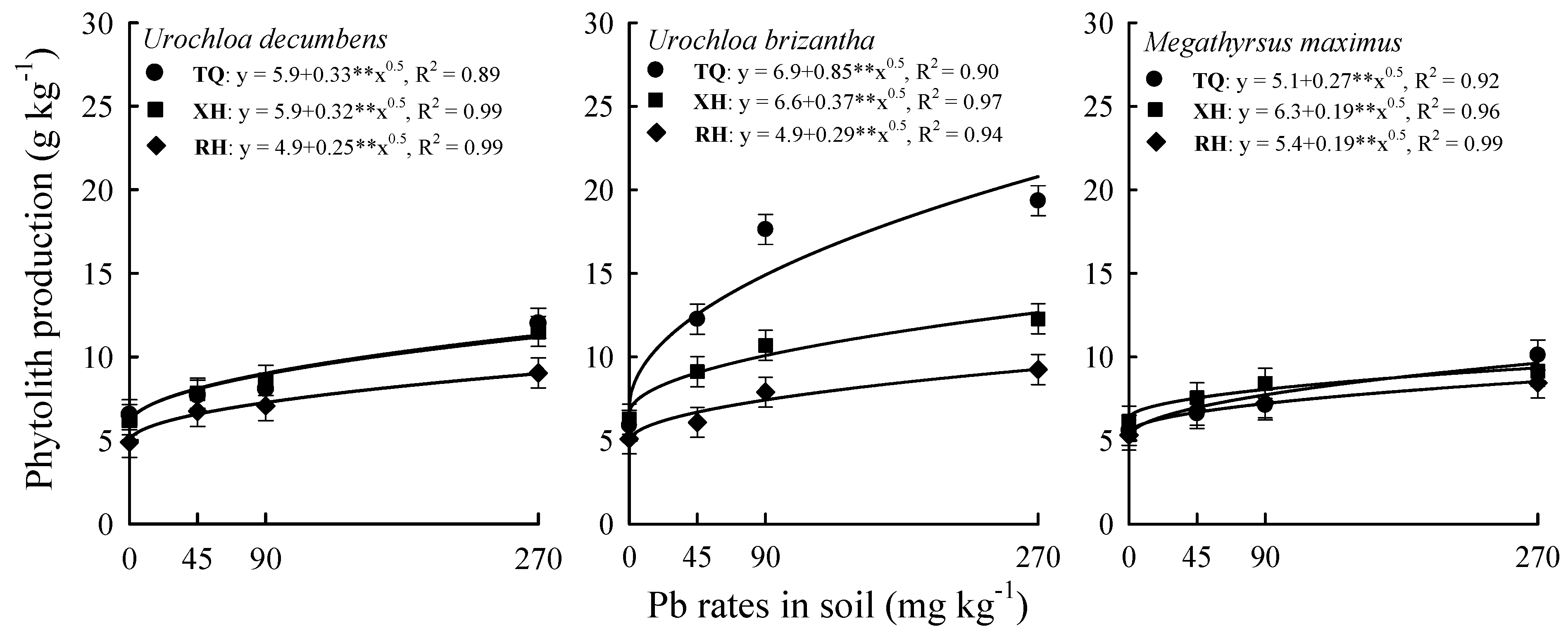
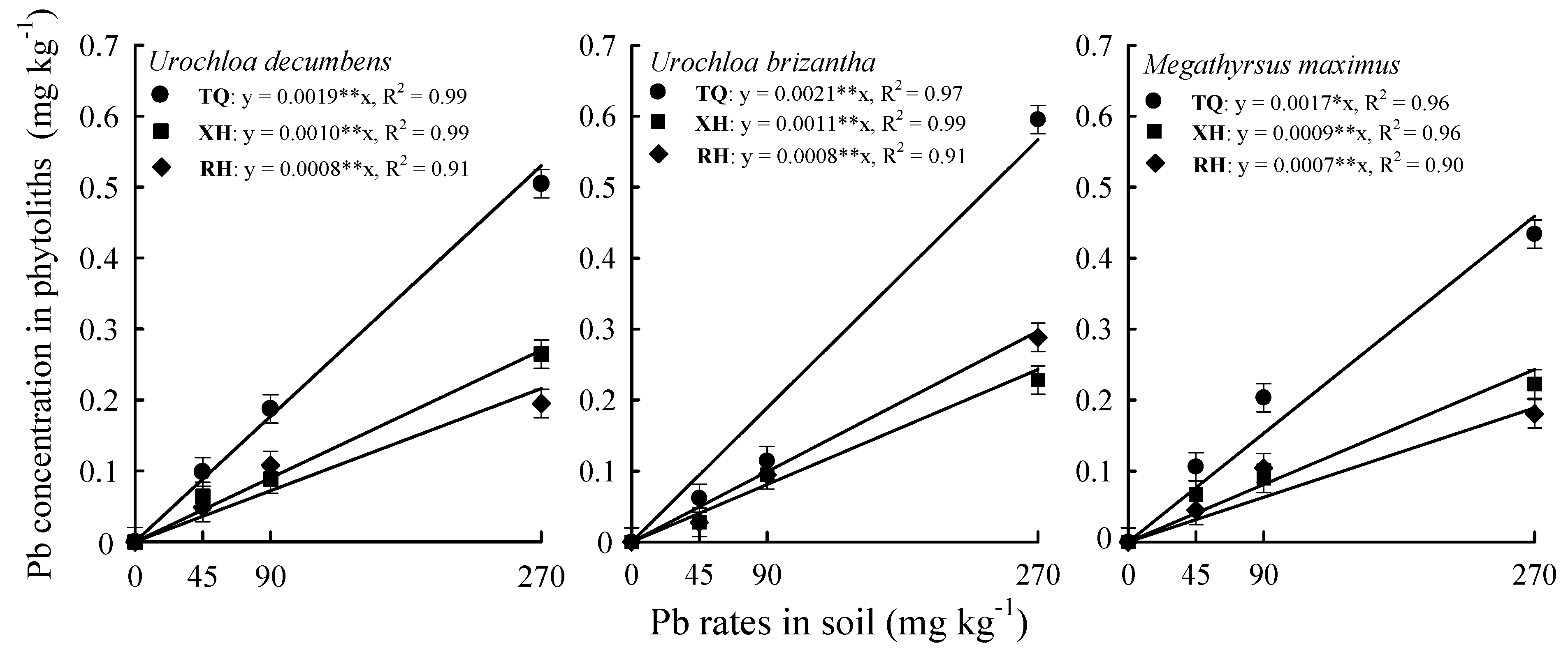
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sperdouli, I. Heavy metal toxicity effects on plants. Toxics 2022, 10, 715. [Google Scholar] [CrossRef] [PubMed]
- Van Der Merwe, M.J.; Osorio, S.; Moritz, T.; Nunes-Nesi, A.; Fernie, A.R. Decreased mitochondrial activities of malate dehydrogenase and fumarase in tomato lead to altered root growth and architecture via diverse mechanisms. Plant Physiol. 2009, 149, 653–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.A.; Piyatida, P.; Silva, J.A.T.; Fujita, M. Molecular mechanism of heavy metal toxicity and tolerance in plants: Central role of glutathione in detoxification of reactive oxygen species and methylglyoxal and in heavy metal chelation. J. Bot. 2012, 2012, 1–37. [Google Scholar] [CrossRef] [Green Version]
- Cannata, M.G.; Carvalho, R.; Bertoli, A.C.; Bastos, A.R.R.; Carvalho, J.G.; Freitas, M.P.; Augusto, A.S. Effects of lead on the content, accumulation, and translocation of nutrients in bean plant cultivated in nutritive solution. Commun. Soil Sci. Plant Anal. 2013, 44, 939–951. [Google Scholar] [CrossRef]
- Nascimento, S.S.; Silva, E.B.; Alleoni, L.R.F.; Grazziotti, P.H.; Fonseca, F.G.; Nardis, B.O. Availability and accumulation of lead for forage grasses in contaminated soil. J. Soil Sci. Plant Nutr. 2014, 14, 783–802. [Google Scholar] [CrossRef]
- Fontenele, N.M.B.; Otoch, M.D.L.O.; Gomes-Rochette, N.F.; Menezes Sobreira, A.C.; Barreto, A.A.G.C.; Oliveira, F.D.B.; Melo, D.F. Effect of lead on physiological and antioxidant responses in two Vigna unguiculata cultivars differing in Pb-ac.cumulation. Chemosphere 2017, 176, 397–404. [Google Scholar] [CrossRef]
- Charkiewic, A.E.; Backstrand, J.R. Lead Toxicity and Pollution in Poland. Int. J. Environ. Res. Public Health 2020, 17, 4385. [Google Scholar] [CrossRef]
- Miranda, L.S.; Anjos, J.A.S.A. Occupational impacts and adaptation to standards in accordance with Brazilian legislation: The case of Santo Amaro, Brazil. Saf. Sci. 2018, 104, 10–15. [Google Scholar] [CrossRef]
- Ng, C.C.; Nasrulhaq, A.; Rahman, M.M.; Abas, M.R.B. Tolerance Threshold and Phyto-assessment of Cadmium and Lead in Vetiver Grass, Vetiveria zizanioides (Linn.) Nash. Chiang Mai J. Sci. 2017, 44, 1367–1378. [Google Scholar]
- Yang, Y.; Jiang, M.; Liao, J.; Luo, Z.; Gao, Y.; Yu, W.; He, R.; Feng, S. Effects of simultaneous application of double chelating agents to pb-contaminated soil on the phytoremediation efficiency of Indocalamus decorus Q. H. Dai and the soil environment. Toxics 2022, 10, 713. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals-concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Kashiflrshad, M. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: A review. Ecotox. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Inam, F.; Deo, S.; Narkhede, N. Analysis of minerals and heavy metals in some spices collected from local market. J. Pharm. Biol. Sci. 2013, 8, 40–43. [Google Scholar] [CrossRef]
- Usman, K.; Abu-Dieyeh, M.H.; Zouari, N.; Al-Ghouti, M.A. Lead (Pb) bioaccumulation and antioxidative responses in Tetraena qataranse. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy metal tolerance in plants: Role of transcriptomics, proteomics, metabolomics, and ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezzopane, J.R.M.; Santos, P.M.; Mendonça, F.M.; Araujo, L.C.; Cruz, P.G. Dry matter production of Tanzania grass as a function of agrometeorological variables. Pesq. Agropec. Bras. 2012, 47, 471–477. [Google Scholar] [CrossRef] [Green Version]
- Pan, W.; Song, Z.; Liu, H.; Müeller, K.; Yang, X.; Zhang, X.; Li, Z.; Liu, X.; Qiu, S.; Hao, Q.; et al. Impact of grassland degradation on soil phytolith carbon sequestration in Inner Mongolian steppe of China. Geoderma 2017, 308, 86–92. [Google Scholar] [CrossRef]
- Ru, N.; Yang, X.; Song, Z.; Liu, H.; Hao, Q.; Liu, X.; Wu, X. Phytoliths and phytolith carbon occlusion in aboveground vegetation of sandy grasslands in eastern Inner Mongolia, China. Sci. Total Environ. 2018, 625, 1283–1289. [Google Scholar] [CrossRef]
- Yang, X.; Song, Z.; Liu, H.; Van Zwieten, L.; Song, A.; Li, Z.; Hao, Q.; Zhang, X.; Wang, H. Phytolith accumulation in broadleaf and conifer forests of northern China: Implications for phytolith carbon sequestration. Geoderma 2018, 312, 36–44. [Google Scholar] [CrossRef]
- Yan, A.; Wang, Y.; Tan, S.N.; Yusof, M.L.M.; Ghosh, S.; Chen, Z. Phytoremediation: A promising approach for revegetation of heavy metal-polluted land. Front. Plant Sci. 2020, 11, 359. [Google Scholar] [CrossRef]
- Gupta, D.K.; Huang, H.G.; Corpas, F.J. Lead tolerance in plants: Strategies for phytoremediation. Environ. Sci. Pollut. Res. Int. 2013, 20, 2150–2161. [Google Scholar] [CrossRef] [PubMed]
- Buján, E. Elemental composition of phytoliths in modern plants (Ericaceae). Quatern. Int. 2013, 287, 114–120. [Google Scholar] [CrossRef]
- Anala, R.; Nambisan, P. Study of morphology and chemical composition of phytoliths on the surface of paddy straw. Paddy Water Environ. 2015, 13, 521–527. [Google Scholar] [CrossRef]
- Olonova, M.V.; Gudkova, P.D.; Shiposha, V.D.; Kriuchkova, E.A.; Mezina, N.S.; Blinnikov, M. Phytoliths from some grasses (Poaceae) in arid lands of Xinjiang, China. Acta Biol. Sibirica. 2021, 7, 345–361. [Google Scholar] [CrossRef]
- Song, Z.; McGrouther, K.; Wang, H. Occurrence, turnover and carbon sequestration potential of phytoliths in terrestrial ecosystems. Earth-Sci. Rev. 2016, 158, 19–30. [Google Scholar] [CrossRef]
- Oliva, S.R.; Mingorance, M.D.; Leidi, E.O. Effects of silicon on copper toxicity in Erica andevalensis Cabezudo and Rivera: A potential species to remediate contaminated soils. J. Environ. Monit. 2011, 13, 591–596. [Google Scholar] [CrossRef]
- Fernandes-Horn, H.M.; Sampaio, R.A.; Horn, A.H.; Oliveira, E.S.A.; Lepsch, I.F.; Bilal, E. Use of Si-Phytoliths in depollution of mining areas in the Cerrado-Caatinga region, MG, Brazil. Int. J. Geomate 2016, 11, 2216–2221. [Google Scholar]
- Su, R.; Ou, Q.; Wang, H.; Luo, Y.; Dai, X.; Wang, Y.; Chen, Y.; Shi, L. Comparison of phytoremediation potential of Nerium indicum with inorganic modifier calcium carbonate and organic modifier mushroom residue to lead-zinc tailings. Int. J. Environ. Res. Public Health 2022, 19, 10353. [Google Scholar] [CrossRef]
- Bhat, S.A.; Bashir, O.; Ul Haq, S.A.; Amin, T.; Rafiq, A.; Ali, M.; Américo-Pinheiro, J.H.P.; Sher, F. Phytoremediation of heavy metals in soil and water: An eco-friendly, sustainable and multidisciplinary approach. Chemosphere 2022, 303, 134788. [Google Scholar] [CrossRef]
- Soil Survey Staff. Soil Taxonomy: Keys to Soil Taxonomy, 12th ed.; Department of Agriculture, Natural Resources and Conservation Service: Washington, DC, USA, 2014; p. 360.
- Teixeira, P.C.; Donagemma, G.K.; Fontana, A.; Teixeira, W.G. Manual for Methods of Soil Analysis, 3rd ed.; Embrapa: Brasília, Brazil, 2017; p. 573. [Google Scholar]
- USEPA. United States Environmental Protection Agency. Microwave Assisted Acid Digestion of Sediments, Sludges, Soils and Oils—Method 3052—SW—846; EPA: Washington, DC, USA, 1994. Available online: http:www.epa.gov/epaosver/hazwaste/test/3052.pdf (accessed on 8 October 2018).
- Santos, S.R.; Silva, E.B.; Alleoni, L.R.F.; Grazziotti, P.H. Citric acid influence on soil phosphorus availability. J. Plant Nut. 2017, 40, 2138–2145. [Google Scholar] [CrossRef]
- Conama; National Environmental Council of Brazil. Resolution no 420/2009. It Provides Criteria and Guiding Values of Soil Quality for the Presence of Chemical Substances and Establishes Guidelines for the Environmental Management of Areas Contaminated by These Substances as a Result of An-thropic Activities. Conama, Brasília, Brazil. 2009. Available online: http://www2.mma.gov.br/port/conama/legiabre.cfm?codlegi=620 (accessed on 8 February 2022).
- Parr, J.F.; Lentfer, C.J.; Boyd, W.E. A comparative analysis of wet and dry ashing techniques for the extraction of phytoliths from plant material. J. Archaeol. Sci. 2001, 28, 875–886. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attribute | Unit | Soil (6) | ||
|---|---|---|---|---|
| TQ | XH | RH | ||
| pH (1) water | - | 5.1 | 5.4 | 5.5 |
| P (2) | mg kg−1 | 0.2 | 0.1 | 0.2 |
| K (2) | mmolc kg−1 | 0.4 | 0.1 | 0.2 |
| Ca (3) | mmolc kg−1 | 6.7 | 4.50 | 8.1 |
| Mg (3) | mmolc kg−1 | 3.5 | 1.8 | 3.9 |
| Al (3) | mmolc kg−1 | 7.8 | 4.2 | 1.6 |
| Cation-exchange capacity | mmolc kg−1 | 40.6 | 71.4 | 49.2 |
| Organic carbon | g kg−1 | 3.5 | 5.8 | 5.2 |
| Pb (4) | mg kg−1 | 0.0 | 0.0 | 0.0 |
| Maximum P adsorption | mg kg−1 | 100 | 200 | 250 |
| Sand (5) | g kg−1 | 830.0 | 580.0 | 310.0 |
| Loam (5) | g kg−1 | 110.0 | 70.0 | 180.0 |
| Clay (5) | g kg−1 | 60.0 | 350.0 | 510.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farnezi, M.M.d.M.; Silva, E.d.B.; Santos, L.L.d.; Silva, A.C.; Grazziotti, P.H.; Alleoni, L.R.F.; Silva, W.C.; Santos, A.A.; Alves, F.A.F.; Bezerra, I.R.S.; et al. Potential of Forage Grasses in Phytoremediation of Lead through Production of Phytoliths in Contaminated Soils. Land 2023, 12, 62. https://doi.org/10.3390/land12010062
Farnezi MMdM, Silva EdB, Santos LLd, Silva AC, Grazziotti PH, Alleoni LRF, Silva WC, Santos AA, Alves FAF, Bezerra IRS, et al. Potential of Forage Grasses in Phytoremediation of Lead through Production of Phytoliths in Contaminated Soils. Land. 2023; 12(1):62. https://doi.org/10.3390/land12010062
Chicago/Turabian StyleFarnezi, Múcio Magno de Melo, Enilson de Barros Silva, Lauana Lopes dos Santos, Alexandre Christofaro Silva, Paulo Henrique Grazziotti, Luís Reynaldo Ferracciú Alleoni, Wesley Costa Silva, Angela Aparecida Santos, Flávio Antônio Fernandes Alves, Iracema Raquel Santos Bezerra, and et al. 2023. "Potential of Forage Grasses in Phytoremediation of Lead through Production of Phytoliths in Contaminated Soils" Land 12, no. 1: 62. https://doi.org/10.3390/land12010062
APA StyleFarnezi, M. M. d. M., Silva, E. d. B., Santos, L. L. d., Silva, A. C., Grazziotti, P. H., Alleoni, L. R. F., Silva, W. C., Santos, A. A., Alves, F. A. F., Bezerra, I. R. S., & Miranda, L. C. (2023). Potential of Forage Grasses in Phytoremediation of Lead through Production of Phytoliths in Contaminated Soils. Land, 12(1), 62. https://doi.org/10.3390/land12010062